Introduction
The semaphorin family is a large super-protein
family containing secreted, transmembrane and
glycosylphosphatidylinositol-linked proteins, and it has been
divided into eight hypotypes based on their structures and amino
acid sequence similarity: Invertebrate semaphorins consist of
classes 1 and 2, while classes 3–7 are vertebrate semaphorins (with
the exception of class 5C semaphorins, which are encoded by viral
genomes) (1). The semaphorin family
was initially identified to be involved in mediating axonal
guidance in the developing nervous system (2). Certain members of the semaphorin family
have been also shown to exert diverse and important functions in
other physiological processes, including heart morphogenesis
(3,4),
vascular growth (5), immune cell
regulation (6) and tumor progression
(7,8).
Important regulatory functions of certain members of the semaphorin
family within tumor angiogenesis have been reported. For example,
semaphorin 3A (Sema3A) inhibits angiogenesis through competition
for vascular endothelial growth factor (VEGF) (9), and Sema3B has also been indicated as a
putative tumor suppressor that inhibits tumor growth and
angiogenesis (10). By contrast,
Sema3C presumably may promote angiogenesis by stimulating integrin
phosphorylation and VEGF120 secretion (11), and Sema4D has been demonstrated to
serve a role in tumor-induced angiogenesis (12).
Sema6D, mapped on chromosome 2, is the best
characterized factor of the class 6 semaphorins, which are
single-pass membrane-bound semaphorins (13). It was reported that Sema6D regulates
the late-phase activity of T cells during the primary immune
response (14), and it functions as a
promoter of tissue remodeling (15)
and tumorigenesis (16).
Additionally, Sema6D also functions as a ligand for plexin-A1
during T cell-dendritic cell interactions (6). Plexin-A1 serves as a main receptor for
Sema6D and contributes to cardiac morphogenesis (17). Notably, plexin-A1 forms complexes with
VEGF receptor-2 (VEGFR-2), which undergoes phosphorylation upon
stimulation by Sema6D (15), and
VEGFR is responsible for the transduction of pro-angiogenic signals
(18). The association between
plexin-A1 expression and gastric carcinoma angiogenesis has been
previously explored by the present authors (19).
In the current study, Sema6D was observed to be
highly expressed in tumor tissue compared with normal gastric
mucosa and was determined to be responsible for tumor promotion.
Its putative receptors were detected, and it was speculated that
plexin-A1 may be the main receptor for Sema6D. Next, the levels of
Sema6D and plexin-A1 were detected, and it was noticed that they
were highly expressed in tumor vascular endothelial cells.
Furthermore, both were positively correlated with VEGFR2. These
observations indicate that they may function as a modifier in the
formation of tumor angiogenesis. This is consistent with a previous
report showing that Sema6D activates VEGFR2 through
plexin-A1-mediated signal transduction and controls cardiac
morphogenesis (20).
Materials and methods
Human tissue specimens
Gastric tissues were obtained from The 266th General
Hospital of People's Liberation Amy (Chengde, China) with the
institutional approval and informed consent of the patients. The
procedures to obtain human gastric tissues were in accordance with
the Ethical Principles for Medical Research Involving Human
Subjects, as formulated in the World Medical Association
Declaration of Helsinki (revised in 2008). During surgical
resection, gastric tumor tissues and normal gastric tissues
(located ~5 cm away from the macroscopic margin of the resected
tumors) were obtained from 10 patients who were diagnosed as
gastric carcinomas by pathologists. The patient ages ranged between
37 and 82 years, with a median age of 58 years. There were six male
and four female patients. None of the patients had received any
chemotherapy or radiotherapy prior to biopsy or surgery.
Reagents
RPMI-1640 medium, fetal bovine serum (FBS), 0.25%
trypsin and 0.02% ethylenediaminetetraacetic acid (EDTA) were
purchased from Gibco (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). SuperScript III First-Strand Synthesis System kit and
GoTaq® qPCR Master Mix were purchased from Promega
Corporation (Madison, WI, USA). Anti-VEGFR2 mouse monoclonal
antibody (ab9530), anti-plexin-A1 rabbit monoclonal antibody
(ab32960) and anti-β-actin mouse monoclonal antibody (ab8226) were
purchased from Abcam (Cambridge, MA, USA), while anti-Sema6D goat
monoclonal antibody (sc-67965) was purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). Peroxidase-conjugated
AffiniPure goat anti-mouse immunoglobulin G (IgG),
peroxidase-conjugated AffiniPure goat anti-rabbit IgG,
peroxidase-conjugated AffiniPure donkey anti-goat IgG, Alexa
Fluor® 488-conjugated AffiniPure goat anti-mouse IgG,
Alexa Fluor® 594-conjugated AffiniPure donkey anti-goat
IgG and Alexa Fluor® 594-conjugated AffiniPure goat
anti-rabbit IgG were purchased from Santa Cruz Biotechnology,
Inc.
Cell culture
Human gastric cancer cell lines (MGC803, HGC27 and
MNK45) and human normal gastric mucosa cell line (GES-1) were
provided by the Academy of Military Medical Sciences (Beijing,
China). The cell lines were cultured in an incubator with an
atmosphere of 5% CO2 at 37°C in RPMI-1640 medium
supplemented with 10% FBS. The cells were then subcultured with
0.25% trypsin and 0.02% EDTA when the cell growth reached 80–90%.
The experiments were carried out when the cells reached logarithmic
growth phase.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was isolated using the
RNAgents® Total RNA Isolation System (Promega
Corporation) with DNase I (Invitrogen; Thermo Fisher Scientific,
Inc.) treatment. RNA (2 µg), oligo(dT)20 primers and the
SuperScript III First-Strand Synthesis System kit were used to
synthesize complementary DNA. qPCR was performed using the
SYBR® Green I dye provided in the GoTaq® qPCR
Master Mix according to the manufacturer's protocol. PCR was
performed under the following conditions: Denaturation at 95°C for
30 sec, followed by 40 cycles at 95°C for 3 sec, 60°C for 30 sec
and 72°C for 45 sec. The results were analyzed using the
comparative quantitative cycle (Cq) method (21), with glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) as an internal control. The results were
normalized to the GAPDH levels using the formula ΔCq = Cq of the
target gene - Cq of GAPDH. The messenger RNA (mRNA) level of the
control group was used as the baseline, and ΔΔCq was calculated
using the formula ΔΔCq = ΔCq of the target gene - ΔCq of the
baseline. The fold-change in mRNA level was calculated as
2−ΔΔCq.
For RT-PCR, the PCR products were resolved in 2%
agarose gels and visualized by staining with ethidium bromide. To
semiquantify the PCR products, the bands representing the amplified
products were analyzed by Quantity One® 1-D analysis
software (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
relative level of the target mRNA expression was defined as the
ratio of the absorbance of the target band to that of the β-actin
band. The primers used in the present study were synthesized by
Shenzhen Huada Gene Technology Co., Ltd. (Shenzhen, China), and
their sequences are presented in Table
I.
 | Table I.Primers used in the present study. |
Table I.
Primers used in the present study.
| Transcript | Primer sequence
(5′-3′) |
|---|
| Sema6D |
TGAGGAGGAAGGTAGCTCAGTG (Sense) |
|
|
CCATCAGCAGCAGTATGTAGGC (Antisense) |
| Plexin-A1 |
TGGACGACCTGTTTGAGACCA (Sense) |
|
|
TGATCACGTTCACCCAGAAGC (Antisense) |
| Plexin-A2 |
CATCYCGTACTGGACCCCAC (Sense) |
|
|
TTTACAACGGCTACAGCGTG (Antisense) |
| Plexin-A4 | TCTCAGTACAACGTGCTG
(Sense) |
|
| TAGCACTGGATCTGATTGC
(Antisense) |
| VEGFR2 |
CTACCAGTACGGCACCACTCAA (Sense) |
|
|
TCTTCCTCCAACTGCCAATACC (Antisense) |
| β-actin |
TGACGTGGACATCCGCAAAG (Sense) |
|
|
CTGGAAGGTGGACAGCGAGG (Antisense) |
| GAPDH |
TGAAGGTCGGAGTCAACGGAT (Sense) |
|
|
CTGGAAGATGGTGATGGGATT (Antisense) |
Western blot analysis
Total proteins were extracted from each group with
radioimmunoprecipitation assay buffer (Thermo Fisher Scientific,
Inc.) and quantified using the Pierce BCA Protein Assay kit (Thermo
Fisher Scientific, Inc.). The proteins were separated by 10% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis and transferred
to polyvinylidene difluoride membranes. Upon blocking with 5%
bovine serum albumin (Beijing Solarbio Science & Technology
Co., Ltd., Beijing, China), the blots were probed with the
appropriate primary antibodies overnight at 4°C. The antibodies
used were anti-Sema6D antibody (1:500 dilution), anti-plexin-A1
antibody (1:1,000 dilution), anti-VEGFR2 antibody (1:500 dilution)
and anti-β-actin antibody (1:1,000 dilution). The membranes were
washed three times for 10 min in Tris-buffered saline containing
Tween 20, and incubated with horseradish peroxidase-conjugated
secondary antibodies [goat anti-mouse (sc-2039; Santa Cruz
Biotechnology, Inc.), goat anti-rabbit (sc-2040; Santa Cruz
Biotechnology, Inc.) or donkey anti-goat (sc-2024; Santa Cruz
Biotechnology, Inc.), correspondingly] at 1:1,000 dilution for 2 h
at 37°C. Immunoreactive bands were detected using Pierce ECL
Western Blotting Substrate (Thermo Fisher Scientific, Inc.) and
imaged using the ImageQuant LAS 4000 system (GE Healthcare Life
Sciences, Chalfont, UK).
Immunohistochemistry and fluorescence
microscopy
Immunohistochemical analysis was performed using
standard techniques. Briefly, paraffin-embedded tissues were cut
into 4-µm-thick sections, deparaffinized and antigen-recovered in
citrate buffer. The sections were blocked for endogenous avidin,
peroxidase and biotin, and then incubated with anti-plexin-A1
antibody (ab32960; Abcam) or anti-Sema6D antibody (sc-67965; Santa
Cruz Biotechnology, Inc.) overnight at 4°C. Upon washing three
times with phosphate-buffered saline (PBS), the staining was
developed using the Universal LSAB™ kit/HRP, Rabbit/Mouse/Goat
(Dako, Glostrup, Denmark) according to the manufacturer's
protocol.
For fluorescence immunohistochemical staining and
microscopy, the sections were fixed in 4% paraformaldehyde for 30
min and then permeabilized in 0.2% Triton X-100 in PBS for 10 min.
The primary antibodies [mouse anti-VEGFR2 antibody (ab9530; Abcam;
1:100 dilution), rabbit anti-plexin-A1 antibody (ab32960; Abcam;
1:200 dilution) and goat anti-Sema6D antibody (sc-67965; Santa Cruz
Biotechnology, Inc.; 1:100 dilution)] were incubated overnight at
4°C. Then, the appropriate Alexa-Fluor®-conjugated
secondary antibodies [Alexa Fluor® 488-conjugated
AffiniPure goat anti-mouse IgG (sc-395764; Santa Cruz
Biotechnology, Inc.), Alexa Fluor® 594-conjugated
AffiniPure donkey anti-goat IgG (sc-362275; Santa Cruz
Biotechnology, Inc.) and Alexa Fluor® 594-conjugated
AffiniPure goat anti-rabbit IgG (sc-362282; Santa Cruz
Biotechnology, Inc.)] were used at 37°C for 2 h at 1:200 dilution.
The nuclei were stained using 4′,6-diamidino-2-phenylindole
(H-1200; Vector Laboratories, Inc., Burlingame, CA, USA).
Fluorescence images were collected under a laser scanning confocal
microscope (Leica Microsystems GmbH, Wetzlar, Germany).
Short hairpin (sh) RNA
transfection
shRNA plasmid vectors were purchased from Shanghai
GenePharma Co., Ltd. (Shanghai, China), including
pGPU6/GFP/Neo-shplexin-A1 (targeting plexin-A1) and
pGPU6/GFP/Neo-shNC (not targeting any gene, which served as
control). The constructs were transfected into MGC803 cells with
jetPRIME® (Polyplus-transfection® SA,
Illkirch, France). The procedure of transfection was performed
according to the manufacturer's protocol. Next, stable cell clones
were selected by treatment with 400–1,000 µM G418 (Beijing Solarbio
Science & Technology Co., Ltd.) for 1 month.
Antibiotic-resistant cell clones were verified by the expression of
green fluorescent protein.
Statistical analysis
All experiments were repeated ≥3 times, unless
otherwise indicated. Data are presented as the mean ± standard
deviation. Statistical analysis involved the use of one-way
analysis of variance and Student's t-test. Statistical analysis was
performed using SPSS version 19.0 (IBM SPSS, Armonk, NY, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Sema6D and its receptor plexin-A1 are
highly expressed in gastric tumor tissues compared with normal
gastric mucosa
In the present study, RT-qPCR was performed to
analyze four cases of gastric carcinoma and the corresponding
cancer-adjacent lesions in gastric cancer. The results demonstrated
that gastric tumor tissues expressed high levels of Sema6D compared
with normal gastric mucosa (Fig. 1A).
The protein expression levels of Sema6D were evaluated by western
blotting (Fig. 1B). Based on the
results of our current and previous studies (13), we speculated that Sema6D may serve a
role in the formation and development of gastric cancer. According
to a previous study, plexin-A1, plexin-A2 and plexin-A4 are all
downstream receptors for Sema6D (22). Plexin-As have been shown to form a
functional complex with neuropilin-1 (NP-1) and/or NP-2 (23) for certain semaphorins. For example,
plexin-As are co-expressed with both Sema3A and NPs in yolk sac
endothelial cells during vasculogenesis (24). However, NPs neither bound to Sema6D
nor influenced the binding of Sema6D to plexin-A1, and another
study also indicated that NPs may function independently (25). Based on the fact that Sema6D functions
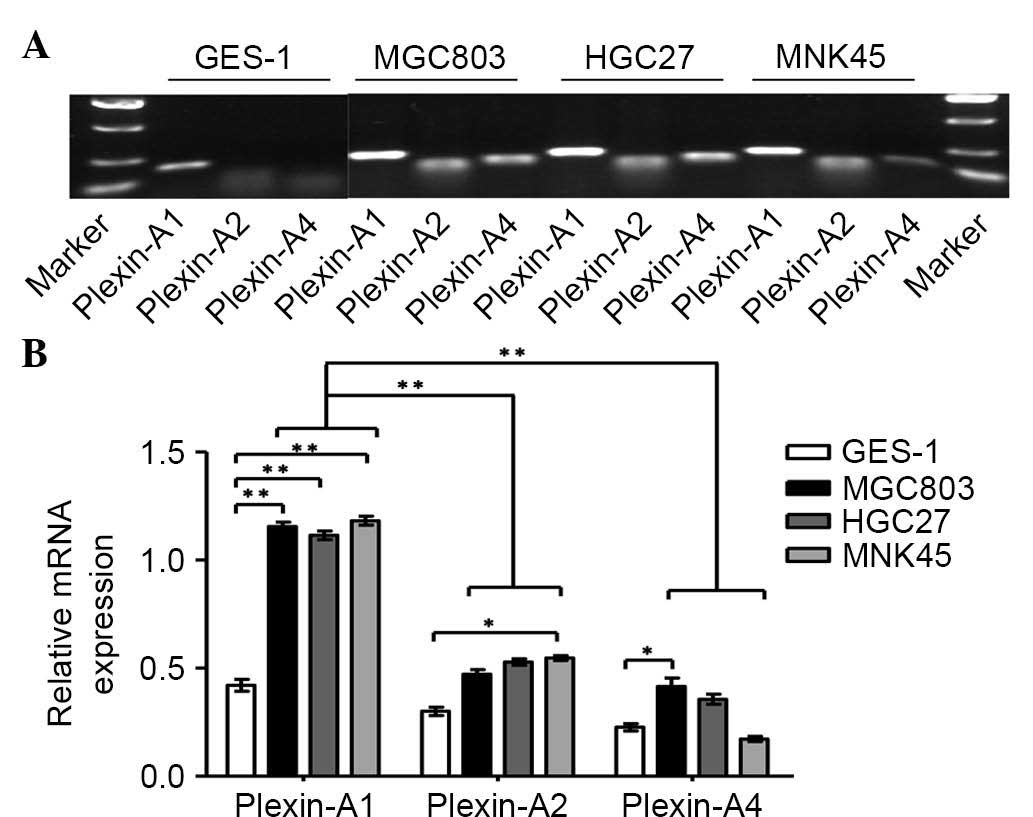
through plexin-As but does not depend on NPs, the present study
next detected the levels of plexin-As in GES-1 cells and in a panel
of gastric cancer cell lines by RT-PCR (Fig. 2A). The results indicated that
plexin-A1 is highly expressed in gastric cancer cells lines
compared with GES-1 cells. However, no significant difference in
the mRNA expression of plexin-A2 or plexin-A4 was identified
between gastric cancer cells and normal gastric mucosa. It was also
observed that plexin-A1 expression was remarkably higher than that
of plexin-A2 and plexin-A4 in the gastric cancer cell lines
(Fig. 2B). Based on previous studies
(13,19) and the current results, we speculate
that plexin-A1 may be the putative receptor for Sema6D. Given the
active roles of Sema6D in regulating tumor development observed in
the present study, the mRNA and protein levels of plexin-A1 were
next detected by qPCR and western blotting, respectively, using
four cases of gastric cancer tissues and corresponding
cancer-adjacent tissues. The results revealed that plexin-A1 was
highly expressed in gastric tumor tissues at the mRNA level
(Fig. 3A). Similar results were
obtained for the plexin-A1 protein expression by western blotting
(Fig. 3B). Therefore, we hypothesized
that Sema6D functions in tumorigenesis using plexin-A1 as a
receptor.
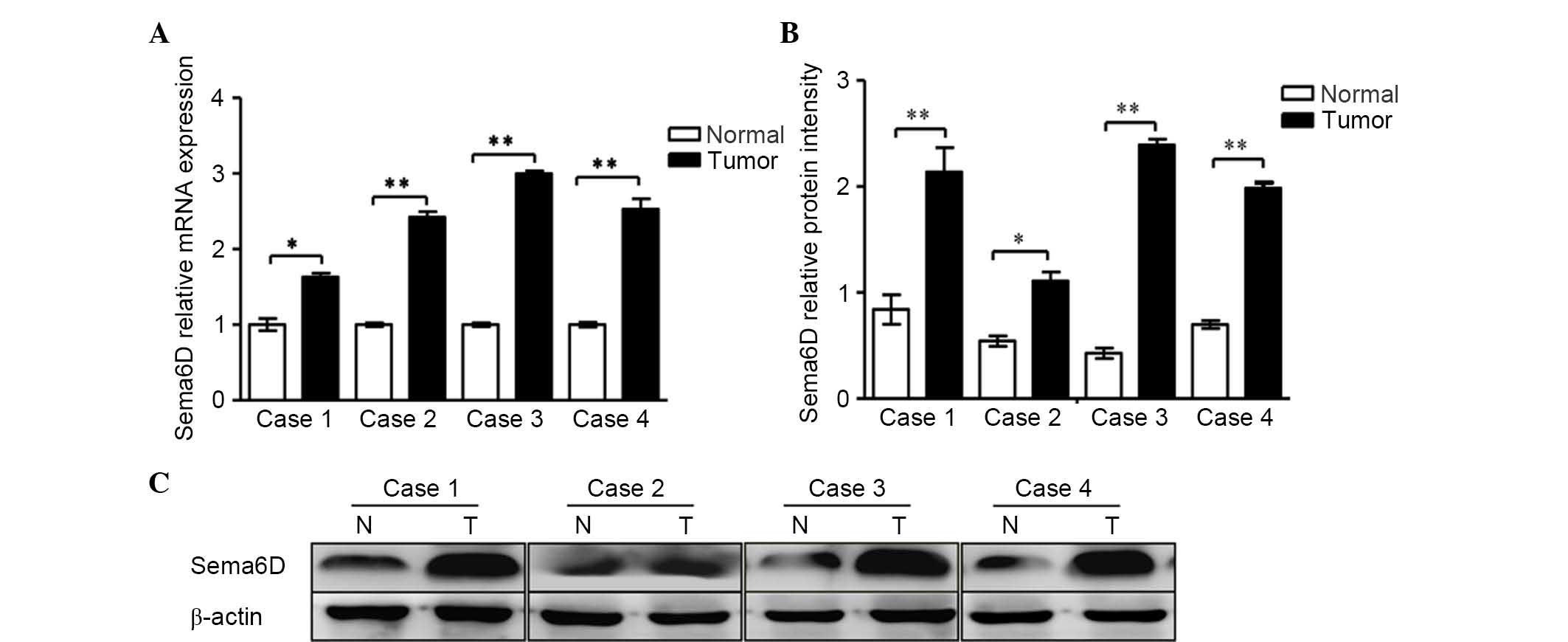 | Figure 1.Sema6D was highly expressed in four
cases of gastric cancer tissue. (A) Reverse
transcription-quantitative polymerase chain reaction analyses of
the mRNA expression of Sema6D in four cases of gastric cancer.
Sema6D mRNA levels increased remarkably by 1.6, 2.4, 2.9 and
2.6-fold in tumor cases 1, 2, 3 and 4, respectively, when compared
with normal gastric mucosa. (B) The relative protein intensities of
Sema6D with respect to the loading control, β-actin, are shown. (C)
The protein expression levels of Sema6D in the four specimens were
enhanced. Data are represented as the mean ± standard deviation
(n=3 for each group, *P<0.05, **P<0.01). Sema, semaphorin;
mRNA, messenger RNA; N, normal; T, tumor. |
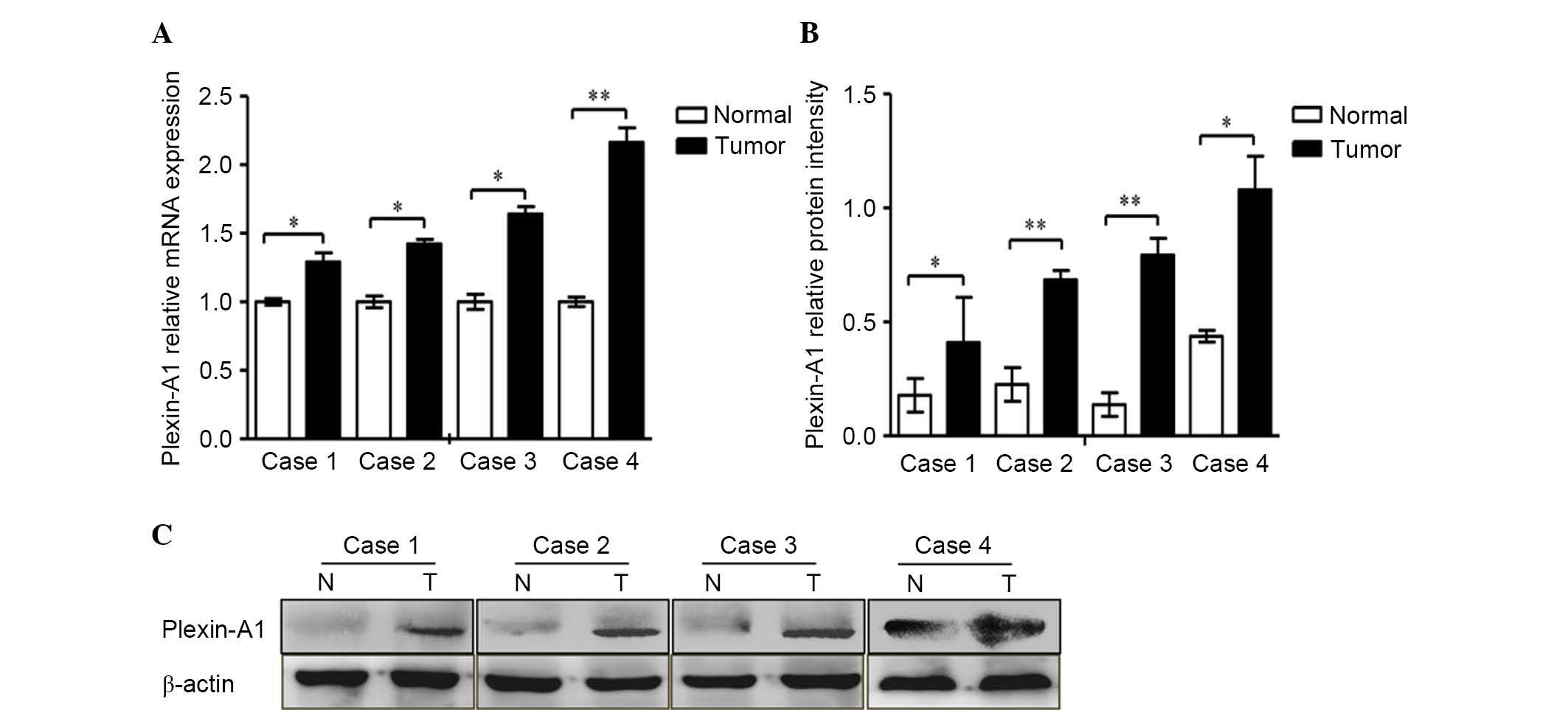 | Figure 3.Plexin-A1 was highly expressed in four
cases of gastric cancer tissue. (A) The mRNA expression of
plexin-A1 was obviously increased in cancer tissue compared with
normal gastric mucosa. Plexin-A1 mRNA levels increased remarkably
by 1.3, 1.4, 1.6 and 2.2-fold in tumors 1, 2, 3 and 4,
respectively, compared with normal gastric tissue. (B) The
histogram indicates the protein levels, which were normalized to
the levels of β-actin protein. (C) The protein expression levels of
plexin-A1 in all four cancer specimens were remarkably enhanced.
Data are represented as the mean ± standard deviation (n=3 for each
group, *P<0.05, **P<0.01). mRNA, messenger RNA; N, normal; T,
tumor. |
Sema6D and its receptor plexin-A1 are
highly expressed in vascular epithelial cells within gastric
cancer
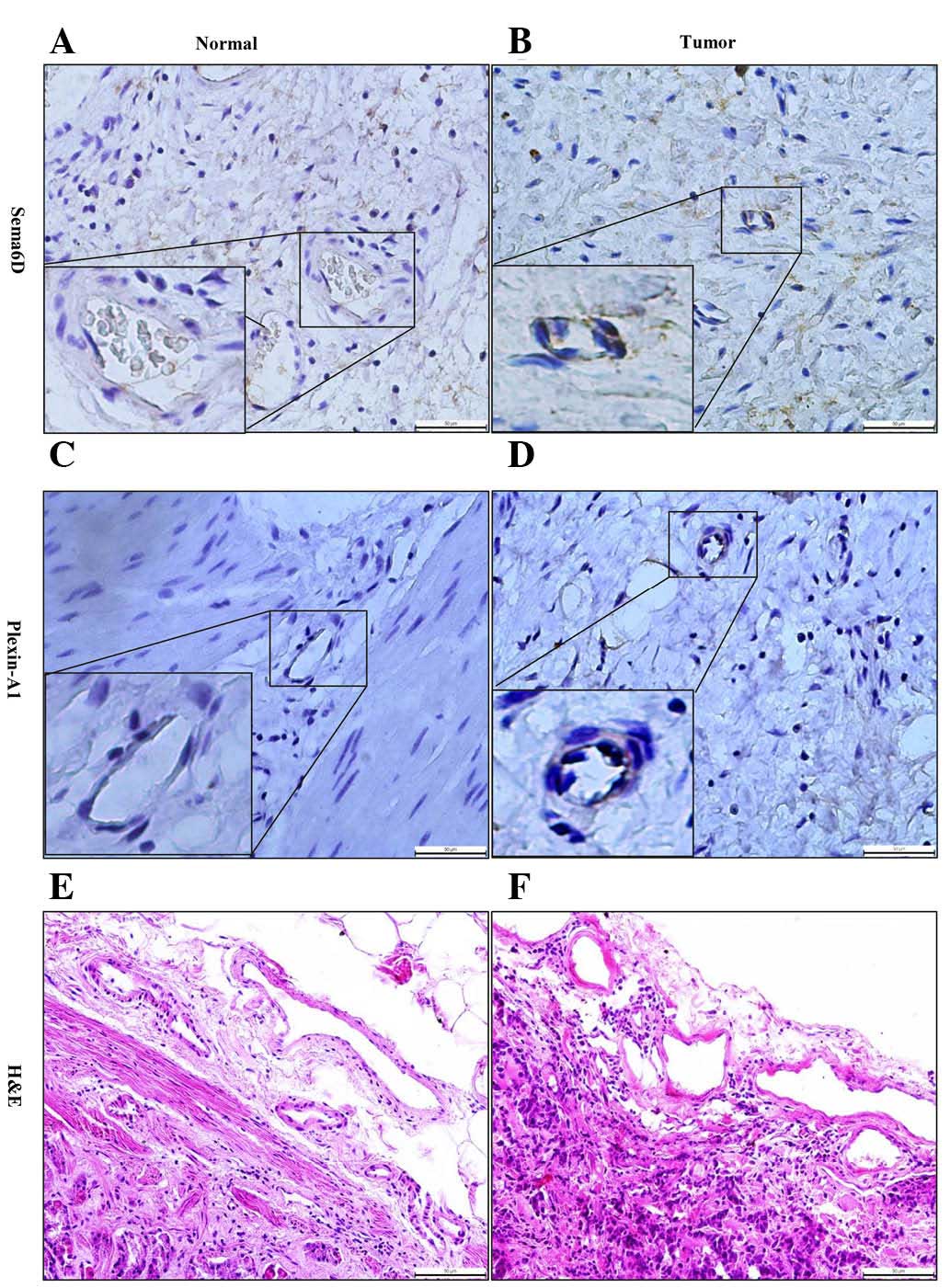
To further address the role of Sema6D and its
receptor plexin-A1 in tumorigenesis, immunohistochemical methods
were used to detect their expression and localization. Notably,
Sema6D and plexin-A1 were mainly located at the membrane and
cytoplasm of gastric carcinoma vascular endothelial cells, which
appeared as brown particles (Fig. 4).
Therefore, we speculated that they may serve a critical role in
tumor angiogenesis. Angiogenesis involves the formation of new
blood vessels from primitive vasculature, which is critical to
numerous physiological processes, and vascular malformation may
lead to several major diseases, particularly tumor progression
(26). Thus, the present study next
attempted to determine the mechanism of angiogenesis promotion
mediated by Sema6D and plexin-A1 in vessel endothelial cells.
Sema6D and plexin-A1 promote tumor
angiogenesis via VEGFR2 signaling
Various GFRs are implicated in angiogenesis,
particularly the VEGFR family of receptor tyrosine kinases
(27). VEGFR1 and VEGFR2 are closely
related receptor tyrosine kinases, and VEGFR2 serves a broader role
than VEGFR1 in angiogenesis (28).
Sema6D was reported to participate in cardiac morphogenesis by
exerting distinct biological activities through its receptor,
plexin-A1, which formed receptor complexes with VEGFR2 in adjacent
regions to the cardiac tube (15).
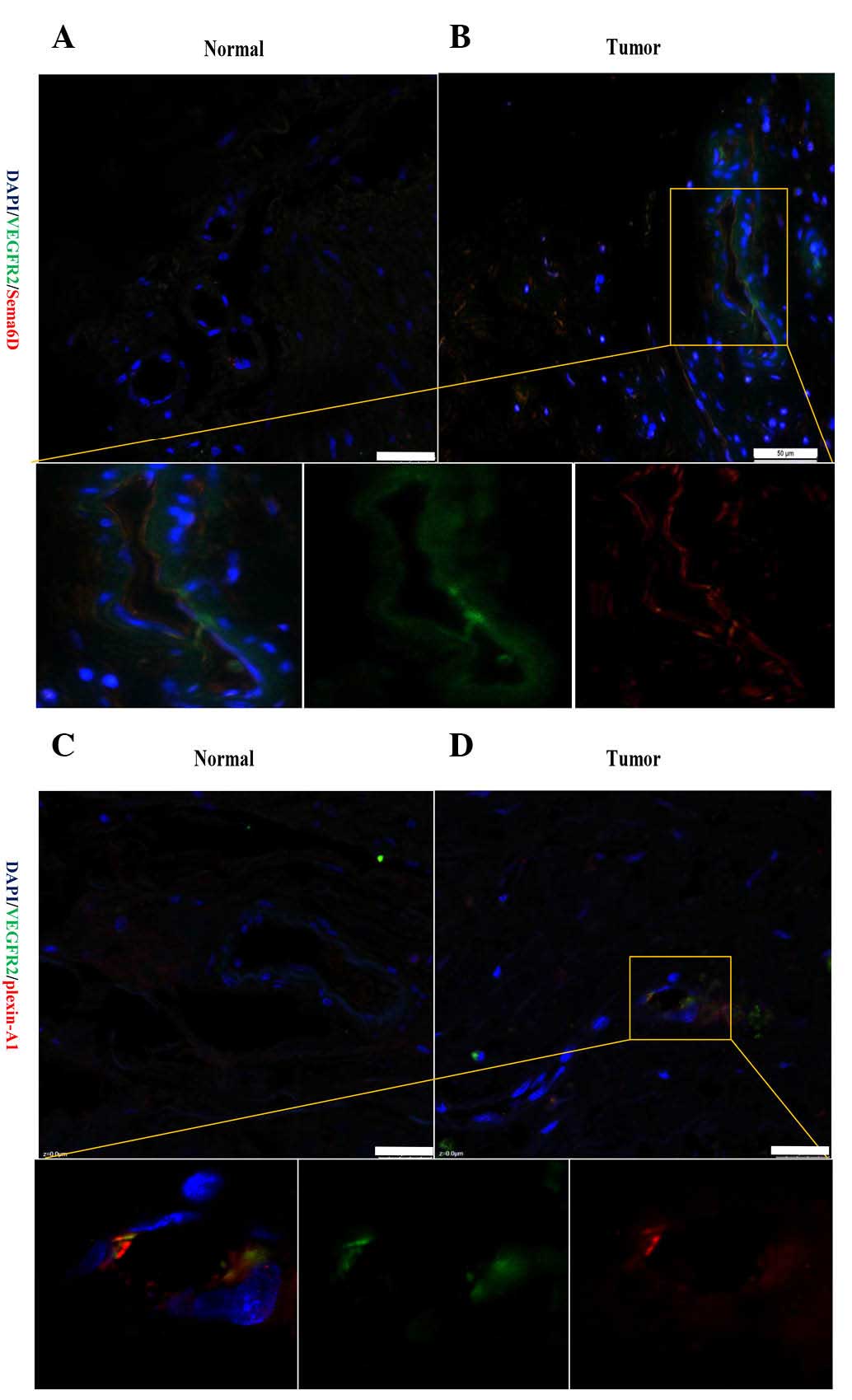
Thus, the current study next detected the association of Sema6D and
plexin-A1 with VEGFR2. An immunofluorescence method was used to
detect the location of Sema6D and plexin-A1 in regards to that of
VEGFR2. Notably, Sema6D and VEGFR2 were observed to be highly
expressed in the membrane and cytoplasm of the same vascular
endothelial cells in cancerous tissue. Representative fluorescence
images revealed that Sema6D was expressed at a high level and was
strongly correlated with the distribution patterns of VEGFR2 in
tumor tissue (Fig. 5A and B).
Similarly, double-labelling of cells for plexin-A1 and VEGFR2 could
also be detected in the section of cancer tissues (Fig. 5C and D). This confirms that
Sema6D/plexin-A1 may be closely associated with tumor angiogenesis
via VEGFR2.
Sema6D, plexin-As and VEGFR2 were further
systematically detected in three gastric cancer cell lines by
RT-PCR (Fig. 6A), and significant
differences in the mRNA expression of Sema6D, plexin-A1 and VEGFR2
were identified in gastric cancer cells compared with normal
gastric mucosa (Fig. 6B). The protein
expression levels of VEGFR2 in MGC803 and GES-1 cells were detected
by western blotting, which revealed that VEGFR2 exhibited a high
expression in MGC803 cells (Fig. 6C).
To further address the interaction between plexin-A1 and VEGFR2,
plexin-A1 was knocked down using shplexin-A1 in MGC803 cells. The
mRNA (Fig. 6D) and protein (Fig. 6E and F) expression levels of VEGFR2
were downregulated with the targeted disruption of plexin-A1. This
finding verified our speculation that plexin-A1 serves its role via
VEGFR2 in tumor angiogenesis.
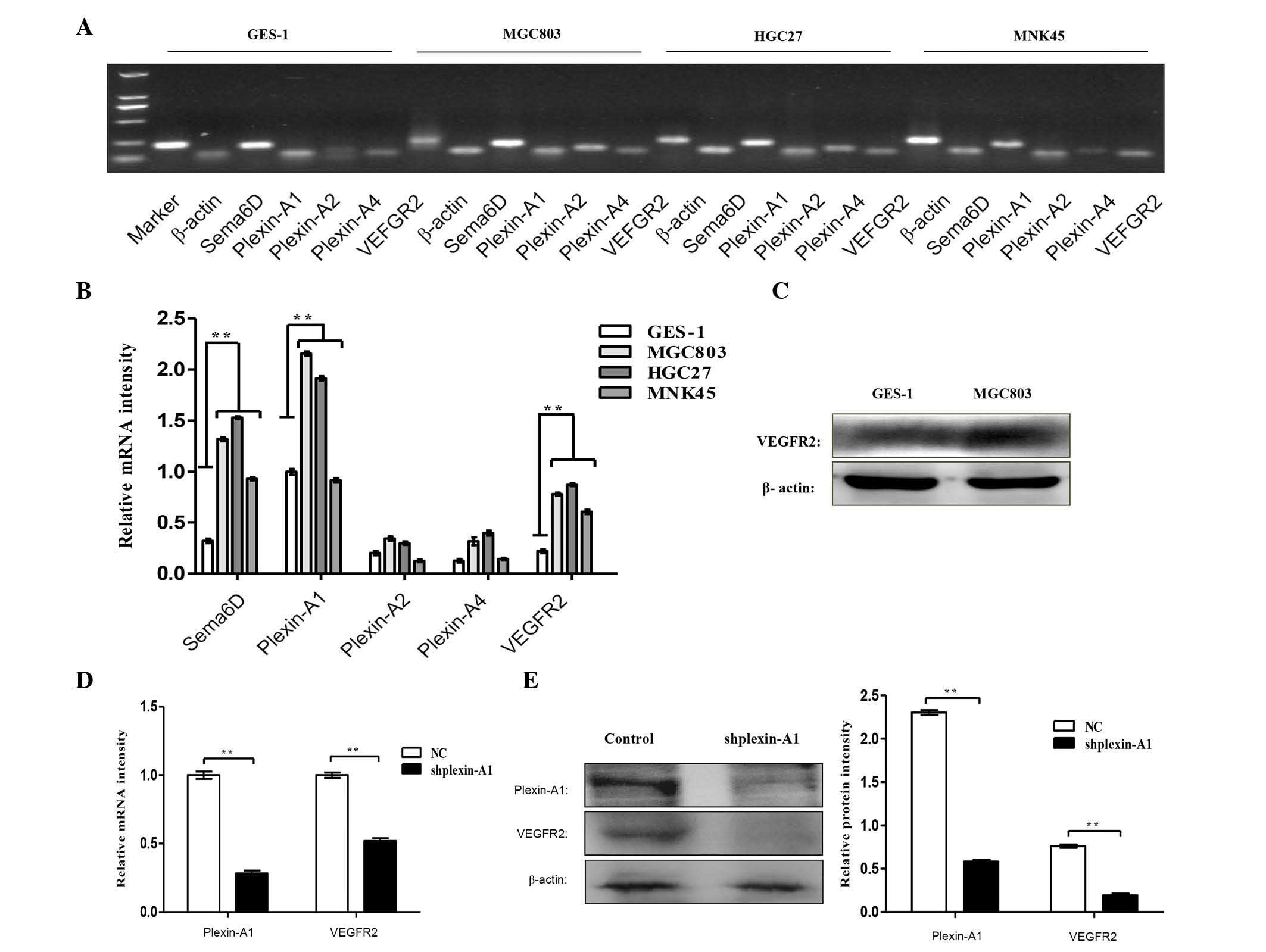 | Figure 6.Sema6D and plexin-A1 promote
angiogenesis through VEGFR2 signaling. (A) RT-PCR analysis detected
the mRNA expression levels of Sema6D, plexin-As and VEGFR2 in three
gastric cancer cell lines and GES-1 cells. (B) The histogram
indicates the relative mRNA expression levels, which were
normalized to the levels of β-actin. (C) Western blot analysis
demonstrated that the protein expression levels of VEGFR2 were
remarkably enhanced in MGC803 gastric cancer cells compared with
GES-1 cells. (D) RT-quantitative PCR analysis of the mRNA
expression levels of plexin-A1 and VEGFR2 upon using shplexin-A1
transfection to knock down plexin-A1. The mRNA expression level of
VEGFR2 was downregulated. (E) Western blot analysis detected the
protein levels of plexin-A1 and VEGFR2, and the protein expression
of VEGFR2 was decreased with targeted disruption of plexin-A1. (F)
The relative protein expression levels of plexin-A1 and VEGFR2 is
shown. Data are represented as the mean ± standard deviation (n=3
for each group, **P<0.01). Sema, semaphorin; VEGFR, vascular
endothelial growth factor receptor; mRNA, messenger RNA; NC,
negative control; sh, short hairpin; RT-PCR, reverse
transcription-polymerase chain reaction. |
Discussion
Gastric cancer is the fifth most common cancer and
the third leading cause of cancer mortality worldwide, with nearly
950,000 new cases and 723,000 mortalities estimated in 2012
(29). Angiogenesis involves the
formation of new blood vessels, which serves a pivotal role in
tumor growth and metastasis (30).
Anti-angiogenic therapies targeting VEGF and its receptors have
been developed in recent years (31).
However, in gastric cancer, the therapeutic role of anti-angiogenic
agents remains to be determined, particularly for patients with
advanced disease, where treatment options are limited.
Despite the fact that Sema6D and plexin-A1 serve a
crucial role in the formation of the nervous system, increasing
evidence suggests their participation in cardiogenesis (15). Ferrara and Kerbel indicated that the
action of VEGF is mainly mediated through VEGFR2, which is present
in tumor endothelial cells (32).
Therefore, the present study aimed to investigate the interaction
of Sema6D/plexin-A1 and VEGFR2.
In conclusion, the current study has provided
evidence that Sema6D and plexin-A1 may regulate angiogenesis in
vitro, and raises the possibility that they may serve a role in
tumor-induced angiogenesis by VEGFR2. Further studies will be
necessary to elucidate whether there is a secreted factor
responsible for the tumor-promotion effect of Sema6D and plexin-A1
in gastric carcinoma, such as VEGFR2, and to determine whether
other members of the semaphorin family and their receptors can
function as tumor stimulators or suppressors in vivo.
Understanding the function of Sema6D and plexin-A1
in the regulation of malignant transformation may aid to unravel
the molecular mechanisms involved in gastric cancer, which may open
novel therapeutic avenues to interfere with this process. Our
studies indicated a direct connection between Sema6D, plexin-A1 and
gastric cancer angiogenesis, and may open novel therapeutic avenues
to interfere with this process. Therefore, future clinical trials
may be undertaken with the blockage of the Sema6D/plexin-A1 pathway
for the treatment of gastric cancer regarding angiogenesis, which
serves an important pathogenic role.
Acknowledgements
The present study was supported by the Key Subjects
in Universities and Colleges of Hebei Province of China (Pathology
and Pathophysiology; Shijiazhuang, China) and the Science and
Technology Support Program of Hebei Province of China
(Shijiazhuang, China; grant no. 13277779D).
References
|
1
|
Kolodkin AL, Matthes DJ and Goodman CS:
The semaphorin genes encode a family of transmembrane and secreted
growth cone guidance molecules. Cell. 75:1389–1399. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Takahashi K, Ishida M, Hirokawa K and
Takahashi H: Expression of the semaphorins Sema 3D and Sema 3F in
the developing parathyroid and thymus. Dev Dyn. 237:1699–1708.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Behar O, Golden JA, Mashimo H, Schoen FJ
and Fishman MC: Semaphorin III is needed for normal patterning and
growth of nerves, bones and heart. Nature. 383:525–528. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gitler AD, Lu MM and Epstein JA: PlexinD1
and semaphorin signaling are required in endothelial cells for
cardiovascular development. Dev Cell. 7:107–116. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gu C, Rodriguez ER, Reimert DV, Shu T,
Fritzsch B, Richards LJ, Kolodkin AL and Ginty DD: Neuropilin-1
conveys semaphorin and VEGF signaling during neural and
cardiovascular development. Dev Cell. 5:45–57. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Suzuki K, Kumanogoh A and Kikutani H:
Semaphorins and their receptors in immune cell interactions. Nat
Immunol. 9:17–23. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tse C, Xiang RH, Bracht T and Naylor SL:
Human Semaphorin 3B (SEMA3B) located at chromosome 3p21.3
suppresses tumor formation in an adenocarcinoma cell line. Cancer
Res. 62:542–546. 2002.PubMed/NCBI
|
|
8
|
Tomizawa Y, Sekido Y, Kondo M, Gao B,
Yokota J, Roche J, Drabkin H, Lerman MI, Gazdar AF and Minna JD:
Inhibition of lung cancer cell growth and induction of apoptosis
after reexpression of 3p21.3 candidate tumor suppressor gene
SEMA3B. Proc Natl Acad Sci USA. 98:13954–13959. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Miao HQ, Soker S, Feiner L, Alonso JL,
Raper JA and Klagsbrun M: Neuropilin-1 mediates
collapsin-1/semaphorin III inhibition of endothelial cell motility:
Functional competition of collapsin-1 and vascular endothelial
growth factor-165. J Cell Biol. 146:233–342. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kigel B, Varshavsky A, Kessler O and
Neufeld G: Successful inhibition of tumor development by specific
class-3 semaphorins is associated with expression of appropriate
semaphorin receptors by tumor cells. PLoS One. 3:e32872008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Banu N, Teichman J, Dunlap-Brown M,
Villegas G and Tufro A: Semaphorin 3C regulates endothelial cell
function by increasing integrin activity. FASEB J. 20:2150–2152.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Basile JR, Castilho RM, Williams VP and
Gutkind JS: Semaphorin 4D provides a link between axon guidance
processes and tumor-induced angiogenesis. Proc Natl Acad Sci USA.
103:9017–9022. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao XY, Chen L, Xu Q and Li YH:
Expression of semaphorin 6D in gastric carcinoma and its
significance. World Gastroenterol. 12:7388–7390. 2006. View Article : Google Scholar
|
|
14
|
O'Connor BP, Eun SY, Ye Z, Zozulya AL,
Lich JD, Moore CB, Iocca HA, Roney KE, Holl EK, Wu QP, et al:
Semaphorin 6D regulates the late phase of CD4+ T cell primary
immune responses. Proc Natl Acad Sci USA. 105:13015–13020. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Toyofuku T, Zhang H, Kumanogoh A,
Takegahara N, Suto F, Kamei J, Aoki K, Yabuki M, Hori M, Fujisawa H
and Kikutani H: Dual roles of Sema6D in cardiac morphogenesis
through region-specific association of its receptor, Plexin-A1,
with off-track and vascular endothelial growth factor receptor type
2. Genes Dev. 18:435–447. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Moriarity BS, Otto GM, Rahrmann EP, Rathe
SK, Wolf NK, Weg MT, Manlove LA, LaRue RS, Temiz NA, Molyneux SD,
et al: A Sleeping Beauty forward genetic screen identifies new
genes and pathways driving osteosarcoma development and metastasis.
Nat Genet. 47:615–624. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Toyofuku T, Zhang H, Kumanogoh A,
Takegahara N, Yabuki M, Harada K, Hori M and Kikutani H: Guidance
of myocardial patterning in cardiac development by Sema6D reverse
signalling. Nat Cell Biol. 6:1204–1211. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kigel B, Rabinowicz N, Varshavsky A,
Kessler O and Neufeld G: Plexin-A4 promotes tumor progression and
tumor angiogenesis by enhancement of VEGF and bFGF signaling.
Blood. 118:4285–4296. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao XY, Chen L, Li YH and Xu Q: PlexinA1
expression in gastric carcinoma and its relationship with tumor
angiogenesis and proliferation. World J Gastroenterol.
13:6558–6561. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Catalano A, Lazzarini R, Di Nuzzo S,
Orciari S and Procopio A: The plexin-A1 receptor activates vascular
endothelial growth factor-receptor 2 and nuclear factor-kappaB to
mediate survival and anchorage-independent growth of malignant
mesothelioma cells. Cancer Res. 69:1485–1493. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Suzuki K, Kumanogoh A and Kikutani H:
Semaphorins and their receptors in immune cell interactions. Nat
Immunol. 9:17–23. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fujii T, Nakao F, Shibata Y, Shioi G,
Kodama E, Fujisawa H and Takagi S: Caenorhabditis elegans PlexinA,
PLX-1, interacts with transmembrane semaphorins and regulates
epidermal morphogenesis. Development. 129:2053–2063.
2002.PubMed/NCBI
|
|
24
|
Herzog Y, Kalcheim C, Kahane N, Reshef R
and Neufeld G: Differential expression of neuropilin-1 and
neuropilin-2 in arteries and veins. Mech Dev. 109:115–119. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ellis LM: The role of neuropilins in
cancer. Mol Cancer Ther. 5:1099–1107. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Peplow PV: Influence of growth factors and
cytokines on angiogenic function of endothelial progenitor cells: A
review of in vitro human studies. Growth Factors. 32:83–116. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Meunier-Carpentier S, Dales JP, Djemli A,
Garcia S, Bonnier P, Andrac-Meyer L, Lavaut MN, Allasia C and
Charpin C: Comparison of the prognosis indication of VEGFR-1 and
VEGFR-2 and Tie2 receptor expression in breast carcinoma. Int J
Oncol. 26:977–984. 2005.PubMed/NCBI
|
|
28
|
Rahimi N: VEGFR-1 and VEGFR-2: Two
non-identical twins with a unique physiognomy. Front Biosci.
11:818–829. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang ZL, Liu ZS and Sun Q: Effects of
thalidomide on angiogenesis and tumor growth and metastasis of
human hepatocellular carcinoma in nude mice. World J Gastroenterol.
11:216–220. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bertolini F, Marighetti P, Martin-Padura
I, Mancuso P, Hu-Lowe DD, Shaked Y and D'Onofrio A: Anti-VEGF and
beyond: Shaping a new generation of anti-angiogenic therapies for
cancer. Drug Discov Today. 16:1052–1060. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ferrara N and Kerbel RS: Angiogenesis as a
therapeutic target. Nature. 438:967–974. 2005. View Article : Google Scholar : PubMed/NCBI
|