Introduction
Our previous studies demonstrated that one of the
main mechanisms of simvastatin (SIM) cytotoxic effects on B16.F10
murine melanoma cells under normoxic conditions is associated with
the suppression of the constitutive tumor cell production of the α
subunit of hypoxia inducible factor (HIF)-1 (1). The normoxic expression of HIF-1 via the
constitutive activation of the Ras-Raf-mitogen-activated protein
kinase kinase-extracellular signal-regulated kinase 1/2 and
phosphatidylinositol-4,5-bisphosphate 3-kinase-AKT signaling
pathways ensures the maintenance of the malignant phenotype of
highly metastatic melanoma cells (2–4). In
addition, the production of HIF-1 in these malignant cells is
further amplified by the hypoxic tumor microenvironment, suggesting
that, under hypoxia, HIF-1 may activate different signaling
pathways to promote cell survival and proliferation (5). Thus, hypoxia-induced HIF-1α
stabilization is followed by the activation of multiple genes
encoding for proteins with critical functions in angiogenesis, cell
proliferation, invasion, metastasis, genetic instability, cell
survival, apoptosis, epithelial-mesenchymal transition and immune
evasion (6–8).
The present study aimed to explore whether the
mechanisms underlying the cytotoxicity of SIM on B16.F10 melanoma
cells cultured under hypoxia also involved the modulation of HIF-1α
expression in these cancer cells. To mimic hypoxia, B16.F10
melanoma cells were cultured in the presence of cobalt chloride
(CoCl2), a chemical inducer of HIF-1α stabilization
(9,10). The mechanisms of cytotoxicity exerted
by SIM on B16.F10 cells grown in the presence of the
CoCl2 were investigated with regard to tumor cell
production of HIF-1α and expression of proteins involved in main
tumorigenic processes coordinated by this protein, including
angiogenesis and inflammation (11,12).
Furthermore, as previous experimental data suggested that nuclear
factor (NF)-κB can also induce overexpression of HIF-1α messenger
RNA (13–15), the expression level of this protein
was analyzed. In addition, the role of oxidative stress in
SIM-induced cytotoxicity was evaluated. The results demonstrated
that, under hypoxia, the main mechanism of SIM cytotoxicity in
B16.F10 melanoma cells is mediated via inhibition of the chemically
induced expression of HIF-1α in these cancer cells. Additionally,
and in accordance with the main function of the inducible
production of this factor, the levels of proteins supporting tumor
angiogenesis and inflammation were also strongly suppressed.
Materials and methods
Cell line and culture conditions
B16.F10 murine melanoma cells (CRL-6475; American
Type Culture Collection, Manassas, VA, USA) were cultured in
Dulbecco's modified Eagle's medium (Lonza Group Ltd., Basel,
Switzerland) supplemented with 10% heat-inactivated fetal bovine
serum (HyClone; GE Healthcare Life Sciences, Logan, UT, USA), 100
IU/ml penicillin, 100 µg/ml streptomycin, 0.25 µg/ml amphotericin
B, 4 mM L-glutamine and 5% NaHCO3 (Lonza Group Ltd.) as
monolayer at 37°C in a 5% CO2 humidified atmosphere. To
obtain hypoxic conditions, cells were incubated in the above
culture medium supplemented with several concentrations of
CoCl2 (100, 200 and 300 µM). It was noticed that 200 µM
CoCl2 was the minimum concentration that allowed the
stimulation of the inducible expression of HIF-1α without causing
any cytotoxic effects on these cancer cells (data not shown).
Stock solutions of SIM
SIM (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
was dissolved in 70% ethanol to prepare stock solutions of 2 mg/ml.
Working solutions were prepared directly into the culture medium.
As controls for ethanol toxicity, cells were incubated with ethanol
at similar concentrations (0.017–0.925%) as those used for the
preparation of SIM working solutions.
Cell proliferation assay
To determine the effects of SIM on cell
proliferation under normoxic and hypoxic conditions, B16.F10
melanoma cells (1,000 cells/well) were seeded in a 96-well plate
for 24 h. Different concentrations of SIM, ranging from 0.5 to 50
µg/ml, were evaluated in triplicate. The range of statin
concentrations was selected based on our previous study regarding
the statin effects on B16.F10 cells (1). The cytotoxicity of the ethanol
concentrations used for the preparation of the different SIM
solutions was also assessed. Cells cultured either in medium or in
medium supplemented with 200 µM CoCl2 were used as
controls for normoxic and hypoxic conditions, respectively. The
proliferative activity of the cells following statin administration
was analyzed with an immunoassay [Cell Proliferation ELISA, BrdU
(colorimetric); Roche Applied Science, Penzberg, Germany] according
to the manufacturer's protocol (16,17). This
method is based on the incorporation of the pyridine analogue
bromodeoxyuridine (BrdU), instead of thymidine, into the DNA of
proliferating cells (16). B16.F10
melanoma cells were incubated with a BrdU solution for 24 h, and
the culture medium was then completely removed from each well.
Following this step, the cells were fixed, and the DNA was
denatured with FixDenat buffer provided in the kit. An anti-BrdU
monoclonal antibody conjugated with peroxidase (anti-BrdU-POD,
#11647229001, Roche Applied Science, dilution, 1:100; part of Cell
Proliferation ELISA, BrdU kit) was added in each well in order to
detect the incorporated BrdU in the newly synthesized cellular DNA.
The antibody was removed after 1 h of incubation at room
temperature, and the cells were then washed three times with PBS.
Next, a peroxidase substrate (3,3′,5,5′-tetramethylbenzidine, part
of Cell Proliferation ELISA, BrdU kit) was added in each well, and
the immune complexes were detected by measuring the absorbance of
the reaction product at 450 nm with a reference wavelength of 655
nm.
Viability assay
The cytotoxic effect of SIM was assessed using a
colorimetric assay (CytoTox 96 Non-Radioactive Cytotoxicity Assay;
Promega Corporation, Madison, WI, USA) based on the measurement of
the catalytic activity of the lactate dehydrogenase (LDH) enzyme
released in the culture medium (18).
The assay was performed following the manufacturer's protocol.
Briefly, B16.F10 melanoma cells (5,000 cells/well) were cultured in
a 96-well plate for 24 h at 37°C. Each statin concentration,
ranging from 0.5 to 50 mg/ml, was added in triplicate. The
cytotoxicity of the ethanol concentrations used for the preparation
of the different SIM solutions was also tested. Cells cultured
either in medium or in medium with 200 µM CoCl2 were
used as controls for the spontaneous release of LDH under normoxic
or hypoxic conditions, respectively. To determine the total release
of LDH in the culture medium, 10 µl of cell lysis buffer (part of
the CytoTox 96 Non-Radioactive Cytotoxicity Assay kit) was added to
the wells containing control cells for 45 min prior to harvesting
the supernatant. At the end of the incubation period, the plate was
centrifuged at 260 × g, and 50 µl of the culture medium from each
well was transferred to an enzymatic assay plate and mixed with an
equal volume of substrate mix (lactate and iodonitrotetrazolium
violet, included in the CytoTox 96 Non-Radioactive Cytotoxicity
Assay kit). After 30 min of incubation at room temperature, the
reaction was stopped by adding the stop solution from the CytoTox
96 Non-Radioactive Cytotoxicity Assay kit, and the absorbance was
measured at 490 nm.
Western blot analysis of the
expression of HIF-1α and NF-κB
The effects of 5 µg/ml SIM on the B16.F10 cell
production of two key players in tumor cell survival, HIF-1 and
NF-κB, was assessed by western blot analysis after 24 h of
incubation at 37°C. To obtain cell lysates (19–21), cells
were washed with PBS, and viable (adherent) tumor cells were
mechanically detached and lysed with cell lysis buffer [10 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, 200 mM NaCl, 1%
Triton X-100, 10 mM MgCl2 and 1 mM dithiothreitol] after
30 min of incubation on ice. cOmplete™ Protease Inhibitor Cocktail
Tablets (Roche Applied Science) were added to the lysis buffer.
Cells lysates were cleared by centrifugation at 18,000 × g for 10
min at 4°C, and the supernatant was collected. The protein content
of the cell lysates was determined by Bradford assay
(Sigma-Aldrich; Merck KGaA). A total of 50 µg of total protein was
loaded per lane onto a 10% polyacrylamide gel. Electrophoresis was
performed at 45 mV, and subsequently the proteins were
electro-transferred onto a nitrocellulose membrane at 100 mV for 40
min. The membranes were blocked with 5% skimmed milk powder
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) in TBS containing
0.1% Tween-20 (TBS-T) for 3 h at room temperature. Subsequently,
the membranes were incubated overnight at 4°C with rabbit
polyclonal anti-HIF-1α (NB100-479, Novus Biologicals, Ltd.,
Cambridge, UK) and mouse monoclonal anti-NF-κB antibodies (sc56735;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA), diluted 500-fold
in 5% skimmed milk powder in TBS-T. For the loading control, a
primary rabbit polyclonal antibody against mouse β-actin (A2103;
Sigma-Aldrich; Merck KGaA), diluted 1,000-fold in TBS-T, was used.
To detect the bound antibodies, the membranes were washed with
TBS-T and incubated with a goat anti-rabbit (sc-2004; Santa Cruz
Biotechnology, Inc.) or a goat anti-mouse horseradish peroxidase
(HRP)-conjugated secondary antibody (sc-2005; Santa Cruz
Biotechnology, Inc.), diluted 4,000-fold in 5% skimmed milk powder
in TBS-T at room temperature for 2 h. Proteins were detected using
SuperSignal West Pico Chemiluminescent Substrate (Thermo Fisher
Scientific, Inc., Waltham, MA, USA), and the membranes were exposed
to an X-ray film (Kodak, Rochester, NY, USA) for 2 min. The films
were developed and analyzed using TotalLab Quant Software version
12 for Windows (TotalLab, Ltd., Newcastle upon Tyne, UK).
Transcription factors levels in tumor cells treated with statin
were compared to those in the controls. The final results are
represented as the mean ± standard deviation (SD) of two
independent experiments.
Array analysis of
inflammatory/angiogenic protein levels
To assess whether SIM could affect the tumor cell
production of proteins involved in angiogenesis and inflammation, a
screening using a protein array (AAM-ANG-1-8; RayBio®
Mouse Angiogenesis Array C1; RayBiotech Inc., Norcross, GA, USA)
was performed as described previously (22). The same tumor cell lysates as those
employed in western blot analysis were used. One array membrane
containing 24 types of primary antibodies against specific mouse
proteins was used per cell lysate. The array membranes were
incubated with 200 µg of cell lysate proteins overnight at 4°C. A
mixture of secondary biotin-conjugated antibodies (included in the
RayBio® Mouse Angiogenesis Array C1 kit) against the
same angiogenic factors as those targeted by the aforementioned
primary antibodies was added to the membranes and incubated for 2 h
at room temperature, followed by incubation with HRP-conjugated
streptavidin (included in the RayBio® Mouse Angiogenesis
Array C1 kit) for additional 2 h at room temperature. Each
incubation step was followed by five washing steps with two washing
buffers included in the kit. Thereafter, the membranes were
incubated with a mixture of two detection buffers (included in the
RayBio® Mouse Angiogenesis Array C1 kit) for 1 min,
exposed to an X-ray film (Kodak) for 1 min, and then the films were
developed. The protein expression level was quantified by measuring
the intensity of the color of each spot on the membranes in
comparison with the positive control spots already bound to the
membranes using TotalLab Quant Software version 12 for Windows.
Each protein level from the statin-treated groups was expressed as
a percentage of the same protein level from the untreated cells
(controls). Each protein from each experimental group was
determined in duplicate.
High-performance liquid chromatography
(HPLC) determination of malondialdehyde (MDA) levels
MDA is the main product of lipid peroxidation
mediated by reactive oxygen species (ROS), and therefore, it is a
good indicator of overall oxidative stress (23). In the present study, MDA levels were
determined according to the HPLC method employed by Karatas et
al (24). Prior to HPLC
quantification of MDA, sample deproteinization was performed as
described previously (24). Then,
samples were centrifuged at 4,500 × g for 5 min, and 100 µl of each
supernatant was used for HPLC analysis. The column type was
LiChrosorb® RP18 (5 µm) (Sigma-Aldrich; Merck KGaA), and
the mobile phase consisted of 30 mM
KH2PO4:methanol in a volume ratio of 65:35.
The flow rate was set at 0.5 ml/min, and MDA was measured using an
ultraviolet detector set at 254 nm. The retention time of MDA was
~5.4 min. Data were expressed as µmol of MDA/mg of protein.
Statistical analysis
Data from different experiments were reported as the
mean ± SD. SIM effects on tumor cells cultured with or without
CoCl2 were analyzed by unpaired Student's t-test. To
analyze the SIM effects on the levels of angiogenic/inflammatory
proteins in cancer cells under both culture conditions, two-way
analysis of variance with Bonferroni correction for multiple
comparisons was used. All statistical analyses were performed with
GraphPad Prism version 6 for Windows (GraphPad Software, Inc., La
Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
SIM cytotoxicity in vitro
The cytotoxic effects of SIM were assessed with
regard to B16.F10 cell proliferation and viability, and the results
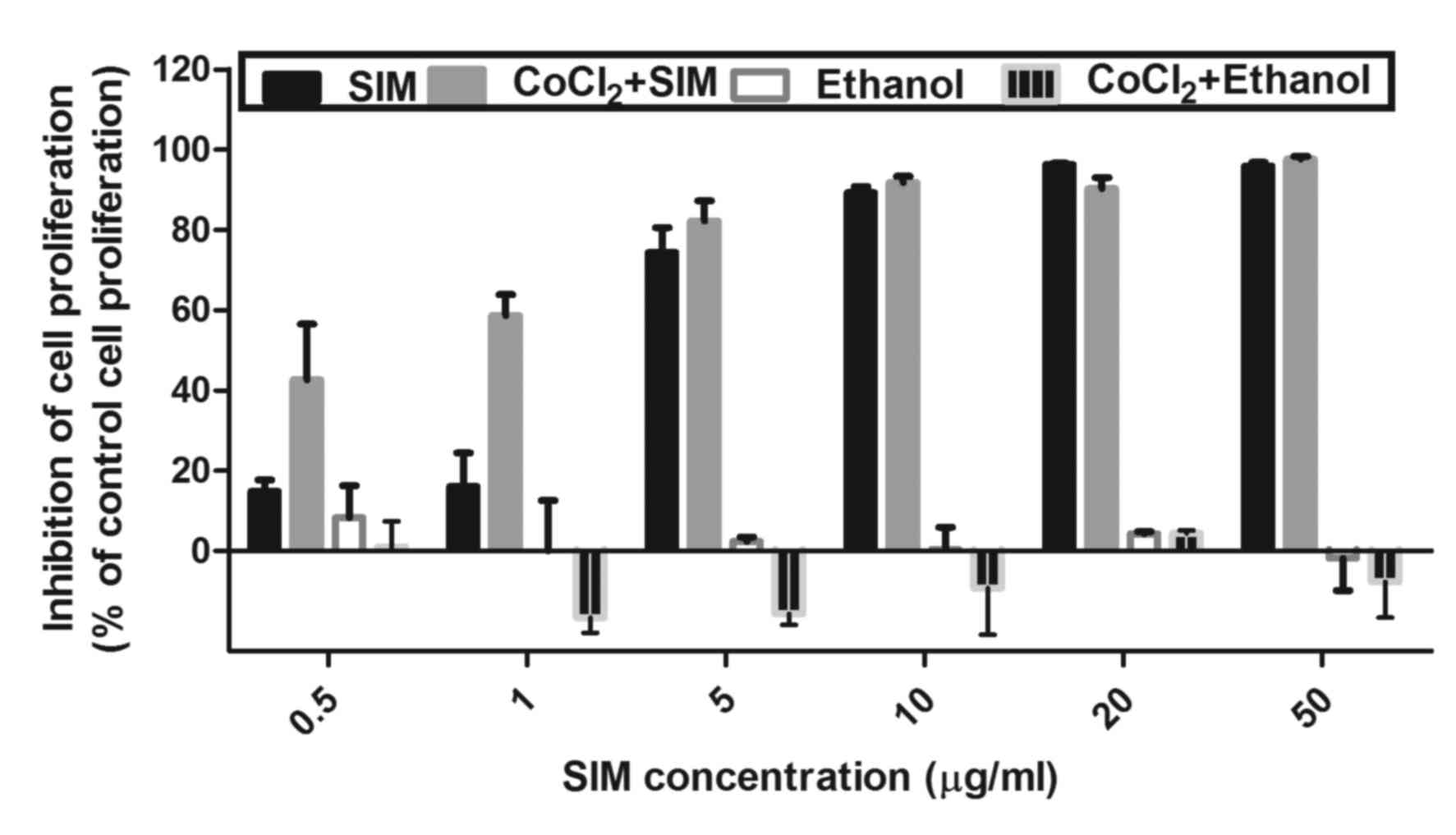
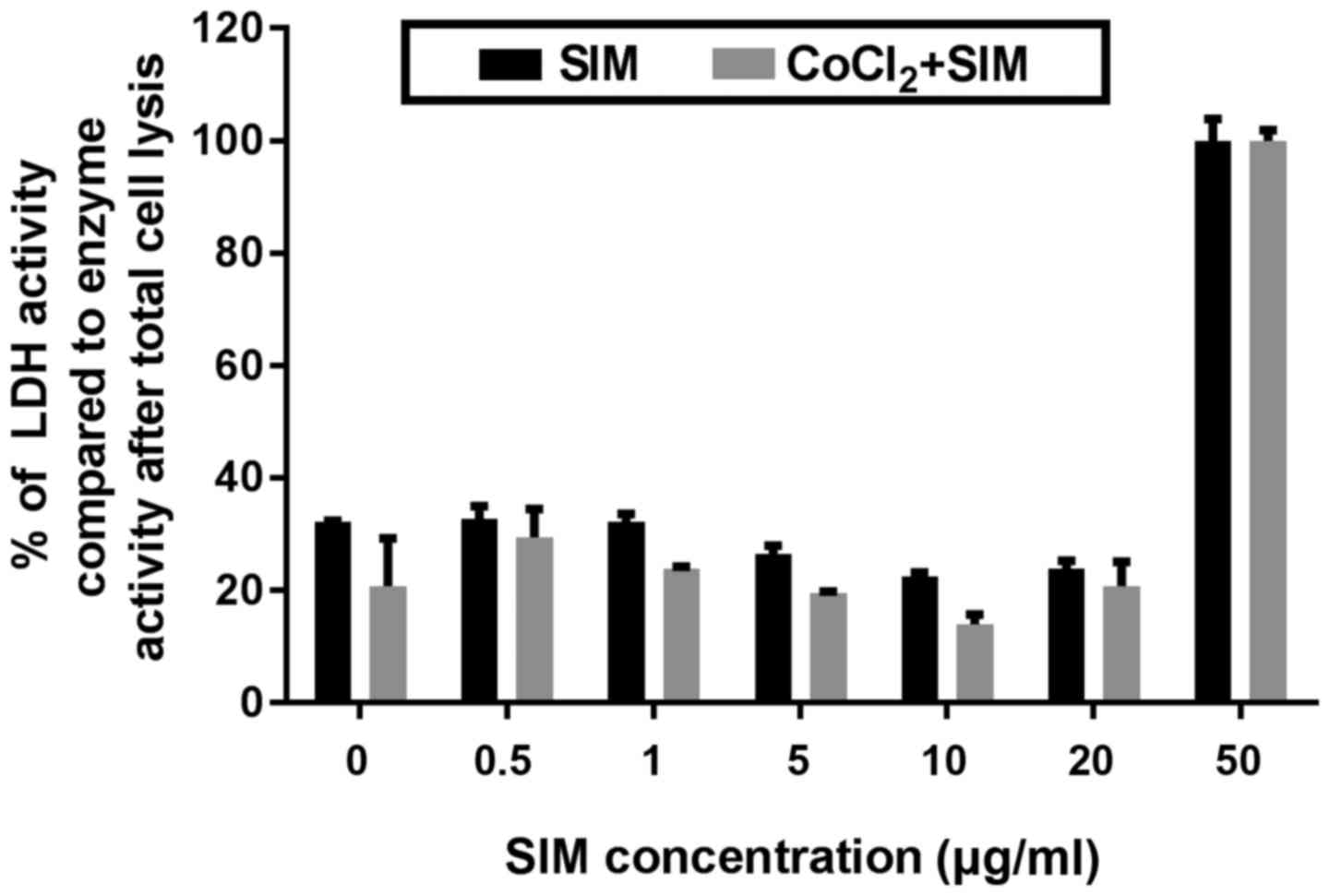
are shown in Figs. 1 and 2.
Effect of SIM on cell
proliferation
To investigate the inhibitory effect of SIM on the
proliferation of B16.F10 murine melanoma cells, these cells were
incubated in the presence of CoCl2 with increasing
concentrations of SIM ranging from 0.5 to 50 µg/ml for 24 h
(Fig. 1). The proliferation rate of
the cells incubated with the same concentrations of SIM under
normoxic conditions was also assessed. Cells incubated only with
culture medium or with culture medium supplemented with 200 µM
CoCl2 were used as controls. The cytotoxicity of the
ethanol concentrations used for SIM solutions preparation was also
tested. No inhibitory effect of any ethanol concentration on tumor
cell proliferation was detected (Fig.
1).
The effect of SIM on cancer cell proliferation was
expressed as the percentage of inhibition of cell proliferation
compared with the proliferation of cells used as controls (Fig. 1). After 24 h of incubation, SIM
inhibited cell proliferation by >70% under normoxic and hypoxic
conditions at concentrations ranging from 5 to 50 µg/ml. Notably,
under hypoxia, a moderate inhibitory effect of the statin (~40–60%
compared with the proliferation of control cells) was also noted at
the lowest drug concentrations tested (0.5 and 1 µg/ml). However,
since 5 µg/ml was the lowest concentration of SIM that exhibited a
strong cytotoxic effect (>70% inhibition of cell proliferation
compared with the proliferation of control cells) (Fig. 1), this concentration was used
throughout the experiments performed for testing the mechanisms of
SIM cytotoxicity.
Effect of SIM on cell viability
The cytotoxic effect of different concentrations of
SIM under both normoxic and hypoxic conditions after 24 h of
incubation were expressed as percentages of catalytic activity of
LDH released in the culture medium. As shown in Fig. 2, the viability of tumor cells grown
under normoxia or hypoxia was not affected by SIM at concentrations
ranging from 0.5 to 20 µg/ml. Only at the highest concentration of
SIM tested (50 µg/ml), the viability of B16.F10 melanoma cells was
totally affected (Fig. 2), since the
activity of LDH released from these treated cells was similar to
that determined upon cell lysis.
Effects of SIM on the expression
levels of HIF-1α and NF-κB
Since HIF-1 and NF-κB serve key roles in the
preservation of the malignant phenotype of cancer cells and in the
survival mechanisms of tumor cells under critical conditions
(25–27), the effects of 5 µg/ml SIM on the
expression of both proteins in B16.F10 melanoma cells were
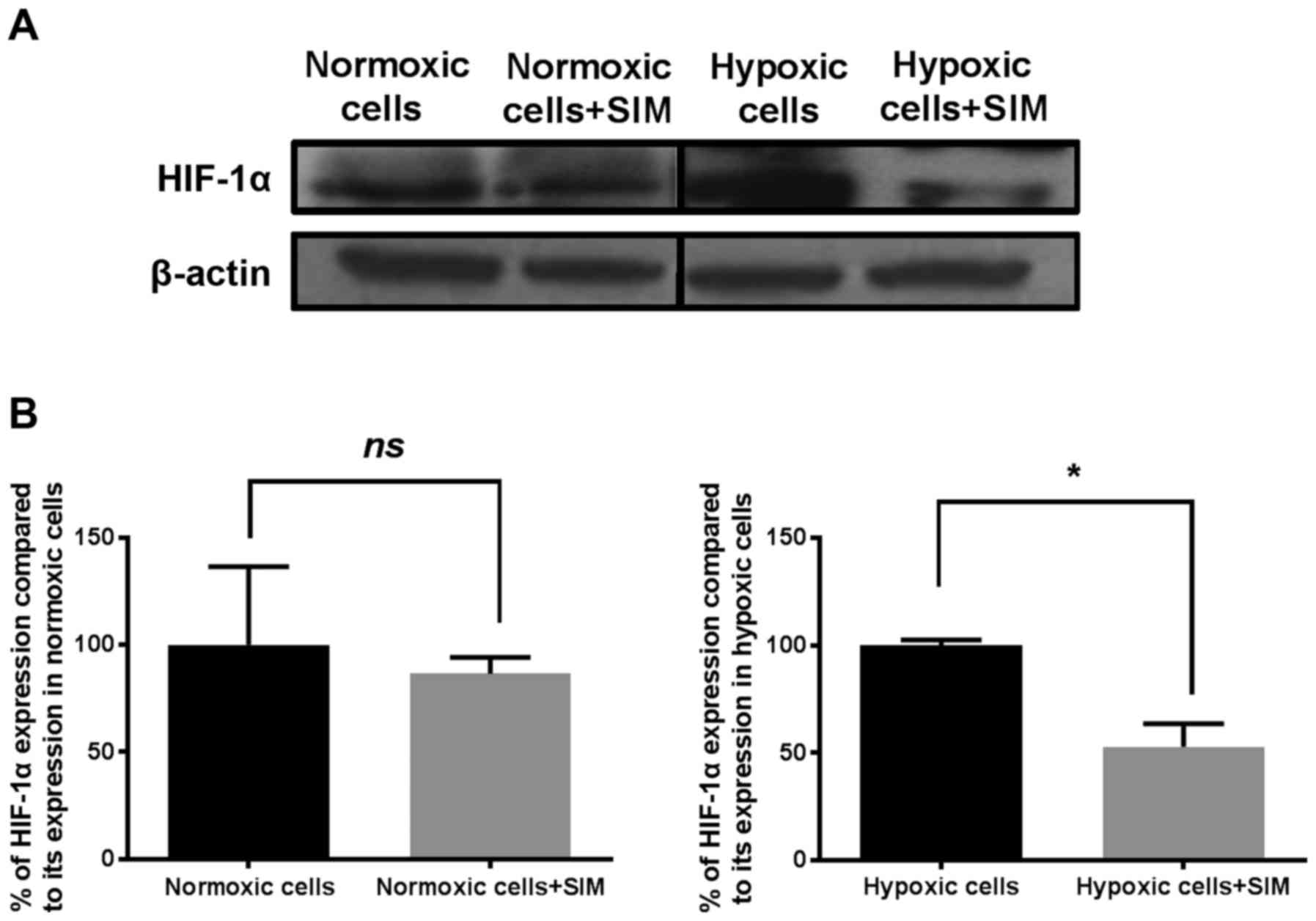
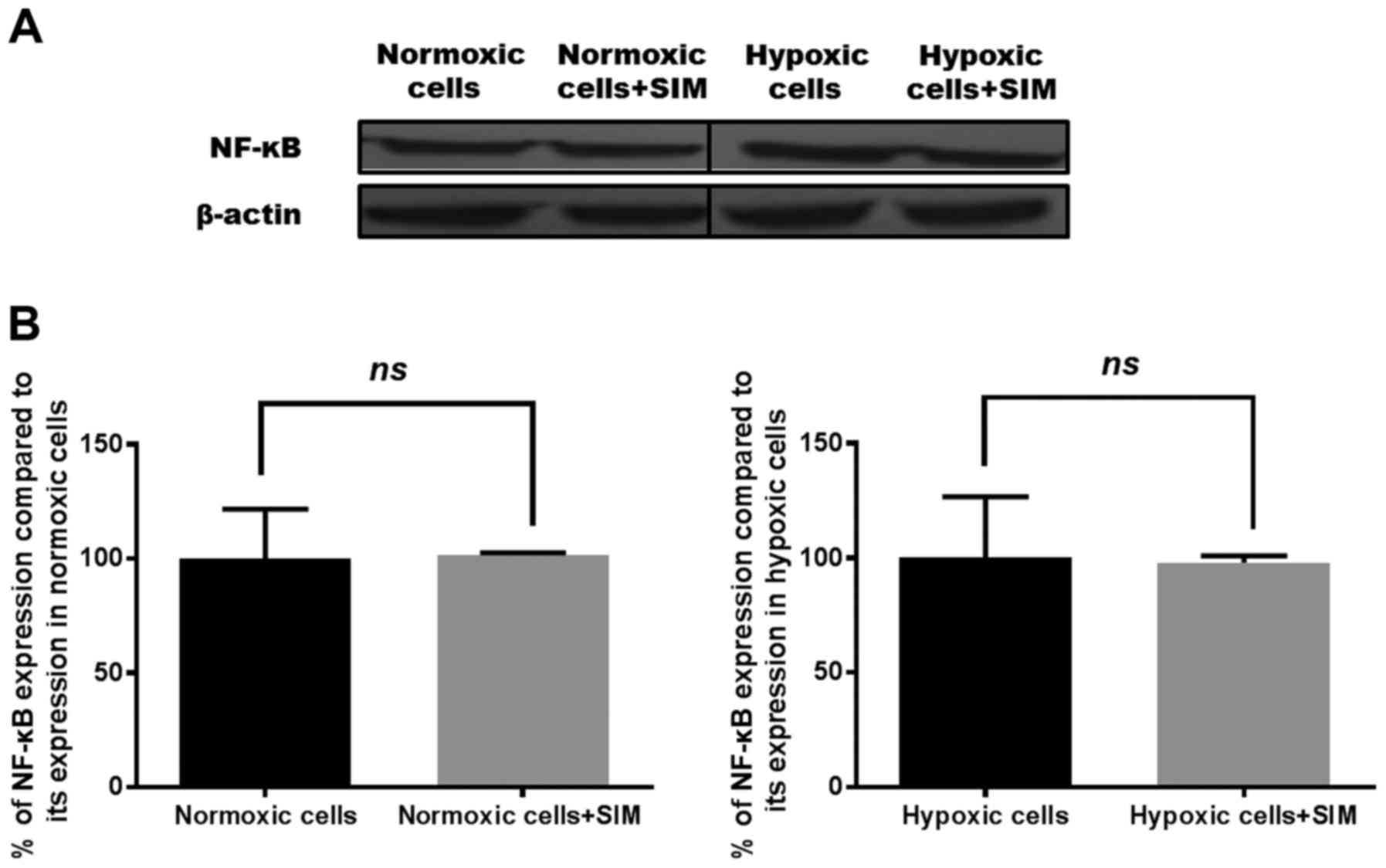
evaluated. The results are shown in Figs.
3 and 4, respectively. As
previously reported (6,10), the present study confirmed that the
addition of 200 µM CoCl2 to melanoma cells increased the
expression level of HIF-1α by 68% compared with its level in the
same cells cultured under normoxia (P<0.05).
Under hypoxic conditions, SIM inhibited HIF-1α
expression by 50% compared with its production in the control
hypoxic cells (P<0.05), while constitutive expression of this
factor was not affected by the statin (Fig. 3B). Our previous results have shown
that SIM inhibited strongly the constitutive expression of HIF-1α
in B16.F10 murine melanoma cells at a concentration 10-fold higher
than that used in the present study (1). Regarding the production of NF-κB, SIM
treatment did not affect the expression levels of this
transcription factor under normoxic or hypoxic conditions (Fig. 4).
Effects of SIM on the
angiogenic/inflammatory capacity of B16.F10 cells
As the production of one of the key regulatory
factors of angiogenesis and inflammation, HIF-1α (1), was inhibited by SIM treatment under
hypoxia, this effect was associated with the levels of proteins
involved in these tumorigenic processes in tumor cells. Therefore,
a screening for 24 angiogenic and inflammatory proteins was
performed via protein arrays (Tables
I and II).
 | Table I.Effects of 5 µg/ml SIM on B16.F10
melanoma cell production of pro-angiogenic/pro-inflammatory
proteins under normoxic and hypoxic conditions. |
Table I.
Effects of 5 µg/ml SIM on B16.F10
melanoma cell production of pro-angiogenic/pro-inflammatory
proteins under normoxic and hypoxic conditions.
|
| Percentage of
reduction (−) or increase (+) in tumor cell production of proteins
involved in tumorangiogenesis/inflammation following SIM treatment,
mean ± SD |
|---|
|
|
|
|---|
|
Pro-angiogenic/pro-inflammatory
proteins | SIM | CoCl2 +
SIM |
|---|
| Granulocyte-colony
stimulating factor |
+19±18.12a |
−83.19±0.54d |
|
Granulocyte-macrophage-colony stimulating
factor |
+22.41±13.20a |
−74.00±1.09d |
| Macrophage-colony
stimulating factor |
+9.27±0.78a |
−82.80±0.13d |
| Insulin growth
factor II |
+4.60±2.16a |
−82.37±0.22d |
| Interleukin 1α |
+18.00±0.01b |
−73.90±0.50d |
| Interleukin 1β |
+22.70±0.93c |
−72.64±0.51d |
| Interleukin 6 |
+13.63±5.36a |
−85.52±0.04d |
| Interleukin 9 |
+7.92±1.27a |
−71.40±0.35d |
| Interleukin 12
p40 |
+80.89±11.48d |
−83.26±0.73d |
| Interleukin 13 |
+6.19±2.86a |
−75.42±0.78d |
| Tumor necrosis
factor α |
+58.87±29.45c |
−83.98±0.17d |
| Monocyte
chemoattractant protein-1 |
+24.07±3.21c |
−82.65±0.14d |
| Eotaxin |
−4.09±3.11a |
−47.31±0.86b |
| Fas ligand |
+5.91±17.93a |
−74.8±0.19d |
| Basic fibroblast
growth factor |
+22.68±7.57a |
−85.19±0.06d |
| Vascular
endothelial growth factor |
+0.92±23.31a |
−72.22±0.66b |
| Leptin |
+8.64±13.19a |
−56.23±1.39a |
| Thrombopoietin |
−7.03±11.62a |
−80.83±0.32d |
| Table II.Effects of 5 µg/ml SIM on B16.F10
melanoma cell production of anti-angiogenic/anti-inflammatory
proteins under normoxic and hypoxic conditions. |
Table II.
Effects of 5 µg/ml SIM on B16.F10
melanoma cell production of anti-angiogenic/anti-inflammatory
proteins under normoxic and hypoxic conditions.
|
| Percentage of
reduction (−) and increase (+) in tumor cell production of proteins
involved in tumor angiogenesis/inflammation following SIM
treatment, mean ± SD |
|---|
|
|
|
|---|
|
Anti-angiogenic/anti-inflammatory
proteins | SIM | CoCl2 +
SIM |
|---|
| Tissue inhibitor of
matrix metalloproteinase 1 |
−6.75±9.18a |
−78.13±0.47c |
| Tissue inhibitor of
matrix metalloproteinase 2 |
−22.72±6.18b |
−82.97±0.10c |
| Platelet factor
4 |
+14.25±8.11a |
−82.13±0.17c |
| Interleukin 12
p70 |
+9.87±2.60a |
−82.32±0.26c |
| Interferon γ |
+17.15±3.92a |
−82.32±0.41c |
| Monokine induced by
interferon γ |
+10.19±1.04a |
−25.09±1.41a |
Notably, the mean average production of angiogenic
and inflammatory proteins tested in cells cultured under
CoCl2-induced hypoxia was significantly higher (by 26%;
P<0.05) than their production in normoxic cells (cells in
culture medium) (Fig. 5). Thus, the
production of 11 out of 24 proteins was stimulated significantly
under CoCl2 treatment. More specifically, the levels of
pro-angiogenic proteins, including granulocyte-colony stimulating
factor (G-CSF), macrophage-CSF (M-CSF), insulin-like growth factor
(IGF)-II, monocyte chemoattractant protein (MCP)-1 and basic
fibroblast growth factor (bFGF), were strongly stimulated in
CoCl2-treated cells (by 50–100%) compared with their
production in control normoxic cells (Fig. 5). The levels of the anti-angiogenic
and anti-inflammatory proteins platelet factor (PF)-4, tissue
inhibitor of metalloproteinase (TIMP)-2 and interleukin (IL)-12p70
were also increased under CoCl2 treatment by 65–95%
compared with their cell production in normoxic cells (Fig. 5). However, the cell production of
important pro-angiogenic [vascular endothelial growth factor
(VEGF)] and inflammatory (IL-1β and IL-6) proteins was reduced by
60% in hypoxic cells in comparison with their normoxic production
(Fig. 5). When tumor cells were
incubated with SIM under normoxic conditions, the overall
production of pro-angiogenic/pro-inflammatory proteins was not
affected by the statin treatment (P>0.05). Furthermore,
incubation of the tumor cells only with SIM stimulated the
production of IL-1α, IL-1β and MCP-1 by 15–30%, while the
expression of IL-12p40 and tumor necrosis factor (TNF)-α was
stimulated by 60–80% (Table I). Only
the level of the anti-angiogenic protein TIMP-2 was significantly
reduced by 22% compared with its level in normoxic cells (Table II).
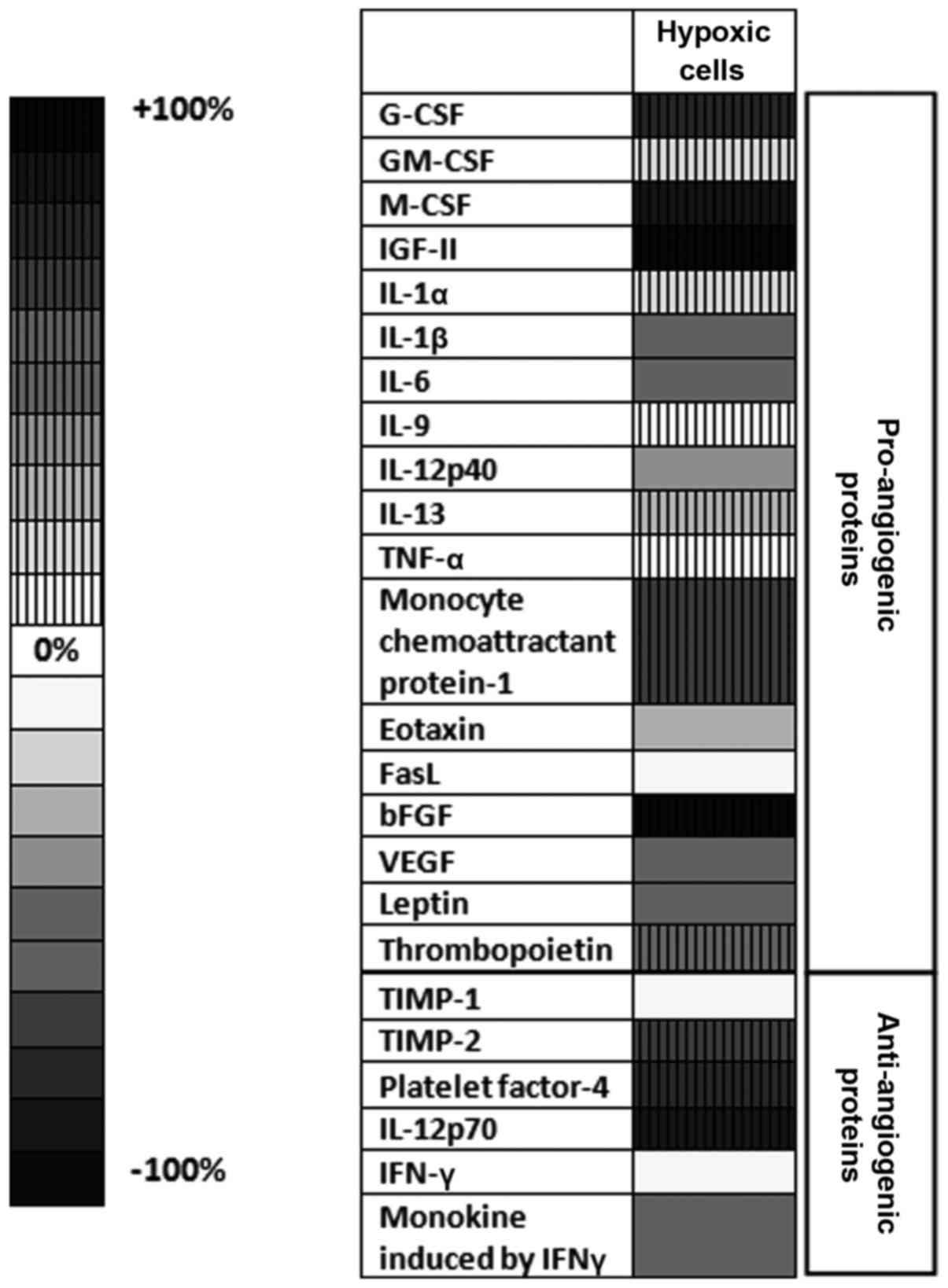 | Figure 5.Effects of CoCl2-induced
hypoxia on the production of angiogenic/inflammatory proteins
in B16.F10 melanoma cells. Results are presented either as a
percentage of reduction in tumor protein levels ranging from 0%
(white) to 100% (black), or as percentage of increase in protein
production ranging from 0% (white with vertical line pattern) to
100% (black with vertical line pattern) in cells treated with
CoCl2 compared with the levels of the same proteins in
normoxic cells. Hypoxic cells, percentages of protein expression
levels in B16.F10 melanoma cells after incubation with 200 µM
CoCl2 for 24 h compared with their levels in normoxic
cells; G-CSF, granulocyte-colony stimulating factor; M-CSF,
macrophage-colony stimulating factor; IGF, insulin-like growth
factor; IL, interleukin; TNF, tumor necrosis factor; FasL, Fas
ligand; bFGF, basic fibroblast growth factor; VEGF, vascular
endothelial growth factor; TIMP, tissue inhibitor of
metalloproteinase; IFN, interferon. |
When SIM was administered to B16.F10 cells under
hypoxia induced by CoCl2, the levels of all
pro-angiogenic/pro-inflammatory proteins were reduced significantly
(Table I), with an overall inhibitory
effect of 75% (P<0.0001) compared with the production of these
proteins in hypoxic cells. More specifically, SIM reduced strongly
(by 70–85%) the production of 16 out of 18
pro-angiogenic/pro-inflammatory proteins (including G-CSF, GM-CSF,
M-CSF, IGF-II, IL-1α, IL-1β, IL-6, IL-9, IL-12p40, IL-13, TNF-α,
MCP-1, Fas ligand, VEGF, bFGF and thrombopoietin) and moderately
(by 45–60%) that of leptin and eotaxin (Table I). However, with the exception of
monokine induced by gamma interferon (MIG), the cell production of
all the anti-angiogenic proteins was also inhibited strongly (by
78–82%) by SIM treatment following CoCl2 administration
(Table II).
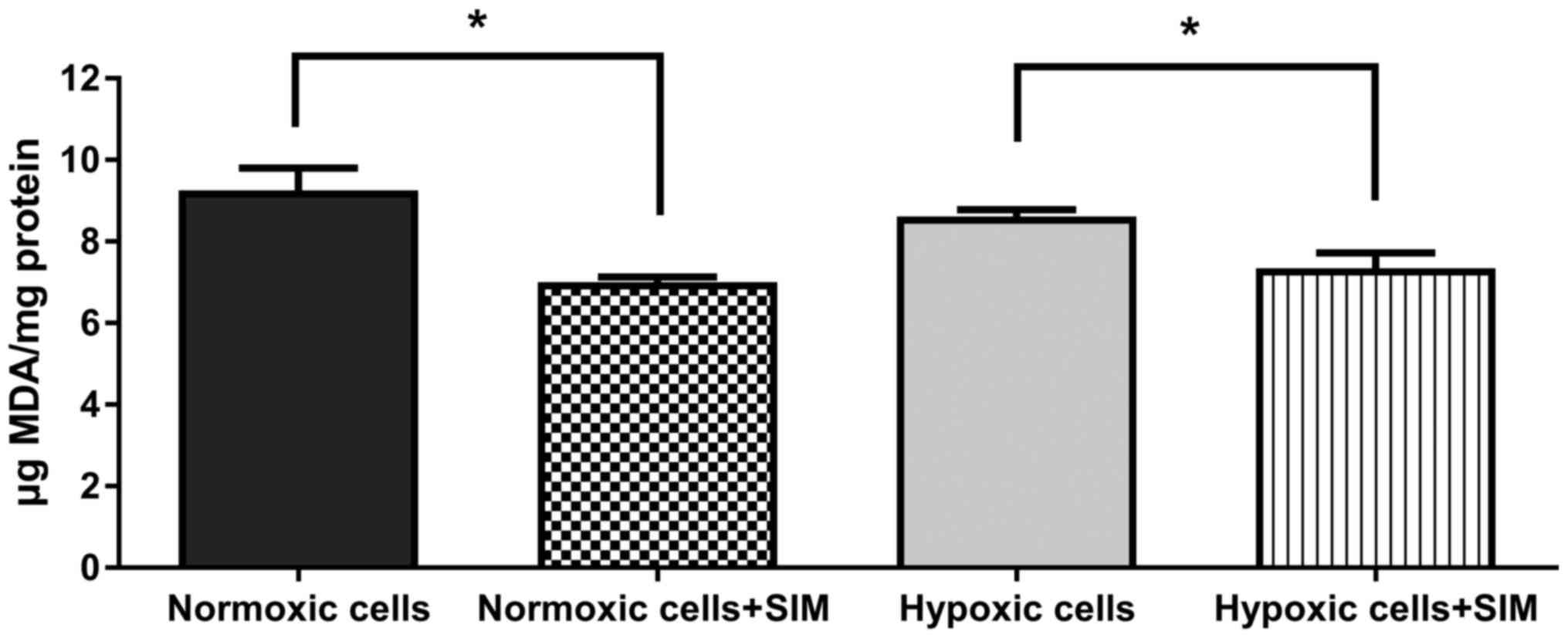
SIM effect on intracellular oxidative
stress
To evaluate whether SIM treatment affected
intracellular oxidative stress in murine melanoma cells cultured
under normoxic and hypoxic conditions, the levels of MDA (a general
marker for oxidative stress in tumor cells) (24) were determined. The data revealed that
5 µg/ml SIM significantly inhibited (P<0.05) the production of
MDA in B16.F10 melanoma cells by 15–25% under both conditions
(Fig. 6). Therefore, the anti-oxidant
activity of SIM may complement the antitumor effects of this statin
on the expression levels of HIF-1α, and angiogenic and inflammatory
proteins in B16.F10 melanoma cells.
Discussion
The present study provides a follow-up of our
earlier observations that inhibition of the constitutive expression
of HIF-1α in B16.F10 murine melanoma cells is one of the main
reasons behind the cytotoxicity caused by lipophilic statins on
these cancer cells (1). Therefore,
the present study evaluated whether this finding could be expanded
under hypoxic conditions, when the constitutive production of
HIF-1α in B16.F10 melanoma cells is accompanied by its induced
expression as a result of CoCl2-mediated transcription
factor stabilization and activation (28). To the best of our knowledge, these
statin effects on melanoma cells under hypoxic conditions have not
been previously described. As expected, culture of melanoma cells
in the presence of CoCl2 increased the expression level
of HIF-1α by 68% compared with its expression in normoxic cells
(Fig. 3A). It is known that
hypoxia-induced expression of this protein in cancer cells triggers
the upregulation of numerous genes encoding for proteins involved
in tumorigenic processes, including angiogenesis, invasion,
metastasis, cell proliferation, genetic instability, cell survival,
apoptosis, epithelial-mesenchymal transition and immune evasion
(29). Thus, this considerable number
of tumorigenic processes controlled by HIF-1 may be affected
simultaneously by the targeting of SIM to suppress this key
regulatory protein expression in melanoma cells. In line with
previous results regarding SIM cytotoxicity in B16.F10 melanoma
cells under normoxia (1,30), SIM also inhibited strongly the
proliferation of hypoxic melanoma cells at concentrations ranging
from 5 to 50 µg/ml (Fig. 1). However,
the viability of melanoma cells was affected only at the highest
SIM concentration tested (50 µg/ml) (Fig.
2), suggesting a limited cytotoxicity of SIM on these cancer
cells. To further investigate the link between the mechanisms of
SIM-induced cytotoxicity in B16.F10 melanoma cells under hypoxia
and targeting of HIF-1α, the cell expression levels of this protein
were assessed. After 24 h of incubation with 5 µg/ml SIM, the cell
production of HIF-1α was reduced by 50% in hypoxic cells (Fig. 3B), while the same concentration of SIM
added to normoxic cells had no effect on the B16.F10 cell
expression of this transcription factor subunit (Fig. 3A and B). This finding suggested that 5
µg/ml SIM was not sufficient to abolish the constitutive expression
of HIF-1α in B16.F10 melanoma cells, as its production in normoxic
cells was not affected by this treatment (Fig. 3A and B). An explanation for this
result may be due to the lack of effects of SIM on the expression
of NF-κB (Fig. 4). The persistent
NF-κB expression in SIM-treated tumor cells may represent an escape
mechanism for maintaining the basal levels of HIF-1α in highly
metastatic cancer cells, since NF-κB is a direct modulator of
HIF-1α expression at both transcriptional and post-translational
levels (14,31). However, since previous studies
demonstrated that the hypoxic switch from the proliferative
phenotype of melanoma cells to an invasive pattern is dependent on
HIF-1α tumor cell production (9), the
SIM-mediated suppression of HIF-1α production may suggest that this
statin is able to control the metastatic capacity of B16.F10
cells.
To link the inhibitory actions of SIM on HIF-1α
production with the statin actions on the capacity of tumor cells
to support vital processes for tumor survival (12,32), 24
proteins involved in angiogenesis and inflammation were assessed in
the present study (Fig. 5 and
Tables I and II). As expected, overexpression of HIF-1α
following CoCl2 treatment stimulated the levels of the
majority of pro-angiogenic and pro-inflammatory proteins, with the
highest enhancement observed in the production of G-CSF, M-CSF,
IGF-II, MCP-1 and bFGF (Fig. 5).
Among these proteins, bFGF (whose whole levels upon
CoCl2 treatment increased by 100% in comparison with its
control levels) is considered one of the most powerful angiogenic
factors in cancer cells regulated by HIF-1α (33–35). By
contrast, the expression levels of the key pro-angiogenic protein
VEGF and important pro-inflammatory cytokines such as IL-6 and
IL-1β were strongly reduced (by 60%) in B16.F10 cells treated with
CoCl2, suggesting that not all
pro-angiogenic/pro-inflammatory pathways are equally modulated in
these cancer cells (36). In
addition, the expression of MIG, an angiostatic protein (37), was suppressed by 60% upon hypoxic
induction of HIF-1α production. However, probably as a compensatory
mechanism for this effect, the levels of the anti-angiogenic
proteins PF-4, TIMP-2 and IL-12p70 were also increased in cells
treated with CoCl2 (10).
When SIM was administered to normoxic melanoma
cells, the overall production of pro-angiogenic proteins was not
affected (Table I). This finding may
be associated with the inability of SIM to affect the constitutive
expression of HIF-1α in melanoma cells (Fig. 3A). Unexpectedly, SIM administrated
concomitantly with CoCl2 to tumor cells induced a ~75%
reduction in pro-angiogenic factors (Table I). This beneficial effect is clearly
associated with the inhibitory effect of SIM on the hypoxia-induced
cell production of HIF-1α (Fig. 3B),
which is the most important regulator of tumor angiogenesis
(32). However, the cell production
of the majority of anti-angiogenic proteins was strongly inhibited
by SIM treatment in the presence of CoCl2 (Table II). This effect may be associated
with the suppressive effects of SIM exerted on other transcription
factors (such as activator protein-1) that control the expression
of the genes encoding for these proteins (38,39). Since
there is a tight correlation between HIF-1α expression and
oxidative stress intensity (40,41), and a
large body of data demonstrated the role of SIM in the modulation
of tumor oxidative stress (1,42,43), the
current study assessed a general marker for tumor oxidative stress,
MDA. The results indicated that 5 µg/ml SIM slightly reduced the
levels of ROS in melanoma cells under both normoxia and hypoxia
(Fig. 6). Although this beneficial
effect is modest, it may contribute to amplify the main inhibitory
action of SIM on the HIF-1α-activated angiogenic and inflammatory
capacity of melanoma cells.
In summary, the present data suggested that
targeting HIF-1α by SIM in hypoxic melanoma cells induced a strong
antitumor response via inhibition of crucial regulators of
tumorigenic processes such as cell proliferation, angiogenesis and
inflammation. It is worth mentioning that the SIM concentration
used in the present study can be easily achieved in vivo,
thus overcoming one of the main limitations of statin use for
cancer therapy (44). Although SIM
was not able to inhibit the constitutive expression of HIF-1α, the
current data may be valuable for future antitumor strategies based
on the combination of this lipophilic statin with drugs that can
counteract the escape mechanisms used by these tumor cells. Thus,
inhibitors of NF-κB production in melanoma cells such as
NEMO-binding domain peptide selective inhibitor and bortezomib
(45,46) may suppress indirectly the constitutive
levels of HIF-1α in cancer cells and help to improve future
anticancer therapies based on lipophilic statins.
Acknowledgements
The present study was supported by grants from the
Romanian National Authority for Scientific Research and Innovation
(project number PN-II-RU-TE-2014-4-1191; contract number 235/2015)
and the Babes-Bolyai University (project code GTC_34033/2013). The
present manuscript is the result of a post-doctoral research study
conducted thanks to the financial support of the Sectoral
Operational Program for Human Resources Development 2007–2013,
which was co-financed by the European Social Fund under the project
POSDRU/159/1.5/S/133391 (project title ‘Doctoral and postdoctoral
excellence programs for training highly qualified human resources
for research in the fields of Life Sciences, Environment and
Earth’). The open access publication fee for the present study was
covered by the national project PN-II-RU-TE-2014-4-1191 (contract
number 235/2015).
References
|
1
|
Alupei MC, Licarete E, Cristian FB and
Banciu M: Cytotoxicity of lipophilic statins depends on their
combined actions on HIF-1α expression and redox status in B16.F10
melanoma cells. Anticancer Drugs. 25:393–405. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mills CN, Joshi SS and Niles RM:
Expression and function of hypoxia inducible factor-1 alpha in
human melanoma under non-hypoxic conditions. Mol Cancer. 8:1042009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yajima I, Kumasaka MY, Thang ND, Goto Y,
Takeda K, Yamanoshita O, Iida M, Ohgami N, Tamura H, Kawamoto Y and
Kato M: RAS/RAF/MEK/ERK and PI3K/PTEN/AKT signaling in malignant
melanoma progression and therapy. Dermatol Res Pract.
2012:3541912012.PubMed/NCBI
|
|
4
|
Kuphal S, Winklmeier A, Warnecke C and
Bosserhoff AK: Constitutive HIF-1 activity in malignant melanoma.
Eur J Cancer. 46:1159–1169. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Masson N and Ratcliffe PJ: Hypoxia
signaling pathways in cancer metabolism: The importance of
co-selecting interconnected physiological pathways. Cancer Metab.
2:32014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kallio PJ, Wilson WJ, O'Brien S, Makino Y
and Poellinger L: Regulation of the hypoxia-inducible transcription
factor 1alpha by the ubiquitin-proteasome pathway. J Biol Chem.
274:6519–6525. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kaelin WG Jr and Ratcliffe PJ: Oxygen
sensing by metazoans: The central role of the HIF hydroxylase
pathway. Mol Cell. 30:393–402. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Semenza GL: Hypoxia-inducible factor 1 and
cancer pathogenesis. IUBMB Life. 60:591–597. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Widmer DS, Hoek KS, Cheng PF, Eichhoff OM,
Biedermann T, Raaijmakers MI, Hemmi S, Dummer R and Levesque MP:
Hypoxia contributes to melanoma heterogeneity by triggering
HIF1α-dependent phenotype switching. J Invest Dermatol.
133:2436–2443. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rodrigues M, Xin X, Jee K,
Babapoor-Farrokhran S, Kashiwabuchi F, Ma T, Bhutto I, Hassan SJ,
Daoud Y, Baranano D, et al: VEGF secreted by hypoxic Müller cells
induces MMP-2 expression and activity in endothelial cells to
promote retinal neovascularization in proliferative diabetic
retinopathy. Diabetes. 62:3863–3873. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Krock BL, Skuli N and Simon MC:
Hypoxia-induced angiogenesis: Good and evil. Genes Cancer.
2:1117–1133. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ling FC, Khochfar J, Baldus SE, Brabender
J, Drebber U, Bollschweiler E, Hoelscher AH and Schneider PM:
HIF-1alpha protein expression is associated with the environmental
inflammatory reaction in Barrett's metaplasia. Dis Esophagus.
22:694–699. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nakayama K: cAMP-response element-binding
protein (CREB) and NF-κB transcription factors are activated during
prolonged hypoxia and cooperatively regulate the induction of
matrix metalloproteinase MMP1. J Biol Chem. 288:22584–22595. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
van Uden P, Kenneth NS and Rocha S:
Regulation of hypoxia-inducible factor-1alpha by NF-kappaB. Biochem
J. 412:477–484. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Culver C, Sundqvist A, Mudie S, Melvin A,
Xirodimas D and Rocha S: Mechanism of hypoxia-induced NF-kappaB.
Mol Cell Biol. 30:4901–4921. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gratzner HG: Monoclonal antibody to
5-bromo- and 5-iododeoxyuridine: A new reagent for detection of DNA
replication. Science. 218:474–475. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Coimbra M, Banciu M, Fens MH, de Smet L,
Cabaj M, Metselaar JM, Storm G and Schiffelers RM: Liposomal
pravastatin inhibits tumor growth by targeting cancer-related
inflammation. J Control Release. 148:303–310. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Singer CA, Figueroa-Masot XA, Batchelor RH
and Dorsa DM: The mitogen-activated protein kinase pathway mediates
estrogen neuroprotection after glutamate toxicity in primary
cortical neurons. J Neurosci. 19:2455–2463. 1999.PubMed/NCBI
|
|
19
|
Yang L, Xiao M, Li X, Tang Y and Wang YL:
Arginine ADP-ribosyltransferase 1 promotes angiogenesis in
colorectal cancer via the PI3K/Akt pathway. Int J Mol Med.
37:734–742. 2016.PubMed/NCBI
|
|
20
|
Kim J, Shao Y, Kim SY, Kim S, Song HK,
Jeon JH, Suh HW, Chung JW, Yoon SR, Kim YS and Choi I:
Hypoxia-induced IL-18 increases hypoxia-inducible factor-1alpha
expression through a Rac1-dependent NF-kappaB pathway. Mol Biol
Cell. 19:433–444. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shi Y, Chang M, Wang F, Ouyang X, Jia Y
and Du H: Role and mechanism of hypoxia-inducible factor-1 in cell
growth and apoptosis of breast cancer cell line MDA-MB-231. Oncol
Lett. 1:657–662. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Banciu M, Schiffelers RM, Fens MH,
Metselaar JM and Storm G: Anti-angiogenic effects of liposomal
prednisolone phosphate on B16 melanoma in mice. J Control Release.
113:1–8. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Del Rio D, Stewart AJ and Pellegrini N: A
review of recent studies on malondialdehyde as toxic molecule and
biological marker of oxidative stress. Nutr Metab Cardiovasc Dis.
15:316–328. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Karatas F, Karatepe M and Baysar A:
Determination of free malondialdehyde in human serum by
high-performance liquid chromatography. Anal Biochem. 311:76–79.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Culver C, Sundqvist A, Mudie S, Melvin A,
Xirodimas D and Rocha S: Mechanism of hypoxia-induced NF-kappaB.
Mol Cell Biol. 30:4901–4921. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nakayama K: cAMP-response element-binding
protein (CREB) and NF-κB transcription factors are activated during
prolonged hypoxia and cooperatively regulate the induction of
matrix metalloproteinase MMP1. J Biol Chem. 288:22584–22595. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vaupel P, Mayer A and Höckel M: Tumor
hypoxia and malignant progression. Methods Enzymol. 381:335–354.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Semenza GL: HIF-1, O(2) and the 3 PHDs:
How animal cells signal hypoxia to the nucleus. Cell. 107:1–3.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Semenza GL: Defining the role of
hypoxia-inducible factor 1 in cancer biology and therapeutics.
Oncogene. 29:625–634. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Glynn SA, O'Sullivan D, Eustace AJ, Clynes
M and O'Donovan N: The 3-hydroxy-3-methylglutaryl-coenzyme A
reductase inhibitors, simvastatin, lovastatin and mevastatin
inhibit proliferation and invasion of melanoma cells. BMC cancer.
8:92008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rius J, Guma M, Schachtrup C, Akassoglou
K, Zinkernagel AS, Nizet V, Johnson RS, Haddad GG and Karin M:
NF-kappaB links innate immunity to the hypoxic response through
transcriptional regulation of HIF-1alpha. Nature. 453:807–811.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Krock BL, Skuli N and Simon MC:
Hypoxia-induced angiogenesis: Good and evil. Genes Cancer.
2:1117–1133. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kerbel RS: Tumor angiogenesis: Past,
present and the near future. Carcinogenesis. 21:505–515. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lieu C, Heymach J, Overman M, Tran H and
Kopetz S: Beyond VEGF: Inhibition of the fibroblast growth factor
pathway and antiangiogenesis. Clin Cancer Res. 17:6130–6139. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Calvani M, Rapisarda A, Uranchimeg B,
Shoemaker RH and Melillo G: Hypoxic induction of an
HIF-1alpha-dependent bFGF autocrine loop drives angiogenesis in
human endothelial cells. Blood. 107:2705–2712. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shirasuna K, Shimamura N, Seno K, Ohtsu A,
Shiratsuki S, Ohkuchi A, Suzuki H, Matsubara S, Nagayama S, Iwata H
and Kuwayama T: Moderate hypoxia down-regulates interleukin-6
secretion and TLR4 expression in human Sw.71 placental cells. Cell
Physiol Biochem. 36:2149–2160. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Addison CL, Arenberg DA, Morris SB, Xue
YY, Burdick MD, Mulligan MS, Iannettoni MD and Strieter RM: The CXC
chemokine, monokine induced by interferon-gamma, inhibits non-small
cell lung carcinoma tumor growth and metastasis. Hum Gene Ther.
11:247–261. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Samten B, Townsend JC, Weis SE, Bhoumik A,
Klucar P, Shams H and Barnes PF: CREB, ATF and AP-1 transcription
factors regulate IFN-gamma secretion by human T cells in response
to mycobacterial antigen. J Immunol. 181:2056–2064. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mann DA, Trim J, Smart D, Wright MC and
Michael JP: Control of TIMP-1 gene transcription in hepatic
myofibroblasts by a combination of AP-1 proteins and novel
transcription factors. Int J Exp Pathol. 81:A18–A19. 2000.
View Article : Google Scholar
|
|
40
|
Bonello S, Zähringer C, BelAiba RS,
Djordjevic T, Hess J, Michiels C, Kietzmann T and Görlach A:
Reactive oxygen species activate the HIF-1alpha promoter via a
functional NFkappaB site. Arterioscler Thromb Vasc Biol.
27:755–761. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pialoux V, Mounier R, Brown AD, Steinback
CD, Rawling JM and Poulin MJ: Relationship between oxidative stress
and HIF-1 alpha mRNA during sustained hypoxia in humans. Free Radic
Biol Med. 46:321–326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Guterres FA, Martinez GR, Rocha ME and
Winnischofer SM: Simvastatin rises reactive oxygen species levels
and induces senescence in human melanoma cells by activation of
p53/p21 pathway. Exp Cell Res. 319:2977–2988. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Qi XF, Kim DH, Yoon YS, Kim SK, Cai DQ,
Teng YC, Shim KY and Lee KJ: Involvement of oxidative stress in
simvastatin-induced apoptosis of murine CT26 colon carcinoma cells.
Toxicol Lett. 199:277–287. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Licarete E, Sesarman A and Banciu M:
Exploitation of pleiotropic actions of statins by using
tumour-targeted delivery systems. J Microencapsul. 32:619–631.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
May MJ, D'Acquisto F, Madge LA, Glöckner
J, Pober JS and Ghosh S: Selective inhibition of NF-kappaB
activation by a peptide that blocks the interaction of NEMO with
the IkappaB kinase complex. Science. 289:1550–1554. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Amiri KI, Horton LW, LaFleur BJ, Sosman JA
and Richmond A: Augmenting chemosensitivity of malignant melanoma
tumors via proteasome inhibition: Implication for bortezomib
(VELCADE, PS-341) as a therapeutic agent for malignant melanoma.
Cancer Res. 64:4912–4918. 2004. View Article : Google Scholar : PubMed/NCBI
|