Introduction
Mammary gland tumors are the most common type of
tumor in female dogs, and ~50% of them are malignant (1,2). Thus, it
is a critical disease in veterinary clinical practice. However,
following common cancer therapy (surgery, chemotherapy or
radiotherapy, or their combinations), certain patients with canine
mammary tumor still develop recurrence and/or metastases (2). As with humans, the presence of cancer
stem cells (CSCs) within the tumor mass is a possible explanation
for this clinical issue. CSCs are a small subset of cancer cells,
which constitute a reservoir of self-sustaining cells with the
exclusive ability to self-renew, thereby maintaining the tumor
(3). CSCs are considered to be
responsible for treatment resistance, recurrence and metastasis in
various human cancers (4–6).
Increasing evidence supports the presence of CSCs in
canine mammary tumors (7–10). Human breast CSCs are primarily
characterized by cluster of differentiation
(CD)44+/CD24−/low cells, sphere formation and
active aldehyde dehydrogenase (11).
These characteristics have also been observed in canine mammary
CSCs; a CD44+/CD24−/low phenotype is also a
specific maker of CSCs in canine mammary carcinoma (8,9,12). Another feature of CSCs is
overexpression of stem cell-associated genes, including
octamer-binding transcription factor 4 (Oct-4) and sex determining
region Y-box 2 (Sox-2) (10).
Furthermore, activation of the Wnt/β-catenin, Hedgehog and Notch
signaling pathways confers resistance and survival to CSCs in human
cancer (13). Similar to what has
been observed in human cancer, canine mammary CSCs upregulate stem
cell-associated genes (9) and
activate the Wnt/β-catenin signaling pathway (14). Canine mammary CSCs are highly
tumorigenic, and exhibit multidrug resistance and invasive
potential (9,15); therefore, they serve an important role
in the treatment resistance, recurrence and metastasis of canine
mammary tumors.
Salinomycin (SAL), a polyether ionophore antibiotic
isolated from Streptomyces albus, is widely used in farm
animals as an anticoccidial drug (16). A previous review demonstrated that SAL
is able to selectively deplete CSCs in different types of human
cancer, including breast, gastric, hepatocellular, pancreatic,
prostate and ovarian (13). SAL was
also revealed to inhibit cell invasion and migration in a variety
of cancer types (17). However, the
mechanism of SAL anticancer activity in CSCs is not yet completely
understood. A previous study demonstrated that SAL is a canonical
Wnt/β-catenin signaling pathway inhibitor, through effecting the
expression of numerous Wnt-associated genes, including β-catenin,
Cyclin D1, fibronectin, lymphoid enhancer-binding factor and
low-density lipoprotein receptor-associated protein 6 (18). In addition, the Wnt/β-catenin
signaling pathway appears to be responsible for the antitumor
effects of SAL on human breast cancer (19).
Based on the aforementioned data, new cytotoxic
drugs that target canine CSCs, in addition to those in humans, are
required. Therefore, the present study focused on whether SAL
inhibits canine mammary CSCs. The effects of SAL on the viability,
sphere-forming ability and invasive potential of cells derived from
canine mammary carcinoma were analyzed. In addition, various
proteins (β-catenin, Cyclin D1 and Oct-4) and the
CD44+/CD24−/low cell population were observed
in response to SAL.
Materials and methods
Cell lines and cell culture
Canine mammary carcinoma CMT7364 cells (originally
from mammary gland carcinoma, extracted during surgical resection
at the Veterinary Teaching Hospital of China Agricultural
University) and CIPp cells (20),
[originally from mammary gland adenocarcinoma, a gift from the
Laboratory of Veterinary Surgery (Graduate School of Agricultural
and Life Sciences, University of Tokyo, Tokyo, Japan)] were grown
in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum (FBS; both Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). All cells used in the present study were
incubated at 37°C in an atmosphere containing 5% CO2.
Ethical approval for the present study was obtained from China
Agricultural University (Beijing, China).
Chemicals and antibodies
SAL, doxorubicin (DOX) and cisplatin (DDP) were
purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). The
antibodies used for western blot analysis were as follows: Primary
antibodies, mouse anti-β-catenin (dilution, 1:1,000; cat. no.
sc-133240); mouse anti-β-actin (dilution, 1:2,000; cat. no.
sc-47,778); mouse anti-Oct-4 (dilution, 1:500; cat. no. sc-5279)
and rabbit anti-Cyclin D1 (dilution, 1:1,000; cat. no. sc-753);
secondary antibodies, goat anti-rabbit IgG-HRP (dilution, 1:2,000;
cat. no. sc-2054) and goat anti-mouse IgG-HRP (dilution, 1:2,000;
cat. no. sc-2005; all Santa Cruz Biotechnology, Inc., Dallas, TX,
USA). Antibodies used in the flow cytometric assay were
phycoerythrin anti-mouse CD24 (dilution, 1:200; cat. no. 553261)
and allophycocyanin anti-mouse/human CD44 (dilution, 1:200; cat.
no. 561859; both BD Biosciences, Franklin Lakes, NJ, USA).
Cell viability assay for
chemoresistance
Cell spheres were obtained as previously described
(9). CMT7364 and CIPp cells and their
spheres were plated on 96-well plates at a density of
3×103 cells/well. Cells were treated with a range of
concentrations of DDP (0–80 µM), DOX (0–40 µM) and SAL (0–20 µM)
for 24, 48 and 72 h. Cell viability was measured using the Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan) assay according to the manufacturer's protocol,
then measuring the optical density (OD) with a microplate reader
(ELx808™; BioTek Instruments, Inc., Winooski, VT, USA)
at 450 nm. Cell viability was calculated according to the following
formula: Cell viability (%)=[(OD of test samples)-(OD of
blank)]/[(OD of control samples)-(OD of blank)]x100. The half
maximal inhibitory concentration (IC50) was calculated
with Prism software (version 6.01; GraphPad Software, Inc., La
Jolla, CA, USA).
Sphere formation assay to identify
CSCs
Cells were pretreated with SAL (0, 3 or 6 µM) for 48
h. Cells were then harvested and plated at a density of
2×103 cells/well on an ultralow attachment 6-well plate
(Corning Incorporated, Corning, NY, USA.) in serum-free DMEM/F12
medium (Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10
ng/ml of basic fibroblast growth factor (PeproTech China, Suzhou,
China), 10 ng/ml of epidermal growth factor (PeproTech China), 4
µg/ml of heparin (Sigma-Aldrich; Merck KGaA) and 50X B-27 (Gibco;
Thermo Fisher Scientific, Inc.). The number of spheres was counted
after a 7-day incubation (at 37°C in atmosphere containing 5%
CO2) using an inverted phase-contrast microscope at ×40
magnification. Five fields of view from each plate were selected
and counted, and the results were expressed as the total number of
spheres.
Cell invasion assay
Tumor cell invasion capacity was analyzed using a
Transwell chamber (cat. no. 3422; Corning Incorporated) according
to the protocol of the manufacturer. In brief, the Transwell plate
was coated with 25% Matrigel (cat. no. 354248; BD Biosciences).
DMEM containing 10% FBS was added to the lower chamber, and
2×103 cells were seeded into the upper chamber with
serum-free DMEM containing SAL (0, 3 or 6 µM). The plate was
incubated for 48 h at 37°C, and the upper cells were then scraped
off with a cotton swab. The cells on the lower side of the insert
were fixed with 5% glutaraldehyde for 10 min, followed by staining
with 1% crystal violet in 2% ethanol for an additional 20 min at
room temperature. The number of cells on the lower side of the
filter were viewed under a microscope at ×400 magnification. Five
fields of view were randomly selected and cells were counted in
each, with the results expressed as the mean number of
cells/field.
Flow cytometry
Spheres treated with SAL (0, 3 or 6 µM) for 48 h at
37°C, and their parental cells which were not treated with SAL,
were enzymatically dissociated with 0.25% Trypsin-EDTA (Gibco;
Thermo Fisher Scientific, Inc.). Following washing with PBS plus 2%
FBS, the cells were incubated with antibodies directed against CD44
and CD24 for 30 min at 4°C according to the manufacturer's
protocol. The cell populations were then analyzed within 30 min by
flow cytometry using a BD FACSCalibur flow cytometer (BD
Biosciences), and data were analyzed with FlowJo (version 10.0.7;
FlowJo, LLC, Ashland, OR, USA).
Western blot analysis
Spheres treated with SAL (0, 3 or 6 µM) for 48 h at
37°C and their parental cells were incubated with
radioimmunoprecipitation assay buffer with phenylmethanesulfonyl
fluoride. Equal amounts of protein (30 µg) were resolved on a 10%
gel using SDS-PAGE, then transferred to polyvinylidene fluoride
(PVDF) membranes (Pall Life Sciences, Port Washington, NY, USA).
The membrane was blocked with 5% milk in PBS-Tween-20 at room
temperature for 1 h. The primary antibodies (anti-β-catenin,
anti-Cyclin D1 and anti-Oct-4) were then diluted in blocking buffer
and added to individual PVDF membranes that were incubated at 4°C
overnight. The next day, the membranes were incubated with
horseradish peroxidase-conjugated secondary antibodies for 2 h at
room temperature and protein bands were detected with an enhanced
chemiluminescence substrate (cat. no. 34080; Thermo Fisher
Scientific, Inc.). β-actin was used as an internal control.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software (version 6.01). Data are presented as the mean ±
standard deviation. A student's test was used when 2 groups were
being compared, and a one-way analysis of variance when 3 groups
were being compared. P<0.05 was considered to indicate a
statistically significant difference.
Results
CMT7364 and CIPp spheres exhibit
characteristics of CSCs
CMT7364 and CIPp spheres (Fig. 1A) derived from parental cells, under
anchorage-independent conditions and in the absence of FBS,
exhibited characteristics of CSCs. Firstly, spheres exhibited a
CD44+/CD24−/low phenotype and expressed an
increased level of CD44 compared with their parental cells
(Fig. 1B). In addition, western blot
analysis demonstrated that stem cell marker Oct-4 and the
Wnt/β-catenin-associated proteins β-catenin and cyclin D1 were
overexpressed in spheres when compared with their parental cells
(Fig. 1C). The IC50 was
then calculated using a CCK-8 assay. The results demonstrated that
the IC50 of DDP and DOX were significantly increased in
spheres compared with their parental cells (P<0.05; Fig. 1D). These results indicate that the
spheres derived from CMT7364 and CIPp cells are CSCs.
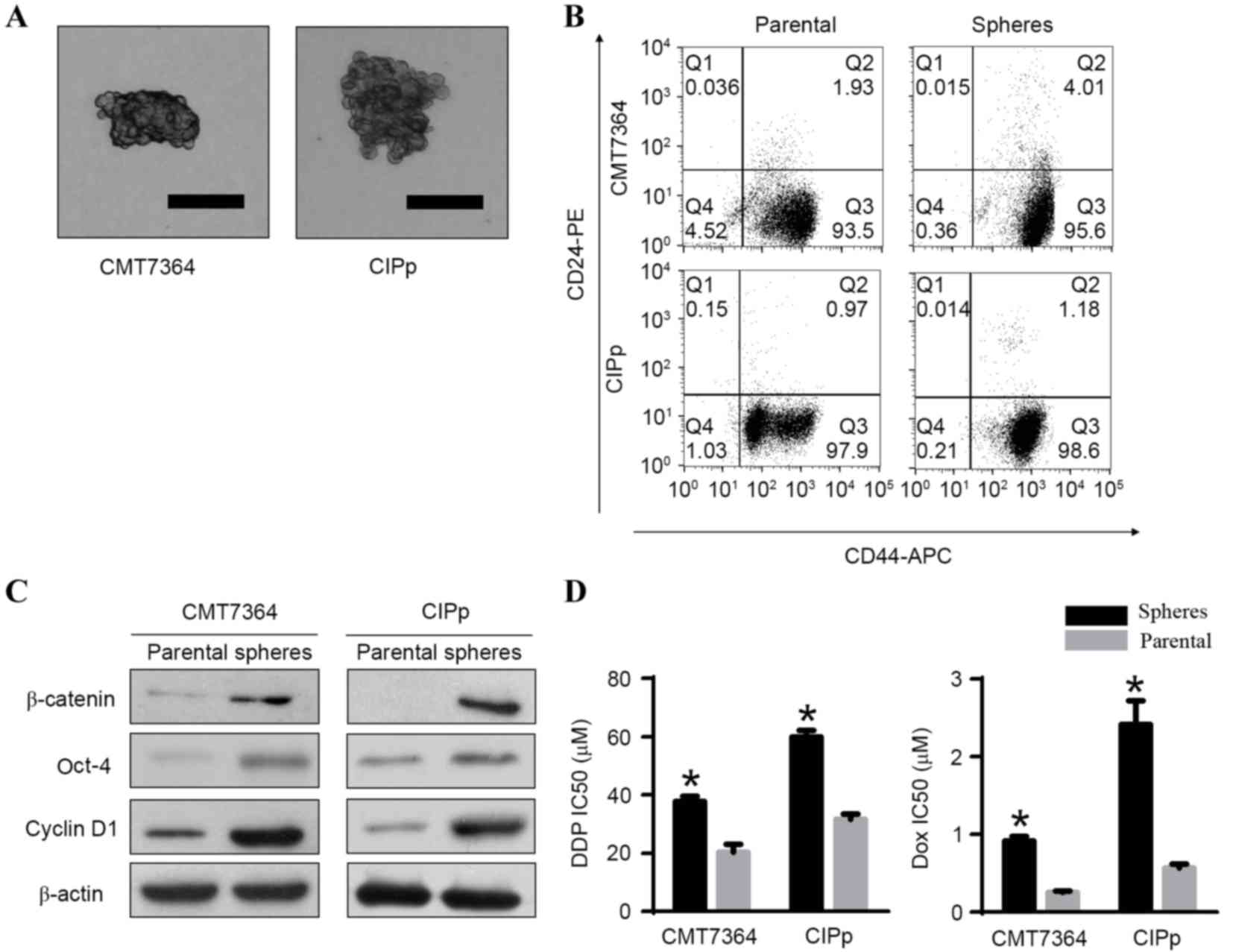 | Figure 1.Canine mammary cancer stem cells were
identified using a sphere-formation assay and flow cytometry, and
their expression of Wnt/β-catenin signaling pathway-associated
proteins and chemoresistance analyzed. (A) Representative
morphology of spheres derived from the canine mammary carcinoma
cell lines CMT7364 and CIPp (scale bar, 100 µM). (B) Flow
cytometric analysis of the expression of CD44 and CD24 in spheres
and their parental cells. The cells in Q3 correspond to
CD44+/CD24−/low cells. Results are
demonstrated as representative images of three independent
experiments. (C) Protein levels of β-catenin, Oct-4 and Cyclin D1
in spheres and their parental cells, measured by western blot
analysis. These proteins were overexpressed in spheres compared
with their parental cells. (D) The 48 h IC50 of DDP and
DOX in spheres and their parental cells. Results are presented as
the mean ± standard deviation (n=3). *P<0.05 vs. the parental
cells. IC50, half maximal inhibitory concentration; CD,
cluster of differentiation; PE, phycoerythrin; APC,
allophycocyanin; Oct-4, octamer-binding transcription factor 4; Q,
quadrant; DDP, cisplatin; DOX, doxorubicin. |
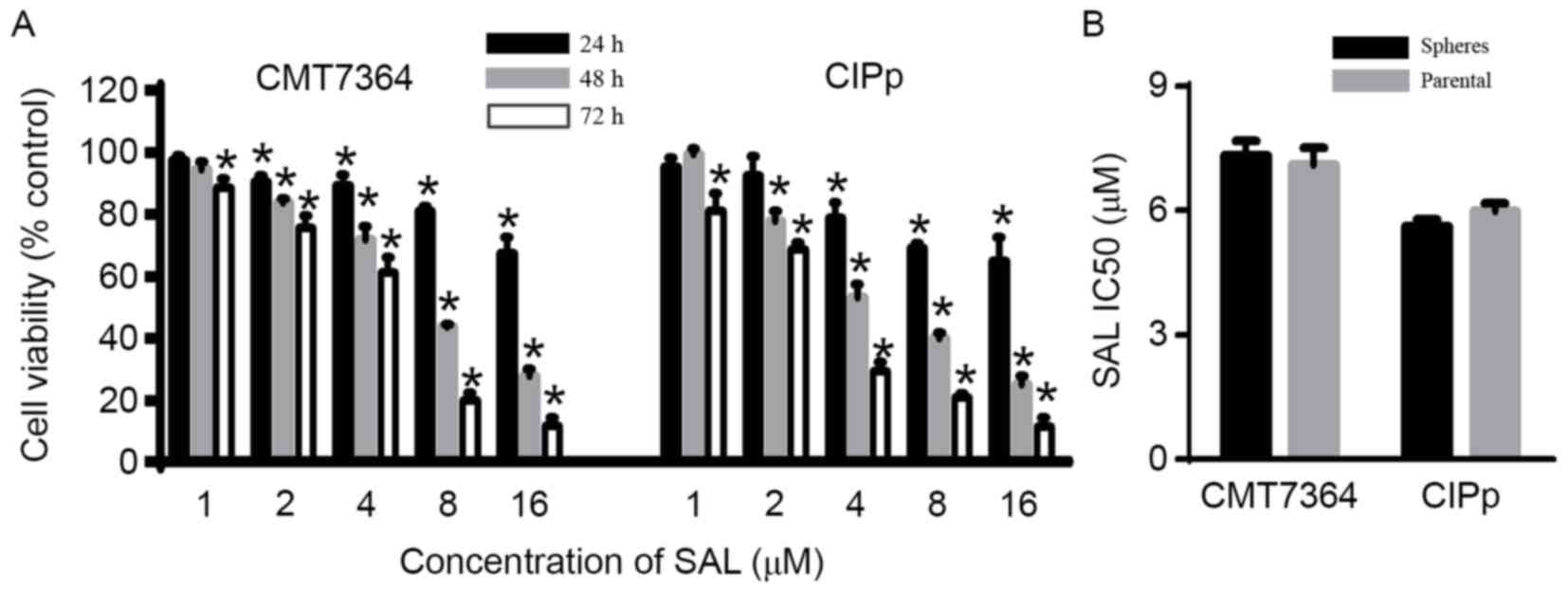
SAL inhibits the viability,
sphere-forming ability and expression of stem cell-associated
proteins of CMT7364 and CIPp spheres
The present study aimed to determine whether SAL
targets canine mammary CSCs. The cytotoxic effect of SAL was
evaluated using the CCK-8 assay. The results demonstrated that SAL
reduced cell viability in a time- and concentration-dependent
manner (P<0.05; Fig. 2A), and the
48 h IC50 of SAL in spheres was similar compared with
that of the parental cells (Fig. 2B).
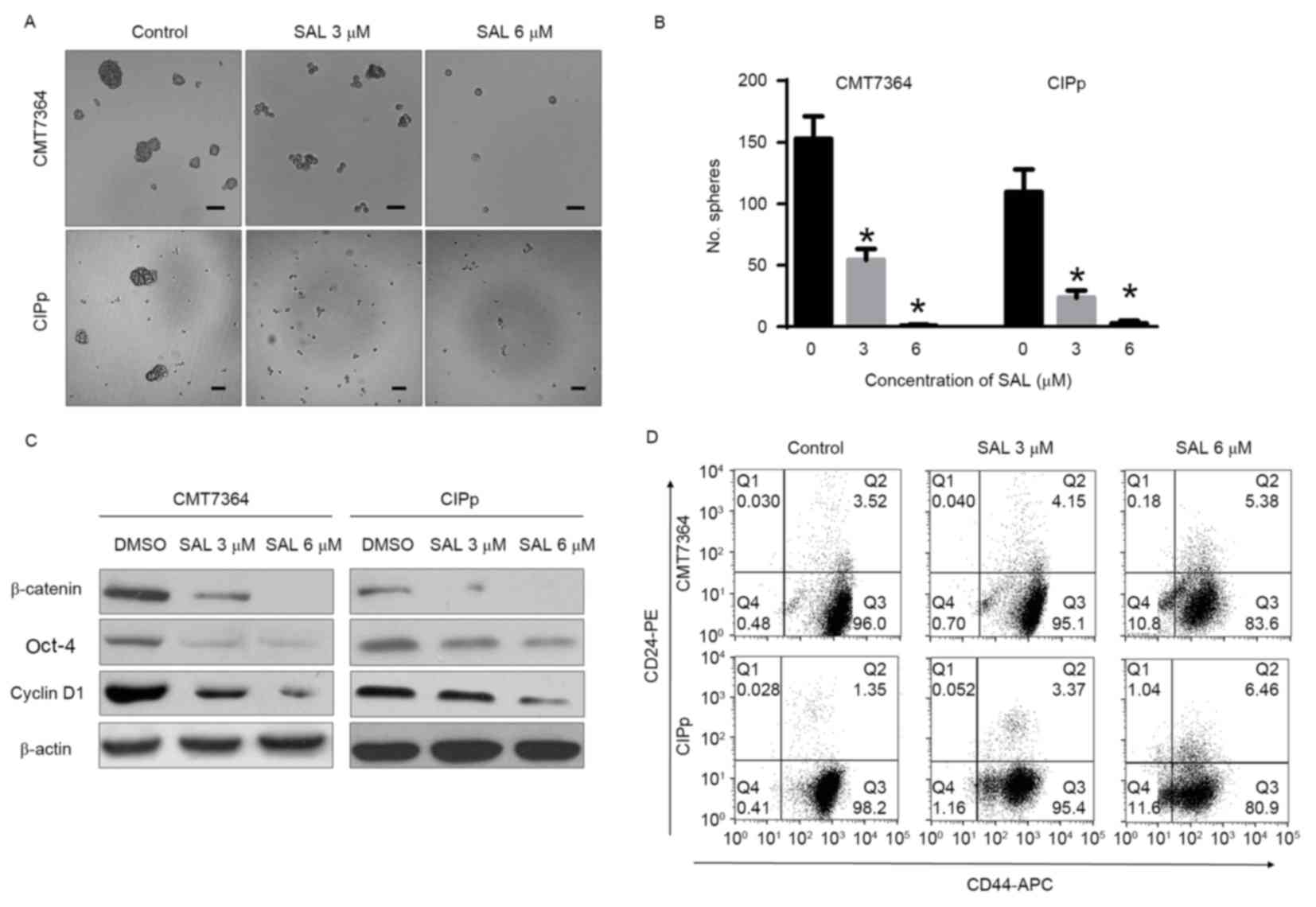
In addition, it was identified that SAL significantly inhibited
canine mammary CSC sphere-formation, reducing the size and number
of spheres (P<0.05; Fig. 3A and
B). Western blot analysis revealed that the protein expression
of β-catenin, Cyclin D1 and Oct-4 in spheres was downregulated
following treatment with SAL (Fig.
3C). To confirm that SAL targets canine mammary CSCs, the
CD44+/CD24−/low phenotype of the spheres was
measured following treatment with SAL. As illustrated in Fig. 3D, SAL reduced the proportion of
CD44+/CD24−/low cells in the overall sphere
population.
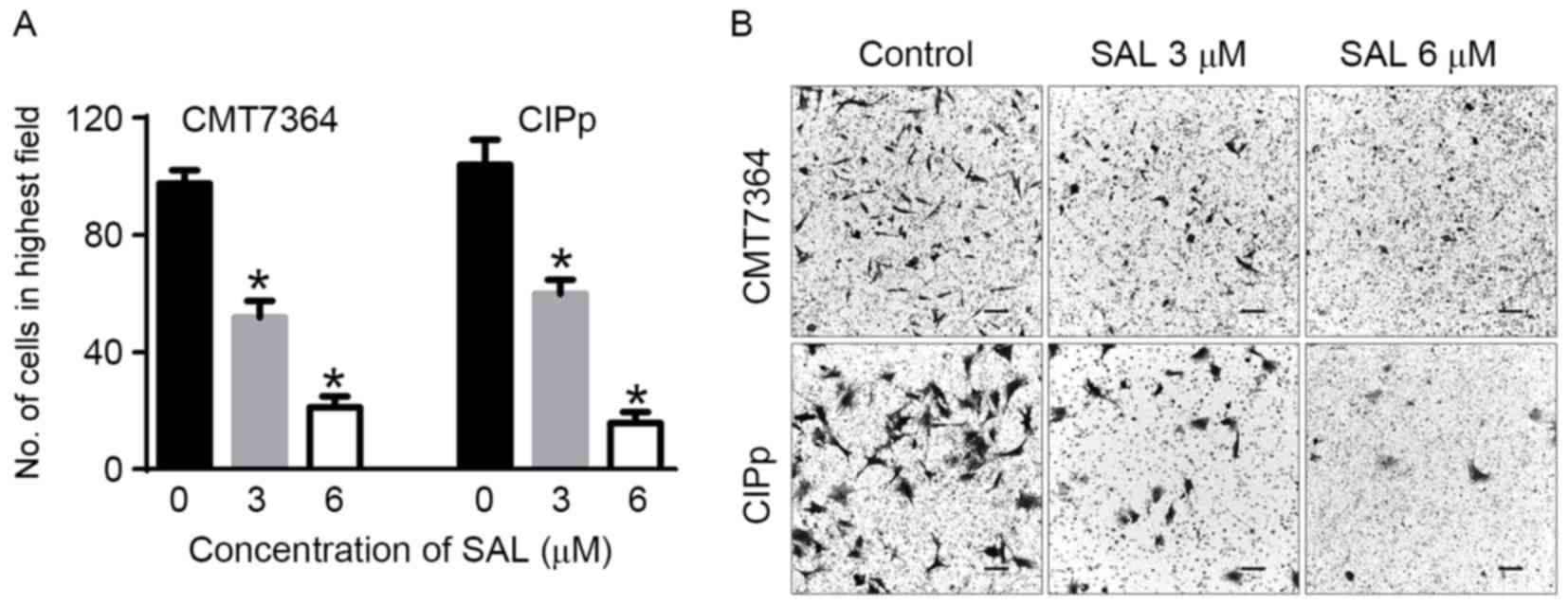
SAL inhibits the invasive potential of
CMT7364 and CIPp spheres
Since CSCs have a high invasive potential, a
Transwell assay was performed to evaluate the effect of SAL on the
invasion of the spheres (Fig. 4).
Compared with untreated cells, the number of cells that invaded the
lower side of the Transwell chamber was significantly lower
(P<0.05).
Discussion
Increasing evidence supports the CSC theory, which
hypothesizes that CSCs are responsible for tumor growth, therapy
resistance, recurrence and metastasis (3,4,6,21,22). In canines, mammary CSCs are associated
with a higher grade of carcinoma and poor prognosis (7,8). In
addition, canine mammary CSCs are typically resistant to common
cancer therapies (9,15). Therefore, it is important to identify
novel drugs targeting CSCs for the eradication of canine, and
human, mammary tumors. A previous review has demonstrated that SAL
selectively depletes CSCs in different types of human cancer,
likely by interfering with ATP-binding cassette drug transporters,
the Wnt/β-catenin signaling pathway and other CSC-associated
signaling pathways (13). Thus, the
present study investigated whether SAL inhibits CSCs in canine
mammary tumors.
The sphere formation assay is a useful tool to
identify CSCs. Previous studies have used this method to analyze
the sphere-forming ability of canine mammary CSCs (15,23). This
method has also been used to investigate novel therapeutic drugs in
humans and canines (14,24). In the present study, it was observed
that spheres from canine mammary carcinoma cell lines overexpressed
the stem cell markers CD44 and Oct-4, and exhibited chemoresistance
to DOX and DDP. These findings indicate that the spheres derived
from CMT7364 and CIPp are CSCs.
Spheres may form single suspended cells under
serum-free conditions, which is important for the self-renewal
capacity of CSCs (9). The present
study demonstrated that SAL reduced cell viability in a time- and
concentration-dependent manner, and also inhibited sphere formation
by reducing the number and size of spheres. In addition, the
IC50 of SAL in spheres and their parental cells were
similar, which indicates that canine mammary CSCs are not resistant
to SAL. A CD44+/CD24−/low phenotype is a
specific maker of CSCs in canine mammary carcinoma (8). In the present study, SAL treatment
reduced the CD44+/CD24−/low cell population.
A previous study identified that Oct-4 is as an adequate marker of
CSCs of canine mammary tumors (25),
and overexpression of Oct-4 has been associated with malignancy and
poor outcomes (26). Thus, the level
of expression change of Oct-4 after drug treatment may be used to
evaluate the ability to specifically target CSCs. In the present
study, the expression of Oct-4 protein was decreased in spheres
following treatment with SAL. These findings suggest that SAL
inhibits CSCs in canine mammary tumors.
The canonical Wnt/β-catenin signaling pathway serves
an essential role in cell proliferation and migration, and in the
self-renewal of CSCs (27). A
previous study has identified that this signaling pathway is
upregulated in human breast CSCs (28). Similar to in humans, the present study
revealed that stem cell-like spheres overexpressed the
Wnt/β-catenin signaling pathway-associated proteins β-catenin and
Cyclin D1, indicating that this signaling pathway is also
upregulated in canine mammary CSCs. Previous studies have
demonstrated that SAL targets CSCs in various tumors, including
gastric cancer, osteosarcoma and lung cancer, by impacting the
Wnt/β-catenin signaling pathway (29–31).
Notably, in the present study the protein level of β-catenin and
Cyclin D1 in spheres was decreased following treatment with SAL.
This suggests that the Wnt/β-catenin signaling pathway is
associated with the effect of SAL on canine mammary CSCs.
Tumor metastasis is a major cause of mortality in
humans and canines with cancer. CSCs possess migration and invasion
potential, driving metastatic tumor formation (21,22).
However, there are few effective treatments for patients with
metastatic disease. Recently, studies have indicated that SAL
inhibits cell migration and invasion in a variety of cancer types
in vitro (17), in addition to
reducing metastasis in vivo (32,33). In
the present study, the invasive ability of canine mammary CSCs was
decreased following treatment with SAL in vitro; however,
whether SAL has inhibits the metastasis of canine mammary tumor
cells requires additional study.
In conclusion, the present study identified that SAL
is an effective inhibitor of canine mammary CSCs in vitro,
indicating that SAL is a promising chemotherapeutic for the
treatment of canine mammary carcinoma.
Acknowledgements
The authors thank the Laboratory of Veterinary
Surgery of the Graduate School of Agricultural and Life Sciences at
the University of Tokyo, for their gift of the canine mammary
carcinoma cell line CIPp. The present study was supported by the
National Natural Science Foundation of China (grant no.
31372489).
References
|
1
|
Vail DM and MacEwen EG: Spontaneously
occurring tumors of companion animals as models for human cancer.
Cancer Invest. 18:781–792. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sleeckx N, De Rooster H, Kroeze EJ
Veldhuis, Van Ginneken C and Van Brantegem L: Canine mammary
tumours, an overview. Reprod Domest Anim. 46:1112–1131. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Clarke MF, Dick JE, Dirks PB, Eaves CJ,
Jamieson CH, Jones DL, Visvader J, Weissman IL and Wahl GM: Cancer
Stem cells-perspectives on current status and future directions:
AACR workshop on cancer stem cells. Cancer Res. 66:9339–9344. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Visvader JE and Lindeman GJ: Cancer stem
cells in solid tumours: Accumulating evidence and unresolved
questions. Nat Rev Cancer. 8:755–768. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hu Y and Fu L: Targeting cancer stem
cells: A new therapy to cure cancer patients. Am J Cancer Res.
2:340–356. 2012.PubMed/NCBI
|
|
6
|
Mitra A, Mishra L and Li S: EMT, CTCs and
CSCs in tumor relapse and drug-resistance. Oncotarget.
6:10697–11711. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Im KS, Jang YG, Shin JI, Kim NH, Lim HY,
Lee SM, Kim JH and Sur JH: CD44+/CD24- cancer stem cells are
associated with higher grade of canine mammary carcinomas. Vet
Pathol. 52:1041–1044. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Magalhães GM, Terra EM, de Oliveira
Vasconcelos R, de Barros Bandarra M, Moreira PR, Rosolem MC and
Alessi AC: Immunodetection of cells with a CD44+/CD24- phenotype in
canine mammary neoplasms. BMC Vet Res. 9:2052013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Michishita M, Akiyoshi R, Yoshimura H,
Katsumoto T, Ichikawa H, Ohkusu-Tsukada K, Nakagawa T, Sasaki N and
Takahashi K: Characterization of spheres derived from canine
mammary gland adenocarcinoma cell lines. Res Vet Sci. 91:254–260.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Leis O, Eguiara A, Lopez-Arribillaga E,
Alberdi MJ, Hernandez-Garcia S, Elorriaga K, Pandiella A, Rezola R
and Martin AG: Sox2 expression in breast tumours and activation in
breast cancer stem cells. Oncogene. 31:1354–1365. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Oliveira LR, Jeffrey SS and Ribeiro-Silva
A: Stem cells in human breast cancer. Histol Histopathol.
25:371–385. 2010.PubMed/NCBI
|
|
12
|
Michishita M, Akiyoshi R, Suemizu H,
Nakagawa T, Sasaki N, Takemitsu H, Arai T and Takahashi K: Aldehyde
dehydrogenase activity in cancer stem cells from canine mammary
carcinoma cell lines. Vet J. 193:508–513. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Naujokat C and Steinhart R: Salinomycin as
a drug for targeting human cancer stem cells. J Biomed Biotechnol.
2012:9506582012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Torres CG, Olivares A and Stoore C:
Simvastatin exhibits antiproliferative effects on spheres derived
from canine mammary carcinoma cells. Oncol Rep. 33:2235–2244.
2015.PubMed/NCBI
|
|
15
|
Pang LY, Cervantes-Arias A, Else RW and
Argyle DJ: Canine mammary cancer stem cells are radio- and chemo-
resistant and exhibit an epithelial-mesenchymal transition
phenotype. Cancers (Basel). 3:1744–1762. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou S, Wang F, Wong ET, Fonkem E, Hsieh
TC, Wu JM and Wu E: Salinomycin: A novel anti-cancer agent with
known anti-coccidial activities. Curr Med Chem. 20:4095–4101. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kopp F, Hermawan A, Oak PS, Herrmann A,
Wagner E and Roidl A: Salinomycin treatment reduces metastatic
tumor burden by hampering cancer cell migration. Mol Cancer.
13:162014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu D, Choi MY, Yu J, Castro JE, Kipps TJ
and Carson DA: Salinomycin inhibits Wnt signaling and selectively
induces apoptosis in chronic lymphocytic leukemia cells. Proc Natl
Acad Sci USA. 108:pp. 13253–13257. 2011; View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lu W and Li Y: Salinomycin suppresses LRP6
expression and inhibits both Wnt/β-catenin and mTORC1 signaling in
breast and prostate cancer cells. J Cell Biochem. 115:1799–1807.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Uyama R, Nakagawa T, Hong SH, Mochizuki M,
Nishimura R and Sasaki N: Establishment of four pairs of canine
mammary tumour cell lines derived from primary and metastatic
origin and their E-cadherin expression. Vet Comp Oncol. 4:104–113.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dalerba P and Clarke MF: Cancer stem cells
and tumor metastasis: First steps into uncharted territory. Cell
Stem Cell. 1:241–242. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li F, Tiede B, Massagué J and Kang Y:
Beyond tumorigenesis: Cancer stem cells in metastasis. Cell Res.
17:3–14. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Grange C, Lanzardo S, Cavallo F, Camussi G
and Bussolati B: Sca-1 identifies the tumor-initiating cells in
mammary tumors of BALB-neuT transgenic mice. Neoplasia.
10:1433–1443. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Simmons MJ, Serra R, Hermance N and
Kelliher MA: NOTCH1 inhibition in vivo results in mammary tumor
regression and reduced mammary tumorsphere-forming activity in
vitro. Breast Cancer Res. 14:R1262012. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ferletta M, Grawé J and Hellmén E: Canine
mammary tumors contain cancer stem-like cells and form spheroids
with an embryonic stem cell signature. Int J Dev Biol. 55:791–799.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang J, Zhang D, Xie F and Lin D: The
potential role of COX-2 in cancer stem cell-mediated canine mammary
tumor initiation: An immunohistochemical study. J Vet Sci.
16:225–231. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Curtin JC and Lorenzi MV: Drug discovery
approaches to target Wnt signaling in cancer stem cells.
Oncotarget. 1:552–566. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao Z, Lu P, Zhang H, Xu H, Gao N, Li M
and Liu C: Nestin positively regulates the Wnt/β-catenin pathway
and the proliferation, survival and invasiveness of breast cancer
stem cells. Breast Cancer Res. 16:4082014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tang QL, Zhao ZQ, Li JC, Liang Y, Yin JQ,
Zou CY, Xie XB, Zeng YX, Shen JN, Kang T and Wang J: Salinomycin
inhibits osteosarcoma by targeting its tumor stem cells. Cancer
Lett. 311:113–121. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mao J, Fan S, Ma W, Fan P, Wang B, Zhang
J, Wang H, Tang B, Zhang Q, Yu X, et al: Roles of Wnt/β-catenin
signaling in the gastric cancer stem cells proliferation and
salinomycin treatment. Cell Death Dis. 5:e10392014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang X, Lou Y, Zheng X, Wang H, Sun J,
Dong Q and Han B: Wnt blockers inhibit the proliferation of lung
cancer stem cells. Drug Des Devel Ther. 9:2399–2407.
2015.PubMed/NCBI
|
|
32
|
Wang F, He L, Dai WQ, Xu YP, Wu D, Lin CL,
Wu SM, Cheng P, Zhang Y, Shen M, et al: Salinomycin inhibits
proliferation and induces apoptosis of human hepatocellular
carcinoma cells in vitro and in vivo. PLoS One. 7:e506382012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Qu H, Ma B, Yuan HF, Wang ZY, Guo SJ and
Zhang J: Effect of salinomycin on metastasis and invasion of
bladder cancer cell line T24. Asian Pac J Trop Med. 8:578–582.
2015. View Article : Google Scholar : PubMed/NCBI
|