Introduction
The Hippo/Yes-associated protein (YAP) signaling
pathway is a highly-conserved signal transduction pathway, which
involves various oncogenes and tumor suppressor genes (1). Central to the signaling pathway is a
core kinase cascade chain, which consists of mammalian STE20-like
kinase 1/2 (MST1/2), human salvador 1 (hSAV1), large tumor
suppressor kinase 1/2 (LATS1/2) and mps 1 binder kinase
activator-like 1 (MOB1), and the downstream transcriptional
coactivator YAP. Additionally, cell cycle protein E (cyclin E) and
cellular inhibitor of apoptosis protein 1 (CIAP1) are regulatory
factors downstream of the signaling pathway (2,3).
Hippo/YAP signaling is a molecular switch for cell
division and apoptosis, and YAP is the core component of the
signaling pathway (4). Under
physiological conditions, YAP is phosphorylated by proteins
upstream of the Hippo/YAP signaling pathway via a series of kinase
cascades (5,6). Initially, MST1/2 binds to hSAV1, which
forms a complex and phosphorylates LATS1/2. YAP is subsequently
phosphorylated by LATS1/2, which is regulated by MOB1.
Phosphorylation of YAP subsequently promotes cytoplasmic
localization of the YAP [serine (ser) 127]/14-3-3 protein complex.
This in turn prevents YAP-mediated transcriptional activation in
the nucleus (5,6) (Fig.
1).
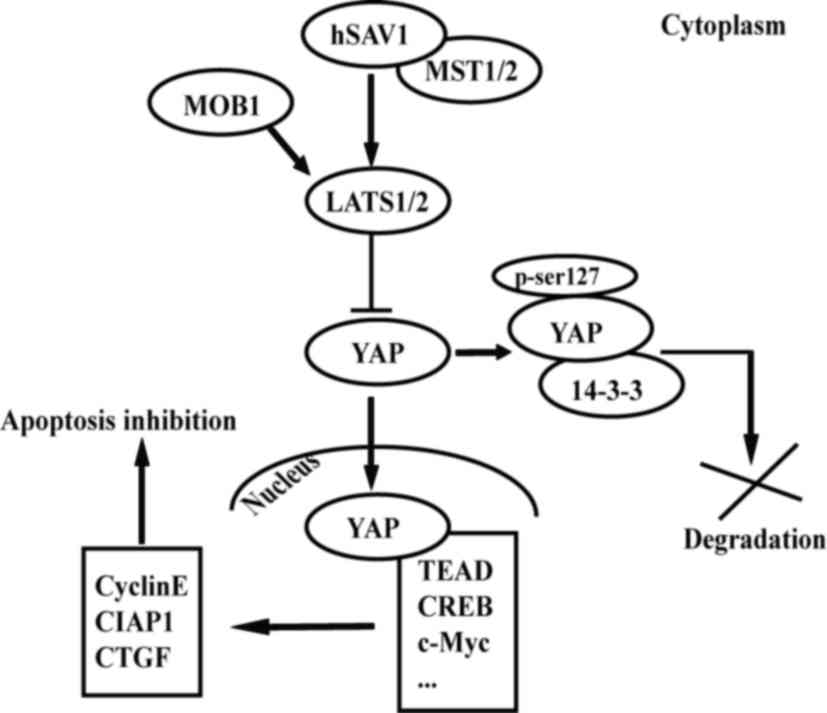 | Figure 1.Hippo/YAP signaling pathway maintains
the balance between cell proliferation and apoptosis. YAP is a key
factor downstream of the signaling pathway. Under physiological
conditions, YAP is phosphorylated through a series of kinase
cascades and is located in the cytoplasm. When MST1/2, hSAV1, MOB1
and LATS1/2 are inactivated or YAP is overexpressed, YAP is
translocated into the nucleus and binds to TEAD. This in turn
promotes the expression of a number of factors (cyclin E, CIAP1 and
CTGF). This also promotes cell proliferation and inhibits
apoptosis. CIAP1, cellular inhibitor of apoptosis protein 1; CREB,
cyclic adenosine monophosphate response element-binding protein;
CTGF, connective tissue growth factor; hSAV1, human salvador 1;
LATS1/2, large tumor suppressor kinase 1/2; MOB1, mps 1 binder
kinase activator-like 1; MST1/2, mammalian STE20-like kinase 1/2;
se, serine; TEAD, TEA domain family member; YAP, yes-associated
protein. |
Additionally, YAP has a WW domain that is able to
directly associate with PPxY motifs in LATS1/2 for signal
transduction. This suggests that Hippo/YAP signaling is also able
to inhibit YAP by non-phosphorylation (7). When Hippo/YAP signaling is aberrant,
overexpressed YAP enters into the nucleus and activates the
transcription factor TEA domain family member (TEAD), which in turn
stimulates the transcription of downstream factors cyclin E, CIAP1
and connective tissue growth factor (CTGF). Therefore, YAP is able
to promote proliferation and inhibit apoptosis (8). Notably, the WW domain of YAP is also
able to promote apoptosis following DNA damage via binding to p73,
which may be associated with the microenvironment or type of tumor
(9).
Hepatocellular carcinoma (HCC) is a common type of
malignant tumor and the third leading cause of cancer-associated
mortality worldwide (10). HCC is
induced by numerous factors and develops in multiple stages, which
hinders early detection. YAP is able to promote HCC cell
proliferation and transformation (a change in cell phenotype)
(11,12). Therefore, investigating the
association between YAP and HCC is important for identifying novel
tumor markers. YAP-associated membrane proteins, including cluster
of differentiation (CD)166 and melanoma cell adhesion molecule
(MCAM), have been reported to be involved in hepatocarcinogenesis
(13). Additionally, a number of
studies have demonstrated that the targeting of Hippo/YAP signaling
may be able to inhibit proliferation in HCC cells (14,15).
Studies have employed two main strategies: i) Targeting components
of the Hippo/YAP signaling pathway, including inhibiting the
activity of YAP or promoting the activity of MST1 or LATS1/2
(16–18); and ii) targeting pathways and factors
associated with the Hippo/YAP signaling pathway, including
Wnt/β-catenin, cyclic adenosine monophosphate response
element-binding protein (CREB) and tribbles homolog 2 (TRIB2)
(12,19,20).
YAP may also be used to determine the prognosis of
HCC. The observation that liver regeneration stimulates the growth
of residual cancer tissue following surgery has been demonstrated
by multiple clinical studies. However, the precise underlying
mechanisms remain unclear (21).
Furthermore, YAP is important for regulating liver size, and
YAP-associated HCC is linked with a poor short-term prognosis,
suggesting that YAP may be utilized as an independent prognostic
marker for HCC (22). In conclusion,
YAP exerts an important role in the development of HCC. Further
elucidation of the underlying molecular mechanisms of the Hippo/YAP
signaling pathway and the interaction between different signaling
pathways and molecules may provide novel methods for the diagnosis,
treatment and prognosis of HCC.
In the present review, the underlying molecular
mechanisms of Hippo/YAP signaling and YAP-associated proteins in
HCC pathogenesis were examined. In addition, the corresponding
advantages of YAP in diagnosis, treatment and determining the
prognosis of HCC were analyzed.
Overexpression of YAP promotes HCC
HCC is a highly malignant and aggressive cancer, and
YAP is overexpressed in 62% of patients with HCC (23). The overexpression of YAP has been
reported in rat and human HCC cells (24). Another study demonstrated that YAP is
present in the HCC cell nuclei during hepatocarcinogenesis
(25). Furthermore, the expression of
YAP has also been associated with the differentiation of HCC cells,
level of serum α-fetoprotein (AFP) and vascular infiltration
(26,27).
The activation of hepatic stellate cells (HSCs) has
been demonstrated to be critical for the development of cirrhosis
from hepatic fibrosis, and that it promotes the proliferation and
survival of HCC cells (28). It has
been reported that a complex loop of regulation between HSCs and
YAP is involved in hepatocarcinogenesis. Initially, the
overexpression of YAP promotes the HCC cells to secrete more CTGF,
which in turn regulates HSC function. The extracellular matrix
(ECM) is subsequently activated by HSCs. As a result of the
increasing activity of ECM in the HCC microenvironment, YAP is
translocated from the cytoplasm into the nucleus, and YAP-mediated
gene transcription is activated (29–31).
Notably, the knockdown of YAP has been observed to inhibit the
pluripotency of embryonic stem (ES) cells. By contrast,
overexpression of YAP inhibits ES cell differentiation (32). These studies suggest that the
overexpression of YAP promotes HCC cell proliferation and
transformation (change in cell phenotype), and that YAP may have a
critical role in the development of hepatocarcinogenesis.
Significance of YAP in the diagnosis of
HCC
Tumor screening tests require markers with high
sensitivity and specificity. YAP is overexpressed in patients with
HCC, which suggests that it can be used as a clinical tumor marker
(23).
A previous study suggested that the joint detection
of YAP and AFP is specific for HCC, and that this joint detection
may improve the diagnosis rate (21).
However, YAP is predominantly located in the nuclei of HCC cells,
which makes detection difficult for diagnosis. By contrast,
membrane proteins involved in the organization of tumor cells are
readily released into the blood. Therefore, utilizing
YAP-associated membrane proteins to aid the clinical diagnosis of
HCC has become an area of interest.
The membrane proteins CD166 and MCAM are potential
candidates for HCC diagnostic markers. CD166 is an upstream
regulatory factor of YAP, and it is also upregulated in HCC cells.
CD166 promotes YAP function in hepatocarcinogenesis (33). MCAM is a specific target of YAP in
HCC, and MCAM expression is upregulated in HCC cells. It has been
reported that the serum levels of MCAM and AFP are positively
correlated (34).
In conclusion, serum CD166 and MCAM may replace YAP
as tumor markers for YAP-associated HCC. The elucidation of the
molecular mechanisms of YAP and YAP-associated membrane proteins
may aid the development of non-invasive tests for the diagnosis of
HCC.
Role of YAP in the treatment of HCC
The Hippo/YAP signaling pathway involves a number of
oncogenes and tumor suppressor genes. The inactivation of this
signaling pathway leads to an imbalance between cell proliferation
and apoptosis, which induces hepatocarcinogenesis (35). Investigating the molecular mechanisms
of the Hippo/YAP pathway and other associated factors is beneficial
for elucidating the novel targets for the treatment of HCC.
Use of Hippo/YAP signaling molecules
as targets for the treatment of HCC
YAP is an oncogene. Inhibiting the activity of YAP
may be beneficial for the treatment of HCC (36). Harvey et al (36) proposed that YAP1 may be a potential
target for tumor treatment. However, it was suggested that although
therapies that target YAP1 would be specific, drug resistance is
likely to occur (36). Studies have
demonstrated that knocking down YAP via short hairpin RNAs and
small interfering RNAs can lead to a decrease in the activity of
cyclin E, and reduce cell proliferation and adhesion (24,37).
Furthermore, it has been observed that phosphorylation of YAP
(ser127) is promoted by overexpression of MST1 (38). This suggests that MST1 overexpression
is able to inhibit hepatocarcinogenesis, therefore, it may be a
potential target for the treatment of HCC (38).
A number of microRNAs (miRs/miRNAs) have also been
implicated in hepatocarcinogenesis. However, the interactions
between YAP and miRNAs remain unclear. A previous study
demonstrated that the ectopic expression of miR-375 decreases the
transcriptional activity of YAP mRNA and inhibits the expression of
endogenous YAP proteins (39).
Additionally, Wang et al (40)
demonstrated that miR-506 inhibits YAP expression by targeting the
3′-untranslated region of YAP mRNA and reduces the proliferation of
HCC cells. This indicates that utilizing miR-375 or miR-506 to
target YAP may be a potential strategy to inhibit the proliferation
of HCC cells.
YAP may be targeted to increase the sensitivity of
tumor cells to chemotherapy. It has been reported that the
expression of YAP mRNA increases when HepG2 cells are treated with
doxorubicin (0.7 µg/ml), and that the chemosensitivity of the cells
increases when expression of YAP is increased (41,42). It
has been observed that during chemotherapy, p53 is able to promote
the expression of YAP by binding to the YAP gene promoter in the
2,300 to 2,105 bp region (41,42). These
studies suggest that increasing the expression levels of the
regulatory factors of the Hippo pathway and reducing the expression
of YAP may be a method for treating HCC. Understanding the precise
mechanisms of these interactions would assist in the development of
novel sensitizers for chemotherapy.
Use of YAP-independent factors as
targets for the treatment of HCC
Hepatocarcinogenesis is regulated by numerous
factors (43). YAP expression is not
only regulated by the Hippo/YAP signaling pathway, but is
associated with the activation of YAP-dependent factors; therefore,
the Hippo/YAP signaling pathway may not be the only target for the
treatment of HCC (33). The main
mechanisms between YAP and YAP-independent transcription factors
are presented in Table I.
 | Table I.Interaction between YAP and
YAP-independent transcription factors in HCC. |
Table I.
Interaction between YAP and
YAP-independent transcription factors in HCC.
| Transcription
factor | Mechanism | (Refs.) |
|---|
| TEAD | Dephosphorylated
YAP binds to TEAD via its WW domain, which promotes cell
proliferation and inhibits apoptosis. | (8) |
| CD166 | CD166 modulates the
activity of YAP at the transcriptional and post-transcriptional
levels. YAP is regulated transcriptionally by CD166 through its
interaction with CREB and post-transcriptionally by CD166-mediated
inhibition of AMOT130, which affects the stability of YAP. | (31) |
| MCAM | High levels of p300
promote the binding of YAP to the MCAM promoter, which subsequently
promotes histone acetylation and polymerase II recruitment through
the dissociation of the deacetylase Sirt1. | (32) |
| CREB | YAP inhibits the
degradation of CREB, which is mediated by BTRC and p38. The
accumulation of CREB stimulates transcription of YAP. | (12) |
|
| TRIB2 | TRIB2 represses the
C/EBPα-mediated inhibition of YAP/TEAD transcriptional activation
in HCC cells. | (18) |
| c-Myc | YAP promotes c-Myc
transcriptional output via c-Abl. c-Myc promotes protein expression
independent of transcription. | (44) |
| Shh | Shh promotes YAP
translocation into the nucleus. Shh stabilizes IRS1, and the
interactions between IRS1 and YAP determine the subcellular
localization of YAP. | (46) |
CREB promotes the transcription of YAP by binding to
its promoter (2,608 to 2,439 bp). YAP inhibits CREB degradation
following phosphorylation by interacting with mitogen-activated
protein kinase 14 and β-transducin repeat-containing E3 ubiquitin
protein ligase (BTRC). The interaction between CREB and YAP is
important for HCC cell survival and change in cell phenotype
(12,44).
The Wnt signaling pathway regulates cell
proliferation, differentiation and activity. Studies have reported
that phosphorylation of YAP inhibits Wnt/β-catenin signaling
(19). Additionally, other studies
have reported that TRIB2 is a direct target of Wnt/T-cell factor
signaling in HCC. It has also been demonstrated that TRIB2 is able
to inhibit BTRC, which reduces CCAAT/enhancer binding protein α
(C/EBPα)-mediated inhibition of the transcriptional activity of
YAP/TEAD. These studies suggest that TRIB2 may be a key modulator
with Wnt/β-catenin, Hippo/YAP and C/EBPα signaling; therefore,
TRIB2 may be another target for the treatment of HCC (20,45,46).
c-Myc may be involved in regulating the
transcriptional activity of YAP. Xiao et al (47) demonstrated that YAP-knockdown inhibits
the expression of c-Myc. Further investigation indicated that
although c-Myc-knockdown has no effect on YAP mRNA levels, it
inhibits the expression of human telomerase reverse transcriptase,
which is a downstream target of YAP.
The interactions between Hippo/YAP signaling and a
number of factors, including CD44 (48), Sonic hedgehog (49), Notch (50), insulin-like growth factor 1
receptor/protein kinase B (AKT) (51)
and phosphatidylinositol 3-kinase/AKT (52), are also important in
hepatocarcinogenesis and represent potential therapeutic targets
for HCC. In conclusion, hepatocarcinogenesis is a complex process,
and understanding the interactions between regulatory signals is
useful for altering Hippo/YAP signaling and for identifying novel
targets for the treatment of HCC (Fig.
2).
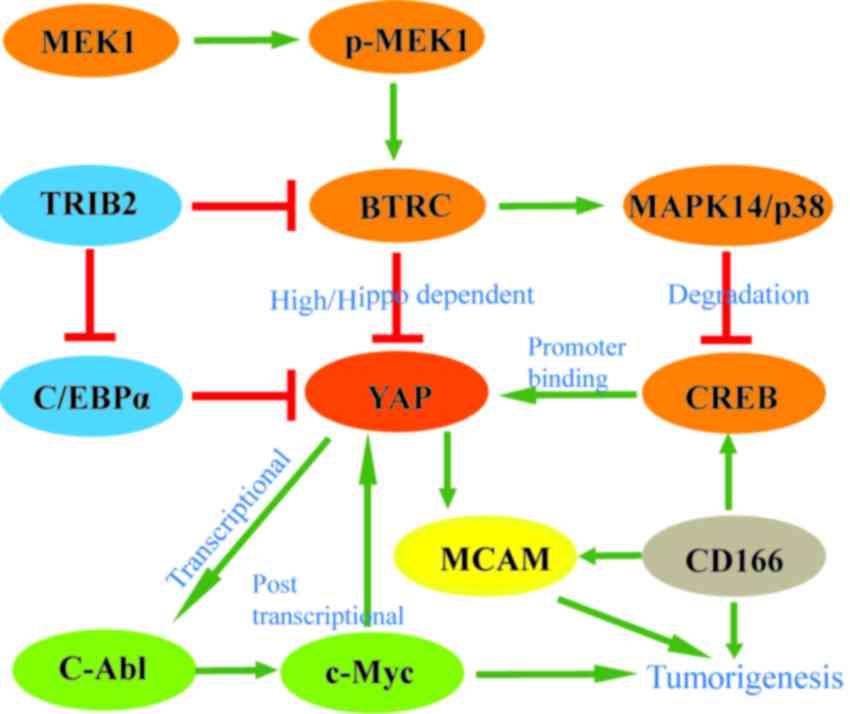 | Figure 2.YAP is regulated by various factors
during hepatocarcinogenesis in addition to the members of the Hippo
signaling pathway. These factors have an important role in
YAP-associated hepatocellular carcinoma. BTRC and TRIB2 inhibit the
overexpression of YAP. The interactions between YAP and other
factors (CREB, c-Myc, CD166 and MCAM) promote hepatocarcinogenesis.
High/hippo dependent refers to the effects of BTRC on YAP, which
are dependent on the Hippo pathway. When BTRC is overexpressed, or
the Hippo pathway is activated and YAP turnover is inhibited;
however, at normal levels of BTRC, YAP is protected from
degradation. BTRC, β-transducin repeat-containing E3 ubiquitin
protein ligase; CD, cluster of differentiation; CREB, cyclic
adenosine monophosphate response element-binding protein; C/EBPα,
CCAAT/enhancer binding protein α; MAPK14, mitogen-activated protein
kinase 14; MCAM, melanoma cell adhesion molecule; p-MEK1,
phosphorylated-mitogen-activated protein kinase; TRIB2, tribbles
homolog 2; YAP, yes-associated protein. |
Role of YAP in determining the prognosis of
HCC
Currently, surgical resection is the preferred
treatment for HCC. However, the 5-year survival rate is only 20 to
40% (53). Following surgery,
residual cancer tissues are highly proliferative, and
overexpression of YAP accelerates the proliferation of HCC cells.
Expression of YAP, a tumor diameter of >5 cm, poor
differentiation and AJCC stage III are independent risk factors of
tumor recurrence post-surgery (4).
Consequently, it is important to investigate the molecular
mechanisms of YAP signaling and its effect in determining the
prognosis of HCC.
Xu et al (23)
analyzed the clinical and pathological data of 166 patients with
HCC and observed that the overexpression of YAP in HCC cells is
associated with a short disease-free survival (DFS) time
(log-rank=3.252; P=0.071) using Kaplan-Meier analysis. The DFS time
was 27.1 months [95% confidence interval (CI), 19.5–34.7] for the
YAP-negative group (n=64) and 14.5 months (95% CI, 10.8–18.2) for
the YAP-positive group (n=102). Therefore, the survival time in HCC
patients with YAP overexpression was significantly reduced compared
with that in patients with YAP-negative HCC following surgery
(23). These findings demonstrated
that there is early cancer recurrence and a poorer prognosis in
YAP-associated HCC, suggesting that altering the expression of YAP
post-surgery may be useful to prolong the survival time of
patients.
Furthermore, F-box and WD repeat domain-containing 7
(Fbxw7), an E3 ubiquitin ligase, may be a potential independent
prognostic marker for YAP-associated HCC. A previous study
demonstrated that Fbxw7 expression inhibits proliferation and
induces apoptosis in HCC cells by promoting the ubiquitination and
proteasomal degradation of YAP, which is crucial for improving the
5-year survival rate (54).
Conclusion
Compared with conventional therapies (radiotherapy
and chemotherapy), targeting YAP and YAP-associated proteins for
the treatment of HCC has the advantage of high selectivity and
specificity (16). YAP and associated
membrane proteins, which are specifically overexpressed in HCC, may
be used as markers to aid in the early diagnosis of HCC.
The molecular mechanisms of Hippo/YAP signaling and
YAP-associated proteins have been gradually elucidated. The factors
identified include YAP, MST1/2, CREB, TRIB2, c-Myc, miR-506, CD44,
Sonic hedgehog, Notch, insulin-like growth factor 1 receptor/AKT,
phosphatidylinositol 3-kinase/AKT and Wnt/β-catenin, which have
been implicated in HCC differentiation, the cell cycle, apoptosis,
migration, invasion, and lymph node and systemic metastasis.
Therefore, investigations into the underlying molecular mechanisms
of interaction between these pathways are beneficial for the
development of high-efficiency and broad-spectrum targeting
drugs.
YAP may be employed as an independent prognostic
marker for HCC. YAP-associated HCC is highly malignant and
aggressive. The recovery rate is low and the prognosis is poor.
Overexpression of YAP during liver regeneration is a leading cause
of early cancer recurrence. This suggests that detecting the change
in YAP expression post-surgery may be beneficial for the early
detection of HCC recurrence and metastasis.
In conclusion, the overexpression of YAP promotes
cell proliferation and phenotypic change in HCC cells. The
interactions between Hippo/YAP signaling and its
associated-proteins, which are involved in hepatocarcinogenesis,
constitute a complex network. The components in this network are
crucial for HCC diagnosis, treatment and prognosis.
The development of HCC treatments targeting YAP may
be a long process. Correlative studies have made considerable
progress. However since the exact molecular mechanisms underlying
the pathogenesis of HCC are not fully understood, the
identification of specific proteins as therapeutic targets for
treatment has considerable limitations. Therefore, elucidating how
YAP is dysregulated in HCC will be pivotal for determining the
factors that should be targeted. Resolving the problem of primary
and acquired drug resistance during YAP-targeting therapy is also a
challenge. Notably, a number of studies have reported that YAP is
able to act as an oncogene and a tumor suppressor gene, indicating
that HCC treatments targeting YAP require further investigation. In
conclusion, whilst the development of therapies involving the
targeting of Hippo/YAP signaling is at its infancy, these therapies
may be novel strategies for treating HCC.
References
|
1
|
Huang J, Wu S, Barrera J, Matthews K and
Pan D: The Hippo signaling pathway coordinately regulates cell
proliferation and apoptosis by inactivating Yorkie, the Drosophila
homolog of YAP. Cell. 122:421–434. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hilman D and Gat U: The evolutionary
history of YAP and the Hippo/YAP pathway. Mol Biol Evol.
28:2403–2417. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yu FX and Guan KL: The Hippo pathway:
Regulators and regulations. Genes Dev. 27:355–371. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zeng Q and Hong W: The emerging role of
the Hippo pathway in cell contact inhibition, organ size control
and cancer development in mammals. Cancer Cell. 13:188–192. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Halder G and Johnson RL: Hippo signaling:
Growth control and beyond. Development. 138:9–22. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Oh H and Irvine KD: Yorkie: The final
destination of Hippo signaling. Trends Cell Biol. 20:410–417. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schlegelmilch K, Mohseni M, Kirak O,
Pruszak J, Rodriguez JR, Zhou D, Kreger BT, Vasioukhin V, Avruch J,
Brummelkamp TR and Camargo FD: YAP1 acts downstream of α-catenin to
control epidermal proliferation. Cell. 144:782–795. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang J, Ji JY, Yu M, Overholtzer M,
Smolen GA, Wang R, Brugge JS, Dyson NJ and Haber DA: YAP-dependent
induction of amphiregulin identifies a non-cell-autonomous
component of the Hippo pathway. Nat Cell Biol. 11:1444–1450. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Strano S and Blandino G: YAP1 meets tumor
suppression. Mol Cell. 27:863–864. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Farazi PA and Depinho RA: Hepatocellular
carcinoma pathogenesis: From genes to environment. Nat Rev Cancer.
6:674–687. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dong J, Feldmann G, Huang J, Wu S, Zhang
N, Comerford SA, Gayyed MF, Anders RA, Maitra A and Pan D:
Elucidation of a universal size-control mechanism in drosophila and
mammals. Cell. 130:1120–1133. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang J, Ma L, Weng W, Qiao Y, Zhang Y, He
J, Wang H, Xiao W, Li L, Chu Q, et al: Mutual interaction between
YAP and CREB promotes tumorigenesis in liver cancer. Hepatology.
58:1011–1020. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tang X, Chen X, Xu Y, Qiao Y, Zhang X,
Wang Y, Guan Y, Sun F and Wang J: CD166 positively regulates MCAM
via inhibition to ubiquitin E3 ligases Smurf1 and βTrCP through
PI3K/AKT and c-Raf/MEK/ERK signaling in Bel-7402 hepatocellular
carcinoma cells. Cell Signal. 27:1694–1702. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Y, Fang R, Cui M, Zhang W, Bai X,
Wang H, Liu B, Zhang X and Ye L: The oncoprotein HBXIP up-regulates
YAP through activation of transcription factor c-Myb to promote
growth of liver cancer. Cancer Lett. 385:234–242. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu G, Wang J, Wu F, Wang N, Zhou W, Wang
Q, Pan W, Ao G and Yang J: YAP and 14–3-3γ are involved in
HS-OA-induced growth inhibition of hepatocellular carcinoma cells:
A novel mechanism for hydrogen sulfide releasing oleanolic acid.
Oncotarget. 7:52150–52165. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Perra A, Kowalik MA, Ghiso E,
Ledda-Columbano GM, Di Tommaso L, Angioni MM, Raschioni C, Testore
E, Roncalli M, Giordano S and Columbano A: YAP activation is an
early event and a potential therapeutic target in liver cancer
development. J Hepatol. 61:1088–1096. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Loforese G, Malinka T, Keogh A, Baier F,
Simillion C, Montani M, Halazonetis TD, Candinas D and Stroka D:
Impaired liver regeneration in aged mice can be rescued by
silencing Hippo core kinases MST1 and MST2. EMBO Mol Med. 9:46–60.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yi J, Lu L, Yanger K, Wang W, Sohn BH,
Stanger BZ, Zhang M, Martin JF, Ajani JA, Chen J, et al: Large
tumor suppressor homologs 1 and 2 regulate mouse liver progenitor
cell proliferation and maturation through antagonism of the
coactivators YAP and TAZ. Hepatology. 64:1757–1772. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Perumal N, Perumal M, Kannan A, Subramani
K, Halagowder D and Sivasithamparam N: Morin impedes Yap nuclear
translocation and fosters apoptosis through suppression of
Wnt/β-catenin and NF-κB signaling in Mst1 overexpressed HepG2
cells. Exp Cell Res. 355:124–141. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang J, Park J, Wei Y, Rajurkar M, Cotton
JL, Fan Q, Lewis BC, Ji H and Mao J: TRIB2 acts downstream of
Wnt/TCF in liver cancer cells to regulate YAP and C/EBPα function.
Mol Cell. 51:211–225. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang YP and Tang DX: Expression of
Yes-associated protein in liver cancer and its correlation with
clinicopathological features and prognosis of liver cancer
patients. Int J Clin Exp Med. 8:1080–1086. 2015.PubMed/NCBI
|
|
22
|
Tordjmann T: Hippo signaling: Liver size
regulation and beyond. Clin Res Hepatol Gastroenterol. 35:344–346.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu MZ, Yao TJ, Lee NP, Ng IO, Chan YT,
Zender L, Lowe SW, Poon RT and Luk JM: Yes-associated protein is an
independent prognostic marker in hepatocellular carcinoma. Cancer.
115:4576–4585. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zender L, Spector MS, Xue W, Flemming P,
Cordon-Cardo C, Silke J, Fan ST, Luk JM, Wigler M, Hannon GJ, et
al: Idetification and validation of oncogenes in liver cancer using
an integrative oncogenomic approach. Cell. 125:1253–1267. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao B, Ye X, Yu J, Li L, Li W, Li S, Yu
J, Lin JD, Wang CY, Chinnaiyan AM, et al: TEAD mediates
YAP-dependent gene induction and growth control. Genes Dev.
22:1962–1971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cai WY, Lin LY, Hao H, Zhang SM, Ma F,
Hong XX, Zhang H, Liu QF, Ye GD, Sun GB, et al: Yes-associated
protein/TEA domain family member and hepatocyte nuclear factor
4-alpha (HNF4α) repress reciprocally to regulate
hepatocarcinogenesis in rats and mice. Hepatology. 65:1206–1221.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lin C, Hu Z, Lei B, Tang B, Yu H, Qiu X
and He S: Overexpression of Yes-associated protein and its
association with clinicopathological features of hepatocellular
carcinoma: A meta-analysis. Liver Int. Mar 26–2017.(Epub ahead of
print). View Article : Google Scholar
|
|
28
|
Song Y, Kim SH, Kim KM, Choi EK, Kim J and
Seo HR: Activated hepatic stellate cells play pivotal roles in
hepatocellular carcinoma cell chemoresistance and migration in
multicellular tumor spheroids. Sci Rep. 6:367502016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mannaerts I, Leite SB, Verhulst S,
Claerhout S, Eysackers N, Thoen LF, Hoorens A, Reynaert H, Halder G
and van Grunsven LA: The Hippo pathway effector YAP controls mouse
hepatic stellate cell activation. J Hepatol. 63:679–688. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Paget S: The distribution of secondary
growths in cancer of the breast. 1989. Cancer Metastasis Rev.
8:98–101. 1989.PubMed/NCBI
|
|
31
|
Okabe H, Beppu T, Hayashi H, Horino K,
Masuda T, Komori H, Ishikawa S, Watanabe M, Takamori H, Iyama K and
Baba H: Hepatic stellate cells may relate to progression of
intrahepatic cholangiocarcinoma. Ann Surg Oncol. 16:2555–2564.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lian I, Kim J, Okazawa H, Zhao J, Zhao B,
Yu J, Chinnaiyan A, Israel MA, Goldstein LS, Abujarour R, et al:
The role of YAP transcription coactivator in regulating stem cell
self-renewal and differentiation. Genes Dev. 24:1106–1118. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ma L, Lin J, Qiao Y, Weng W, Liu W, Wang J
and Sun F: Serum CD166: A novel hepatocellular carcinoma tumor
marker. Clin Chim Acta. 441:156–162. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang J, Tang X, Weng W, Qiao Y, Lin J, Liu
W, Liu R, Ma L, Yu W, Yu Y, et al: The membrane protein melanoma
cell adhesion molecule (MCAM) is a novel tumor marker that
stimulates tumorigenesis in hepatocellular carcinoma. Oncogene.
36:5781–5795. 2015. View Article : Google Scholar
|
|
35
|
Camargo FD, Gokhale S, Johnnidis JB, Fu D,
Bell GW, Jaenisch R and Brummelkamp TR: YAP1 increases organ size
and expands undifferentiated progenitor cells. Curr Bio1.
17:2054–2060. 2007. View Article : Google Scholar
|
|
36
|
Harvey KF, Zhang X and Thomas DM: The
Hippo pathway and human cancer. Nat Rev Cancer. 13:246–257. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu AM, Xu MZ, Chen J, Poon RT and Luk JM:
Targeting YAP and Hippo signaling pathway in liver cancer. Expert
Opin Ther Targets. 14:855–868. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhou D, Conrad C, Xia F, Park JS, Payer B,
Yin Y, Lauwers GY, Thasler W, Lee JT, Avruch J and Bardeesy N: Mst1
and Mst2 maintain hepatocyte quiescence and sup-press
hepatocellular carcinoma development through inac-tivation of the
Yap1 oncogene. Cancer Cell. 16:425–438. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu AM, Poon RT and Luk JM: MicroRNA-375
targets Hippo-signaling effector YAP in liver cancer and inhibits
tumor properties. Biochem Biophys Res Commun. 394:623–627. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang Y, Cui M, Sun BD, Liu FB, Zhang XD
and Ye LH: miR-506 suppresses proliferation of hepatoma cells
through targeting YAP mRNA 3′UTR. Acta Pharmacol Sin. 35:1207–1214.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bai N, Zhang C, Liang N, Zhang Z, Chang A,
Yin J, Li Z, Luo N, Tan X, Luo N, et al: Yes-associated protein
(YAP) increases chemosensitivity of hepatocellular carcinoma cells
by modulation of p53. Cancer Biol Ther. 14:511–520. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang W and Cohen SM: The Hippo pathway
acts via p53 and microRNAs to control proliferation and
proapoptotic gene expression during tissue growth. Biol Open.
2:822–828. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shimizu D, Inokawa Y, Sonohara F, Inaoka K
and Nomoto S: Search for useful biomarkers in hepatocellular
carcinoma, tumor factors and background liver factors (Review).
Oncol Rep. 37:2527–2542. 2017.PubMed/NCBI
|
|
44
|
Noubissi FK, Elcheva I, Bhatia N, Shakoori
A, Ougolkov A, Liu J, Minamoto T, Ross J, Fuchs SY and Spiegelman
VS: CRD-BP mediates stabilization of betaTrCP1 and c-myc mRNA in
response to beta-catenin signalling. Nature. 441:898–901. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Thompson MD and Monga SP: Wnt/beta-catenin
signaling in liver health and disease. Hepatology. 45:1298–1305.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Qiao Y, Zhang Y and Wang J: Ubiquitin E3
ligase SCF(β-TRCP) regulates TRIB2 stability in liver cancer cells.
Biochem Biophys Res Commun. 441:555–559. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xiao W, Wang J, Ou C, Zhang Y, Ma L, Weng
W, Pan Q and Sun F: Mutual interaction between YAP and c-Myc is
critical for carcinogenesis in liver cancer. Biochem Biophys Res
Commun. 439:167–172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xu Y, Stamenkovic I and Yu Q: CD44
attenuates activation of the Hippo signaling pathway and is a prime
therapeutic target for glioblastoma. Cancer Res. 70:2455–2464.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Fernandez LA, Northcott PA, Dalton J,
Fraga C, Ellison D, Angers S, Taylor MD and Kenney AM: YAP1 is
amplified and upregulated in hedgehog-associated medulloblastomas
and mediates Sonic hedgehog-driven neural precursor proliferation.
Genes Dev. 23:2729–2741. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yu JZ, Poulton J, Huang YC and Deng WM:
The hippo pathway promotes Notch signaling in regulation of cell
differentiation, proliferation, and oocyte polarity. PLoS One.
3:e17612008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kim D, Shu S, Coppola MD, Kaneko S, Yuan
ZQ and Cheng JQ: Regulation of proapoptotic mammalian ste20-like
kinase MST2 by the IGF1-Akt pathway. PLoS One. 5:e96162010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yuan Z, Kim D, Shu S, Wu J, Guo J, Xiao L,
Kaneko S, Coppola D and Cheng JQ: Phosphoinositide 3-kinase/Akt
inhibits MST1-mediated pro-apoptotic signaling through
phosphorylation of threonine 120. J Biol Chem. 285:3815–3824. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Joliat GR, Allemann P, Labgaa I,
Demartines N and Halkic N: Treatment and outcomes of recurrent
hepatocellular carcinomas. Langenbecks Arch Surg. May 11–2017.(Epub
ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tu K, Yang W, Li C, Zheng X, Lu Z, Guo C,
Yao Y and Liu Q: Fbxw7 is an independent prognostic marker and
induces apoptosis and growth arrest by regulating YAP abundance in
hepatocellular carcinoma. Mol Cancer. 13:1102014. View Article : Google Scholar : PubMed/NCBI
|














