Introduction
Ovarian cancer, together with endometrial cancer and
cervical carcinoma, are the three most common gynecological
malignant cancers in the female genital system (1). Ovarian cancer ranks the fifth of all
female malignancies, causing 14,270 mortalities in the USA per year
(1,2).
Management of ovarian cancer generally consists of surgery followed
by chemotherapy (3–6). Although surgery and platinum-based
chemotherapy currently provide the cornerstone of standard ovarian
cancer management pathways, novel therapy strategies are urgently
required to improve outcomes (5,6). To
develop effective ovarian cancer treatments, elucidation of the
molecular pathogenesis of ovarian cancer is required.
MicroRNAs (miRNAs or miRs) are a class of small RNAs
of ~18–22 nucleotides in length (7).
miRNAs suppress protein expression by inhibiting messenger RNA
(mRNA) translation or inducing mRNA degradation by binding to the
3′-untranslated region (3′-UTR) of their target mRNAs (7). Deregulation or dysfunction of miRNAs
contribute to cancer development (7–12). A
number of miRNAs have been identified as highly up- or
downregulated in ovarian cancer (13–19).
However, their roles in the pathogenesis of ovarian cancer remain
unclear, and whether other miRNAs may be involved in the
pathogenesis of ovarian cancer remains unknown.
Previous studies demonstrated that miR-520b was
downregulated in breast cancer cells and contributed to the
migration of highly metastatic breast cancer cells via a network
that involves two target genes, hepatitis B virus X-interacting
protein (HBXIP) and interleukin (IL)-8, in which HBXIP promotes
cell migration by upregulation of IL-8, which is mediated by
nuclear factor-κB (20).
In the present study, the role of miR-520b in
ovarian cancer was investigated. The data indicated that miR-520b
promoted cellular proliferation, and suggested that the targeted
gene of miR-520b may be ring finger protein 216 (RNF216). The
present results may provide a novel therapeutic target for ovarian
cancer.
Materials and methods
Cell culture and transfection
The human ovarian cancer cell lines SKOV3, Hey and
OVCAR3 as well as the human embryonic kidney (HEK) 293 cell line
were purchased from the Cell Bank of the Chinese Academy of
Sciences (Shanghai, China). The cell lines were maintained in RPMI
1640 medium (Invitrogen; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) in the presence of 10% heat-inactivated fetal bovine serum
(Biological Industries, Beit Haemek, Israel), 100 IU/ml penicillin
and 100 ng/ml streptomycin in a humidified 5% (v/v) atmosphere of
CO2 at 37°C. Transfection was performed using
Lipofectamine 2000 reagent (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) according to the manufacturer's
protocol.
Patients
Surgical specimens from 30 ovarian cancer patients
and matched tumor adjacent normal tissues were obtained
postoperatively from April to June, 2008 from the Department of
Obstetrics and Gynecology of Changhai Hospital, Second Military
Medical University (Shanghai, China). All diagnoses were based on
pathological and/or cytological evidence. Tissues were obtained
prior to chemotherapy and radiotherapy, and were immediately frozen
and stored at −80°C prior to being subjected to reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis. Informed consent was obtained from each patient, and the
study was approved by the Ethics Committee of the Second Military
Medical University.
RT-qPCR
RT-qPCR analysis was performed on a 7500 Real-Time
PCR System (Applied Biosystems; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). miR-520b expression was assessed using a
mirVanaTM qRT-PCR miRNA Detection kit (Ambion; Thermo Fisher
Scientific, Inc., Austin, TX, USA). The primers were designed and
synthesized by Shanghai Shenggong Biology Engineering Technology
Service, Ltd. (Shanghai, China). U6 was used as an internal
control. The relative messenger RNA (mRNA) levels of RNF216 were
normalized to the mRNA levels of the housekeeping gene
glyceraldehyde 3-phosphate dehydrogenase (GAPDH), and calculated by
the 2−ΔΔCq method (21).
The primers used were as follows: GAPDH forward,
5′-CCATGTTCGTCATGGG-TGT GAA CCA-3′ and reverse,
5′-GCCAGTAGAGGCAGGGATGATGTTG-3′; RNF216 forward,
5′-TCTTCCTGTCCTTAGTCGGGAG-3′ and reverse,
5′-CTGGGCAGCTGGTTTGATGA-3′; and U6 forward,
5′-GTGGACCGCACAAGCTCGCT-3′ and reverse,
5′-TTGTTGAACGGCACTGTGTATAGCA-3′. A 2-step processwas used for
denaturation, annealing and extension. i) The reaction temperature
was increased to 95°C for 10 sec to melt all dsDNA. ii) The
temperature was lowered to 60°C for 30 sec to promote primer
binding to the template and subsequent elongation occurred due to
sufficient activity of the DNA polymerase at this temperature.
Steps 1–2 were repeated, for 40 cycles for GAPDH and RNF216, and
for 45 cycles for miR-520b.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
For MTT assay, 5×103 cells/well were seeded in
triplicate in a 96-well plate with complete growth medium. Cells
were counted over 5 days using an MTT assay kit (Promega
Corporation, Madison, WI, USA), as described previously (10,22–25). The
data were measured with a microtiter plate reader with 570-nm
filters (Promega Corporation).
miRNAs antisense, miRNAs mimics,
oligonucleotides and overexpression plasmids
miRNAs mimics (miR-520b mimics) and miRNAs antisense
oligonucleotides (miR-520b ASO) were obtained from Shanghai
GenePharma Co., Ltd. (Shanghai, China). miRNAs ASO, miRNAs mimics
and negative control (NC) were transfected into cells at a
concentration of 50 nM using Lipofectamine 2000 transfection
reagent (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) according to the manufacturer's protocol. At 48 or 72 h later,
cells were collected for further experiments. The overexpression
plasmid pcDNA3.1-RNF216 and the knockdown plasmid pcDNA3.1-RNF216
KD were constructed and confirmed by Shanghai Shenggong Biology
Engineering Technology Service, Ltd.
miRNAs targets prediction
TargetScanHuman (http://www.targetscan.org/vert_61/) (26–29) was
applied to identify the potential target of miR-520b.
RNF216 3′-UTR reporter analysis
The RNF216 3′-UTR reporter plasmids (pRL-RNF216)
were constructed by Shanghai Shenggong Biology Engineering
Technology Service, Ltd. Mutations in the miR-520b target regions
of the RNF216 3′-UTR were generated using QuikChange Multi
Site-Directed Mutagenesis kit (Stratagene; Agilent Technologies,
Inc., Santa Clara, CA, USA). Renilla luciferase reporter
plasmids (3.6 fmol) and pGL3-control (500 ng used for
normalization; Promega Corporation) were transfected with
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) into HEK293 (6×104 cells/well). Cells were
collected after 48 h for analysis using the
Dual-Luciferase® Reporter Assay System (Promega
Corporation) (30). The sequence of
the primers used for cloning RNF216 3′UTR were as follows: Forward,
5′-CGACGCGTGGAGGGCCAGATGTGCCCATC-3′; and reverse,
5′-GGGTTTAAACATGTTTAAAATTCTGATGTCATTTATTGG-3′.
Immunohistochemistry (IHC)
IHC staining was performed as described previously
(31). Briefly, 4-µm thick sections
were cut, and an anti-RNF216 antibody (cat. no. HPA018955; 1:1,000;
Sigma-Aldrich, St. Louis, MO, USA) was applied. Subsequent
counterstaining was performed with hematoxylin. The immunostaining
results for RNF216 were evaluated using a semi-quantitative scoring
system as described previously (32),
which calculated the staining intensity and the percentage of
positive cells. IHC staining was scored according to the following
criteria: -, 0–10% of the nucleated cells were stained; +, 10–40%
of the nucleated cells were stained; ++; 40–70% of the nucleated
cells were stained; and +++, 70–100% of the nucleated cells were
stained. RNF216 expression was considered to be observed when the
score was ≥+. Alternatively, the IHC score of RNF216 expression was
- to + and ++ to +++, which represented low and high expression,
respectively.
Statistical analysis
Data were presented as the mean ± standard deviation
from ≥3 independent experiments. The differences between groups
were analyzed using two-tailed Student's t test when only
two groups were compared. The differences between groups were
analyzed using analysis of variance when ≥3 groups were compared.
Correlation analysis was performed by two-tailed Person's
correlation coefficient analysis. Patients survival was determined
by Kaplan-Meier analysis. Statistical analyses were performed using
SPSS software version 17.0 (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
High expression of miR-520b in ovarian
cancer
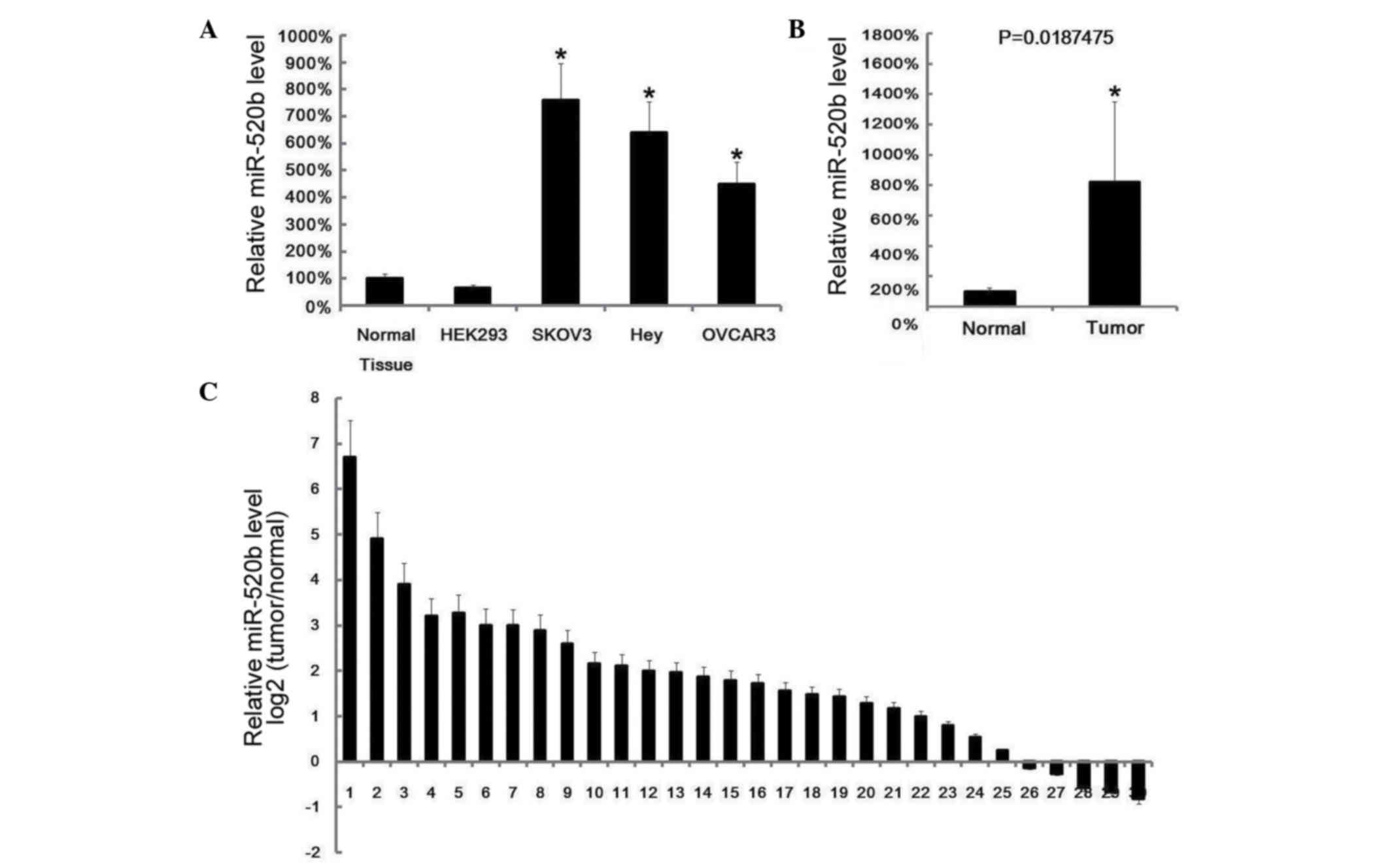
miR-520b expression was examined in human ovarian
cancer cell lines (SKOV3, Hey and OVCAR3) and ovarian cancer
tissues by RT-qPCR. It was observed that miR-520b exhibited high
expression levels in ovarian cancer cell lines and ovarian cancer
tissues (Fig. 1A and B). In total, 30
pairs of ovarian cancer tissues and matched adjacent normal tissues
were collected from 30 ovarian cancer patients, and it was observed
that, in 25 pairs, miR-520b expression was higher than in matched
normal tissues (Fig. 1C).
Downregulation of miR-520b inhibits
cells proliferation, while upregulation of miR-520b promotes cells
proliferation
To investigate the role of miR-520b in ovarian
cancer, miR-520b expression was suppressed in SKOV3 and Hey cells
by ASO transfection. At 48 h after transfection, the suppressed
miR-520b levels in SKOV3 and Hey cells were confirmed by RT-qPCR
(Fig. 2A). Next, cell proliferation
was assayed by MTT analysis, and it was observed that
downregulation of miR-520b inhibited cell proliferation (Fig. 2B). Subsequently, miR-520b was
overexpressed by transfection of miR-520b mimics into HEK293 cells
(Fig. 2C). Similarly, 48 h later,
cell proliferation was evaluated by MTT analysis, and it was
observed that overexpression of miR-520b promoted cell growth
(Fig. 2D).
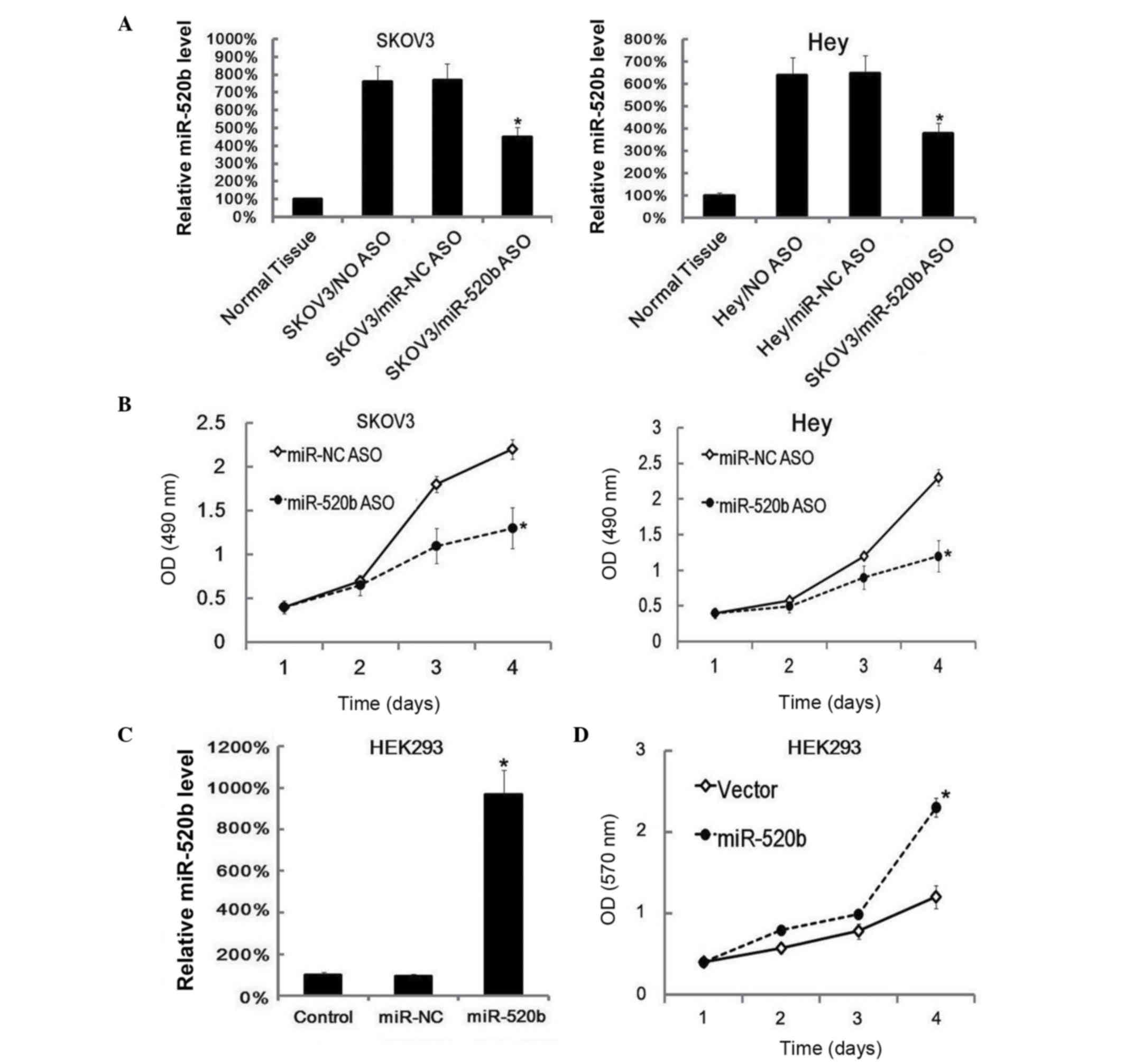 | Figure 2.miR-520b downregulation reduced cell
proliferation, while miR-520b upregulation promoted cell growth.
(A) SKOV3 and Hey cells (6×105 cells/well) were
transfected with miR-520b ASO or miR-NC ASO separately, and 48 h
later, the expression of miR-520b was assayed by RT-qPCR. Data were
normalized to U6 small nuclear RNA. The miR-520b expression level
in ovarian cancer control samples was arbitrarily defined as 100%.
Data are represented as the mean ± SD of three separate
experiments. (B) Following miR-520b ASO transfection, cell
proliferation was assayed with MTT at the indicted times. Data are
represented as the mean ± SD of three separate experiments. (C)
HEK293 cells (6×105 cells/well) were transfected with
miR-520b mimics, and 48 h later, the expression of miR-520b was
assessed by RT-qPCR. Data were normalized to U6 small nuclear RNA.
The miR-520b expression level in ovarian cancer control samples was
arbitrarily defined as 100%. Data are represented as the mean ± SD
of three separate experiments. (D) At 24 h after miR-520b mimics
transfection, cell proliferation at different times was assayed
with MTT. Data are represented as the mean ± SD of three separate
experiments. *P<0.05 vs. control. miR, microRNA; NC, negative
control; ASO, antisense oligonucleotide; NO, untreated with ASO or
non-specific oligonucleotide; OD, optical density; HEK, human
embryonic kidney; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; SD, standard deviation; MTT,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide. |
RNF216 is targeted by miR-520b
The potential target genes of miR-520b were explored
using TargetScanHuman software, which predicted ~884 genes (data
not shown). Of these, the presented study focused on an unreported
gene, RNF216. The binding wild type (WT) and mutant sites in RNF216
are indicated in Fig. 3A. The WT
3′-UTR of RNF216 and its mutated version were cloned into
luciferase reporter plasmids. miR-520b and the reporter plasmids
were co-transfected into HEK293 cells. The results revealed that,
in WT 3′-UTR, miR-520b reduced the luciferase activity, while in
the mutated version, the difference between miR-NC and miR-520b was
not significant (P>0.05; Fig.
3B).
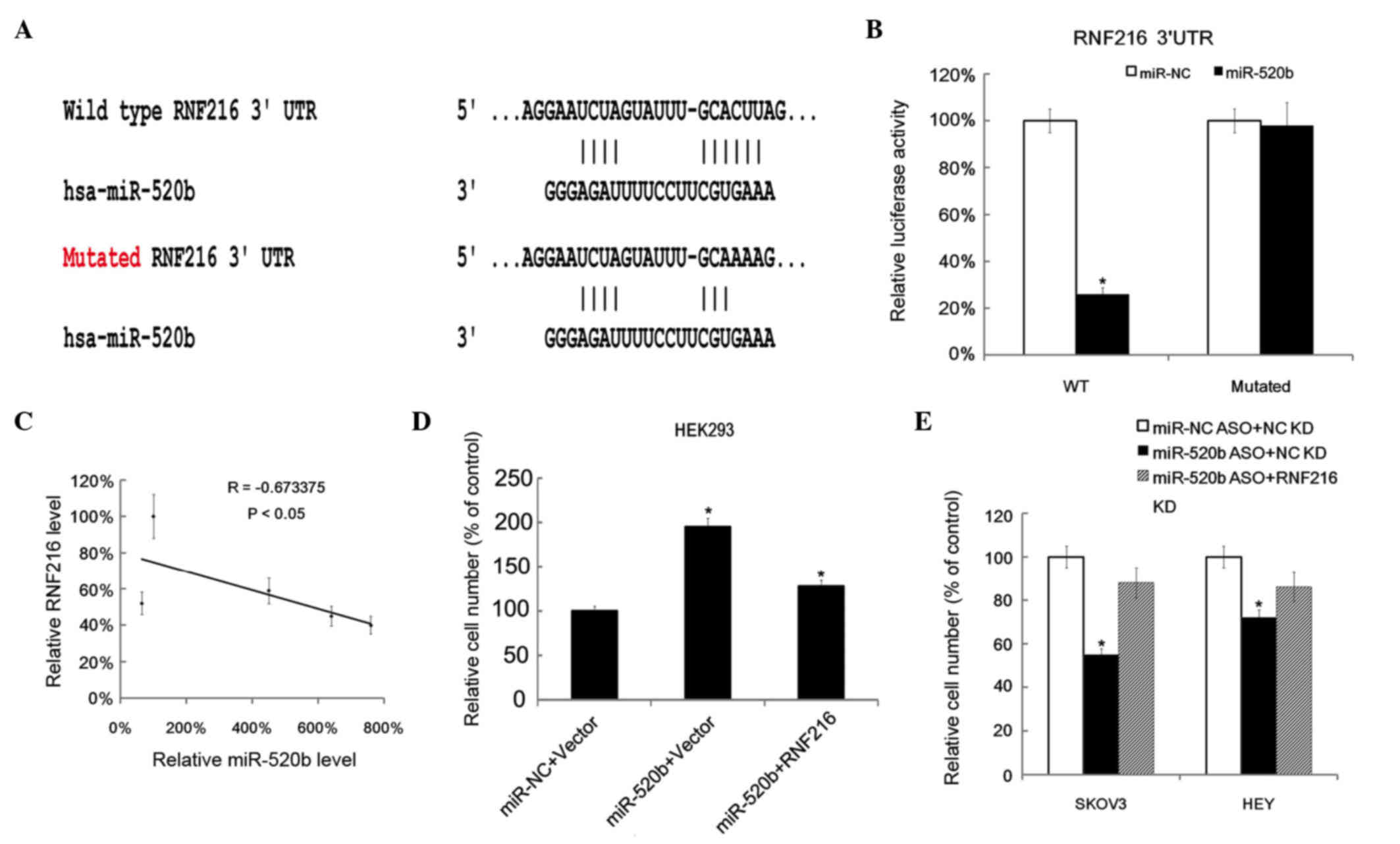 | Figure 3.miR-520b-targeted genes prediction and
validation. (A) Putative genes targeted by miR-520b were predicted
by TargetScanHuman. The binding site of the putative targeted gene
RNF216 and the mutated site of RNF216 were shown. (B) The RL
reporter plasmids (RL-control, RL-RNF216 and RL-mutated RNF216) as
well as miR-520b or miR-NC were co-transfected into HEK293 cells,
along with a firefly luciferase reporter (pGL control) for
normalization. Luciferase activities were measured after 48 h, and
the ratio of RL activity vs. firefly luciferase activity in the
miR-520b-treated group was calculated and compared with that in the
in miR-NC group (which was arbitrary defined as 100%). Data are
represented as the mean ± SD of three separate experiments. (C)
Inverse correlation between miR-520b and RNF216 levels in ovarian
cancer tissues (n=5). Statistical analysis was performed using
Pearson's correlation coefficient. (D) HEK293 cells
(6×105 cells/well) were transfected with an RNF216
overexpression plasmid (pcDNA3.1-RNF216), and 12 h later, the cells
were transfected with miR-520b mimics. After 12 h, cell
proliferation was evaluated by MTT assay. The relative cell number
in the miR-NC + vector (which served as NC) group was defined as
100%. (E) SKOV3 and Hey cells (6×105 cells/well) were
transfected with pcDNA3.1-RNF216 knockdown plasmid, followed by
transfection with miR-520b ASO. Cell proliferation was evaluated by
MTT assay 24 h later. The relative cell number in the miR-NC ASO +
empty vector (which served as NC) group was defined as 100%.
*P<0.05 vs. control. RNF216, ring finger protein 216; UTR,
untranslated region; hsa, Homo sapiens; miR, microRNA; NC,
negative control; ASO, antisense oligonucleotide; WT, wild type;
HEK, human embryonic kidney; KD, knowckdown; RL, Renilla
luciferase; MTT,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide. |
Then, the correlation between miR-520b level and
RNF216 in ovarian cancer tissues was explored. Pearson's
correlation coefficient analysis revealed that RNF216 was inversely
correlated with miR-520b level in ovarian cancer tissues (Fig. 3C). Next, miR-520b mimics and RNF216
overexpression plasmid were co-transfected into HEK293 cells.
RNF216 overexpression partly reduced the cell proliferation effect
of miR-520b (Fig. 3D). As expected,
RNF216 downregulation partly reduced the growth inhibitory effect
of miR-520b ASO in SKOV3 and Hey cells (Fig. 3E).
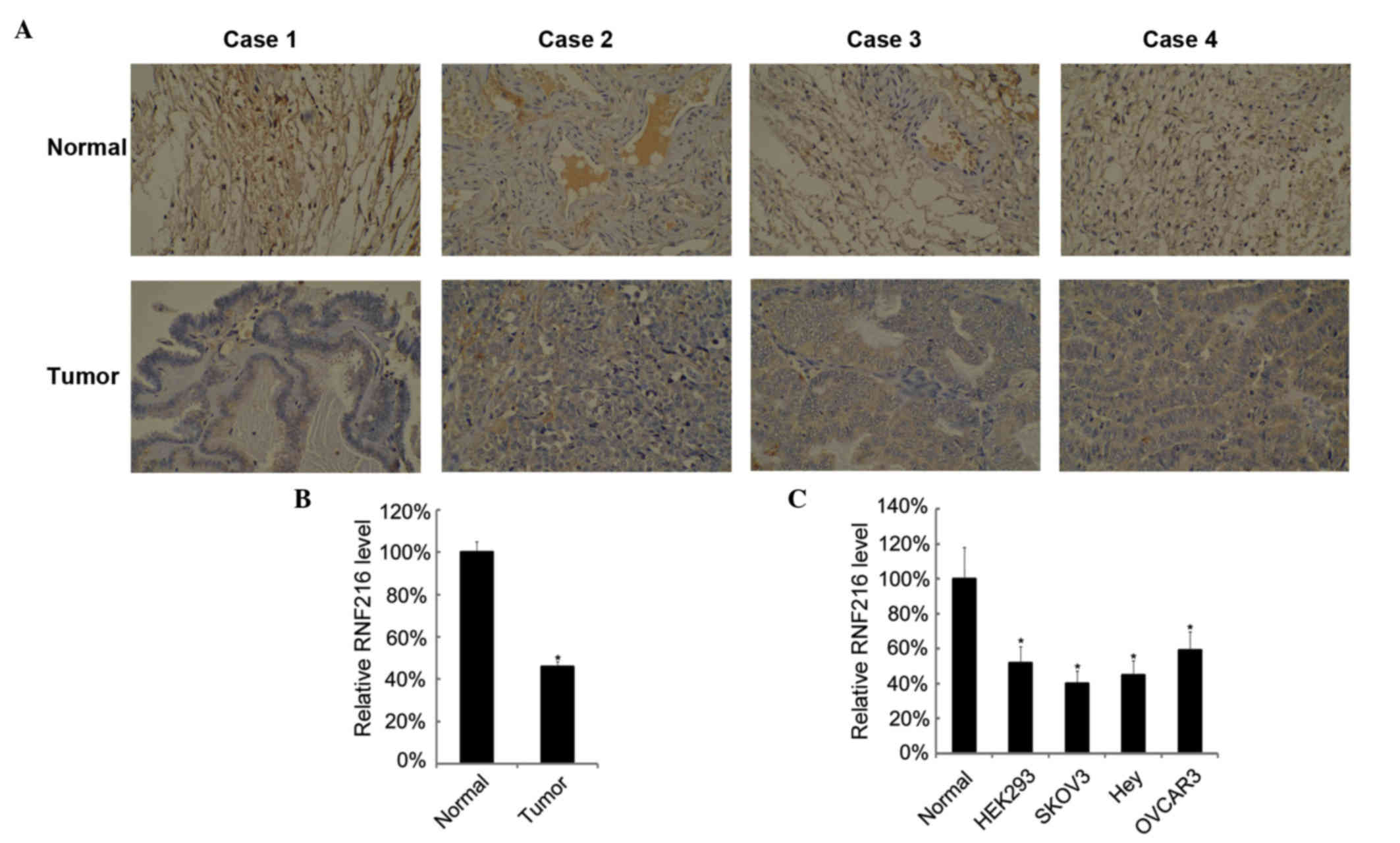
Low expression of RNF216 in ovarian
cancer
Next, the mRNA and protein levels of RNF216 in
ovarian cancer tissues and cell lines were examined. There were
lower levels of RNF216 in ovarian cancer tissues than in normal
ovarian tissues. Four representative cases are shown in Fig. 4A. The mean levels of RNF216 in ovarian
cancer tissues were lower than those in matched tumor adjacent
normal tissues (Fig. 4B). RNF216
expression in ovarian cancer cell lines was also assessed, and
RNF216 displayed lower expression levels in ovarian cancer cell
lines (Fig. 4C).
Discussion
In the present study, high expression of miR-520b
and low expression of RNF216 was detected in ovarian cancer
tissues, and RNF216 was observed to be inversely correlated with
miR-520b level. In cell experiments, the miR-520b level was
regulated by miRNAs mimics or ASO transfection, and it was
demonstrated that downregulation of miR-520b inhibited cell
proliferation, whereas upregulation of miR-520b promoted cell
proliferation. RNF216 3′-UTR reporter analysis revealed that
miR-520b could target RNF216 in vitro. These data led to the
conclusion that high expression of miR-520b in ovarian cancer
promoted cell growth via RNF216. To the best of our knowledge, the
present study is the first to demonstrate that miR-520b targets
RNF216 in ovarian cancer.
The present data indicated that the expression of
miR-520b was higher in ovarian cancer tissues than that in normal
control tissues. However, a previous study reported that miR-520b
was downregulated in breast cancer cells, which contributed to the
migration of these cells (20). In
hepatocellular carcinoma, miR-520b was downregulated in tumor
tissues and hepatoma cell lines, and ectopic expression of miR-520b
inhibited the growth of hepatoma cells in vitro and in
vivo (33). This discrepancy may
be explained by the genes targeted by miR-520b. In breast cancer
cells, miR-520b targeted HBXIP and IL-8, while in hepatocellular
carcinoma, miR-520b targeted mitogen-activated protein kinase
kinase kinase 2 (MEKK2) and cyclin D1 (33). The present study demonstrated that
miR-520b targeted RNF216. Whether miR-520b targeted HBXIP, IL-8,
MEKK2 and cyclin D1 in ovarian cancer requires further
investigation.
The bioinformatics algorithm used in the present
study predicted 884 genes potentially targeted by miR-520b,
including RNF216. There were two reasons for selecting RNF216 for
further investigation. One is that RNF216 has not been reported yet
in ovarian cancer. The other is that data from the preliminary test
(microarray) revealed low RNF216 expression in ovarian cancer.
In conclusion, the present data demonstrated the
role of miR-520b in ovarian cancer, and suggested that high
expression of miR-520b in ovarian cancer promoted cell growth via
RNF216. The present report is a preliminary study to outline the
function of miR-520b. Despite certain discrepancy with the
literature, the findings from the current study may provide the
basis for future cancer studies.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhao YN, Chen GS and Hong SJ: Circulating
MicroRNAs in gynecological malignancies: From detection to
prediction. Exp Hematol Oncol. 3:142014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rooth C: Ovarian cancer: Risk factors,
treatment and management. Br J Nurs. 22:S23–S30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bristow RE, Tomacruz RS, Armstrong DK,
Trimble EL and Montz FJ: Survival effect of maximal cytoreductive
surgery for advanced ovarian carcinoma during the platinum era: A
meta-analysis. J Clin Oncol. 20:1248–1259. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Winter-Roach BA, Kitchener HC and Lawrie
TA: Adjuvant (post-surgery) chemotherapy for early stage epithelial
ovarian cancer. Cochrane Database Syst Rev. 3:CD0047062012.
|
|
6
|
Winter-Roach BA, Kitchener HC and
Dickinson HO: Adjuvant (post-surgery) chemotherapy for early stage
epithelial ovarian cancer. Cochrane Database Syst Rev: CD004706.
2009. View Article : Google Scholar
|
|
7
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hou J, Lin L, Zhou W, Wang Z, Ding G, Dong
Q, Qin L, Wu X, Zheng Y, Yang Y, et al: Identification of miRNomes
in human liver and hepatocellular carcinoma reveals miR-199a/b-3p
as therapeutic target for hepatocellular carcinoma. Cancer Cell.
19:232–243. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li D, Liu X, Lin L, Hou J, Li N, Wang C,
Wang P, Zhang Q, Zhang P, Zhou W, et al: MicroRNA-99a inhibits
hepatocellular carcinoma growth and correlates with prognosis of
patients with hepatocellular carcinoma. J Biol Chem.
286:36677–36685. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mendell JT and Olson EN: MicroRNAs in
stress signaling and human disease. Cell. 148:1172–1187. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Iorio MV, Visone R, Di Leva G, Donati V,
Petrocca F, Casalini P, Taccioli C, Volinia S, Liu CG, Alder H, et
al: MicroRNA signatures in human ovarian cancer. Cancer Res.
67:8699–8707. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Resnick KE, Alder H, Hagan JP, Richardson
DL, Croce CM and Cohn DE: The detection of differentially expressed
microRNAs from the serum of ovarian cancer patients using a novel
real-time PCR platform. Gynecol Oncol. 112:55–59. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chung YW, Bae HS, Song JY, Lee JK, Lee NW,
Kim T and Lee KW: Detection of microRNA as novel biomarkers of
epithelial ovarian cancer from the serum of ovarian cancer
patients. Int J Gynecol Cancer. 23:673–679. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shapira I, Oswald M, Lovecchio J, Khalili
H, Menzin A, Whyte J, Dos Santos L, Liang S, Bhuiya T, Keogh M, et
al: Circulating biomarkers for detection of ovarian cancer and
predicting cancer outcomes. Br J Cancer. 110:976–983. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Suryawanshi S, Vlad AM, Lin HM,
Mantia-Smaldone G, Laskey R, Lee M, Lin Y, Donnellan N, Klein-Patel
M, Lee T, et al: Plasma microRNAs as novel biomarkers for
endometriosis and endometriosis-associated ovarian cancer. Clin
Cancer Res. 19:1213–1224. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zheng H, Zhang L, Zhao Y, Yang D, Song F,
Wen Y, Hao Q, Hu Z, Zhang W and Chen K: Plasma miRNAs as diagnostic
and prognostic biomarkers for ovarian cancer. PLoS One.
8:e778532013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kan CW, Hahn MA, Gard GB, Maidens J, Huh
JY, Marsh DJ and Howell VM: Elevated levels of circulating
microRNA-200 family members correlate with serous epithelial
ovarian cancer. BMC Cancer. 12:6272012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu N, Zhang J, Cui W, Kong G, Zhang S, Yue
L, Bai X, Zhang Z, Zhang W, Zhang X and Ye L: miR-520b regulates
migration of breast cancer cells by targeting hepatitis B
X-interacting protein and interleukin-8. J Biol Chem.
286:13714–13722. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
van Meerloo J, Kaspers GJ and Cloos J:
Cell sensitivity assays: The MTT assay. Methods Mol Biol.
731:237–245. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Han ZB, Yang Z, Chi Y, Zhang L, Wang Y, Ji
Y, Wang J, Zhao H and Han ZC: MicroRNA-124 suppresses breast cancer
cell growth and motility by targeting CD151. Cell Physiol Biochem.
31:823–832. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song B, Zhang C, Li G, Jin G and Liu C:
MiR-940 inhibited pancreatic ductal adenocarcinoma growth by
targeting MyD88. Cell Physiol Biochem. 35:1167–1177. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu N, Zhang C, Bai C, Han YP and Li Q:
MiR-4782-3p inhibited non-small cell lung cancer growth via USP14.
Cell Physiol Biochem. 33:457–467. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Grimson A, Farh KK, Johnston WK,
Garrett-Engele P, Lim LP and Bartel DP: MicroRNA targeting
specificity in mammals: Determinants beyond seed pairing. Mol Cell.
27:91–105. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Garcia DM, Baek D, Shin C, Bell GW,
Grimson A and Bartel DP: Weak seed-pairing stability and high
target-site abundance decrease the proficiency of lsy-6 and other
microRNAs. Nat Struct Mol Biol. 18:1139–1146. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Grentzmann G, Ingram JA, Kelly PJ,
Gesteland RF and Atkins JF: A dual-luciferase reporter system for
studying recoding signals. RNA. 4:479–486. 1998.PubMed/NCBI
|
|
31
|
Xu XS, Wang L, Abrams J and Wang G:
Histone deacetylases (HDACs) in XPC gene silencing and bladder
cancer. J Hematol Oncol. 4:172011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lv T, Yuan D, Miao X, Lv Y, Zhan P, Shen X
and Song Y: Over-expression of LSD1 promotes proliferation,
migration and invasion in non-small cell lung cancer. PLoS One.
7:e350652012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang W, Kong G, Zhang J, Wang T, Ye L and
Zhang X: MicroRNA-520b inhibits growth of hepatoma cells by
targeting MEKK2 and cyclin D1. PLoS One. 7:e314502012. View Article : Google Scholar : PubMed/NCBI
|