Introduction
Gastric cancer is the fourth most frequently
diagnosed cancer and the second leading cause of cancer-associated
mortality worldwide (1,2). Despite recent advances in adjuvant and
neo-adjuvant therapy and improved understanding of gastric cancer
biology, progress in the treatment of gastric cancer has been
limited. Gastric cancer has a high recurrence rate and >50% of
patients succumb to this disease within 3 years of diagnosis
(3), highlighting the requirement for
novel treatment regimens (4).
The use of natural products in gastric cancer
therapy has been the focus of a number of studies (5–9); licorice
is a traditional herbal medicine that originates from the licorice
root, and is one of the most commonly prescribed herbs in China for
the treatment of various diseases, including microbial infections
and cancer (10,11). Licochalcone A (LCA;
C21H22O4; molecular weight, 338.4)
is the primary active compound in the licorice species
Glycyrrhiza glabra and is an estrogenic flavonoid with
antiparasitic, antibacterial and antitumor properties (12–14). It
has been demonstrated previously that LCA is the most cytotoxic
licorice compound, and is able to inhibit the growth of gastric
cancer cells by arresting cell cycle progression and inducing
apoptosis (15). The current standard
chemotherapeutic regimen for gastric cancer is 5-fluorouracil
(5-FU)-based combined chemotherapy, and various herbal extracts are
widely used for the treatment of gastric cancer, including
paclitaxel and curcumin (16,17). In addition, in cases where a novel
therapeutic is used for the treatment of gastric cancer, 5-FU is
frequently used as a combination therapy (18). In the present study, the inhibitory
effects of LCA on gastric cancer cells alone and in combination
with 5-FU were evaluated.
Materials and methods
Cell culture and reagents
The SGC7901 (moderately differentiated) and MKN-45
(poorly differentiated) human gastric cancer cell lines were
purchased from the American Type Culture Collection (Manassas, VA,
USA) and cultured in RPMI-1640 (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Thermo Fisher Scientific, Inc), 1% Gibco® Glutamax™
(Thermo Fisher Scientific, Inc.) and 1% penicillin and
streptomycin, and maintained in an incubator at 37°C with a
humidified atmosphere containing 5% CO2.
Cell proliferation assay
SGC7901 and MKN-45 cells were seeded in 96-well
plates at a density of 1,000 cells/well with 100 µl complete
culture medium. Following adhesion for 24 h, cells were treated
with LCA (Phytomarker Ltd., Tianjin, China) or LCA plus 5-FU
(Shanghai Xudong Haipu Pharmaceutical Co., Ltd., Shanghai, China)
at various concentrations (diluted to 15.625, 31.25, 62.5, 125 and
187.5 µg/ml in complete medium). In a previous study (15), the selected concentration of LCA was
25 µM. Cells that were not exposed to LCA or 5-FU were used as
negative controls (blank). Following LCA or LCA plus 5-FU
treatment, the supernatant was removed, 100 µl RPMI-1640 medium
containing 10 µl Cell Counting Kit-8 (CCK8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was added to each well and the
cells were incubated at 37°C for 3 h. The culture plates were then
agitated for 10 min at room temperature 20°C and optical density
(OD) values were read at 450 nm using a microplate reader
(SpectraMax 190; Molecular Devices, LLC, Sunnyvale, CA, USA).
Cell cycle analysis
Cells (3×105) were plated in RPMI-1640
medium and then treated with 25 µM LCA and 15.625 µg/ml LCA plus
5-FU. The cells were harvested at 0 and 48 h, suspended in 300 µl
PBS, mixed with cold ethanol (700 µl) and incubated at 4°C
overnight. Following centrifugation at 4°C 1,000 × g for 5
min, the pellet was washed with cold PBS, resuspended in 500 µl PBS
and incubated with 50 µl RNase (final concentration, 20 µg/ml) at
4°C for 30 min. Subsequently, the cells were incubated with
propidium iodide (final concentration, 50 µg/ml) at 4°C for 30 min
in the dark. The cell cycle distribution was then determined using
a FACSAria Flow Cytometer (BD Biosciences, Franklin Lakes, NJ,
USA).
Apoptosis analysis
LCA and LCA plus 5-FU-induced apoptosis in SGC7901
and MKN-45 cells was determined using flow cytometry with the
Annexin V-fluorescein isothiocyanate (FITC) Apoptosis Detection kit
(BioVision, Inc., Milpitas, CA, USA) according to the
manufacturer's protocol. Briefly, 3×105 cells were
plated and treated with 25 µM LCA or 15.625 µg/ml LCA plus 5-FU for
24, 48 and 72 h. To prepare for staining analysis, cells were
harvested, washed in PBS and incubated with Annexin V and propidium
iodide in binding buffer at room temperature for 10 min in the
dark. The stained cells were analyzed using a FACSAria flow
cytometer.
Western blot analysis
SGC7901 and MKN-45 whole cell lysates were produced
using radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Shanghai, China) to lyse the cells, followed by
centrifugation at 4°C and 12,000 × g for 5 min and
collection of the supernatant fraction for immunoblotting. The
proteins (50 µg) were separated using 5–10% SDS-PAGE and
transferred onto a nitrocellulose membrane. Following blocking with
5% nonfat milk in blocking buffer (20 mM Tris-buffered saline
containing 0.1% Tween-20; pH 7.5), the membranes were incubated
with primary antibody at 4°C overnight and then incubated at room
temperature for 1 h with the appropriate horseradish
peroxidase-conjugated secondary antibody. The primary antibodies
were anti-B-cell-lymphoma-2 (Bcl-2; #ab7973; Abcam, Cambridge, UK),
anti-Bcl-2-associated X protein (Bax; #2772; Cell Signaling
Technology, Inc., Danvers, MA, USA), anti-caspase 3 (#ab44976;
Abcam), anti-poly ADP ribose polymerase (PARP; #AP102-1; Beyotime
Institute of Biotechnology), anti-tumor protein (p)53 (#op43T;
Merck Millipore, Darmstadt, Germany), anti-p27 (#2747-1; Epitomics,
Burlingame, CA, USA), anti-p21 (#ab109520; Abcam), anti-mouse
double minute 2 homolog (MDM2; #ab16895, Abcam) and anti-GAPDH
(#AP0063; Bioworld Technology Inc., St. Louis Park, MN, USA). GAPDH
antibodies were diluted to 1:5,000 in blocking solution. The
dilutions for all other primary antibodies were as follows: Caspase
3 (dilution, 1:1,000), Bcl-2, Bax, p21, p27, p53, PARP (dilution,
1:500) and MDM2 (dilution, 1:50). The secondary antibodies included
horseradish peroxidase-linked anti-mouse immunoglobulin G
(dilution, 1:5,000; sc-358914) and anti-rabbit immunoglobulin G
(dilution, 1:4,000; sc-2357) (Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) at room temperature for 1 h. The immunoreactive
bands were visualized using the Enhanced Chemiluminescence plus
Western Blotting Detection system, which was purchased from Bio-Rad
Laboratories, Inc. (Hercules, CA, USA). GAPDH was used as a loading
control for each sample.
Statistical analysis
The experimental data were all expressed as the mean
± standard deviation, and statistical differences between groups
were evaluated with a two-tailed Student's t-test using SPSS
version 13 (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
LCA induces inhibition of gastric
cancer cell proliferation
The cytotoxicity of LCA and LCA plus 5-FU against
the SGC7901 and MKN-45 gastric cancer cell lines was evaluated.
Cell proliferation was observed at 24, 48 and 72 h following
treatment. MKN45 cells were moderately sensitive to 5-FU, and
SGC7901 cells were relatively resistant to 5-FU. Therefore, the
most appropriate concentration of 5-FU was determined as 15.625
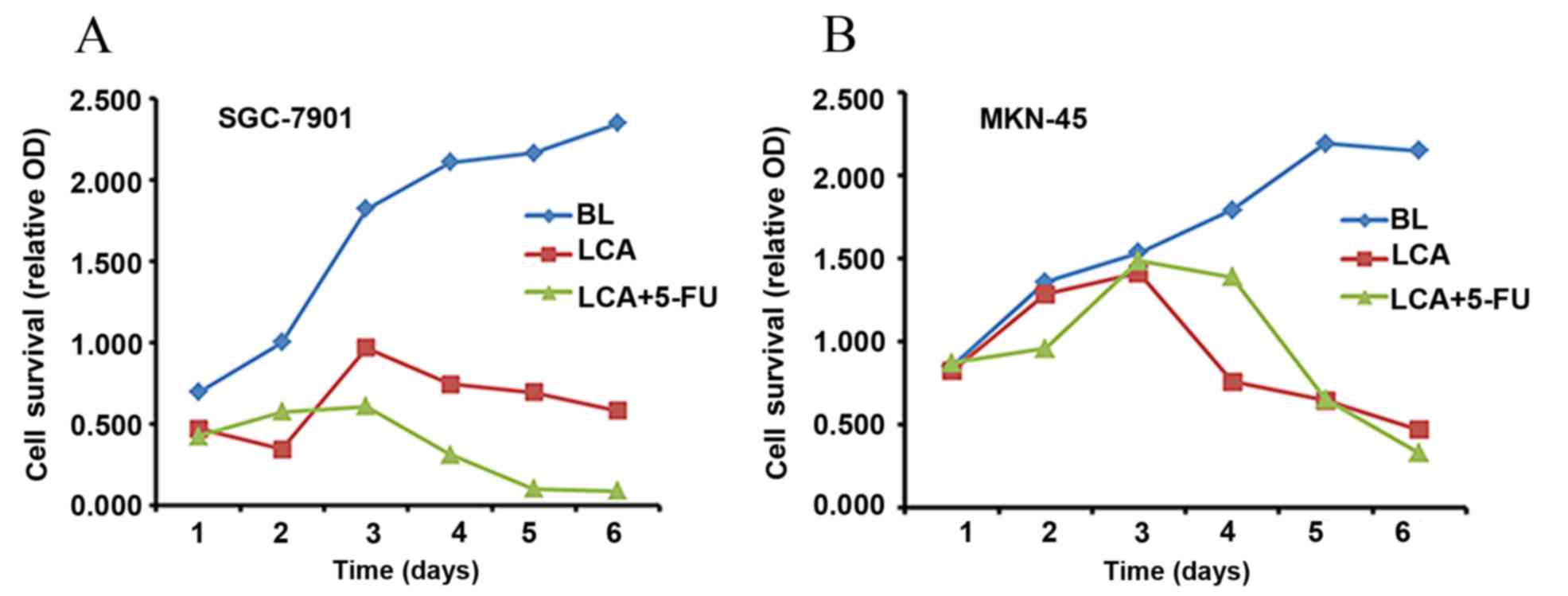
µg/ml. Fig. 1 presents the relative
OD values of SGC-7901 and MKN-45 cells following exposure to the
blank (BL; negative control), LCA and LCA plus 5-FU. Compared with
the control group, cancer cell proliferation was significantly
inhibited by LCA alone (SGC7901, P=0.011; MKN-45, P=0.016) and LCA
in combination with 5-FU (SGC7901, P=0.004; MKN-45, P=0.029). This
inhibitory effect was more pronounced in the combination group,
compared with LCA alone (Fig. 1).
LCA-induces G2/M phase
arrest and 5-FU induces G0/G1 phase arrest in
human gastric cancer cells
The results from a previous study revealed that LCA
may induce G2/M phase arrest by affecting the levels of
cyclin A, cyclin B, retinoblastoma and MDM2 (9). The present study further examined the
effects of LCA alone or in combination with 5-FU on the cell cycle
distribution of gastric cancer cells. The cell cycle distribution
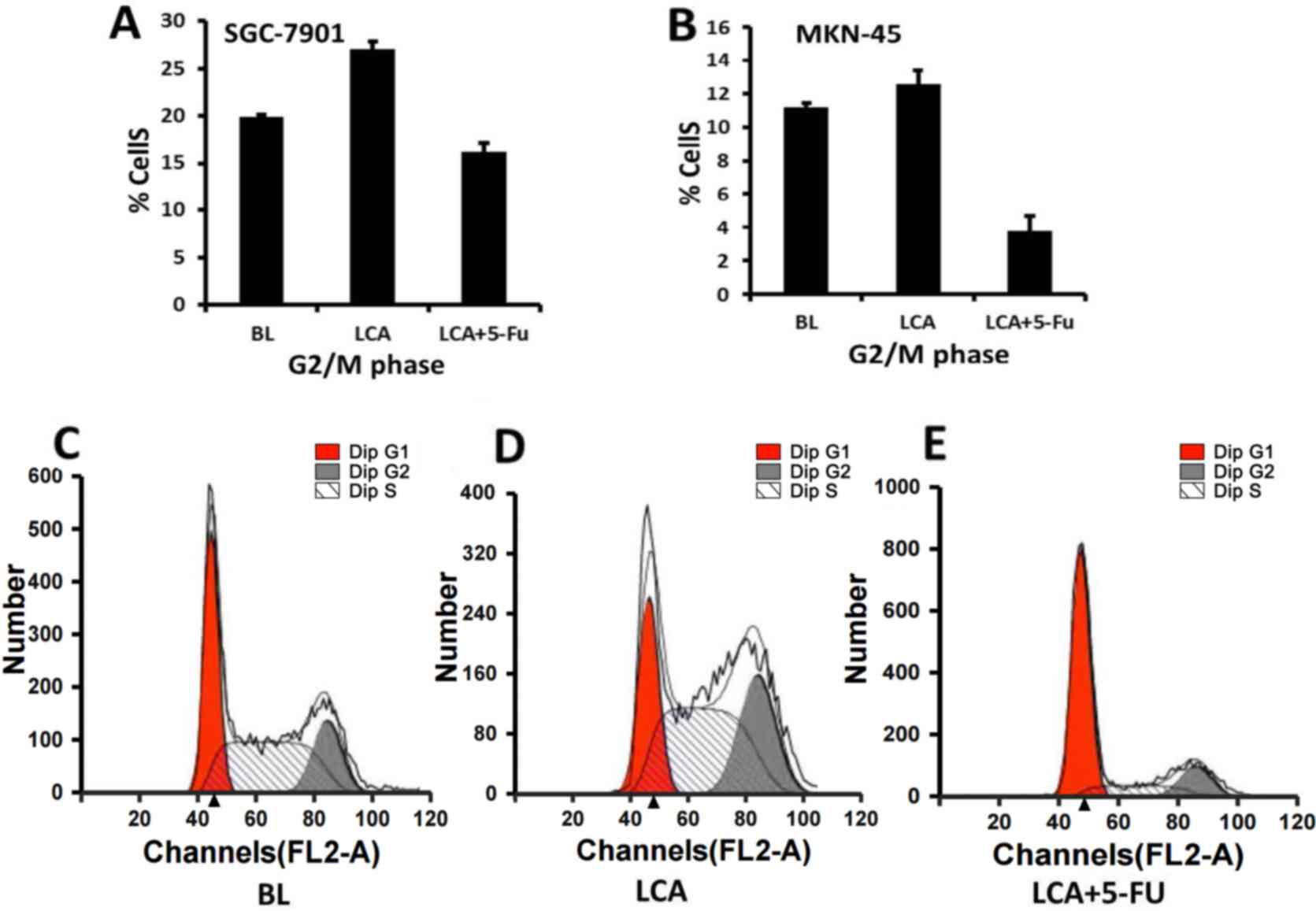
of the cells following exposure to LCA or LCA plus 5-FU for 48 h is
presented in Table I. The SGC-7901
cells treated with LCA exhibited significant G2/M phase
arrest compared with the cells treated with LCA plus 5-FU
(27.04±0.81 vs. 16.25±0.90%, respectively; P<0.001; Fig. 2A) or the blank cells (27.04±0.81 vs.
19.87±0.23%, respectively; P<0.001). A higher proportion of the
SGC7901 cells treated with LCA plus 5-FU exhibited
G0/G1 arrest compared with the blank cells
(67.67±0.77 vs. 37.66±0.46%, respectively; P<0.001) or LCA alone
(67.67±0.77 vs. 26.32±0.77%, respectively; P<0.001).
G2/M phase arrest was also observed in the MKN-45 cells
treated with LCA and with LCA and 5-FU in combination (12.58±0.77
vs. 3.79±0.33%, respectively; P<0.001; Fig. 2B) for 48 h. The MKN-45 cells treated
with LCA also exhibited increased G2/M arrest compared
with the blank cells (12.58±0.77 vs. 11.21±0.68%, respectively);
however, this result was not statistically significant (P=0.084).
By contrast, the MKN-45 cells treated with LCA plus 5-FU exhibited
a significantly lower frequency of G0/G1
arrest compared with the blank cells (50.60±1.62 vs. 63.21±0.84%,
respectively; P<0.001) or LCA alone (50.60±1.62 vs. 68.12±0.94%,
respectively; P<0.001). Fig. 2C-E
present the cell cycle distribution of the SGC7901 cells following
exposure to the blank (Fig. 2C), LCA
(Fig. 2D) and LCA combined with 5-FU
(Fig. 2E). These data suggest that
LCA-induced growth inhibition in gastric cancer cells may be
associated with the induction of G2/M arrest in the two
gastric cancer lines, whereas 5-FU inhibited cell proliferation by
inducing cell cycle arrest during the G0/G1
phase in the moderately differentiated SGC-7901 cells. This result
is concordant with a previous study, which demonstrated that 5-FU
was able to induce G0/G1 arrest in tumor
cells (19).
 | Table I.Cell cycle distribution of SGC-7901
and MKN-45 human gastric cancer cells following exposure to LCA or
LCA plus 5-FU for 48 h. |
Table I.
Cell cycle distribution of SGC-7901
and MKN-45 human gastric cancer cells following exposure to LCA or
LCA plus 5-FU for 48 h.
| Cell line | Cell cycle
stage | BL, (mean ±
SD)% | LCA, (mean ±
SD)% | LCA+5-FU, (mean ±
SD)% |
|---|
| SGC-7901 |
G0/G1 | 37.66±0.46 | 26.32±0.77 | 67.67±0.77 |
|
| S | 42.47±0.63 | 46.65±0.57 | 16.08±0.51 |
|
|
G2/M | 19.87±0.23 | 27.04±0.81 | 16.25±0.90 |
| MKN-45 |
G0/G1 | 63.21±0.84 | 68.12±0.94 | 50.60±1.62 |
|
| S | 25.58±1.43 | 19.31±0.34 | 45.61±1.88 |
|
|
G2/M | 11.21±0.68 | 12.58±0.77 |
3.79±0.33 |
LCA and its combination with 5-FU
induces apoptosis in human gastric cancer cells
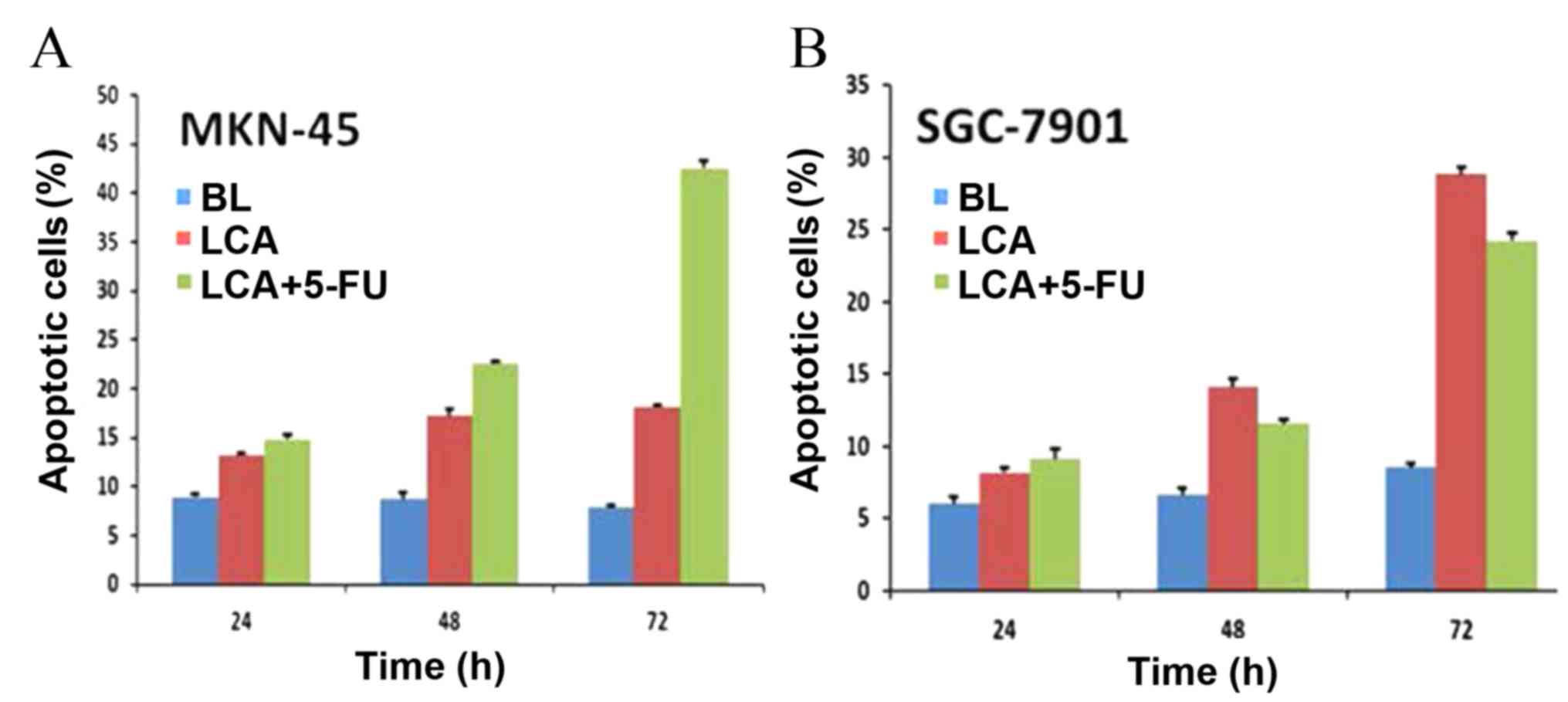
To determine whether LCA is able to inhibit growth
in gastric cancer cells by inducing apoptosis, SGC7901 and MKN-45
cells were treated with LCA (25 μM) and LCA plus 5-FU (15.625
µg/ml) as aforementioned. Apoptotic cell numbers were assessed
using an Annexin V-FITC Apoptosis Detection kit. The results
revealed that exposure of MKN-45 cells to LCA resulted in a
significant increase of apoptotic cells in a time-dependent manner
(13.27±0.24% apoptotic cells at 24 h, 17.21±0.65% at 48 h and
18.16±0.22% at 72 h; P<0.001). The exposure of MKN-45 cells to
LCA plus 5-FU also significantly increased the number of apoptotic
cells over time (14.88±0.39% apoptotic cells at 24 h, 22.62±0.13%
at 48 h and 42.55±0.72% at 72 h; P<0.001), and more apoptotic
cells were detected when the MKN-45 cells were exposed to LCA plus
5-FU as compared with LCA treatment alone (Fig. 3A). The SGC-7901 cells also exhibited
dose-dependent apoptosis when exposed to LCA or LCA plus 5-FU;
however, this varied from the results observed in MKN-45 cells.
SGC-7901 cells exposed to LCA alone exhibited higher levels of
apoptosis compared with those treated with LCA plus 5-FU at 48 and
72 h (14.07±0.56 vs. 11.56±0.26% at 48 h, P<0.001; 28.82±0.48
vs. 24.23±0.51% at 72 h, P<0.001) (Fig. 3B). This may be due to the relative
resistance of SGC7901 cells to 5-FU (20). The number of apoptotic cells was
markedly higher in cell samples treated with LCA compared with the
blank controls (6±0.47 vs. 8.19±0.30% apoptotic cells at 24 h,
6.62±0.47 vs. 14.07±0.56% at 48 h and 8.52±0.24 vs. 28.82±0.48% at
72 h; P<0.001). This effect was more pronounced in cell samples
treated with LCA plus 5-FU, except for SGC-7901 cells, which were
relatively resistant to 5-FU. These results indicated that LCA was
able to effectively induce apoptosis in two gastric cancer cell
lines and that combining LCA with 5-FU induced more apparent
apoptosis.
Effects of LCA on the expression
levels of apoptosis and cell cycle-associated proteins in gastric
cancer cells
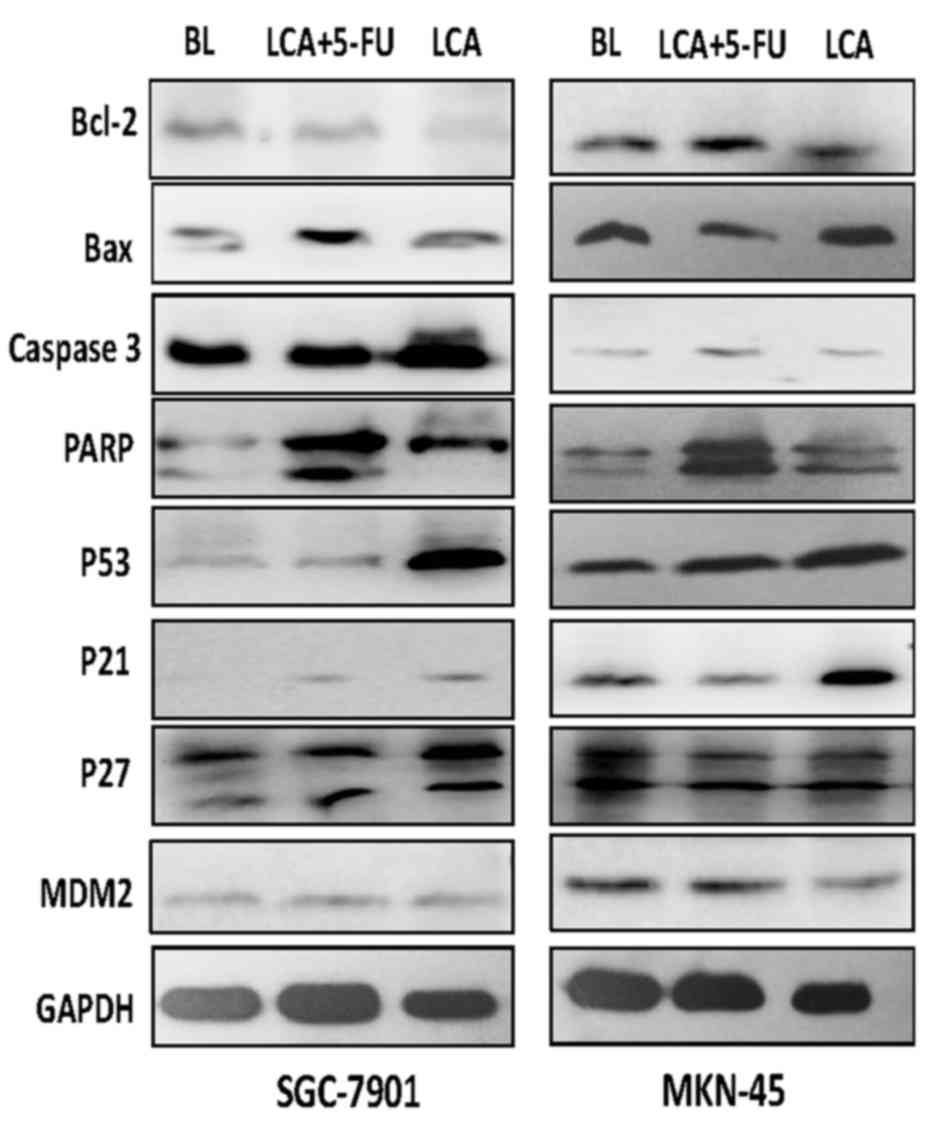
The Bcl-2 family proteins are important in the
regulation of apoptosis (21). A
previous study revealed that LCA induces a dose-dependent increase
in the expression levels of Bax and a decrease in the levels of
Bcl-2 (15). The current study
further analyzed the levels of Bcl-2 and Bax in cells treated with
blank control, LCA and LCA plus 5-FU. Exposure of the SGC-7901
cells to LCA resulted in a significant increase (P<0.001) in the
expression levels of Bax protein and decreased levels of Bcl-2. A
similar result was observed when SGC-7901 cells were exposed to LCA
plus 5-FU, and the increase in Bax expression levels was more
marked. The protein expression levels of Bax and Bcl-2 were similar
to the blank control group following exposure to LCA or LCA plus
5-FU in the MKN-45 cells (Fig.
4).
Caspase-3, an executioner caspase that is activated
by caspase-9, cleaves a broad spectrum of cellular target proteins,
including nuclear PARP, which leads to a cell-death cascade
(22). To clarify the mechanism
underlying the apoptotic signaling pathway activation mediated by
LCA and 5-FU, the effects of LCA and LCA plus 5-FU on the
activation of caspase-3 and PARP were examined. The gastric cancer
cells that were exposed to LCA exhibited increased PARP cleavage. A
trend of increased caspase-3 expression levels was observed in
SGC-7901 cells, and to a lesser degree in MKN-45 cells. The
increase of PARP and caspase 3 expression levels in gastric cancer
cells that were exposed to LCA plus 5-FU was more marked compared
with cells treated with LCA alone (Fig.
4). This suggests that the canonical mitochondrial apoptotic
signaling pathway is activated by LCA and that 5-FU may enhance the
effects of LCA.
As p21 and p27 are important in the regulation of
the cell cycle (23), the effects of
LCA and 5-FU on the expression levels of these proteins were
examined. As presented in Fig. 4,
treatment with LCA resulted in an increase in the expression levels
of p21 protein in the SGC-7901 and MKN-45 gastric cancer cell
lines. However, no statistically different change was observed in
p27 expression levels following treatment with LCA, but there
remained a trend of increase in SGC-7901 cells. These observations
suggest that increased p21 and p27 expression levels be important
for LCA-induced G2/M phase arrest in human gastric
cancer cells. P53 and MDM2 also serve essential roles in the
regulation of the cell cycle (23).
Treatment with LCA alone or LCA plus 5-FU resulted in increased p53
protein expression levels (Fig. 4),
suggesting that LCA may affect the levels of p53 in gastric cancer
cells.
Discussion
The roots and rhizomes of licorice
(Glycyrrhiza) species have been used globally as a natural
sweetener and herbal medicine. Licorice root is a traditional
medicine used primarily for the treatment of hepatitis C, peptic
ulcers, and pulmonary and skin diseases (24). LCA is a natural phenol licorice
compound isolated from the roots of Glycyrrhiza with
numerous biological functions, including anti-angiogenesis,
antiparasitic, antioxidant, antibacterial and anti-inflammatory
activities (25). In the current
study, the anti-cancer effects of LCA alone and in combination with
5-FU on gastric cancer cells were evaluated.
Gastric cancer is an aggressive disease with a poor
prognosis, even when the lesions are completely resected without
distant metastasis (26). A
meta-analysis of phase 2 and 3 randomized gastric cancer trials
demonstrated that combination chemotherapy improves the overall
survival rates compared with single-agent chemotherapy or best
supportive care (1,27). However, the resulting toxicity
remained significant and the survival benefit was modest (4). Therefore, the potential use of natural
products in the treatment of gastric cancer has been the focus of
previous studies (5–9). In the present study, LCA was able to
inhibit cell proliferation, block cell cycle progression at the
G2/M transition and induce apoptosis in gastric cancer
cells. Thus, LCA may be a potential novel therapeutic agent for
gastric cancer. In addition, 5-FU induced
G0/G1 arrest in gastric cancer lines and
enhanced the effects of growth inhibition and apoptosis, and may
therefore be used as a combination treatment.
The dysregulation of the cell cycle is correlated
with the process of tumor development (28). Cyclin-dependent kinases (CDKs) are key
regulators of the cell cycle (29).
This process also involves positive and negative regulatory
factors, including the cyclin dependent kinase inhibitors (CKIs)
p21 and p27 (30). p53 is activated
in response to DNA damage and activates p21, which binds to and
inhibits several CDKs resulting in cell cycle arrest and the
induction of apoptosis in invading tumors (31). p27 is activated by TGF-β and also
inhibits several CDKs, whereas MDM2 is a negative regulator of p53
and triggers cell cycle progression by reversing p53-mediated cell
cycle arrest and apoptosis (32).
Cyclins and CDKs are overexpressed and dysregulated in a variety of
types of human cancer and p21 and p27 are frequently mutated and
exhibit reduced expression levels (33). Human tumor formation often involves
the deactivation of the tumor suppressor p53 and overexpression of
the oncogene MDM2 (34,35). The present study demonstrated that LCA
treatment was able to increase the levels of p21, p27 and p53 and
decrease the levels of MDM2 in gastric cancer cells, an effect that
was amplified by treatment with LCA in combination with 5-FU.
The impairment of apoptosis (programmed cell death)
is important in the pathogenesis and progression of cancer, and is
a major barrier to effective treatment (27). Apoptosis is regulated by members of
the Bcl-2 family that function upstream of the interleukin-1
converting enzyme family of cysteine proteases, also termed
caspases (36,37). During the induction of apoptosis,
caspase proteases are sequentially activated and subsequently
cleave several cellular substrates, including PARP, a nuclear
enzyme involved in DNA repair, maintenance of genome integrity and
post-translational modification of protein ribosylation (38). The current study demonstrated that LCA
treatment increased the levels of Bax, caspase3 and cleaved-PARP
and decreased the levels of Bcl-2 in gastric cancer cells. In
addition, LCA may function synergistically with 5-FU.
In conclusion, the present study demonstrated that
LCA exhibits antitumor activity in vitro by inhibiting cell
proliferation, arresting cell cycle progression and inducing
apoptosis in gastric cancer cells. 5-FU may enhance the antitumor
effects of LCA, and further studies on the antitumor activity of
LCA anti-tumor activity in vivo are required.
Acknowledgements
This study was supported by The National Natural
Science Foundation of China (grant no. 81101648). The authors
acknowledge American Journal Experts for the English language
editing revision of the original manuscript.
References
|
1
|
Bang YJ, Van Cutsem E, Feyereislova A,
Chung HC, Shen L, Sawaki A, Lordick F, Ohtsu A, Omuro Y, Satoh T,
et al: Trastuzumab in combination with chemotherapy versus
chemotherapy alone for treatment of HER2-positive advanced gastric
or gastro-oesophageal junction cancer (ToGA): A phase 3,
open-label, randomised controlled trial. Lancet. 376:687–697. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kamangar F, Dores GM and Anderson WF:
Patterns of cancer incidence, mortality, and prevalence across five
continents: Defining priorities to reduce cancer disparities in
different geographic regions of the world. J Clin Oncol.
24:2137–2150. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zare A, Mahmoodi M, Mohammad K, Zeraati H,
Hosseini M and Naieni K Holakouie: Factors affecting the survival
of patients with gastric cancer undergone surgery at iran cancer
institute: Univariate and multivariate analyses. Iran J Public
Health. 43:800–808. 2014.PubMed/NCBI
|
|
4
|
Schwartz GK, Winter K, Minsky BD, Crane C,
Thomson PJ, Anne P, Gross H, Willett C and Kelsen D: Randomized
phase II trial evaluating two paclitaxel and cisplatin-containing
chemoradiation regimens as adjuvant therapy in resected gastric
cancer (RTOG-0114). J Clin Oncol. 27:1956–1962. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zan X, Cui F, Li Y, Yang Y, Wu D, Sun W
and Ping L: Hericium erinaceus polysaccharide-protein HEG-5
inhibits SGC-7901 cell growth via cell cycle arrest and apoptosis.
Int J Biol Macromol. 76:242–253. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rasul A, Yu B, Khan M, Zhang K, Iqbal F,
Ma T and Yang H: Magnolol, a natural compound, induces apoptosis of
SGC-7901 human gastric adenocarcinoma cells via the mitochondrial
and PI3K/Akt signaling pathways. Int J Oncol. 40:1153–1161.
2012.PubMed/NCBI
|
|
7
|
Lee Y: Induction of apoptosis by
S-allylmercapto-L-cysteine, a biotransformed garlic derivative, on
a human gastric cancer cell line. Int J Mol Med. 21:765–770.
2008.PubMed/NCBI
|
|
8
|
Ma K, Liu Y, Zhu Q, Liu CH, Duan JL, Tan
BK and Zhu YZ: H2S donor, S-propargyl-cysteine, increases CSE in
SGC-7901 and cancer-induced mice: Evidence for a novel anti-cancer
effect of endogenous H2S? PLoS One. 6:e205252011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ma J, Liu C, Chen Y, Jiang J and Qin Z:
Cellular and molecular mechanisms of the Ganoderma applanatum
extracts induces apoptosis on SGC-7901 gastric cancer cells. Cell
Biochem Funct. 29:175–182. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cuendet M, Guo J, Luo Y, Chen S, Oteham
CP, Moon RC, van Breemen RB, Marler LE and Pezzuto JM: Cancer
chemopreventive activity and metabolism of isoliquiritigenin, a
compound found in licorice. Cancer Prev Res (Phila). 3:221–232.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tamir S, Eizenberg M, Somjen D, Stern N,
Shelach R, Kaye A and Vaya J: Estrogenic and antiproliferative
properties of glabridin from licorice in human breast cancer cells.
Cancer Res. 60:5704–5709. 2000.PubMed/NCBI
|
|
12
|
Kim YH, Shin EK, Kim DH, Lee HH, Park JH
and Kim JK: Antiangiogenic effect of licochalcone A. Biochem
Pharmacol. 80:1152–1159. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee CK, Son SH, Park KK, Park JH, Lim SS,
Kim SH and Chung WY: Licochalcone A inhibits the growth of colon
carcinoma and attenuates cisplatin-induced toxicity without a loss
of chemotherapeutic efficacy in mice. Basic Clin Pharmacol Toxicol.
103:48–54. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yo YT, Shieh GS, Hsu KF, Wu CL and Shiau
AL: Licorice and licochalcone-A induce autophagy in LNCaP prostate
cancer cells by suppression of Bcl-2 expression and the mTOR
pathway. J Agric Food Chem. 57:8266–8273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xiao XY, Hao M, Yang XY, Ba Q, Li M, Ni
SJ, Wang LS and Du X: Licochalcone A inhibits growth of gastric
cancer cells by arresting cell cycle progression and inducing
apoptosis. Cancer Lett. 302:69–75. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Takeyoshi I, Makita F, Tanahashi Y,
Iwazaki S, Ogawa T, Tomizawa N, Nakamura S, Ishikawa H, Ohya T,
Kakinuma S, et al: A Phase II Study of Weekly Paclitaxel and
doxifluridine combination chemotherapy for advanced/recurrent
gastric cancer. Anticancer Res. 31:287–291. 2011.PubMed/NCBI
|
|
17
|
Cai XZ, Huang WY, Qiao Y, Du SY, Chen Y,
Chen D, Yu S, Che RC, Liu N and Jiang Y: Inhibitory effects of
curcumin on gastric cancer cells: A proteomic study of molecular
targets. Phytomedicine. 20:495–505. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ye YW, Hu S, Shi YQ, Zhang XF, Zhou Y,
Zhao CL, Wang GJ, Wen JG and Zong H: Combination of the FGFR4
inhibitor PD173074 and 5-fluorouracil reduces proliferation and
promotes apoptosis in gastric cancer. Oncol Rep. 30:2777–2784.
2013.PubMed/NCBI
|
|
19
|
Liang X, Tang J, Liang Y, Jin R and Cai X:
Suppression of autophagy by chloroquine sensitizes
5-fluorouracil-mediated cell death in gallbladder carcinoma cells.
Cell Biosci. 4:102014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang CP, Wu BH, Wang BQ, Fu MY, Yang M,
Zhou Y and Liu F: Overexpression of ECRG4 enhances chemosensitivity
to 5-fluorouracil in the human gastric cancer SGC-7901 cell line.
Tumour Biol. 34:2269–2273. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Adams JM and Cory S: The Bcl-2 protein
family: Arbiters of cell survival. Science. 281:1322–1326. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fan Y, Liu C, Huang Y, Zhang J, Cai L,
Wang S, Zhang Y, Duan X and Yin Z: Dipyrithione induces cell-cycle
arrest and apoptosis in four cancer cell lines in vitro and
inhibits tumor growth in a mouse model. BMC Pharmacol Toxicol.
14:542013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fu Z, Ren L, Wei H, Lv J, Che X, Zhu Z,
Jia J, Wang L, Lin G, Lu R and Yao Z: Effects of Tyroserleutide on
phosphatidylinositol 3′-kinase/AKT pathway in human hepatocellular
carcinoma cell. J Drug Target. 22:146–155. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
As MN and Hosseinzadeh H: Review of
pharmacological effects of Glycyrrhiza sp. and its bioactive
compounds. Phytother Res. 22:709–724. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cho JJ, Chae JI, Yoon G, Kim KH, Cho JH,
Cho SS, Cho YS and Shim JH: Licochalcone A, a natural chalconoid
isolated from Glycyrrhiza inflata root, induces apoptosis via Sp1
and Sp1 regulatory proteins in oral squamous cell carcinoma. Int J
Oncol. 45:667–674. 2014.PubMed/NCBI
|
|
26
|
Schuhmacher C, Gretschel S, Lordick F,
Reichardt P, Hohenberger W, Eisenberger CF, Haag C, Mauer ME, Hasan
B, Welch J, et al: Neoadjuvant chemotherapy compared with surgery
alone for locally advanced cancer of the stomach and cardia:
European organisation for research and treatment of cancer
randomized trial 40954. J Clin Oncol. 28:5210–5218. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wagner AD, Grothe W, Haerting J, Kleber G,
Grothey A and Fleig WE: Chemotherapy in advanced gastric cancer: A
systematic review and meta-analysis based on aggregate data. J Clin
Oncol. 24:2903–2909. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hallett RM, Huang C, Motazedian A, Auf der
Mauer S, Pond GR, Hassell JA, Nordon RE and Draper JS:
Treatment-induced cell cycle kinetics dictate tumor response to
chemotherapy. Oncotarget. 6:7040–7052. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Malumbres M: Cyclin-dependent kinases.
Genome Biol. 15:1222014. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yadav V, Sultana S, Yadav J and Saini N:
Gatifloxacin induces S and G2-phase cell cycle arrest in pancreatic
cancer cells via p21/p27/p53. PLoS One. 7:e477962012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin HP, Lin CY, Huo C, Hsiao PH, Su LC,
Jiang SS, Chan TM, Chang CH, Chen LT, Kung HJ, et al: Caffeic acid
phenethyl ester induced cell cycle arrest and growth inhibition in
androgen-independent prostate cancer cells via regulation of Skp2,
p53, p21Cip1 and p27Kip1. Oncotarget. 6:6684–6707. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Datto MB, Li Y, Panus JF, Howe DJ, Xiong Y
and Wang XF: Transforming growth factor beta induces the
cyclin-dependent kinase inhibitor p21 through a p53-independent
mechanism. Proc Natl Acad Sci USA. 92:5545–5549. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Asghar U, Witkiewicz AK, Turner NC and
Knudsen ES: The history and future of targeting cyclin-dependent
kinases in cancer therapy. Nat Rev Drug Discov. 14:130–146. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Park MT and Lee SJ: Cell Cycle and Cancer.
J Biochem Mol Biol. 36:60–65. 2003.PubMed/NCBI
|
|
35
|
Nag S, Zhang X, Srivenugopal KS, Wang MH,
Wang W and Zhang R: Targeting MDM2-p53 interaction for cancer
therapy: Are we there Yet? Curr Med Chem. 21:553–574. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Adams JM and Cory S: The Bcl-2 apoptotic
switch in cancer development and therapy. Oncogene. 26:1324–1337.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zimmermann KC, Bonzon C and Green DR: The
machinery of programmed cell death. Pharmacol Ther. 92:57–70. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jo EH, Kim SH, Ra JC, Kim SR, Cho SD, Jung
JW, Yang SR, Park JS, Hwang JW, Aruoma OI, et al: Chemopreventive
properties of the ethanol extract of chinese licorice
(Glycyrrhizauralensis) root: Induction of apoptosis and G1 cell
cycle arrest in MCF-7 human breast cancer cells. Cancer Lett.
230:239–247. 2005. View Article : Google Scholar : PubMed/NCBI
|