Introduction
Prostate cancer is the second most common type of
malignancy in the male population worldwide, and is the third
leading cause of cancer-associated mortality in male patients
(1). Surgery and chemotherapy are the
main treatment strategies for prostate cancer (2–4). Due to
the malignant migration and invasion potential of human prostate
cancer cells, the current therapy for this type of cancer remains
unsatisfactory (5). In addition, the
molecular mechanism behind the progression of prostate cancer
remains largely unclear.
MicroRNAs (miRNAs) are a class of small non-coding
RNAs, which control protein-coding gene expression by directly
binding the 3′-untranslated regions (3′-UTRs) of these genes
(6,7).
Accumulating evidence has indicated that miRNAs are important
regulatory factors involved in various biological processes,
including cell differentiation, growth, metastasis and apoptosis
(8,9).
Aberrant expression of miRNAs has been identified in numerous
cancers, including lung and breast cancer and hepatocellular
carcinoma (10–15). Numerous miRNAs may be regarded as
potential and novel biomarkers for the diagnosis, therapy and
prognosis of numerous types of cancer (16,17).
A large number of miRNAs have been identified to be
aberrantly expressed in human prostate cancer, including miR-22,
miR-23b/27b, miR-103, miR-135b, miR-181, miR-192 and miR-613
(18–24). These miRNAs perform tumor-suppressive
or oncogenic roles in the regulation of prostate cancer cell
growth, invasion, migration and apoptosis. The mature sequence of
miR-20b is located at chromosome Xq26.2, which is a fragile
chromosomal region associated with numerous types of human cancer
(25). However, the role of miR-20b
in prostate cancer remains unknown.
The present study investigated the potential effect
of miR-20b on prostate cancer. Firstly, the expression of miR-20b
was examined in prostate cancer tissues and adjacent normal
prostate tissues. MTT and wound-healing assays were then used to
detect the effect of miR-20b on prostate cancer cell growth and
migration. Finally, phosphatase and tensin homolog (PTEN) was
identified as a potential target gene of miR-20b by bioinformatics
analysis, dual-luciferase reporter assay and western blot
analysis.
Materials and methods
Clinical specimens
A total of 35 pairs of prostate cancer tissues and
adjacent normal prostate tissues were obtained from the Institute
of Urology, First Affiliated Hospital of Nanchang University
(Nanchang, China). The patients with prostate cancer did not
receive chemotherapy or radiotherapy prior to prostatectomy. All
samples were immediately snap-frozen in liquid nitrogen until RNA
extraction. The histological diagnosis of each sample was confirmed
simultaneously by two pathologists using hematoxylin and eosin
staining. Written consent forms were obtained from each patient,
and the present study was approved by the Ethics Committee of First
Affiliated Hospital of Nanchang University (Nanchang, China).
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA of each sample was isolated using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), following the manufacturer's protocol. Total RNA (1 µg) was
treated with DNase to remove contaminating DNA. Subsequently, the
RNA was reverse transcribed into first-strand complementary DNA
(cDNA) using M-MLV reverse transcriptase (Promega Corporation,
Madison, WI, USA) for mRNA expression analysis. The miScript
reverse transcription kit (Qiagen GmbH, Hilden, Germany) was used
to reverse total RNA into cDNA for miRNA expression analysis.
RT-qPCR reaction was performed with the Power SYBR-Green PCR Master
Mix (Applied Biosystems; Thermo Fisher Scientific, Inc.) according
to the manufacturer's protocol on the Roche LightCycler 480
Real-Time PCR Machine (Roche Diagnostics, Basel, Switzerland). The
qPCR conditions consisted of a uracil-N-glycosylase carry-over
protection step of 55°C for 2 min, 10 min of DNA polymerase
activation at 95°C, followed by 40 cycles of 95°C for 10 sec and
60°C for 30 sec. The gene or miRNA primers were obtained from
Invitrogen (Thermo Fisher Scientific, Inc.). The sequences of
primers are shown in Table I. U6 and
GAPDH were used as the internal controls. The relative gene and
miRNA expression levels were calculated using the 2−ΔΔCq
method (26). Each experiment was
conducted at least three times.
 | Table I.Primers for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Forward (5′-3′) | Reverse (5′-3′ |
|---|
| miR-20b |
GAGGACGGAACCGGAAAC | Universal primer |
| PTEN |
GTGCAGATAATGACAAG |
GATTTGACGGCTCCTCT |
| U6 | CTCGCTTCGGCAGCAC |
ACGCTTCACGAATTTGC |
| GAPDH |
CAAGGTCATCCATGACAA |
GTCCACCACCCTGTTGCTG |
Cell culture and transfection
Two human prostate cancer cell lines (VCaP and PC-3)
were purchased from Shanghai Institute of Biological Sciences,
Chinese Academy of Science (Shanghai, China). All cells were
cultured in RPMI-1640 (Invitrogen; Thermo Fisher Scientific, Inc.)
containing 10% fetal bovine serum (Invitrogen; Thermo Fisher
Scientific, Inc.) and 1% penicillin/streptomycin (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany). Cells were maintained at 37°C in a
humidified atmosphere with 5% CO2.
A total of four pairs of inhibitors (inhibitor
1#-4#) and corresponding inhibitor controls (inhibitor control
1#-4#) of miR-20b were obtained from Shanghai GenePharma Co., Ltd.
(Shanghai, China). The sequences were as follows: Inhibitor 1#,
5′-UGCUCAUAGUGCAGGUAGUU-3′ and inhibitor control 1#,
5′-GAUGAGCAACAUUGAGGACU-3′; inhibitor 2#,
5′-GCAGGUAGUUUUGGCAUGAC-3′ and inhibitor control 2#,
5′-AGCCCGGAUUACCUGUAGCU-3′; inhibitor 3#,
5′-UCUACUGUAGUAUGGGCACU-3′ and inhibitor control 3#,
5′-GACCCUCAUUCCACGCAUC-3′; inhibitor 4#, 5′-UAUGGGCACUUCCAGUACU-3′
and inhibitor control 4#, 5′-CAUACAUUACCCGAAGUCUA-3′. The
inhibitors and inhibitor controls were transfected into VCaP and
PC-3 cells using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) at a concentration of 50 nM, according to
the manufacturer's protocol.
Cell proliferation assay
Cell proliferation was examined using an MTT assay.
Briefly, ~8,000 VCaP and PC-3 cells/well were seeded onto 96-well
plates. After 24 h, cells were transfected with miR-20b inhibitor
and inhibitor control for 48 h and then treated with 20 ml/well MTT
(Sigma-Aldrich; Merck KGaA; 5 mg/ml). Following incubation for 4 h
at 37°C, the free supernatant of cells was discarded and the
formazan products were dissolved by 150 µl dimethyl sulfoxide.
Finally, the optical density (OD) at 450 nm was detected by a
microplate reader (Synergy™ HT Multi-Mode Microplate Reader; BioTek
Instruments, Inc., Winooski, VT, USA).
Cell migration assay
A wound-healing assay was used to assess prostate
cancer cell migration ability of miR-20b. VCaP and PC-3 cells were
plated onto 12-well plates and transfected with miR-20b inhibitor
and inhibitor control. Following transfection for 24 h at 37°C in a
humidified atmosphere with 5% CO2, wounds were made with
a yellow pipette tip among cells in each well. Images were captured
in five random visual fields (magnification, ×100) at 0 and 12 h by
a Leica DMI 6000B microscope (Leica Microsystems, Inc., Buffalo
Grove, IL, USA) subsequent to wounding in order to determine the
width of wound healing.
Western blot analysis
The protein of cells was extracted using lysis
buffer (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. 45 µg of protein samples in each group
was subjected to 10% SDS-PAGE, and then transferred to
polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA,
USA). Subsequent to blocking with 5% non-fat milk for 2 h at 37°C,
the membrane was incubated with the rabbit monoclonal anti-PTEN
antibodies (dilution, 1:500; cat. no. ab32199; Abcam, Cambridge,
UK) overnight at 4°C, followed by anti-rabbit horseradish
peroxidase-linked secondary antibodies (dilution, 1:2,000; cat. no.
ab6721; Abcam) for 1 h at 37°C. The bands were obtained using
AnalySIS 3.0 image analysis system (Soft Imaging System GmbH,
Münster, Germany), and the protein density was quantified with
Odyssey v1.2 software (LI-COR Biosciences, Lincoln, NE, USA).
Bioinformatics analysis
The potential target of miR-20b was created by
combining three public algorithms, which were TargetScan
(http://www.targetscan.org/), miRanda
(http://www.targetscan.org/) and PicTar
(http://pictar.mdc-berlin.de/). The
putative genes that were predicted by the three algorithms were
accepted and the candidates were chosen based on the gene
function.
Dual-luciferase reporter assay
The mRNA 3′-UTR of PTEN containing the
predicted binding region or mutated binding region was sub-cloned
into a psiCHECK-2 luciferase reporter vector (Promega Corporation).
The psiCHECK-2 vector containing wild-type (WT) or mutant (MUT)
mRNA 3′-UTRs of PTEN with the miR-20b inhibitors or inhibitor
controls were co-transfected into VCaP and PC-3 cells using
Lipofectamine 2000 at 37°C in a humidified atmosphere with 5%
CO2. Luciferase activities were detected by the
Dual-Luciferase Reporter Assay System (Promega Corporation) 48 h
post-transfection, according to the manufacturer's protocol.
Statistical analysis
Each experiment was performed in triplicate for
biological repeat. Statistical analysis was performed using SPSS
17.0 (SPSS, Inc., Chicago, IL, USA). Data are presented as the mean
± standard deviation. Statistical differences were analyzed using
Student's t-test or one-way analysis of variance. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-20b is significantly upregulated
in prostate cancer tissues
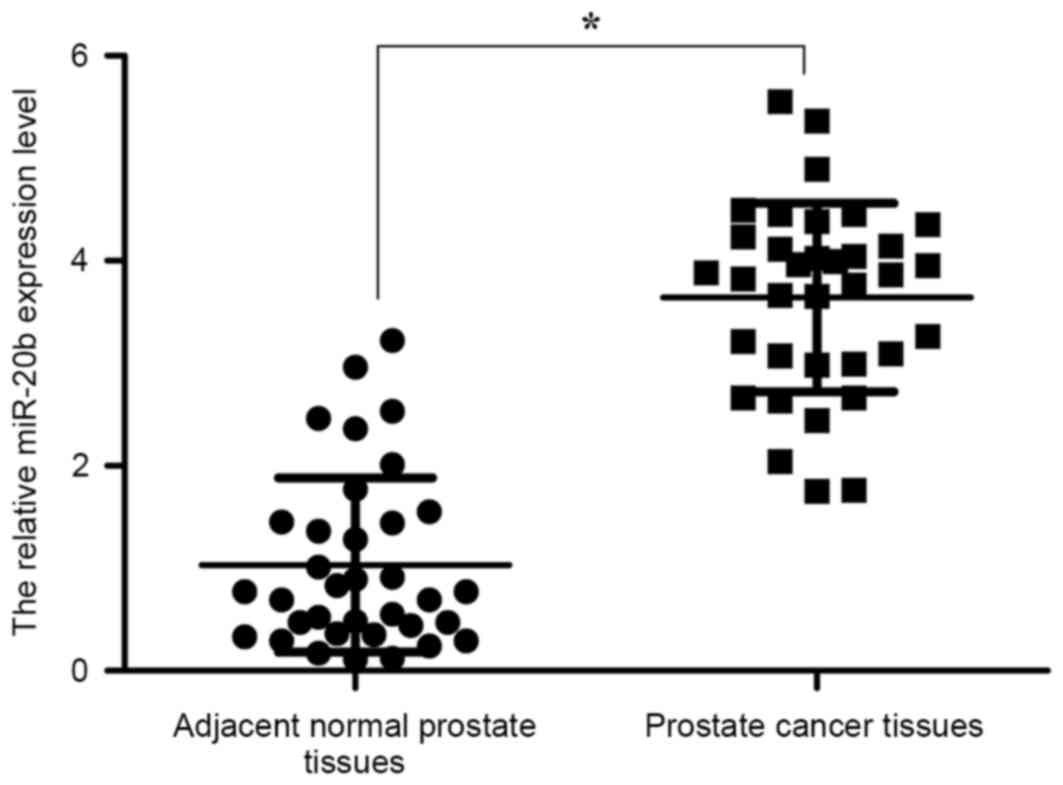
The expression level of miR-20b in 35 pairs of
prostate cancer tissues and adjacent normal prostate tissues was
analyzed by RT-qPCR. The results revealed that the miR-20b
expression was significantly upregulated in prostate cancer
tissues, compared with adjacent normal prostate tissues (P<0.05;
Fig. 1). The data indicated that the
upregulation of miR-20b may be involved in the development of human
prostate cancer.
Knockdown of miR-20b inhibits prostate
cancer cell proliferation
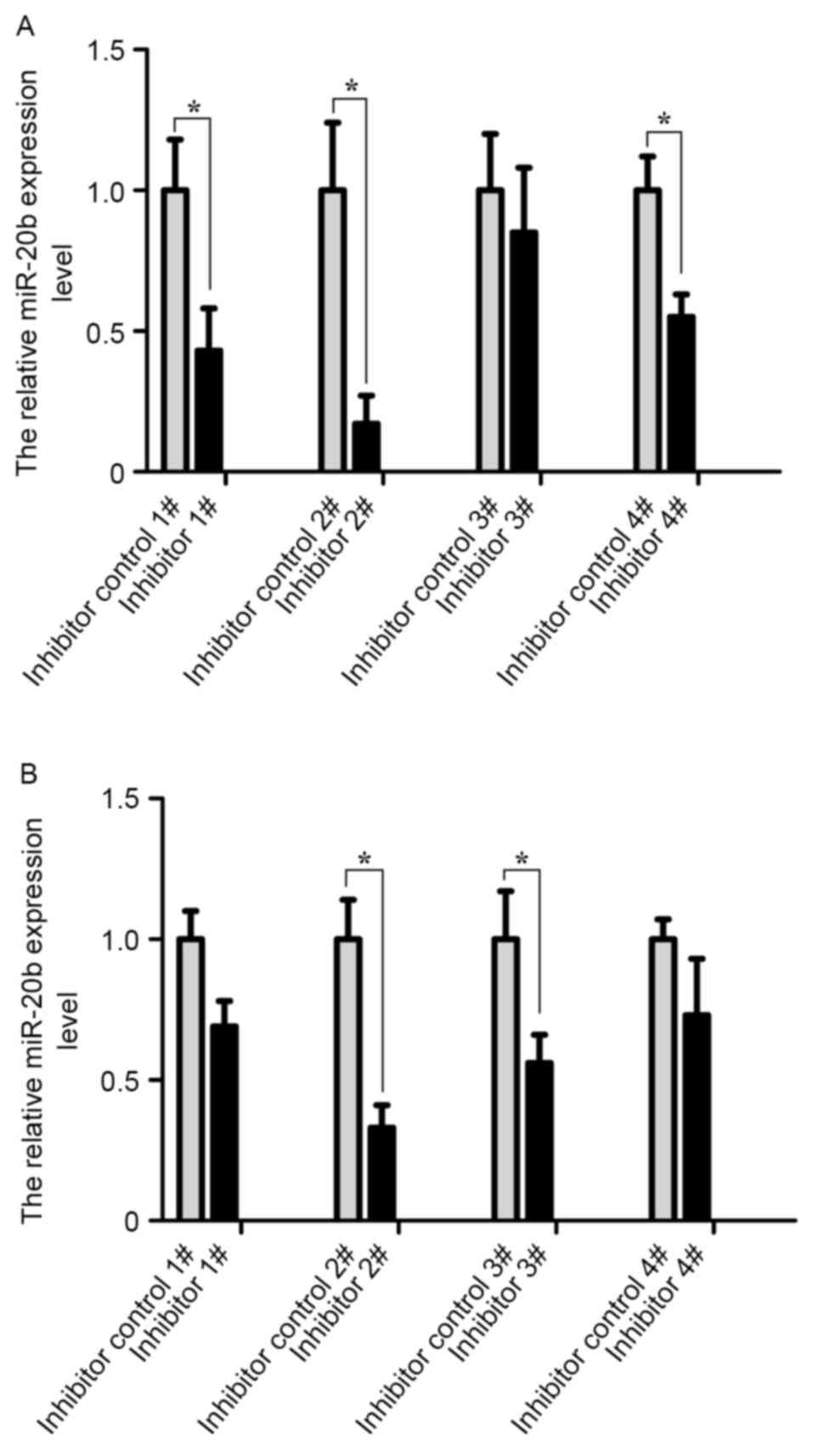
A total of four pairs of inhibitors and inhibitor
controls of the miR-20b were transfected into VCaP and PC-3 cells.
The transfection efficiency was analyzed by RT-qPCR at 24 h
post-transfection. As shown in Fig.
2, the transfection efficiency of cells was the highest in the
inhibitor 2# group compared with other groups (P<0.05).
Therefore, the miR-20b inhibitor 2# group was selected for
subsequent experiments.
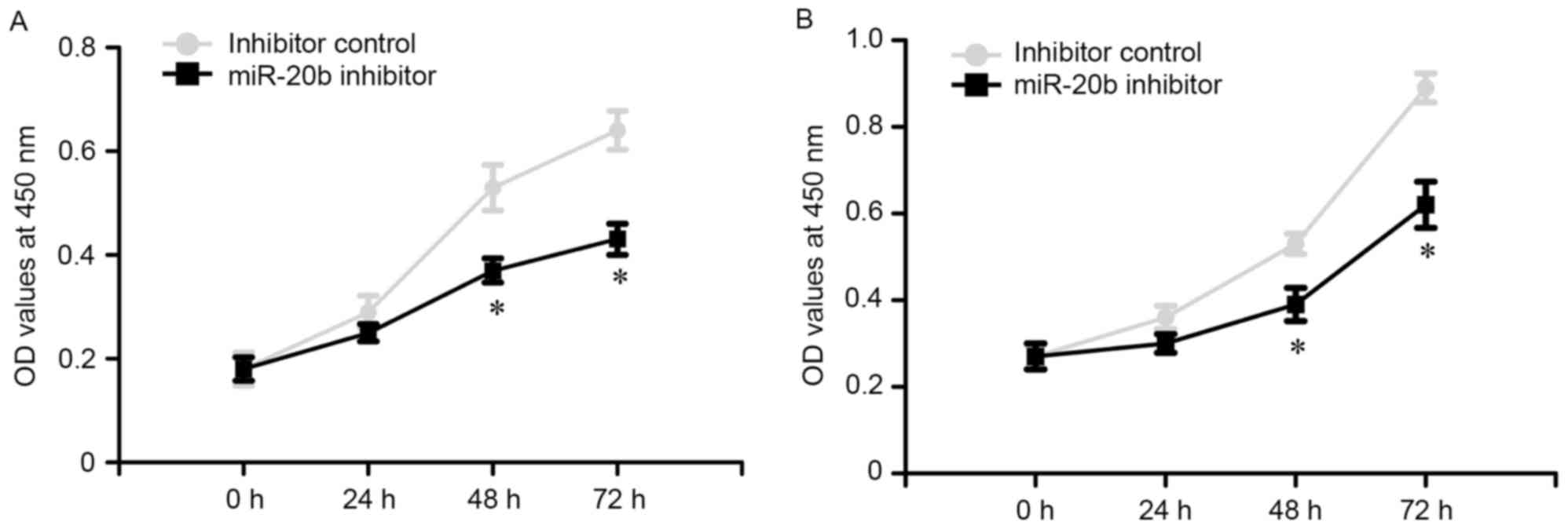
To evaluate the potential role of miR-20b on the
proliferation of prostate cancer cells, a MTT assay was performed.
The OD values of VCaP and PC-3 cells were measured at 0, 24, 48 and
72 h following transfection. The results revealed that miR-20b
inhibitor reduced the growth of VCaP and PC-3 cells, as compared
with the inhibitor control (Fig. 3;
P<0.05).
Knockdown of miR-20b suppresses
prostate cancer cell migration
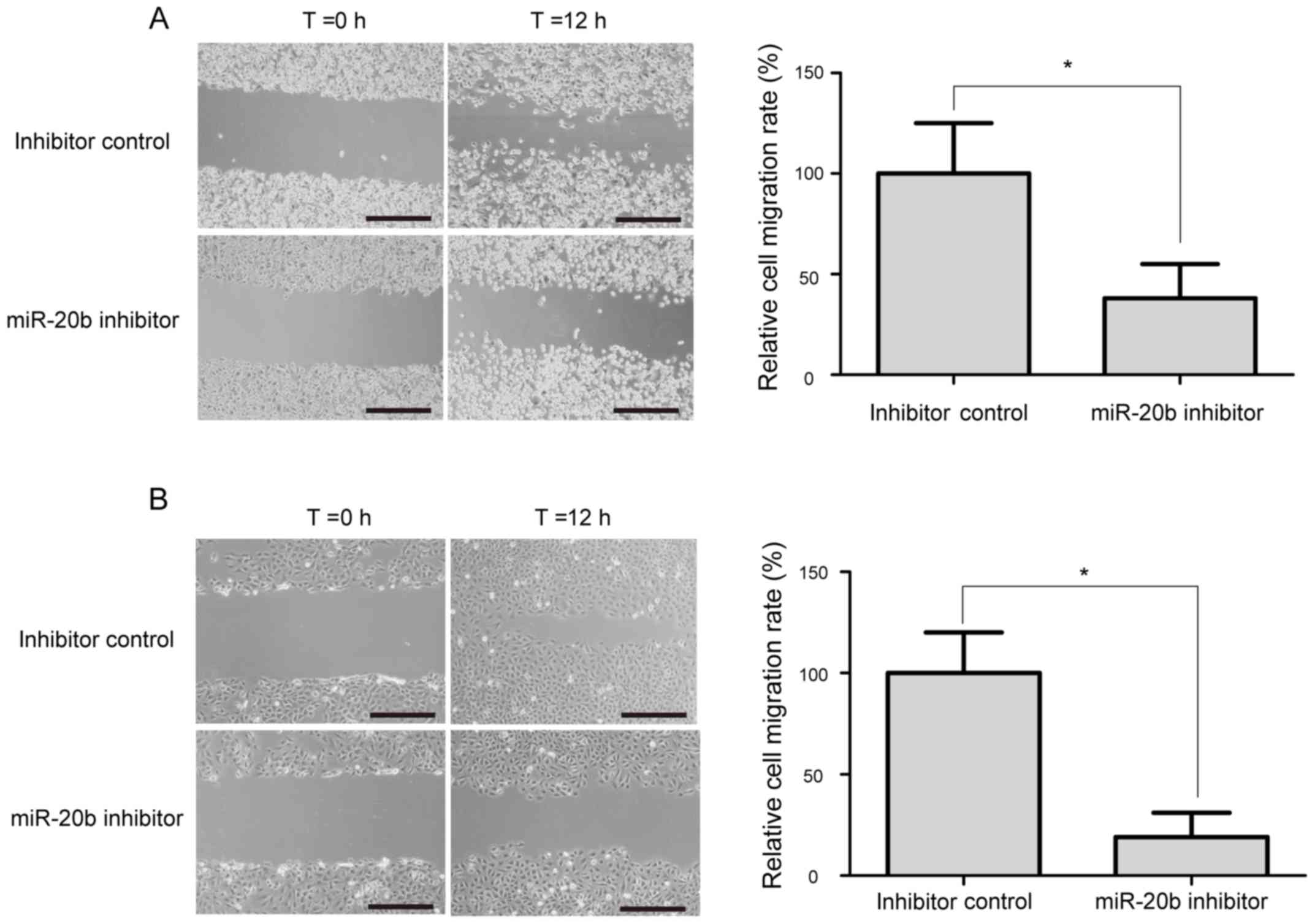
The wound-healing assay was performed to evaluate
the potential role of miR-20b on the migration of prostate cancer
cell. As shown in Fig. 4, a
significant decrease in cell migration ability was observed in
miR-20b inhibitor-treated VCaP and PC-3 cells, compared with
inhibitor control-transfected cells (P<0.05).
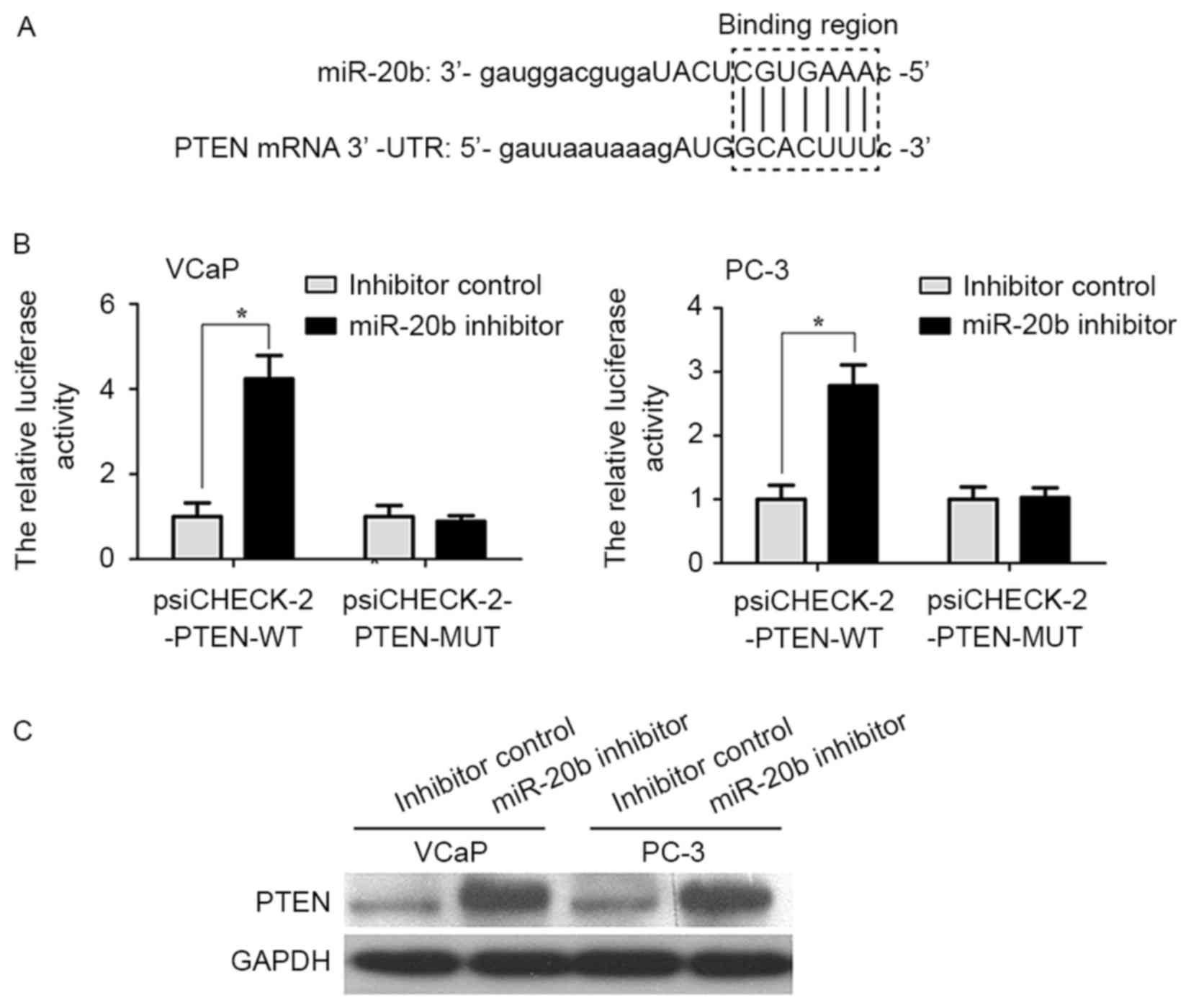
PTEN is a direct target gene of
miR-20b in prostate cancer cells
Bioinformatics analysis predicted that PTEN was a
target gene of miR-20b (Fig. 5A). In
order to provide additional direct evidence for the association
between PTEN and miR-20b, the binding region of miR-20b in the mRNA
3′UTR of PTEN was characterized by the dual-luciferase reporter
assay. As shown in Fig. 5B, the
relative luciferase activity of the psiCHECK-2-PTEN-WT vector in
VCaP and PC-3 cells was increased compared with in the
psiCHECK-2-PTEN-MUT vector (P<0.05). In addition, it was
examined whether miR-20b inhibits endogenous PTEN expression in
VCaP and PC-3 cells. Compared with inhibitor control, endogenous
PTEN protein expression levels were significantly increased in VCaP
and PC-3 cells transfected with miR-20b inhibitor compared with
that in inhibitor control-transfected cells (Fig. 5C; P<0.05).
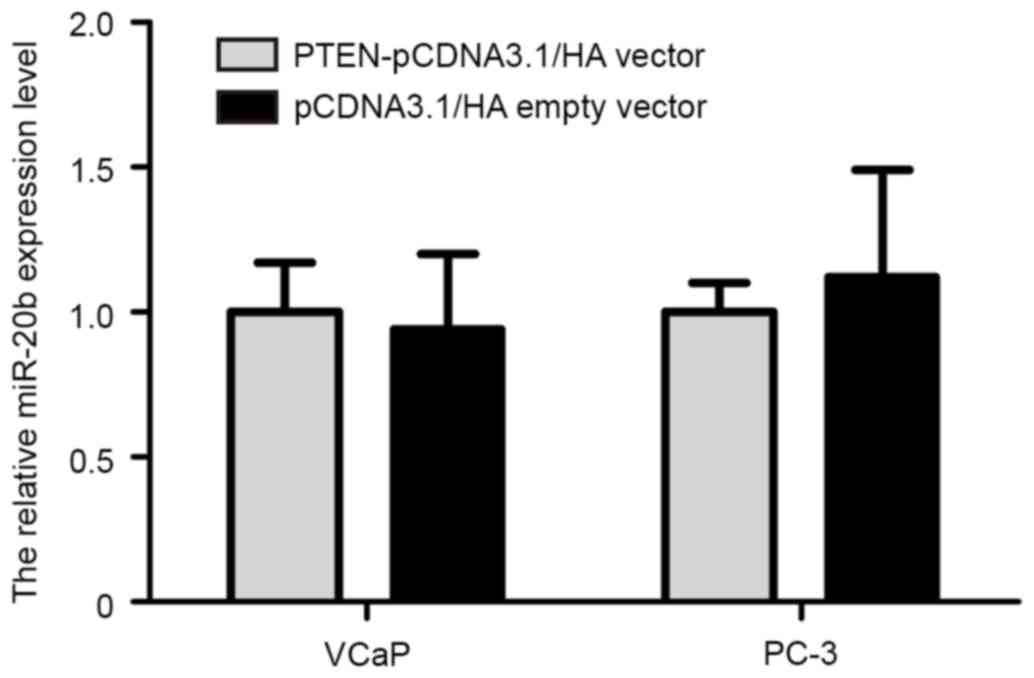
Accumulating evidence has indicated that certain
target genes may also reverse regulate expression of miRNA. In
order to investigate whether PTEN may regulate miR-20b expression
in prostate cancer cells, PTEN overexpression plasmid
(PTEN-pCDNA3.1/HA) was synthesized (Invitrogen; Thermo Fisher
Scientific, Inc.). The PTEN-pCDNA3.1/HA and pCDNA3.1/HA empty
vectors were transfected into VCaP and PC-3 cells using
Lipofectamine 2000, according to the manufacturer's protocol. The
results demonstrated that restoration of PTEN expression in VCaP
and PC-3 cells did not alter the expression level of miR-20b
(Fig. 6).
Discussion
Previous studies have reported that miR-20b is
upregulated in gastric and cervical cancer (27,28).
Studies have indicated that miR-20b functions as an oncogene in
numerous types of cancer. miR-20b promotes breast cancer cell
growth, partly by targeting the PTEN gene (29). miR-20b suppresses PTEN expression
resulting in B7-H1 overexpression in advanced colorectal cancer
(30). miR-20b presents high
expression level and has oncogenic potential in human T-cell
leukemia (31). However, the
expression and function of miR-20b has not been documented in
prostate cancer.
In the present study, miR-20b appeared to be
markedly upregulated in prostate cancer tissues compared with
adjacent normal prostate tissues, which is consistent with previous
studies (27–29,31). The
proliferation and migration abilities of human prostate cancer
cells transfected with miR-20b inhibitor were significantly reduced
compared with the inhibitor control, which indicated that miR-20b
acts as an oncogene in prostate cancer cells. These results were
also similar to observations in colorectal and breast cancer
(32,33), in which miR-20b expression was
upregulated, and knockdown of miR-20b inhibited cellular
proliferation and migration.
The PTEN gene is a classic tumor suppressor gene in
various human cancers, which is located at chromosome 10q23.31
(34). PTEN functions as a negative
regulator of the phosphoinositide 3-kinase/Akt pathway through
dephosphorylation of phosphatidylinositol 3,4,5 trisphosphate, and
is involved in regulation of cellular proliferation, metastasis and
apoptosis during progression of cancers (34). PTEN has been reported to be regulated
by numerous miRNAs in multiple cancers, including colorectal
carcinoma, glioma, ovarian and breast cancer (29,35–37). In
the present study, it was also determined that miR-20b inhibited
PTEN expression by directly binding to the mRNA 3′-UTR of PTEN in
VCaP and PC-3 cells. Additionally, restoration of PTEN expression
did not affect endogenous miR-20b expression in VCaP and PC-3
cells. Above all, the present study indicated that miR-20b serves
as an important oncogene in promoting prostate cancer cell growth
and migration by directly regulating PTEN.
In conclusion, the present study identified that
miR-20b expression was significantly upregulated in prostate cancer
tissues. Notably, knockdown of miR-20b expression exhibits an
anti-tumor effect in vitro. miR-20b acts as an oncogene that
performs a critical role in the growth and migration of prostate
cancer cell by targeting PTEN. Therefore, miR-20b may serve as a
novel biomarker and potential therapeutic target in prostate
cancer.
Acknowledgements
The present study was supported by the Natural
Science Foundation of Jiangxi province (grant no.
20151BAB205017).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Silvestri I, Cattarino S, Aglianò AM,
Collalti G and Sciarra A: Beyond the immune suppression: The
immunotherapy in prostate cancer. Biomed Res Int. 2015:7949682015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rose JN and Crook JM: The role of
radiation therapy in the treatment of metastatic castrate-resistant
prostate cancer. Ther Adv Urol. 7:135–145. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nazim SM and Abbas F: Role of surgery in
locally advanced prostate cancer. Pak J Med Sci. 31:710–716.
2015.PubMed/NCBI
|
|
5
|
De Marzo AM, DeWeese TL, Platz EA, Meeker
AK, Nakayama M, Epstein JI, Isaacs WB and Nelson WG: Pathological
and molecular mechanisms of prostate carcinogenesis: Implications
for diagnosis, detection, prevention, and treatment. J Cell
Biochem. 91:459–477. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mo YY: MicroRNA regulatory networks and
human disease. Cell Mol Life Sci. 69:3529–3531. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Denli AM, Tops BB, Plasterk RH, Ketting RF
and Hannon GJ: Processing of primary microRNAs by the
Microprocessor complex. Nature. 432:231–235. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Miska EA: How microRNAs control cell
division, differentiation and death. Curr Opin Genet Dev.
15:563–568. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Winter J, Jung S, Keller S, Gregory RI and
Diederichs S: Many roads to maturity: MicroRNA biogenesis pathways
and their regulation. Nat Cell Biol. 11:228–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang Y, Cui Y, Li Z, Jiao Z, Zhang Y, He
Y, Chen G, Zhou Q, Wang W and Zhou X: Radiation-induced miR-208a
increases the proliferation and radioresistance by targeting p21 in
human lung cancer cells. J Exp Clin Cancer Res. 35:72016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim G, An HJ, Lee MJ, Song JY, Jeong JY,
Lee JH and Jeong HC: Hsa-miR-1246 and hsa-miR-1290 are associated
with stemness and invasiveness of non-small cell lung cancer. Lung
Cancer. 91:15–22. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Seviour EG, Sehgal V, Lu Y, Luo Z, Moss T,
Zhang F, Hill SM, Liu W, Maiti SN, Cooper L, et al: Functional
proteomics identifies miRNAs to target a p27/Myc/phospho-Rb
signature in breast and ovarian cancer. Oncogene. 35:8012016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rinnerthaler G, Hackl H, Gampenrieder SP,
Hamacher F, Hufnagl C, Hauser-Kronberger C, Zehentmayr F, Fastner
G, Sedlmayer F, Mlineritsch B and Greil R: miR-16-5p is a
stably-expressed housekeeping microRNA in breast cancer tissues
from primary tumors and from metastatic sites. Int J Mol Sci.
17:pii: E1562016. View Article : Google Scholar
|
|
14
|
Ge Y, Yan X, Jin Y, Yang X, Yu X, Zhou L,
Han S, Yuan Q and Yang M: MiRNA-192 [corrected] and miRNA-204
directly suppress lncRNA HOTTIP and interrupt GLS1-mediated
glutaminolysis in hepatocellular carcinoma. PLoS Genet.
11:e10057262015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gao F, Sun X, Wang L, Tang S and Yan C:
Downregulation of MicroRNA-145 caused by Hepatitis B virus X
protein promotes expression of CUL5 and contributes to pathogenesis
of Hepatitis B virus-associated hepatocellular carcinoma. Cell
Physiol Biochem. 37:1547–1559. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bertoli G, Cava C and Castiglioni I:
MicroRNAs: New biomarkers for diagnosis, prognosis, therapy
prediction and therapeutic tools for breast cancer. Theranostics.
5:1122–1143. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ruan K, Fang X and Ouyang G: MicroRNAs:
Novel regulators in the hallmarks of human cancer. Cancer Lett.
285:116–126. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pasqualini L, Bu H, Puhr M, Narisu N,
Rainer J, Schlick B, Schäfer G, Angelova M, Trajanoski Z, Börno ST,
et al: miR-22 and miR-29a are members of the androgen receptor
cistrome modulating LAMC1 and Mcl-1 in prostate cancer. Mol
Endocrinol. 29:1037–1054. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun T, Yang M, Chen S, Balk S, Pomerantz
M, Hsieh CL, Brown M, Lee GM and Kantoff PW: The altered expression
of MiR-221/−222 and MiR-23b/−27b is associated with the development
of human castration resistant prostate cancer. Prostate.
72:1093–1103. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fu X, Zhang W, Su Y, Lu L, Wang D and Wang
H: MicroRNA-103 suppresses tumor cell proliferation by targeting
PDCD10 in prostate cancer. Prostate. 76:543–551. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Aakula A, Leivonen SK, Hintsanen P,
Aittokallio T, Ceder Y, Børresen-Dale AL, Perälä M, Östling P and
Kallioniemi O: MicroRNA-135b regulates ERα, AR and HIF1AN and
affects breast and prostate cancer cell growth. Mol Oncol.
9:1287–1300. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tong SJ, Liu J, Wang X and Qu LX:
microRNA-181 promotes prostate cancer cell proliferation by
regulating DAX-1 expression. Exp Ther Med. 8:1296–1300. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sun J, Fan Z, Lu S, Yang J, Hao T and Huo
Q: MiR-192 suppresses the tumorigenicity of prostate cancer cells
by targeting and inhibiting nin one binding protein. Int J Mol Med.
37:485–492. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ren W, Li C, Duan W, Du S, Yang F, Zhou J
and Xing J: MicroRNA-613 represses prostate cancer cell
proliferation and invasion through targeting Frizzled7. Biochem
Biophys Res Commun. 469:633–638. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Saleiban A, Faxälv L, Claesson K, Jönsson
JI and Osman A: miR-20b regulates expression of
proteinase-activated receptor-1 (PAR-1) thrombin receptor in
melanoma cells. Pigment Cell Melanoma Res. 27:431–441. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xue TM, Tao LD, Zhang M, Xu GC, Zhang J
and Zhang PJ: miR-20b overexpression is predictive of poor
prognosis in gastric cancer. Onco Targets Ther. 8:1871–1876. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li MY and Hu XX: Meta-analysis of microRNA
expression profiling studies in human cervical cancer. Med Oncol.
32:5102015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou W, Shi G, Zhang Q, Wu Q, Li B and
Zhang Z: MicroRNA-20b promotes cell growth of breast cancer cells
partly via targeting phosphatase and tensin homologue (PTEN). Cell
Biosci. 4:622014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhu J, Chen L, Zou L, Yang P, Wu R, Mao Y,
Zhou H, Li R, Wang K, Wang W, et al: MiR-20b, −21 and −130b inhibit
PTEN expression resulting in B7-H1 over-expression in advanced
colorectal cancer. Hum Immunol. 75:348–353. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Landais S, Landry S, Legault P and Rassart
E: Oncogenic potential of the miR-106-363 cluster and its
implication in human T-cell leukemia. Cancer Res. 67:5699–5707.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yamaguchi T, Iijima T, Wakaume R,
Takahashi K, Matsumoto H, Nakano D, Nakayama Y, Mori T, Horiguchi S
and Miyaki M: Underexpression of miR-126 and miR-20b in hereditary
and nonhereditary colorectal tumors. Oncology. 87:58–66. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ahmad A, Ginnebaugh KR, Sethi S, Chen W,
Ali R, Mittal S and Sarkar FH: miR-20b is up-regulated in brain
metastases from primary breast cancers. Oncotarget. 6:12188–12195.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li J, Yen C, Liaw D, Podsypanina K, Bose
S, Wang SI, Puc J, Miliaresis C, Rodgers L, McCombie R, et al:
PTEN, a putative protein tyrosine phosphatase gene mutated in human
brain, breast, and prostate cancer. Science. 275:1943–1947. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huse JT, Brennan C, Hambardzumyan D, Wee
B, Pena J, Rouhanifard SH, Sohn-Lee C, le Sage C, Agami R, Tuschl T
and Holland EC: The PTEN-regulating microRNA miR-26a is amplified
in high-grade glioma and facilitates gliomagenesis in vivo. Genes
Dev. 23:1327–1337. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu W, Yang J, Feng X, Wang H, Ye S, Yang
P, Tan W, Wei G and Zhou Y: MicroRNA-32 (miR-32) regulates
phosphatase and tensin homologue (PTEN) expression and promotes
growth, migration, and invasion in colorectal carcinoma cells. Mol
Cancer. 12:302013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang H, Kong W, He L, Zhao JJ, O'Donnell
JD, Wang J, Wenham RM, Coppola D, Kruk PA, Nicosia SV and Cheng JQ:
MicroRNA expression profiling in human ovarian cancer: MiR-214
induces cell survival and cisplatin resistance by targeting PTEN.
Cancer Res. 68:425–433. 2008. View Article : Google Scholar : PubMed/NCBI
|