Introduction
Acute promyelocytic leukemia (APL) is a clonal
disease characterized by the dysregulated proliferation of abnormal
promyelocytes. A non-random chromosomal translocation breakpoint,
t(15;17)(q22;q21), occurs in almost every APL patient, which
produces the promyelocytic leukemia (PML)-retinoic acid receptor-α
(RAR-α) fusion gene, resulting in the expression of the PML-RARα
fusion protein (1). Other fusion
genes that cause APL also exist, including promyelocytic leukemia
zinc finger protein-RARα and nucleophosmin-RARα (2). The RARα gene is a target of chromosomal
rearrangements in all cases of acute promyelocytic leukemia (APL),
with each fusion protein containing the same portion of the RARα
polypeptide (which includes the DNA-binding and ligand-binding
domains (3), a fact that highlights
the key roles served by RARα in the development of APL (4,5).
RARα belongs to the nuclear receptor family and has
a proven role in the differentiation of myeloid cell lines; it is
also a transcriptional regulator that binds to specific retinoic
acid response elements (RAREs) present in the promoters of
RARα-target genes as a heterodimer with nuclear receptor retinoid X
receptor-α (RXRα) (6). Numerous
observations indicate that PML-RARα is a potent repressor of
promelocyte differentiation (7,8). PML-RARα
contributes to leukemogenesis by competitive binding of RAREs,
either as a homodimer or as a heterodimer with wild-type (WT) RARα
(9,10), thereby repressing gene transcription
essential for myeloid differentiation. In the past few years, the
development of curative approaches for APL has been the paradigm
(11), but the specific mechanistic
network of APL is still not fully understood (12–14). It is
therefore urgent to understand more about the molecular mechanism
of APL in order to develop improved, targeted drugs for the
treatment of APL.
Nuclear location signal (NLS)-RARα is a cleavage
variant of PML-RARα, formed on cleavage by neutrophilelastase (NE),
an early-myeloid-specific serine protease, whose production is
maximal in promyelocytes. NE-deficient animals were reported to be
protected from development of APL (15,16). The
NLS is a short sequence of amino acids that mediates the transport
of nuclear proteins into the nucleus. Typically, deletion of the
NLS disrupts nuclear import (17);
however, the addition of NLS to RARα alters the localization of
this fusion protein. Several reports indicated that protein
localization alters protein functions (18–20). We
hypothesized that the genes transcribed by RARα and NLS-RARα were
similar and perhaps that NLS-RARα had a gain of function compared
to WTRARα. NLS-RARα, following cleavage by NE, may have increased
function compared with RARα, possibly dependent on protein
localization and RARE-binding activity; similarities exist between
NLS-RARα and the PML-RARα fusion protein. The aim of the present
study was to examine the localization of NLS-RARα and the
functional changes that the addition of this signal mediated. These
findings may provide important information on the occurrence and
development of APL.
Materials and methods
Cell culture and plasmid
construction
The NB4, K562 and HEK293 cell lines were purchased
from the Shanghai Institutes for Biological Sciences (Shanghai,
China). The NB4 and K562 cell lines were cultured in RMPI-1640
medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.); the HEK 293 cell line was cultured in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.) containing 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.). All cells were cultured in an environment with
5% CO2 at 37°C, and the culture liquid was changed every day. The
culture liquid was changed every day. The RARα and RXRα expression
constructs used for in vitro translation were generated by
polymerase chain reaction (PCR) amplification of the corresponding
regions of complementary DNA (cDNA) in NB4 cells and cloned into
the expression vectors PCMV-HA and PCMV-mycvectors, respectively
(BioVector NTCC, Inc., Beijing, China). The thermocycling
conditions of RARα were: 95°C for 5 min, 98°C for 10 sec, 62°C for
35 sec, 72°C for 90 sec and 72°C for 5 min. The thermocycling
conditions of RXRα were: 95°C for 5 min, 98°C for 10 sec, 59.5°C
for 35 sec, 72°C for 90 sec and 72°C for 5 min. The DNA polymerase
was purchased from Takara Biotechnology Co., Ltd., Dalian, China.
The primers of RARα and RXRα were: RARα forward,
5′-CCGGTCGACAGATGTACGAGAGTGTAGAAG-3′ (EcoRV sequence
underlined) and reverse, 5′-CCGGATATCGTCACATGGTCGGTAG-3′
(SalI sequence underlined); and RXRα forward,
5′-CCTGAATTCATGGACACCAAACATTTCCTGC-3′ (EcoRI sequence
underlined) and reverse, 5′-CCTGCGGCCGCCTAAGTCATTTGGTGCGGC-3′
(NotI sequence underlined). All primers were synthesized by
Takara Biotechnology Co., Ltd. NLS-RARα was amplified using
pcDNA3.1-PML-RARα as a template, and then subcloned into PCMV-HA
vectors.
RNA extraction and plasmid
construction
Total RNA from NB4 cells was extracted using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific Inc.) and reverse
transcribed using a PrimeScript RT reagent kit (Takara
Biotechnology Co., Ltd.). The PCR reaction mixture contained 2 µl
cDNA, 1 µl (20 µM) of each primer and 25 µl PremixTaq enzyme
(Takara Biotechnology Co., Ltd.), with sterile water added up to a
final volume of 50 µl. The reaction conditions were:
Pre-denaturation at 95°C for 5 min, and then 35 cycles of
denaturation at 98°C for 10 sec, annealing at 62°C (for
RARα)/59.5°C (for RXRα) for 90 sec, extension at 72°C for 30 sec.
The PCR products were separated on 1.5% agarose gels and stained
with ethidium bromide. The fragments were then recycled from the
gel and for RARα, the fragment and a PCMV-HA empty vector was
digested with EcoRV and SalI (Takara Biotechnology
Co., Ltd.); for RXRα, the fragment and PCMV-myc empty vector was
digested with EcoRI and NotI (Takara Biotechnology
Co., Ltd.), and the digested fragments and vector were ligated with
T4 ligase for 6 h. The ligation product was then transformed into
competent Escherichia coli. The transformation process was
as follows: The ligation product was added to competent E.
coli DH5α, which were placed in an ice bath for 30 min, heat
shocked immediately for 90 sec at 42°C, placed into an ice bath for
2 min, and then the present study added 800 µl antibiotic-free
lysogeny broth. After this step, the broth underwent shaking for
30–60 min at 37°C and 220 rpm; centrifugation for 5 min at 1,0006.2
× g produced ~100 µl supernatant. Following the resuspension of the
precipitate with the supernatant, and the coating of the LB plates
with 50 µg/mlkanamycin, the plates were placed in a culture
incubator at 37°C. Transformed colonies were selected using the
kanamycin and, 12 h later, were shaken in 5 ml lysogenybroth at
37°C. After 12 h, the plasmids were extracted using a
PureYield™ Plasmid Maxiprep System kit purchased from
Promega Corporation (Madison, WI, USA) and sent to Takara
Biotechnology Co., Ltd. for sequencing.
Transfection and preparation of the
cell extracts
A total of 1×107 HEK293 cells were seeded
into10-cm dishes. Prior to transfection, the cells were cultured
with DMEM for 1 h then transiently transfected with
PCMV-HA-NLS-RARα (8 µg) and PCMV-myc-RXRα (8 µg) with
Lipofectamine® 2000 (30 µl; Invitrogen; Thermo Fisher
Scientific, Inc.) when cell confluence reached over 80%. At 6 h
after transfection, DMEM was replaced with DMEM supplemented with
10% fetal bovine serum. After 2 days, the cells were collected into
Eppendorf tubes and centrifuged at 96.8 × g for 5 min at 4°C,
washed three times with cold PBS and suspended in
radioimmunoprecipitation assay buffer and phenylmethylsulfonyl
fluoride (at a ratio of 100:1) on ice. This lysis buffer contained
cell extraction reagents A and B that used low osmotic pressure
conditions to lyse cell membranes, releasing cytoplasmic protein.
The extraction of cell proteins was achieved using a Protein
Extraction kit purchased from the Beyotime Institute of
Biotechnology (Haimen, China) and was performed according to the
manufacturer's protocol. Nuclear proteins were obtained using a
nuclear protein extraction reagent kit (P0027) purchased from the
Beyotime Institute of Biotechnology in accordance with the
manufacturer's protocol. The cells were then centrifuged at 15,000
× g for 15 min at 37°C, the supernatant was collected and the
protein concentration was measured using a BCA assay kit (P0009)
purchased from the Beyotime Institute of Biotechnology in
accordance with the manufacturer's protocol.
Co-immunoprecipitation (Co-IP) and
western blot analysis
A total of 1×107 cells were seeded in
10-cm dishes and 48 h after transfection, cells were lysed using IP
lysis buffer from an NP-40 kit (P0013F) purchased from the Beyotime
Institute of Biotechnology. The cell extracts were then incubated
with the appropriate antibodies, if adding anti-RARα antibody at a
given step, the present study would use anti-RXRα antibody in the
following step and vice versa, or non-specific controls.
Immunocomplexes were recovered by protein A/G agarose beads and
resolved by electrophoresis. Cell protein (40 µg) from each group
was separated by 10% SDS-PAGE and then transferred onto a
polyvinylidene difluoride (PVDF) membrane. These membranes were
incubated for 3 h at room temperature in 5% skimmed milk [diluted
with Tris-buffered-saline with Tween-20 (TBST)] for membrane
blocking. The primary antibodies used were anti-RARα rabbit
polyclonal antibody (cat. no. sc-551); anti-RXRα (cat. no.
sc-46659) mouse monoclonal antibody (both diluted 1:1,000; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA). Membranes were
incubated with primary antibodies overnight at 4°C and then
incubated with secondary antibody (goat anti-rabbit antibody;
1:2,000; cat. no. 127760; Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd., Beijing, China) for1 h at room
temperature. After washing three times with TBST, immunoreactive
complexes were visualized using ECL Chemiluminescence system
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). β-actin served as
an internal positive control.
Immunofluorescence (IF)
microscopy
Transfected HEK293 cells were cytospun onto slides,
fixed in 4% formaldehyde for 20 min, washed with PBS containing
0.1% Tween-20, permeabilized with 0.1% Triton X-100 solution for 10
min and washed three times with PBS. Cells were then blocked with
10% goat serum for 30 min at room temperature. After blocking,
cells were immunolabelled with anti-RARα 1:200 diluted rabbit
polyclonal antibodies (cat. no. sc-551) and anti-RXRα 1:200 diluted
mouse monoclonal antibodies (cat. no. sc-46659) raised in the lab
at 4°C overnight, washed three times in PBS and then relabelled
with fluorescein isothiocyanate (FITC)-coupled goat anti-rabbit
(cat. no. ZF-0314; 1:200; Zhongshan Golden bridge Biotechnology
Co., Ltd.) and tetramethylrhodamine (TRITC)-coupled goat anti-mouse
antibodies (cat. no. ZF-0313; 1:200; Zhongshan Golden bridge
Biotechnology Co., Ltd.) at room temperature for 1 h. For nucleus
staining, immunolabeled cells were incubated with DAPI or propidium
iodide. A laser scanning confocal microscope was used to observe
the cells.
Electrophoretic mobility shift assay
(EMSA) and reporter gene assays
For the EMSA, the nuclear extract was obtained from
HEK293 cells transiently transfected with PCMV-HA and PCMV-myc
expression vector(s) containing NLS-RARα and RXRα by washing cells
in each group with ice-cold phosphate-buffered saline and lysing
them in RIPA-1640 solution (Beyotime Institute of Biotechnology)
containing a protease inhibitor cocktail for 20 min at 4°C. After
24 h, 10−6 M all-trans retinoic acid (ATRA) was added to
the dishes. RARα and RXRα genes were presented to the variety of
the consensus binding sequences. Biotin-labeled DNA probes were
synthesized by Takara Biotechnology Co., Ltd. and mixed with
nuclear extract in the binding buffer as aforementioned, with 0.02
µg/µl poly (deoxyinosinic-deoxycytidylic) acid at room temperature.
DNA-protein complexes and free DNA were separated on a 3.75%
polyacrylamide gel. HEK293 cells were transiently transfected with
luciferase reporter plasmids [RARE-2-Tk-Luc and DR5-(or 8)-Tk-Luc]
and pCMV-HA-NLS-RARα, pCMV-HA-RARα using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). The luciferase
reporter plasmids were donated by Professor Huguesde The (Institut
Universitaire d'Hématologie, Université Paris-Diderot, Paris,
France). After 48 h, cells were lysed and normalized luciferase
activities were determined.
Statistical analysis
Independent sample t-test was used to compare the
means between two groups. All statistical analyses were performed
using the SPSS 18.0 software package (SPSS, Inc., Chicago, IL,
USA). All data and results presented are representative of, or
calculated from, at least three independentexperiments. P<0.05
was considered to indicate a statistically significant
difference.
Results
Plasmid construction
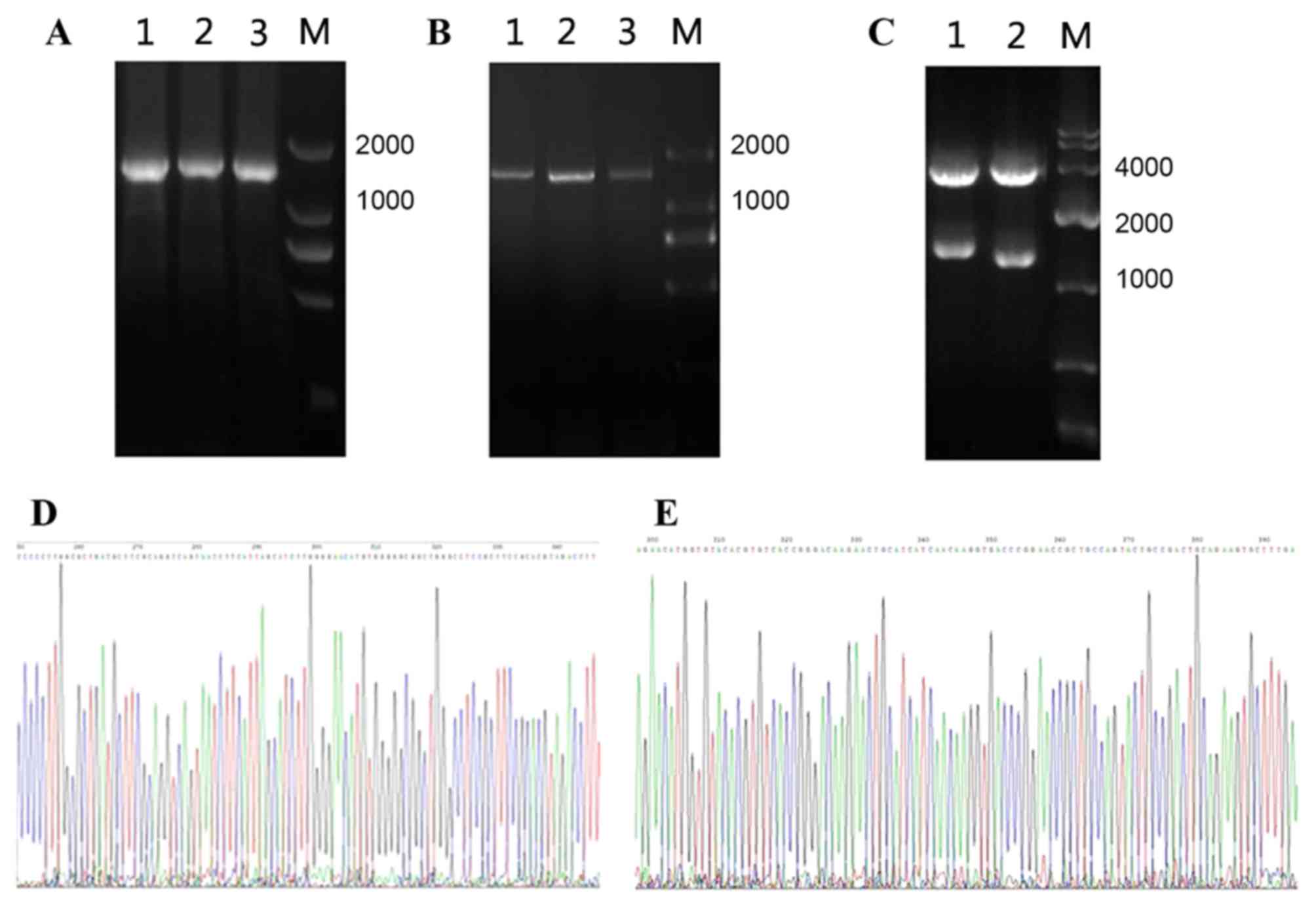
Firstly, RNA was extracted from APL cell line NB4,
which was reverse transcribed into cDNA and then amplified by PCR
(Fig. 1A and B). The lengths of RARα,
RXRα were 1,449 and 1,389 bp respectively. Next, the fragments were
digested with the appropriate restriction endonucleases. Sequencing
results verified that RARα and RXRα expression plasmids had been
constructed (Fig. 1).
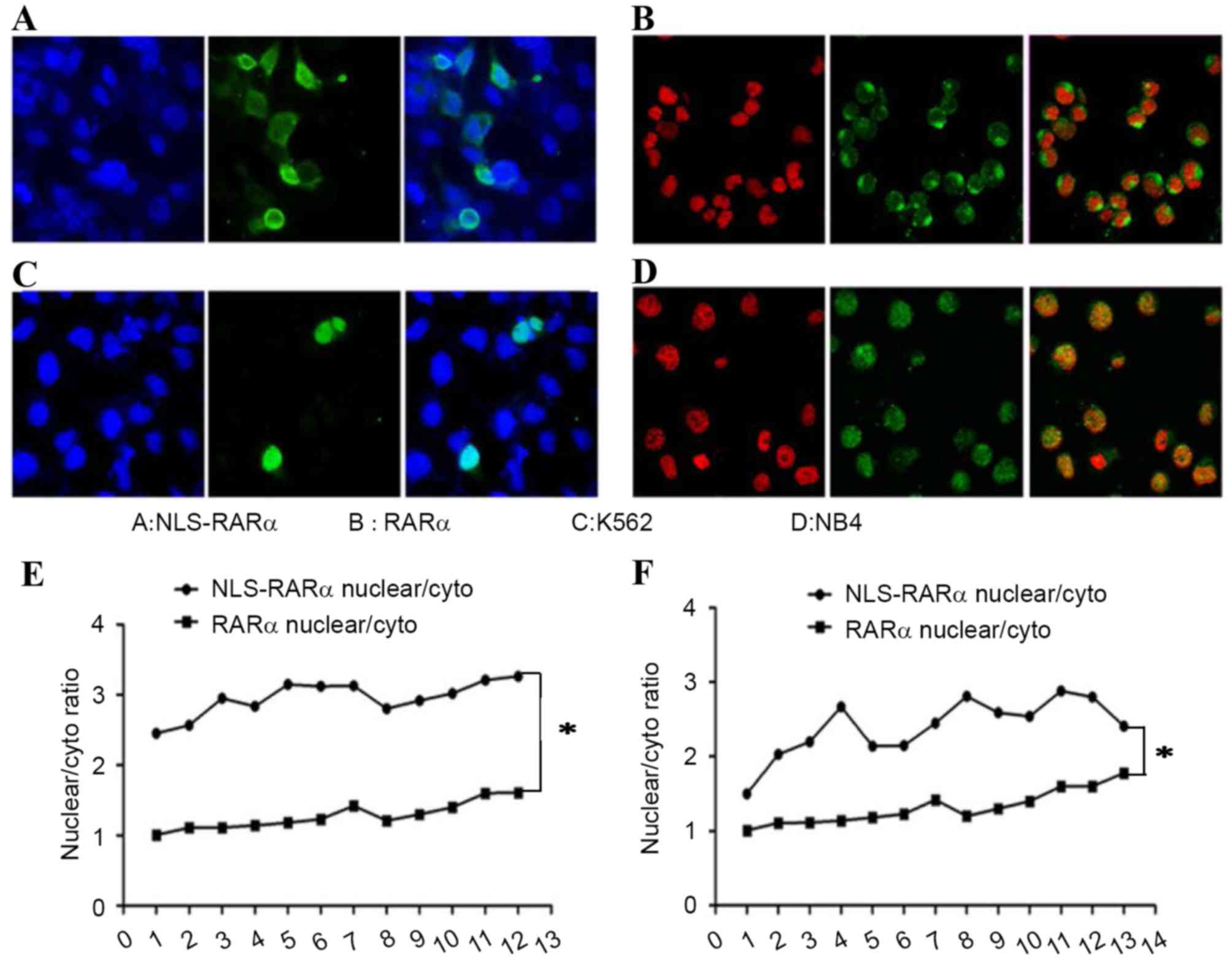
IF result of RARα and NLS-RARα in
different cell lines
RARα belongs to a family of nuclear receptors, and
was revealed to localize to the nucleus and the cytoplasm. Although
NLS-RARα localized to the nucleus and cytoplasm, the cytoplasmic
level was far lower compared with the nuclear levels (Fig. 2A and B). Similar results were also
obtained for leukemia cell lines (Fig. 2C
and D). Calculation of the nuclear fluorescence intensity
revealed that the level of NLS-RARα was increased compared with
that of RARα, and the difference was statistically significant
(P<0.05; Fig. 2E and F).
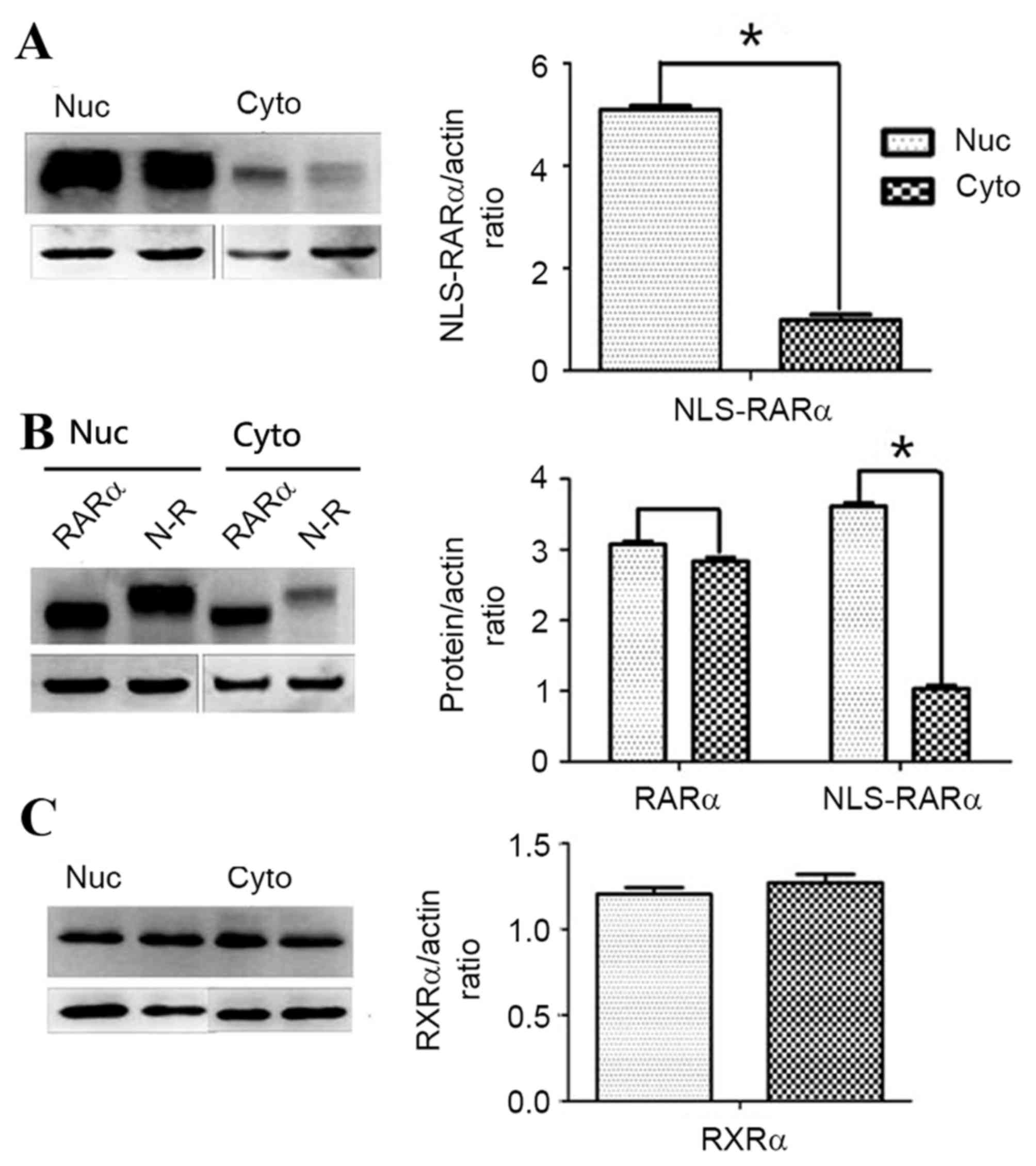
Western blot analysis of RARα and
NLS-RARα
Western blot analysis confirmed the results of IF;
owing to the presence of the NLS, NLS-RARα localized to the
nucleus. There was a significantly increased level NLS-RARα present
in the nucleus compared with that in the cytoplasm (P, 0.05;
Fig. 3A and B). RXRα, like RARα,
belongs to the nuclear receptor family, but was expressed equally
in the nucleus and cytoplasm (Fig.
3C). Western blot analysis revealed that, the distribution of
RXRα was almost equal in the nucleus and cytoplasm.
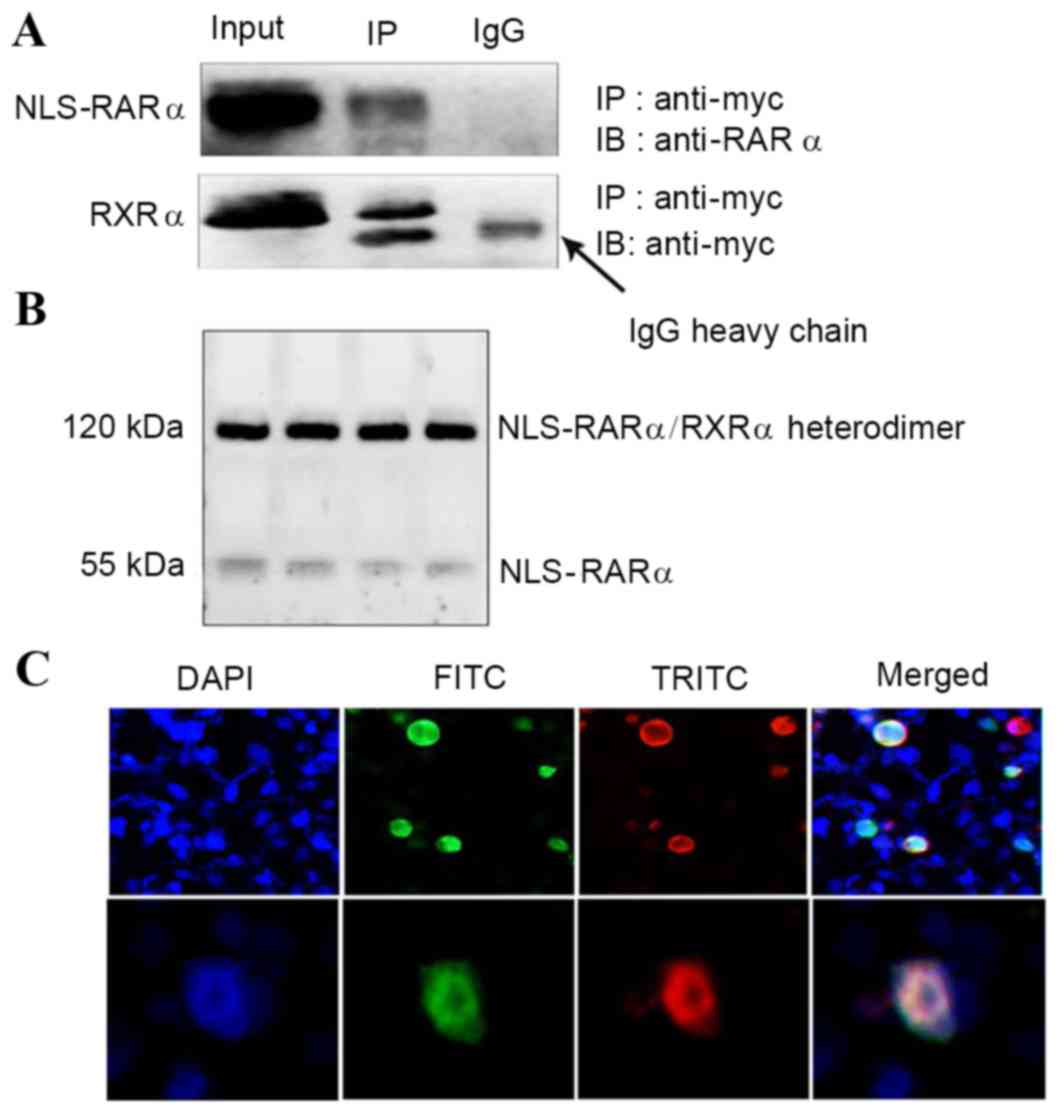
Interaction between NLS-RARα and
RXRα
The retinoid signal is transduced by two families of
nuclear receptors, RARs and RXRs, which formed RXR/RAR
heterodimers. This means that RARα and RXRα interact with each
other (21). However, whether
NLS-RARα and RXRα could interact together was unclear.
Co-imunopreciptation results revealed that NLS-RARα and RXRα did
interact (Fig. 4A). Further
experiments indicated that NLS-RARα and RXRα heterodimerized
(Fig. 4B) and IF demonstrated that
the two proteins co-localized (Fig.
4C).
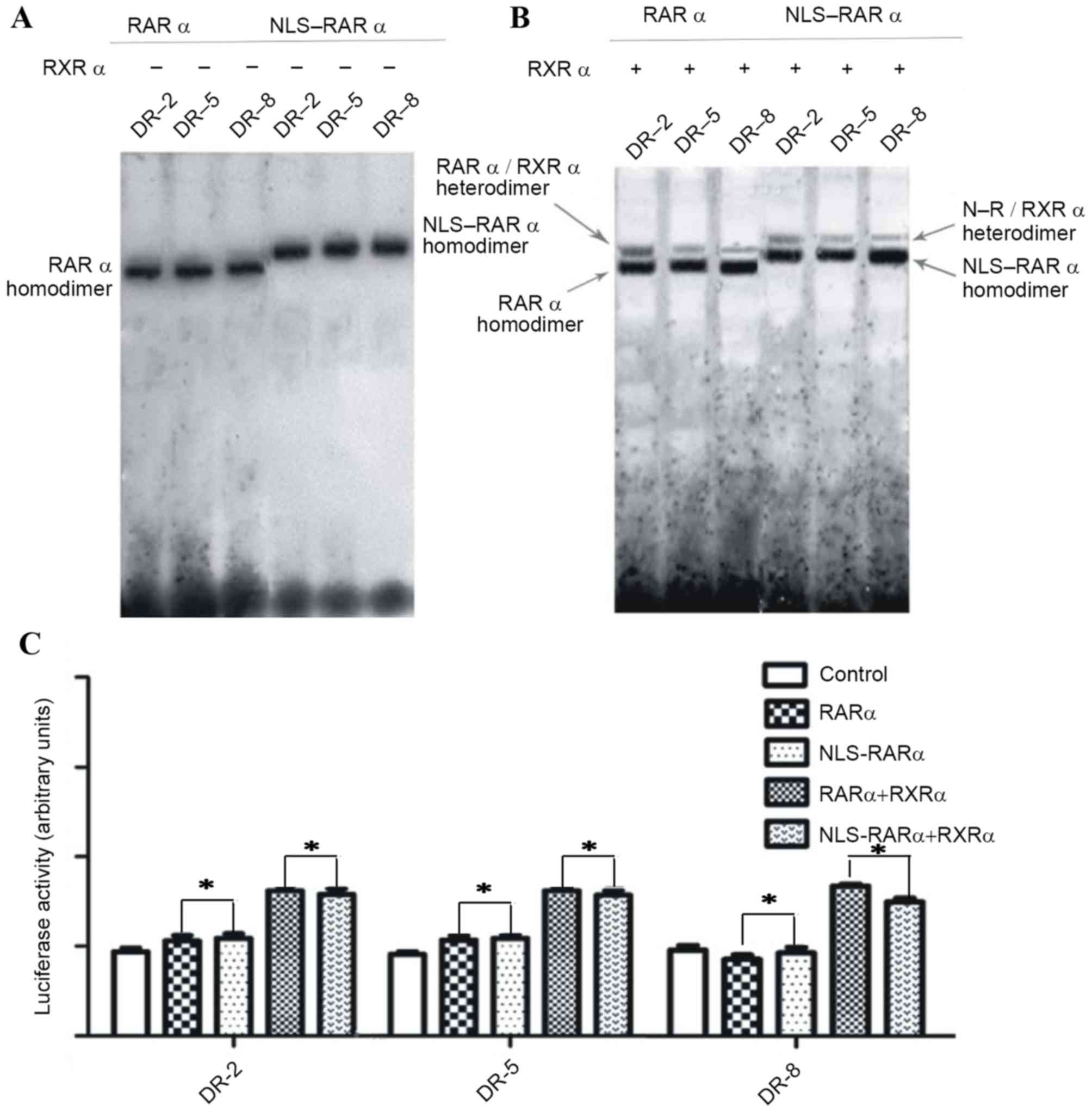
NLS-RARα may bind and regulate the
activity of RAREs
Previous studies demonstrated that RARα and RXRα
could bind to specific DNA sequences or RAREs as asymmetric,
oriented heterodimers in the presence of ATRA (21). Typically, RAREs are composed of two
direct repeats of a core hexameric motif, PuG (G/T)TCA (20). Classical RAREs are 2, 5 and
8-bp-spaced direct repeats (referred to as DR-2, DR-5 and DR-8,
respectively), and in the present study, WTRARα was revealed to
bind DR-2, DR-5 and DR-8 in the presence of ATRA. NLS-RARα could
also bind DR-2, DR-5 and DR-8 (Fig. 5A
and B). The binding of NLS-RARα to DR-2 and DR-5 was a little
weaker than WTRARα, but this was not a statistically significant
difference (P>0.05). Furthermore, results of a luciferase
reporter assay revealed that NLS-RARα bind the luciferase reporter
plasmids and mediate their activity (P<0.05; Fig. 5C).
Discussion
APL is characterized by the aberrant proliferation
of myelocyte precursor cells, whose maturation is blocked at the
promyelocyte stage of granulocytic differentiation (22). The molecular marker of APL is the
t(15;17)(q22;q21) translocation, which yields an aberrant PML-RARα
fusion protein (23). In the absence
of appropriate and prompt treatment, disseminated intravascular
coagulation, owing to the secretion of plasminogen activators and
lysosomal enzymes by leukemia cells, causes serious bleeding
(24). In 1985, Huang et al
(25) demonstrated that ATRA could
induce complete remission, initiating a new era of APL therapy.
However, a number of questions remain unanswered concerning APL.
For example, the exact roles that alterations in RARα serve in the
transformation process and the associated molecular mechanisms
remain unknown. In the present study, the NLS-RARα fusion protein
was revealed to be potentially involved in the pathogenesis of APL.
NLS-RARα has previously been demonstrated to promote proliferation
and inhibit differentiation in the APL HL-60 cell line (26). The aim of the present study was to
explain the localization of NLS-RARα and assess its role as a
variant transcriptional factor.
RARα belongs to nuclear receptor family and
heterodimerizes with RXRα (27). The
RARα/RXRα heterodimer recognizes the RAREs DR-2 and DR-5 (25). RAREs can be identified in the promoter
regions of most RA-responsive genes. In the absence of agonist,
RARα represses the transcription of its targets via the recruitment
of several co-repressor proteins, including nuclear receptor
co-repressor 2, which maintains the chromatin in an inactive state
(13,28,29).
In the present study, NLS-RARα was observed to
localize to the nucleus to a greater degree than WTRARα (Fig. 1). Changes in protein localization have
been demonstrated to alter protein function, as the internal
environment of the nucleus and cytoplasm are markedly different
(17). The present study demonstrated
the presence of an interaction between NLS-RARα and RXRα, and that
NLS-RARα and RXRα could form heterodimers, which enabled them to
bind RAREs. A previous study revealed that the PML-RARα fusion
protein could competitively bind RXRα in APL, forming the
PML-RARα-RXRα oligomer, which could bind an extended repertoire of
response elements (8). In the present
study, the transcriptional activities of the NLS-RARα variant were
assessed; results from the EMSA and luciferase reporter assay
indicated that the transcriptional activity of NLS-RARα was the
same as WTRARα, as NLS-RARα could also bind 2-DR, 5-DR and 8-DR
target probes, pointing to relaxed structural constraints for DNA
binding and clarifying the function of this oncoprotein.
RARα binds to several genes involved in cell cycle
regulation and transformation, including tumor necrosis factor
superfamily member 13 (aproliferation-inducing ligand) (30), cyclin D1 and fibroblast growth factor
18 (31). The NLS-RARα fusion protein
may interrupt the interaction between RARα and its target proteins
through its stronger binding affinity to RAREs. How NLS-RARα
competitively binds to RARα target genes and proteins requires
further study.
In conclusion, the variant protein NLS-RARα promoted
proliferation and inhibited differentiation in leukemia cells, and
its localization was altered by the addition of NLS. As RARα is a
transcriptional factor, the transcriptional activities of NLS-RARα
were assessed and revealed to be similar to WTRARα. Therefore
NLS-RARα may serve a role in APL pathogenesis by competitively
binding RARα-target proteins as a variant transcriptional factor;
however, further research is required to identify the proteins
involved. The present study illuminated the function of NLS-RARα in
APL, the molecular mechanism network of APL formation, and may aid
with a greater understanding of APL.
Acknowledgements
The authors would like to thank Professor de The
(Institut Universitaire d'Hématologie, Université Paris-Diderot,
Paris, France) for providing the luciferase report plasmid. This
study was supported by grants from the National Natural Science
Foundation of China (grant no. 81171658) and the Natural Science
Foundation Project of Chongqing Science and Technology Commission
(grant no. 2011BA5037).
References
|
1
|
de Thé H, Le Bras M and
Lallemand-Breitenbach V: The cell biology of disease: Acute
promyelocytic leukemia, arsenic, and PML bodies. J Cell Biol.
198:11–21. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zelent A, Guidez F, Melnick A, Waxman S
and Licht JD: Translocations of the RARalpha gene in acute
promyelocytic leukemia. Oncogene. 20:7186–7203. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Melnick A and Licht JD: Deconstructing a
disease: RARalpha, its fusion partners and their roles in the
pathogenesis of acute promyelocytic leukemia. Blood. 93:3167–3215.
1999.PubMed/NCBI
|
|
4
|
Chang KS, Stass SA, Chu DT, Deaven LL,
Trujillo JM and Freireich EJ: Characterization of a fusion cDNA
(RARA/myl) transcribed from the t(15;17) translocation breakpoint
in acute promyelocytic leukemia. Mol Cell Biol. 12:800–810. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou GB, Chen SJ and Chen Z: Acute
promyelocytic leukemia: A model of molecular target based therapy.
Hematology. 10 Suppl 1:S270–S280. 2005. View Article : Google Scholar
|
|
6
|
Raelson JV, Nervi C, Rosenauer A,
Benedetti L, Monczak Y, Pearson M, Pelicci PG and Miller WH Jr: The
PML/RAR alpha Oncoprotein is a direct molecular target of retinoic
acidin acute promyelocytic leukemia cells. Blood. 88:2826–2832.
1996.PubMed/NCBI
|
|
7
|
Zhou J, Pérès L, Honoré N, Nasr R, Zhu J
and de Thé H: Dimerization-induced corepressor binding and relaxed
DNA-binding specificity are critical for PML/RARA-induced
immortalization. Proc Natl Acad Sci USA. 103:pp. 9238–9243. 2006,
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kamashev D, Vitoux D and De Thé H:
PML-RARA-RXR oligomers mediate retinoid and rexinoid/cAMP
cross-talk in acute promyelocytic leukemia cell differentiation. J
Exp Med. 199:1163–1174. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu J, Nasr R, Pérès L, Riaucoux-Lormière
F, Honoré N, Berthier C, Kamashev D, Zhou J, Vitoux D, Lavau C and
de Thé H: RXR is an essential component of the oncogenic PML/RARA
complex in vivo. Cancer Cell. 12:23–35. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vitaliano-Prunier A, Halftermeyer J,
Ablain J, de Reynies A, Peres L, Le Bras M, Metzger D and de Thé H:
Clearance of PML/RARA-bound promoters suffice to initiate APL
differentiation. Blood. 124:3772–3780.. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ablain J and de The H: Revisiting the
differentiation paradigm in acute promyelocytic leukemia. Blood.
117:5795–5802. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang Y, Qiu J, Chen G and Dong S:
Coiled-coil domain of PML is essential for the aberrant dynamics of
PML-RARalpha, resulting in sequestration and decreased mobility of
SMRT. Biochem Biophys Res Commun. 365:258–265. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
de Thé H and Chen Z: Acute promyelocytic
leukemia: Novel insights into the mechanisms of cure. Nat Rev
Cancer. 10:775–783. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ablain J and de Thé H: Retinoic acid
signaling in cancer: The parable of acute promyelocytic leukemia.
Int J Cancer. 135:2262–2272. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lane AA and Ley TJ: Neutrophil elastase
cleaves PML-RARalpha and is important for the development of acute
promyelocytic leukemia in mice. 115:305–318. 2003.
|
|
16
|
Hayakawa F and Privalsky ML:
Phosphorylation of PML by mitogen-activated protein kinases plays a
key role in arsenic trioxide-mediated apoptosis. Cancer Cell.
5:389–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cokol M, Nair R and Rost B: Finding
nuclear localization signals. EMBO Rep. 1:411–415. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang XW, Yan XJ, Zhou ZR, Yang FF, Wu ZY,
Sun HB, Liang WX, Song AX, Lallemand-Breitenbach V, Jeanne M, et
al: Arsenic trioxide controls the fate of the PML-RARalpha
oncoprotein by directly binding PML. Science. 328:240–243.. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu TT, Chen C, Chen SM, Xu Y, Wang Y, Chen
Z, Wang F, Xiao BK and Tao ZZ: Nuclear translocation of telomerase
reverse transcriptase is a critical process in lymphatic metastasis
of nasopharyngeal carcinoma. Oncol Lett. 9:265–269. 2015.PubMed/NCBI
|
|
20
|
Sánchez-Quesada C, López-Biedma A and
Gaforio JJ: The differential localization of a methyl group confers
a different anti-breast cancer activity to two triterpenes present
in olives. Food Funct. 6:249–256. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Altucci L, Leibowitz MD, Ogilvie KM, de
Lera AR and Gronemeyer H: RAR and RXR modulation in cancer and
metabolic disease. Nat Rev Drug Discov. 6:793–810. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
McCulloch D, Brown C and Iland H: Retinoic
acid and arsenic trioxide in the treatment of acute promyelocytic
leukemia: Current perspectives. Onco Targets Ther. 10:1585–1601.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kakizuka A, Miller WH Jr, Umesono K,
Warrell RP Jr, Frankel SR, Murty VV, Dmitrovsky E and Evans RM:
Chromosomal translocation t(15;17) in human acute promyelocytic
leukemia fuses RAR alpha with a novel putative transcription
factor, PML. Cell. 66:663–674. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Choudhry A and DeLoughery TG: Bleeding and
thrombosis in acute promyelocytic leukemia. Am J Hematol.
87:596–603. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang ME, Ye YC, Chen SR, Chai JR, Lu JX,
Zhoa L, Gu LJ and Wang ZY: Use of all trans retinoic acid in the
treatment of acute promyelocytic leukemia. Blood. 72:567–572.
1988.PubMed/NCBI
|
|
26
|
Hu XX, Zhong L, Zhang X, Gao YM and Liu
BZ: NLS-RARα promotes proliferation and inhibits differentiation in
HL-60 cells. Int J Med Sci. 11:247–254. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
de Thé H, Lavau C, Marchio A, Chomienne C,
Degos L and Dejean A: The PML-RAR alpha fusion mRNA generated by
the t(15;17) translocation in acute promyelocytic leukemia encodes
a functionally altered RAR. Cell. 66:675–684. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zelent A, Guidez F, Melnick A, Waxman S
and Licht JD: Translocations of the RARalpha gene in acute
promyelocytic leukemia. Oncogene. 20:7186–7203. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bastien J and Rochette-Egly C: Nuclear
retinoid receptors and the transcription of retinoid-target genes.
Gene. 328:1–16. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Planelles L, Medema JP, Hahne M and
Hardenberg G: The expanding role of APRIL in cancer and immunity.
Curr Mol Med. 8:829–844. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Delacroix L, Moutier E, Altobelli G,
Legras S, Poch O, Choukrallah MA, Bertin I, Jost B and Davidson I:
Cell-specific interaction of retinoic acid receptors with target
genes in mouse embryonic fibroblasts and embryonic stem cells. Mol
Cell Biol. 30:231–244. 2010. View Article : Google Scholar : PubMed/NCBI
|