Introduction
Multiple myeloma (MM) is a malignancy, which is
characterized by plasma cell hyperplasia in the bone marrow and
strain integrity of monoclonal immunoglobulin (IgG, IgA, IgD or
IgE) or Bence Jones protein (free monoclonal light chains κ or γ)
overexpression (1). Despite active
chemotherapy regimes and autologous stem cell transplantation, the
majority of MM cases result in mortalities due to tumor recurrence
and metastasis, with the incidence and mortality rate on the
increase year by year (2). Invasion
is a prerequisite of metastasis, which is a leading cause of
mortality from tumors and MM (3,4).
Therefore, inhibiting cell invasion may benefit patients who have
relapsed or have metastatic MM. However, the precise molecular
mechanisms of cell invasion in MM remain to be elucidated.
Transforming growth factor β1 (TGFβ1) is
ubiquitously expressed. TGFβ1 has been reported to stimulate the
production of angiogenic factors and has a critical role in the
development and progression of MM (5). In a previous study of aggressive
myeloma, cells secreting TGFβ1 induced tumor epithelial-mesenchymal
transition (EMT) and promoted tumor metastasis (6). SMAD family member 2 (Smad2), which
mediates TGFβ1 signaling, has also been reported to be involved in
the migration of tumor cells (7).
The role of matrix metalloproteinases (MMPs) in
tumor cell metastasis has been well studied, and they been
demonstrated to degrade the basement membrane and extracellular
matrix (ECM), which is associated with tumor invasion and
metastasis (8). MMP3, a member of the
family of matrix metalloproteinases, promotes tumor cell invasion
and metastasis along the basement membrane via degradation of the
ECM (9). It has also been reported
that TGFβ1 stimulates the expression of MMP3 in human corneal
epithelial cells (10); however, to
the best of our knowledge, this has not been documented in MM
cells.
In the present study, the role of SB431542 in the
inhibition of cell invasion in multiple myeloma cells and the
mechanism of this process were investigated. Furthermore, this
present study provided evidence for the involvement of the
TGFβ1/Smad2/MMP3 signaling pathway in the SB431542-induced
reduction in cell invasion in multiple myeloma RPMI 8226 cells.
Materials and methods
Cell culture and reagents
RPMI 8226 cells were purchased from American Type
Culture Collection (Manassas, VA, USA). Cells were cultured with
RPMI 1640 medium with 15% fetal bovine serum (both Hyclone; GE
Healthcare Life Sciences, Logan, USA), in a humidified incubator at
37°C with 5% CO2 for 2 to 3 days. Cells in the
logarithmic growth phase were used for subsequent experiments. The
primary antibodies anti-glyceraldehyde-3-phosphate dehydrogenase
(GAPDH; cat. no. sc-47724; dilution, 1:1,000; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), anti-TGFβ1 (cat. no. sc-146;
dilution, 1:500; Santa Cruz Biotechnology, Inc.), anti-smad2 (cat.
no. 12,052), anti-p-smad2 (cat. no. 11,958) and anti-MMP3 (cat. no.
14351S) (both dilution, 1:1,000) were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). The bovine anti-goat IgG
horseradish peroxidase-conjugated secondary antibody (cat. no.
sc-2352; dilution, 1:1,000) was purchased from Santa Cruz
Biotechnology, Inc.
Cell counting kit-8 (CCK-8) cell
proliferation assay
RPMI 8226 cells (1×104/100 µl) were
seeded in a 96-well plate for 24 h and subsequently incubated at
37°C in the presence and absence of SB431542 (1.0, 10, 100 and
1,000 nmol/ml; Med Chem Express Co., Monmouth Junction, NJ, USA)
for 12, 24 and 48 h. CCK-8 solution (10%; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was added, and the plate was
incubated at 37°C for 3 h. The optical density (OD) of each well
was measured at 450 nm using a microplate reader (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The OD values were
calculated based on the formula: Proliferation inhibition
rate=(1-mean OD experimental value/mean OD control value)x100%.
Cell motility assay
Cell invasion was assessed by Transwell assay (pore
size, 8 µm; Corning Inc., Corning, NY, USA). In brief, cells
(1×104/well) were treated with SB431542 or
lentiviral-TGFβ1 vectors and the untreated cells (6×104)
were seeded in the upper chamber. The cells were attached to porous
polycarbonate membranes, which were coated with Matrigel basement
membrane matrix. The lower chamber was filled with RPMI 1640 medium
with 15% fetal bovine serum. Cells that failed to attach to
polycarbonate membranes were removed by washing with PBS following
30 h incubation at 37°C, whilst invaded cells were fixed with 4%
paraformaldehyde for 30 min and subsequently stained with crystal
violet dissolved in 1% SDS for 30 min at 20°C. The number of cells
in 5 randomly selected visual fields at ×200 magnification under a
Olympus CX23 light microscope (Olympus Corporation, Tokyo, Japan)
was recorded.
Western blot analysis
Cell lysis buffer [20 mmol/l Tris (pH 7.5), 150
mmol/l NaCl, 1% Triton X-100, protease and phosphate inhibitors
(0.7 µg/ml) was added to the cells, which were incubated on ice for
30 min. The cells were subsequently centrifuged at 6,720 × g at 4°C
for 12 min, and the supernatants were obtained. The concentration
of the total protein was measured using the BCA protein assay
(Thermo Fisher Scientific, Inc., Waltham, MA, USA), and protein was
heated to 100°C with 4 µl loading buffer for 8 min. Protein (40 µg)
was subsequently separated using SDS-PAGE (8% gel) and transferred
to polyvinylidene difluoride membranes. The membranes were blocked
with 5% fat-free milk or 5% bovine serum albumin (Beyotime
Institute of Biotechnology, Haimen, China) for 2 h. The membranes
were subsequently incubated with the previously mentioned primary
antibodies overnight at 4°C and secondary antibodies for 2 h at
room temperature. The blots were visualized using the
Millipore-Immobilon ECL kit (EMD Millipore, Billerica, MA, USA) and
the blots were quantified with Quantity One software (version 4.62;
Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Preparation and transfection of the
lentiviral-TGFβ1 vector
Double digests of the PLVX-IRES-mcherry plasmid
(Laboratory of Neuron Biotechnology Company, Shanghai, China) and
vector (GV287), containing the TGFβ1 gene, with AgeI (5
U/µl) and EcoRI (20 U/µl) (both Beyotime Institute of
Biotechnology) were performed. The product was recovered, purified
and mixed with 1 µl T4 DNA ligase at 16°C for 6 h. The
transformation of competent DH5α cells (Takara Bio, Inc., Otsu,
Japan) was performed. Briefly, the plasmids 1 ng (5 ul) were cooled
in ice for 2 min and 100 µl of competent cells were added to the
plasmids. The cells were kept in an ice bath for 5 min and then
plated onto agar plates containing X-Gal, IPTG and ampicillin.
White colonies were selected for. Recombinant positive clones were
identified by polymerase chain reaction and restriction analysis.
The PCR thermocycling conditions used were as follows: Denaturation
step, 95°C for 5 min; annealing step, 55°C for 30 sec; elongation
step, 72°C for 1 min for 35 cycles; 72°C for 10 min; and held at
4°C. A Taq DNA polymerase kit (Takara Bio, Inc.) was used for PCR.
Primer sequences were as follows: TGFβ1 forward,
5′-ATAAGCTTGATATCGAATTCCACAGAGCCTTCTCGG−3′ and reverse,
5′-GAGGATCCCCGGGTACCGGTCGCCACCATGGCGCTCTTCGTGCGGC-3′. Plasmids were
sent to Biotechnology Co. Ltd. (Shanghai, China) for sequencing.
RPMI 8226 cells, in the logarithmic growth phase, were seeded in
6-well plates (2×105 cells/ml) and cultured for 24 h.
The lentiviral-TGFβ1 vector was added with a multiplicity of
infection value of 80 to RPMI 8226 cells. RPMI 8226 cells were also
transfected with GV287 vectors. Non-transfected RPMI 8226 cells
served as negative controls. Transfection efficiency was detected
by fluorescence microscopy and western blotting at 6 days
post-transfection.
Statistical analysis
All experiments were independently performed in
triplicate. Student's t-test was performed to compare between two
independent sample groups. All statistical analyses were performed
using GraphPad Prism software (version 5.01; GraphPad Software,
Inc., La Jolla, CA, USA). One-way analysis of variance (ANOVA) was
used in conjunction with Fisher's least significant difference
post-hoc test to analyze the difference between the concentration
groups, and one-way ANOVA with Tukey's test was performed to
compare the differences among different treatment groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
SB431542 reduces cell proliferation
and invasion in RPMI 8226 cells
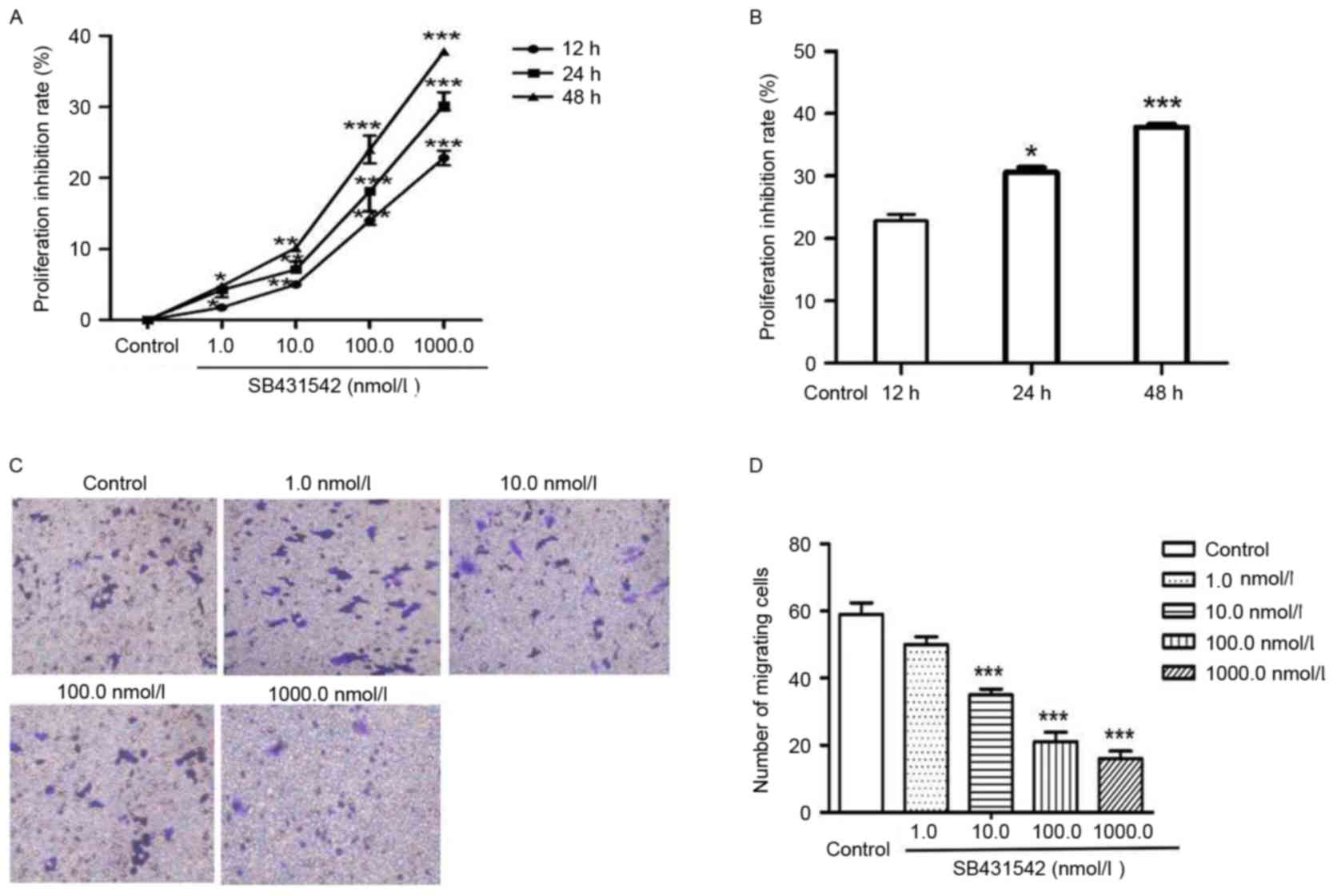
To investigate the function of SB431542 on cell
proliferation and invasion, RPMI 8226 cells were treated with 1,
10, 100 and 1,000 nmol/ml SB431542 (Fig.
1A). A significant dose-dependent decrease in cell
proliferation was observed between 1 and 1,000 nmol/l SB431542. The
cells were subsequently treated with 1,000 nmol/ml SB431542 for 12,
24 and 48 h (Fig. 1B). Cell
proliferation was also inhibited by SB431542 in a dose- and
time-dependent manner. Transwell assay was performed to assess cell
invasion (Fig. 1C). The number of
migrating cells in the control and SB431542 treatment groups (1,
10, 100 and 1,000 nmol/ml) was 59±6, 50±4, 35±3, 21±5 and 16±4,
respectively (Fig. 1D). A significant
difference was observed between the control and the 10, 100 and
1,000 nmol/ml SB431542 treatment groups (P<0.05). No significant
difference was observed between the control and the lowest SB431542
dosage group (1 nmol/ml).
SB431542 reduces TGFβ1 accumulation,
Smad2 activation and MMP3 expression
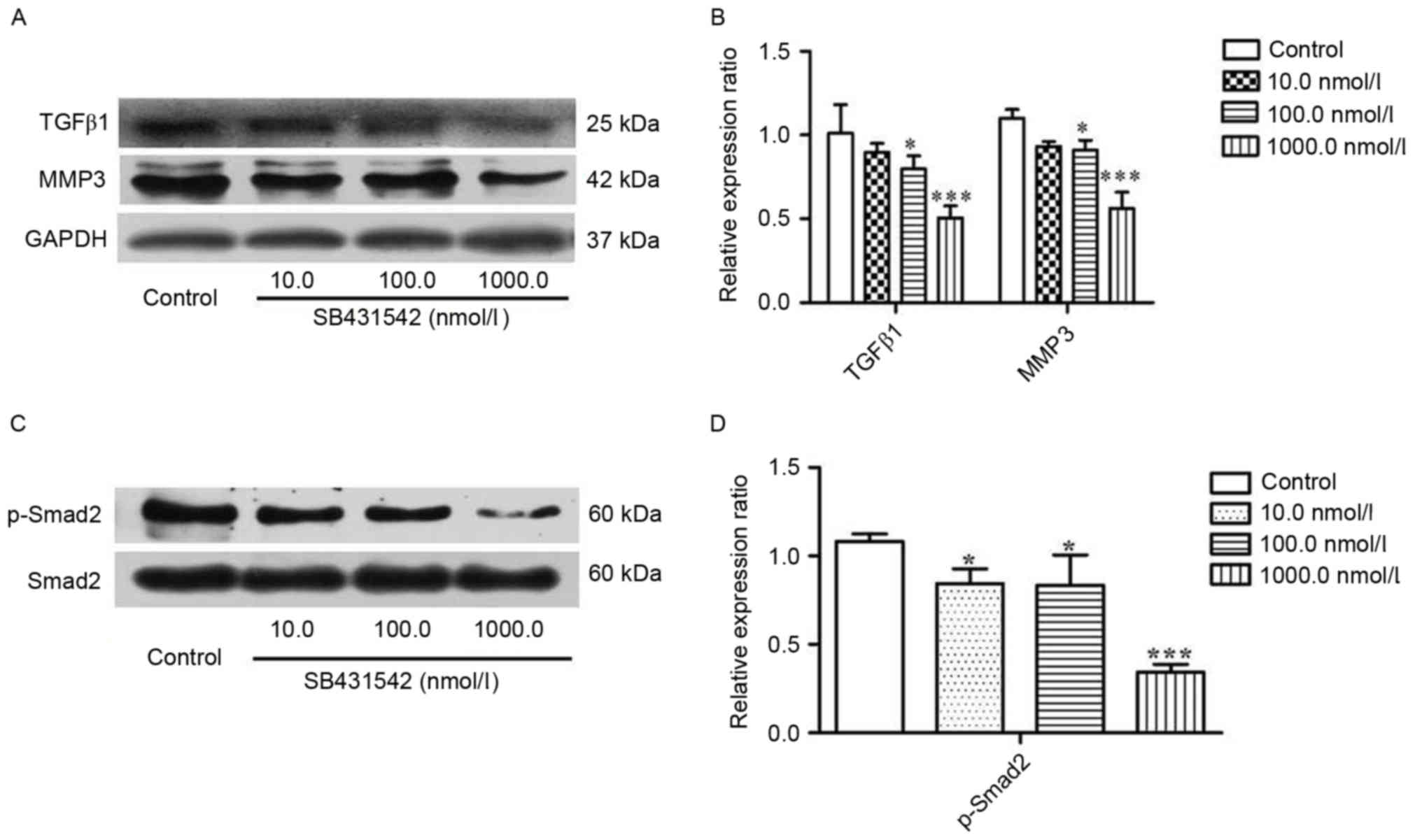
To investigate the effect of SB431542 on the
accumulation of TGFβ1 and MMP3 in MM cells, RPMI 8226 cells were
treated with 10, 100 and 1,000 nmol/ml SB431542 for 48 h. Western
blotting was performed. The results demonstrated that SB431542
significantly decreased the accumulation of TGFβ1 and MMP3 at 100
and 1,000 nmol/ml when compared with the control (Fig. 2A and B). Smad2, as a signaling
molecule, can be initially activated by TGFβ1. Therefore, the
effect of SB431542 on the expression of Smad2 and phosphorylated
(p)-Smad2 in RPMI 8226 cells was investigated. There was no
difference in the levels of Smad2 between the different SB431542
treatment groups and the control, but a significant increase in the
level of p-Smad2 was demonstrated (Fig.
2C and D), indicating that SB431542 was able to repress the
TGFβ1/Smad2 signaling pathway in RPMI 8226 cells.
Increased expression of p-smad2 and
MMP3 in RPMI 8226 cells overexpressing TGFβ1
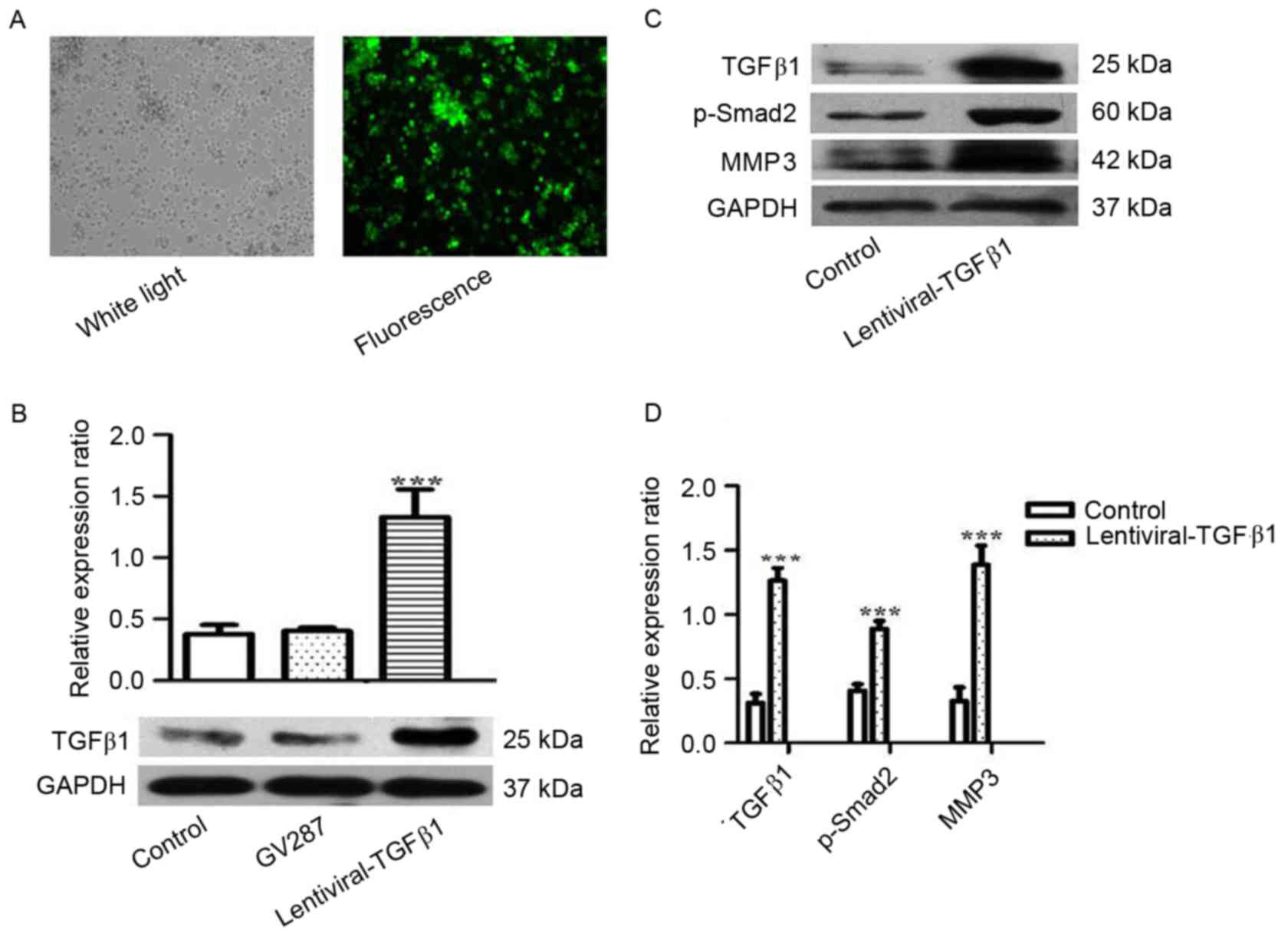
To investigate the role of TGFβ1 in the metastasis
of MM, RPMI 8226 cells were treated with lentiviral-TGFβ1 vectors
to stably express TGFβ1. Transfection efficiency was observed under
fluorescence microscopy and detected by western blotting (Fig. 3A and B). The expression of TGFβ1,
p-Smad2 and MMP3 was analyzed, and increased expression of p-smad2
and MMP3 was observed in the cells overexpressing TGFβ1 compared
with the control (Fig. 3C and D).
Cell invasion and MMP3 expression in
TGFβ1 overexpressing RPMI 8226 cells is partly inhibited by
SB431542
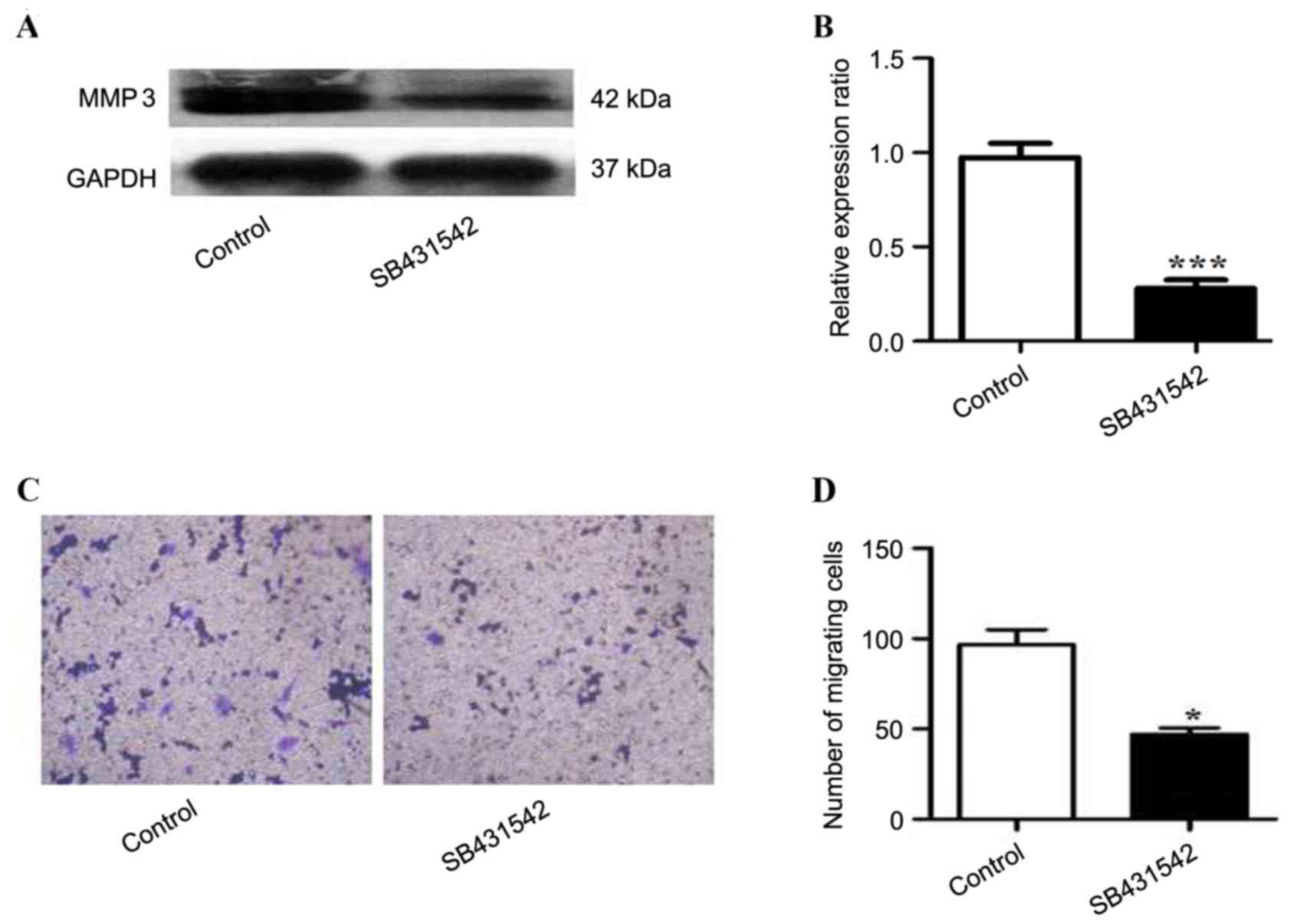
To investigate the role of SB431542 in RPMI 8226
cells overexpressing TGFβ1, the expression of MMP3 in the cells
treated with SB431542 was analyzed. A significant decrease in MMP3
expression was observed in SB431542 treated group when compare to
the control (Fig 4A and B). Cell
invasion was assessed by Transwell assay, and a decrease in the
number of migrating cells was observed in the SB431542 treated
group (Fig. 4C and D).
Discussion
Human MM is a hematological malignancy, and at
present there is limited treatment available (11), with the majority of cases leading to
mortality due to metastasis. A requirement of the metastatic
process is for cells to acquire invasive capability (12). Therefore, it is a priority to
elucidate the mechanism of metastasis in MM. In the present study,
the mechanism of metastasis in MM was investigated. It was
demonstrated that SB431542 significantly inhibited proliferation
and cell invasion in RPMI 8226 cells. Additionally, it was
demonstrated that treating RPMI 8226 cells with SB431542
significantly decreased TGFβ1 expression, Smad2 phosphorylation and
MMP3 expression. Furthermore, overexpression of TGFβ1 resulted in
increased expression of p-Smad2 and MMP3, which could be partly
inhibited by SB431542. Taken together, the results demonstrated
that SB431542 reduced cell invasion in RPMI 8226 cells via the
TGFβ1/Smad2/MMP3 signaling pathway.
SB431542 is a small molecular inhibitor of TGFβ1
receptor, and its role in inducing anti-tumor immunity has been
previously reported (13). Matsuyama
et al (14) demonstrated that
SB431542 exerts an anti-tumor effect primarily by inhibiting the
proliferation of human osteosarcoma. Likewise, the present study
demonstrated that the proliferation of MM cells was inhibited by
SB431542 in a dose- and time-dependent manner. A number of studies
have reported that TGFβ1 has growth-stimulating effects on several
cell lines, including hepatic stellate cells (15), skin fibroblast (16) and bone marrow-derived mesenchymal stem
cells (17). In the present study,
TGFβ1 expression was inhibited by SB431542 in RPMI 8226 cells,
leading to an increase in cell proliferation.
The role of TGFβ1 in promoting tumor metastasis has
also been reported in a number of studies (18–20).
Likewise, the results of the present study also demonstrated the
role of TGFβ1 in promoting tumor metastasis in RPMI 8226 cells.
When the expression of TGFβ1 was inhibited by SB431542, the number
of migrating cells decreased; however, the reverse effect was
observed following overexpression of TGFβ1. The mechanism of TGFβ1
in regulating cell invasion was also investigated, and this present
study demonstrated that MMP3 may be involved and act as a
downstream target of TGFβ1/Smad2 signaling.
MMP3 is a member of the MMP family and is a type of
proteolytic enzyme involved in the degradation of structural
components of the ECM (21), and in
the release of precursor forms of growth factors in order to
degrade cell-cell and cell-ECM adhesion structure (22). This mechanism contributes to tumor
invasion and metastasis. In the present study it was observed that
SB431542 was able to significantly decrease MMP3 expression,
indicating that SB432542 is able to inhibit cell invasion in RPMI
8226 cells. The role of MMPs in promoting cell invasion and tumor
metastasis has been previously reported (23–25), and
is associated with EMT. It has also been reported that MMP3 is a
target and regulator of EMT (26,27).
Furthermore, in the present study, it was demonstrated that
overexpression of TGFβ1 increased MMP3 expression, which may be
partly inhibited by SB431542. Therefore, it was concluded that MMP3
expression was regulated by TGFβ1, and that MMP3 is downstream of
TGFβ1/Smad2 signaling.
In conclusion, overcoming cell invasion and tumor
metastasis is a clinical challenge that requires to be addressed
and SB431542 may be a potential and valuable drug.
Acknowledgements
The present study was supported by the Medical
research project of the Health Bureau of Chongqing (Chongqing,
China; grant no. 2012-2-390).
References
|
1
|
Solomon A, Frangione B and Franklin EC:
Bence Jones proteins and light chains of immunoglobulins.
Preferential association of the V lambda VI subgroup of human light
chains with amyloidosis AL (lambda). J Clin Invest. 70:453–460.
1982. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kumar SK, Dispenzieri A, Lacy MQ, Gertz
MA, Buadi FK, Pandey S, Kapoor P, Dingli D, Hayman SR, Leung N, et
al: Continued improvement in survival in multiple myeloma: Changes
in early mortality and outcomes in older patients. Leukemia.
28:1122–1128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gupta GP and Massagué J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Aggarwal R, Ghobrial IM and Roodman GD:
Chemokines in multiple myeloma. Exp Hematol. 34:1289–1295. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jurczyszyn A, Czepiel J, Biesiada G,
Gdula-Argasińska J, Cibor D, Owczarek D, Perucki W and Skotnicki
AB: HGF, sIL-6R and TGF-β1 play a significant role in the
progression of multiple myeloma. J Cancer. 5:518–524. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Toh B, Wang X, Keeble J, Sim WJ, Khoo K,
Wong WC, Kato M, Prevost-Blondel A, Thiery JP and Abastado JP:
Mesenchymal transition and dissemination of cancer cells is driven
by myeloid-derived suppressor cells infiltrating the primary tumor.
PLoS Biol. 9:e10011622011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ungefroren H, Groth S, Sebens S, Lehnert
H, Gieseler F and Fändrich F: Differential roles of Smad2 and Smad3
in the regulation of TGF-β1-mediated growth inhibition and cell
migration in pancreatic ductal adenocarcinoma cells: Control by
Rac1. Mol Cancer. 10:672011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: Regulators of the tumor microenvironment. Cell.
141:52–67. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Deryugina EI and Quigley JP: Matrix
metalloproteinases and tumor metastasis. Cancer Metastasis Rev.
25:9–34. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim HS, Shang T, Chen Z, Pflugfelder SC
and Li DQ: TGF-beta1 stimulates production of gelatinase (MMP-9),
collagenases (MMP-1, -13) and stromelysins (MMP-3, -10, -11) by
human corneal epithelial cells. Exp Eye Res. 79:263–274. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hideshima T, Richardson P, Chauhan D,
Palombella VJ, Elliott PJ, Adams J and Anderson KC: The proteasome
inhibitor PS-341 inhibits growth, induces apoptosis, and overcomes
drug resistance in human multiple myeloma cells. Cancer Res.
61:3071–3076. 2001.PubMed/NCBI
|
|
12
|
Yilmaz M and Christofori G: EMT, the
cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev.
28:15–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tanaka H, Shinto O, Yashiro M, Yamazoe S,
Iwauchi T, Muguruma K, Kubo N, Ohira M and Hirakawa K: Transforming
growth factor β signaling inhibitor, SB-431542, induces maturation
of dendritic cells and enhances anti-tumor activity. Oncol Rep.
24:1637–1643. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Matsuyama S, Iwadate M, Kondo M, Saitoh M,
Hanyu A, Shimizu K, Aburatani H, Mishima HK, Imamura T, Miyazono K
and Miyazawa K: SB-431542 and Gleevec inhibit transforming growth
factor-beta-induced proliferation of human osteosarcoma cells.
Cancer Res. 63:7791–7798. 2003.PubMed/NCBI
|
|
15
|
He Y, Huang C, Sun X, Long XR, Lv XW and
Li J: MicroRNA-146a modulates TGF-beta1-induced hepatic stellate
cell proliferation by targeting SMAD4. Cell Signal. 24:1923–1930.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu X, Li P, Liu P, Xiong R, Zhang E, Chen
X, Gu D, Zhao Y, Wang Z and Zhou Y: The essential role for c-Ski in
mediating TGF-beta1-induced bi-directional effects on skin
fibroblast proliferation through a feedback loop. Biochem J.
409:289–297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bergfeld SA and DeClerck YA: Bone
marrow-derived mesenchymal stem cells and the tumor
microenvironment. Cancer Metastasis Rev. 29:249–261. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sawada T, Kimura K, Nishihara T, Onoda N,
Teraoka H, Yamashita Y, Yamada N, Yashiro M, Ohira M and Hirakawa
K: TGF-beta1 down-regulates ICAM-1 expression and enhances liver
metastasis of pancreatic cancer. Adv Med Sci. 51:60–65.
2006.PubMed/NCBI
|
|
19
|
Biswas S, Guix M, Rinehart C, Dugger TC,
Chytil A, Moses HL, Freeman ML and Arteaga CL: Inhibition of TGF-β
with neutralizing antibodies prevents radiation-induced
acceleration of metastatic cancer progression. J Clin Invest.
127:11162017. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bacman D, Merkel S, Croner R, Papadopoulos
T, Brueckl W and Dimmler A: TGF-beta receptor 2 downregulation in
tumour-associated stroma worsens prognosis and high-grade tumours
show more tumour-associated macrophages and lower TGF-beta1
expression in colon carcinoma: A retrospective study. BMC Cancer.
7:1562007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Radisky DC, Levy DD, Littlepage LE, Liu H,
Nelson CM, Fata JE, Leake D, Godden EL, Albertson DG, Nieto MA, et
al: Rac1b and reactive oxygen species mediate MMP-3-induced EMT and
genomic instability. Nature. 436:123–127. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Casey TM, Eneman J, Crocker A, White J,
Tessitore J, Stanley M, Harlow S, Bunn JY, Weaver D, Muss H and
Plaut K: Cancer associated fibroblasts stimulated by transforming
growth factor beta1 (TGF-beta 1) increase invasion rate of tumor
cells: A population study. Breast Cancer Res Treat. 110:39–49.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fuxe J, Vincent T and de Herreros A
Garcia: Transcriptional crosstalk between TGF-β and stem cell
pathways in tumor cell invasion: Role of EMT promoting Smad
complexes. Cell Cycle. 9:2363–2374. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen S, Zhu J, Zuo S, Ma J, Zhang J, Chen
G, Wang X, Pan Y, Liu Y and Wang P: 1,25(OH)2D3 attenuates
TGF-β1/β2-induced increased migration and invasion via inhibiting
epithelial-mesenchymal transition in colon cancer cells. Biochem
Biophys Res Commun. 468:130–135. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Blavier L, Lazaryev A, Shi XH, Dorey FJ,
Shackleford GM and DeClerck YA: Stromelysin-1 (MMP-3) is a target
and a regulator of Wnt1-induced epithelial-mesenchymal transition
(EMT). Cancer Biol Ther. 10:198–208. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Robichaud N, Del Rincon SV, Huor B, Alain
T, Petruccelli LA, Hearnden J, Goncalves C, Grotegut S, Spruck CH,
Furic L, et al: Phosphorylation of eIF4E promotes EMT and
metastasis via translational control of SNAIL and MMP-3. Oncogene.
34:2032–2042. 2015. View Article : Google Scholar : PubMed/NCBI
|