Introduction
Cervical cancer is one of the most common types of
gynecological malignancy with high morbidity and mortality, and
seriously affects the living quality of females worldwide (1). The pathogenesis of cervical cancer
remains unknown and may be connected with numerous factors (e.g.
recurrent genetic alterations, microenvironment and lifestyle)
(2,3).
According to the latest Cancer Statistics between 2000 and 2011 in
China (4), there were ~98,900 new
cases of cervical cancer, with >30,500 mortalities from the
disease.
Angiogenesis is defined as the formation of new
blood vessels from pre-existing vessels, and is essential for the
delivery of nutrients and oxygen to cells that are distant from
existing blood vessels (5).
Physiological angiogenesis is a sequence of cellular events
comprising vascular initiation, formation, maturation, remodeling
and regression, which are regulated to supply tissue requirements
(5–7).
It has been demonstrated that angiogenesis is essential for the
growth, metastasis and development of cancer (8). The biochemical stimulation of
angiogenesis is induced by angiogenic factors, including vascular
endothelial growth factor (VEGF), fibroblast growth factor (FGF),
interleukin (IL-)8, angiopoietins, matrix metalloproteinases
(MMPs), cadherins and integrins (9–11).
Thymic stromal lymphopoietin (TSLP) is a cytokine
produced by stromal cells, epithelial cells, fibroblasts,
keratinocytes and basophils (12–14). TSLP
triggers T helper 2 cell cytokines, including thymus and activation
regulated chemokine, and is associated with airway inflammatory
disease, allergic responses, immunoglobulin E production and
eosinophilia (12,13,15). The
TSLP receptor (TSLPR) is a typical heterodimeric cytokine receptor
consisting of a TSLP binding subunit (TSLPRα) and the α-subunit of
the IL-7 receptor (IL-7Rα) (16,17). Our
previous study demonstrated that TSLP secreted by cervical cancer
cells promotes angiogenesis (18). In
addition, an increased level of TSLP in cancer lesions, mediated by
hypoxia, was a regulator of the progression of cervical cancer, and
functioned by recruiting and enabling tumor-associated eosinophils
(EOS) to promote the growth of the cervical cancer cells (19). However, it remains unknown whether and
how TSLP, derived from cervical cells, regulates angiogenesis by
modulating the crosstalk between cervical cancer cells and EOS.
Therefore, the aim of the present study was to investigate the
effect of TSLP on the secretion of angiogenic factors VEGF, FGF and
IL-8 from EOS, and cervical cancer cells, and on the angiogenesis
of human umbilical vein endothelial cells (HUVECs) in
vitro.
Materials and methods
Cell culture
Cervical epidermoid carcinoma HeLa and CasKi cells
(Type Culture Collection of the Chinese Academy of Sciences,
Shanghai, China) were cultured in RPMI-1640 medium (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 5%
fetal bovine serum (FBS; Hyclone; GE Healthcare Life Sciences,
Logan, UT, USA).
HL-60 cells, a promyelocytic leukemia cell line,
were purchased from the Type Culture Collection of the Chinese
Academy of Sciences. According to a previous study (19), the HL-60-eosinophils (HL-60E) were
generated by incubating HL-60 cells for 2 months at pH 7.7 in
RPMI-1640 medium containing 25 mM N-(2-hydroxyethyl)
piperazine-N'-3-propane-sulfonic acid (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) and subsequently culturing for 7 days in
RPMI-1640 medium with 0.5 mM butyric acid (Sigma-Aldrich; Merck
KGaA). HUVECs (Type Culture Collection of the Chinese Academy of
Sciences) were maintained as monolayers in Dulbecco's modified
Eagle medium/F-12 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS. These cells were incubated at 37°C in a
humidified atmosphere containing 5% CO2.
Co-culture of HL-60E with HeLa or
CasKi cells
HeLa and CasKi cells (2×105 cells/well)
were cultured with HL-60E cells (2×105 cells/well) in a
humidified incubator with 5% CO2 at 37°C for 48 h and
subsequently incubated with or without recombinant human TSLP
protein (rhTSLP; 100 ng/ml; R&D Systems, Inc., Minneapolis, MN,
USA), anti-human TSLP or anti-TSLPR neutralizing antibodies (α-TSLP
or α-TSLPR; 5 µg/ml; R&D Systems, Inc.) in a humidified
incubator containing 5% CO2 at 37°C for 48 h, and 1%
PBS, as a negative control. Supernatants of the co-culture system
were selected for ELISA analysis or tube formation assay. Each
experiment was performed in six parallel wells and repeated five
times.
ELISA
HL-60E cells (2×105 cells/well) were
seeded in 24-well plates and treated with rhTSLP (1, 10 or 100
ng/ml) or 1% PBS (negative control) at 37°C for 48 h. Subsequently,
the cell culture supernatants were harvested, centrifuged at 300 ×
g at 4°C for 10 min to remove cellular debris and stored at −80°C
until use. The secretion level of IL-8, VEGF and FGF was determined
using human IL-8 (cat. no. EH005-96), VEGF (cat. no. EH015-96) and
FGF (cat. no. EH022-96) ELISA kits (Shanghai ExCell Biology, Inc.,
Shanghai, China), respectively, according to the manufacturer's
protocol. In addition, the protein concentration of the control, 1,
10 and 100 ng/ml rhTSLP groups was determined and the cytokine
level of each group was calculated as the ratio of the cytokine
concentration of the supernatant to the protein concentration. The
experiment was repeated three times. In addition, the level of
IL-8, VEGF and FGF in the supernatants of the co-culture system
with HeLa or CasKi cells as described above was determined using
ELISA. Each experiment was performed in six parallel wells and
repeated five times.
Tube formation assay (Matrigel
assay)
HUVECs (2×104 cells/well) were seeded in
96-well plates on growth factor-reduced Matrigel in RPMI-1640
medium (5% FBS) with recombinant human VEGF protein (10 ng/ml; a
positive control; R&D Systems, Inc.). Subsequently, HUVECs were
stimulated with the supernatants from HeLa (following treatment
with or without α-TSLP or α-TSLPR as aforementioned), CasKi
(following treatment with or without α-TSLP or α-TSLPR as
aforementioned), HL-60E, co-culture of HL-60E and HeLa, or
co-culture of HL-60E and CasKi cells; the RPMI-1640 medium with 5%
FBS was used as a negative control. After 16 h, in five randomly
chosen optical fields with light microscopy, capillary-like tube
formation was quantified by counting the numbers of
junctions/enclosed circles (original magnification, ×100). Each
experiment was performed in six parallel wells and repeated five
times.
Statistical analysis
All values are presented as the mean ± standard
error of the mean. The data were analyzed using GraphPad Prism
(version 5; GraphPad Software, Inc., La Jolla, CA, USA) by one-way
ANOVA using Tukey's post-hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
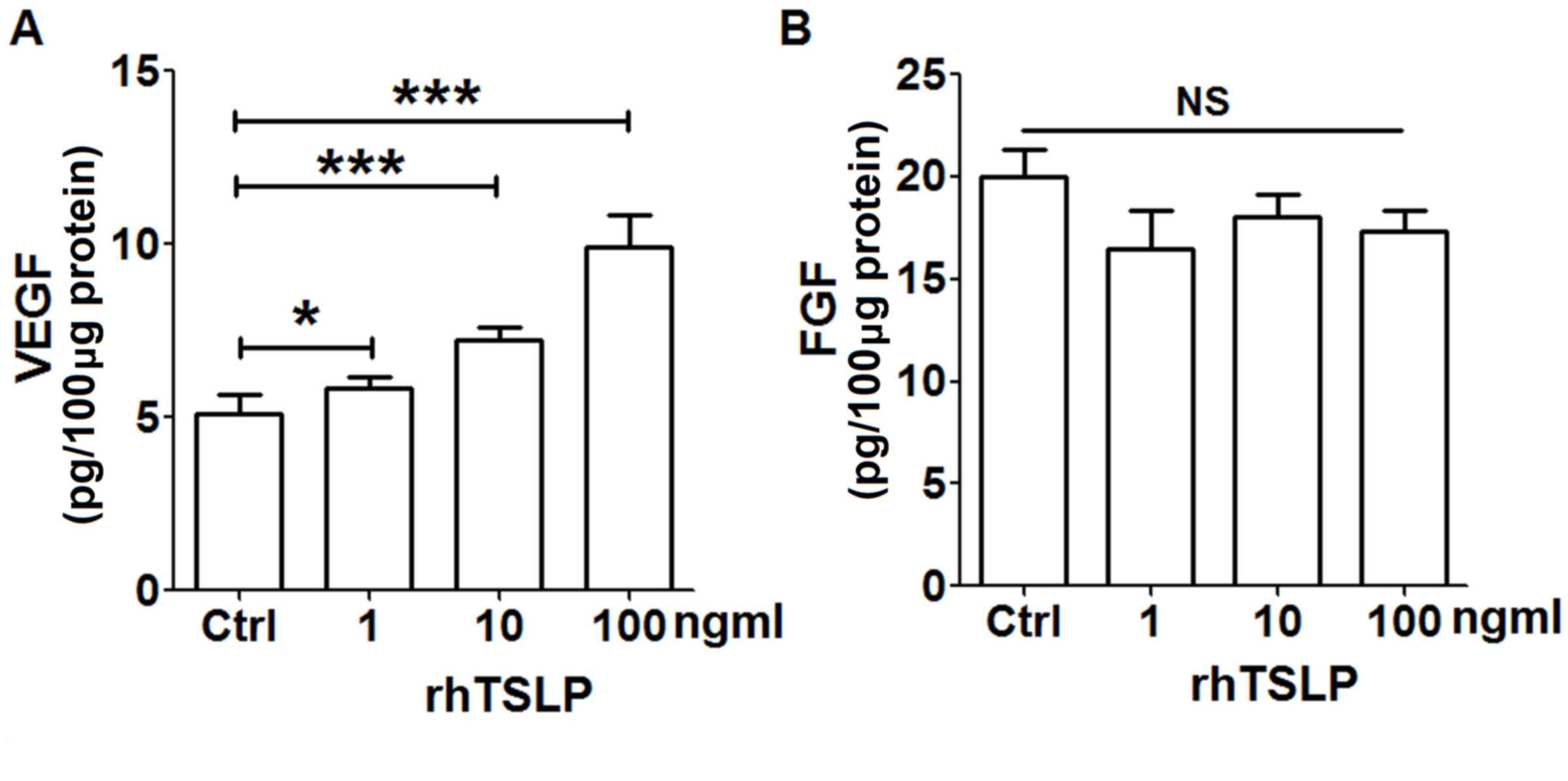
rhTSLP promotes VEGF secretion of
HL-60E cells
In order to explore the secretion of TSLP on
tumor-associated angiogenic factors from EOS, HL-60E cells were
treated with different concentrations of rhTSLP protein for 48 h.
As presented in Fig. 1, rhTSLP
stimulated the production of VEGF from HL-60E cells in a
dose-dependent manner (Fig. 1A;
P<0.05 or P<0.001 compared with control). Compared with the
control group, there was no significant alteration in the level of
FGF secreted from HL-60E following stimulation with rhTSLP
(Fig. 1B; P>0.05 compared with
control). In addition, the secretion of IL-8 in HL-60E was
undetected (data not shown). The results of the present study
demonstrated that exogenous TSLP upregulated VEGF secretion of
HL-60E cells, and may be involved in the promotion of
angiogenesis.
Co-culture of HL-60E with cervical
cancer cells increases the production of IL-8 and VEGF
In order to imitate the local immune environment, a
co-culture system of HL-60E and HeLa or CasKi cells was
constructed. As presented, HeLa or CasKi cells alone secreted an
increased level of IL-8 and VEGF compared with that secreted by
HL-60E cells alone (Fig. 2A and B).
Following co-culture of HL-60E and HeLa, there were increased
levels of IL-8 (Fig. 2A; P<0.001
compared with HeLa cells alone or HL-60E alone) and VEGF (Fig. 2B; P<0.01 compared with HeLa cells
alone; P<0.001 compared with HL-60E alone), but no change in FGF
production (Fig. 2C; P>0.05
compared with HeLa cells alone or HL-60E alone). Notably, IL-8,
VEGF and FGF levels were increased in co-culture of HL-60E and
CasKi cells (Fig. 2; P<0.01
compared with CasKi cells alone for VEGF and FGF; P<0.001
compared with CasKi cells alone for IL-8; P<0.001 compared with
HL-60E alone for IL-8 and VEGF). The results of the present study
indicated that the interaction between EOS and cervical cancer
cells in cancer lesions may involve the secretion of an increased
level of tumor-associated angiogenic factors.
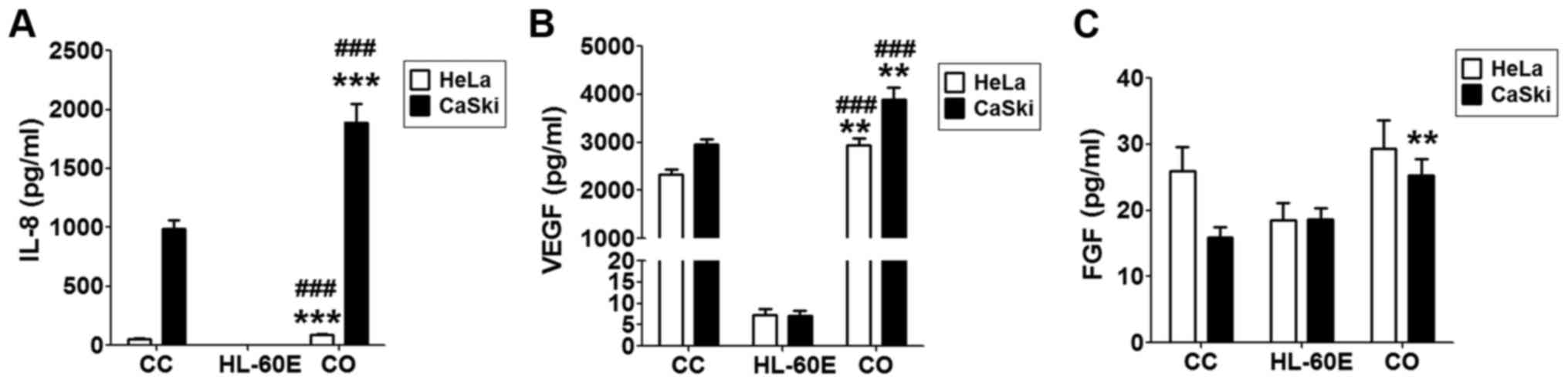 | Figure 2.Co-culture of HL-60E with cervical
cancer cells increases the production of IL-8 and VEGF. HL-60E were
co-cultured with HeLa or CaSki cells for 48 h, and the secretion
level of (A) IL-8, (B) VEGF and (C) FGF were analyzed using ELISA.
The data are expressed as the mean ± standard error of the mean.
**P<0.01 or ***P<0.001 vs. CC, determined using a one-way
analysis of variance; ###P<0.001 vs. HL-60E alone,
determined using a one-way analysis of variance. CC, HeLa or CaSki
cells alone; HL-60E, HL-60-eosinophils alone; CO, co-culture group
of HL-60E with HeLa (black) or CaSki (white) cells; IL,
interleukin; VEGF, vascular endothelial growth factor; FGF,
fibroblast growth factor. |
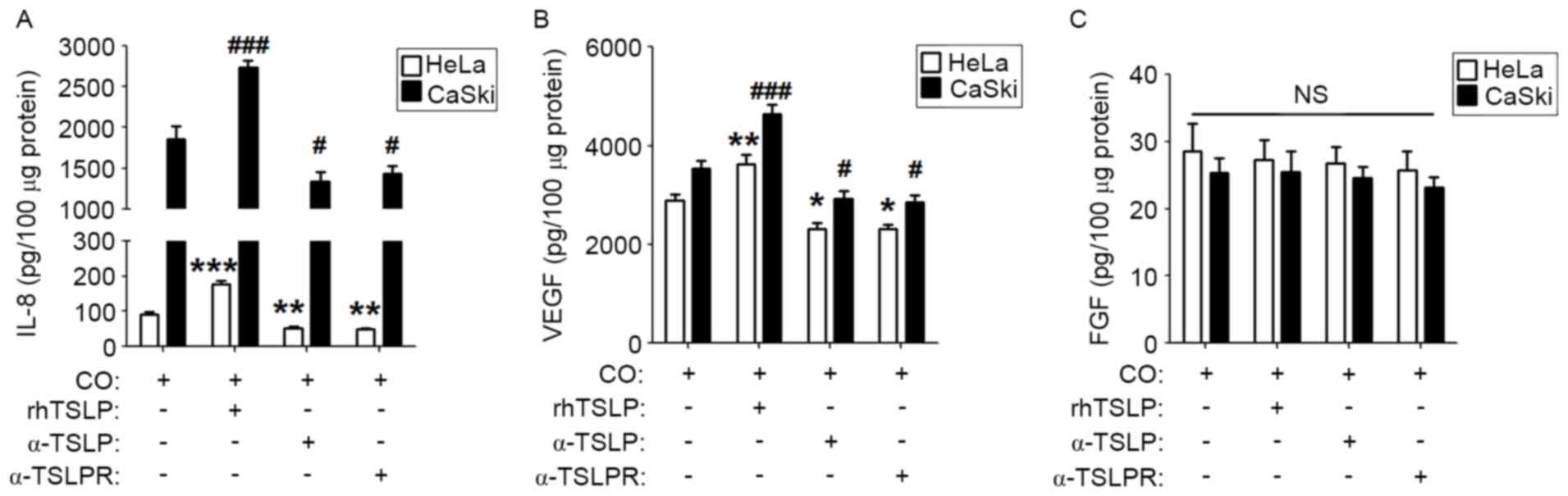
TSLP-derived of cervical cancer cells
upregulates the secretion of IL-8 and VEGF
To explore whether TSLP secreted by cervical cancer
cells is involved in regulating the level of IL-8, VEGF and FGF in
a co-culture system, rhTSLP, α-TSLP or α-TSLPR was added to the
co-culture system. As presented, rhTSLP increased IL-8 and VEGF
levels in both co-culture systems compared with the control
co-culture groups (Fig. 3A and B;
P<0.01 or P<0.001). Inhibiting the TSLP signal with α-TSLP or
α-TSLPR resulted in the decreased secretion of IL-8 and VEGF in the
co-culture systems (Fig. 3A and B;
P<0.05 or P<0.01). However, neither rhTSLP nor TSLP signal
inhibitors affected the FGF production in the co-culture system
(Fig. 3C; P>0.05). These results
suggested that endogenous TSLP secreted by cervical cancer cells
causes an increased production of angiogenic factors in the
co-culture system between cervical cancer cells and EOS.
TSLP stimulates angiogenesis by
strengthening the interaction between cervical cancer cells and
EOS
To identify the function of cervical cancer cells
and EOS in the regulation of angiogenesis in HUVECs, HUVECs were
stimulated with the supernatants from HeLa (S-HeLa), CasKi
(S-CasKi), HL-60E (S-HL-60E), co-culture of HL-60E and HeLa
(S-H+H), or co-culture of HL-60E and CasKi (S-H+C), and the tube
formation of HUVECs were subsequently analyzed. As a positive
control, rhVEGF protein significantly stimulated angiogenesis of
HUVECs compared with the control (Fig. 4A
and B; P<0.001). Compared with the control, S-HeLa, S-CasKi
or S-HL-60E resulted in an increase in angiogenesis of HUVECs in
vitro (Fig. 4A and B; P<0.05).
Compared with S-HeLa, S-CasKi and S-HL-60E alone, S-H+H and S-H+C
exhibited a significantly increased stimulatory effect on tube
formation of HUVECs (Fig. 4A and B;
P<0.01 or P<0.001).
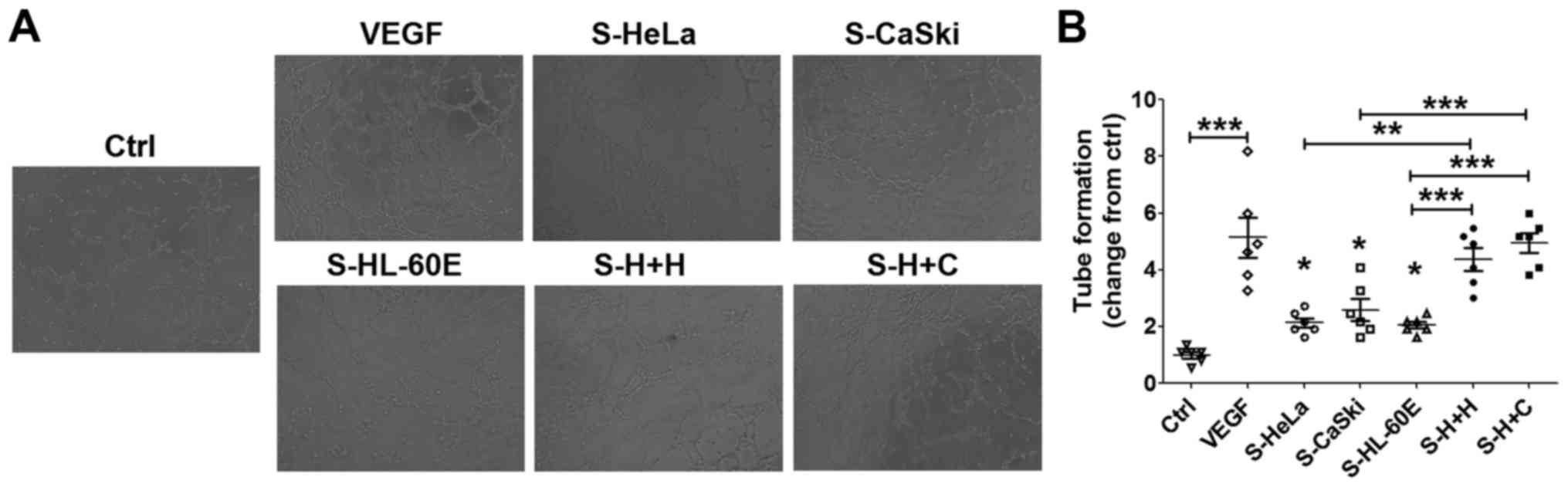 | Figure 4.Cervical cancer cells and EOS
promotes angiogenesis of HUVECs. (A) HUVECs were treated with the
supernatants from HL-60E cells, HeLa, CaSki cells, the culture
system of HL-60E cells and HeLa or CaSki cells, or with rhVEGF (10
ng/ml) as the positive control. Original magnification, ×100. (B)
Subsequently, the tube formation assay was performed to analyze the
angiogenesis of HUVECs. The data are expressed as the mean ±
standard error of the mean. *P<0.05, **P<0.01 and
***P<0.001 (one-way analysis of variance). Ctrl, control;
RhVEGF, recombinant human VEGF; S-HeLa, the supernatant from HeLa
cells; S-CasKi, supernatant from CasKi cells; S-HL-60E, supernatant
from HL-60E cells; S-H+H, the supernatant from the co-culture of
HL-60E and HeLa cells; S-H+C, supernatant from co-culture of HL-60E
and CasKi cells; EOS, eosinophils; rh, recombinant; VEGF, vascular
endothelial growth factor; HUVECs, human umbilical vein endothelial
cells. |
Subsequent analysis revealed that these effects may
be abrogated by inhibiting TSLP or TSLPR (Fig. 5A and B; P<0.01 compared with
control). The results of the present study suggested that the
interaction between HL-60E and cervical cancer cells promotes
angiogenesis of HUVECs in vitro. Furthermore, TSLP may
trigger this process by producing IL-8 and VEGF secretion.
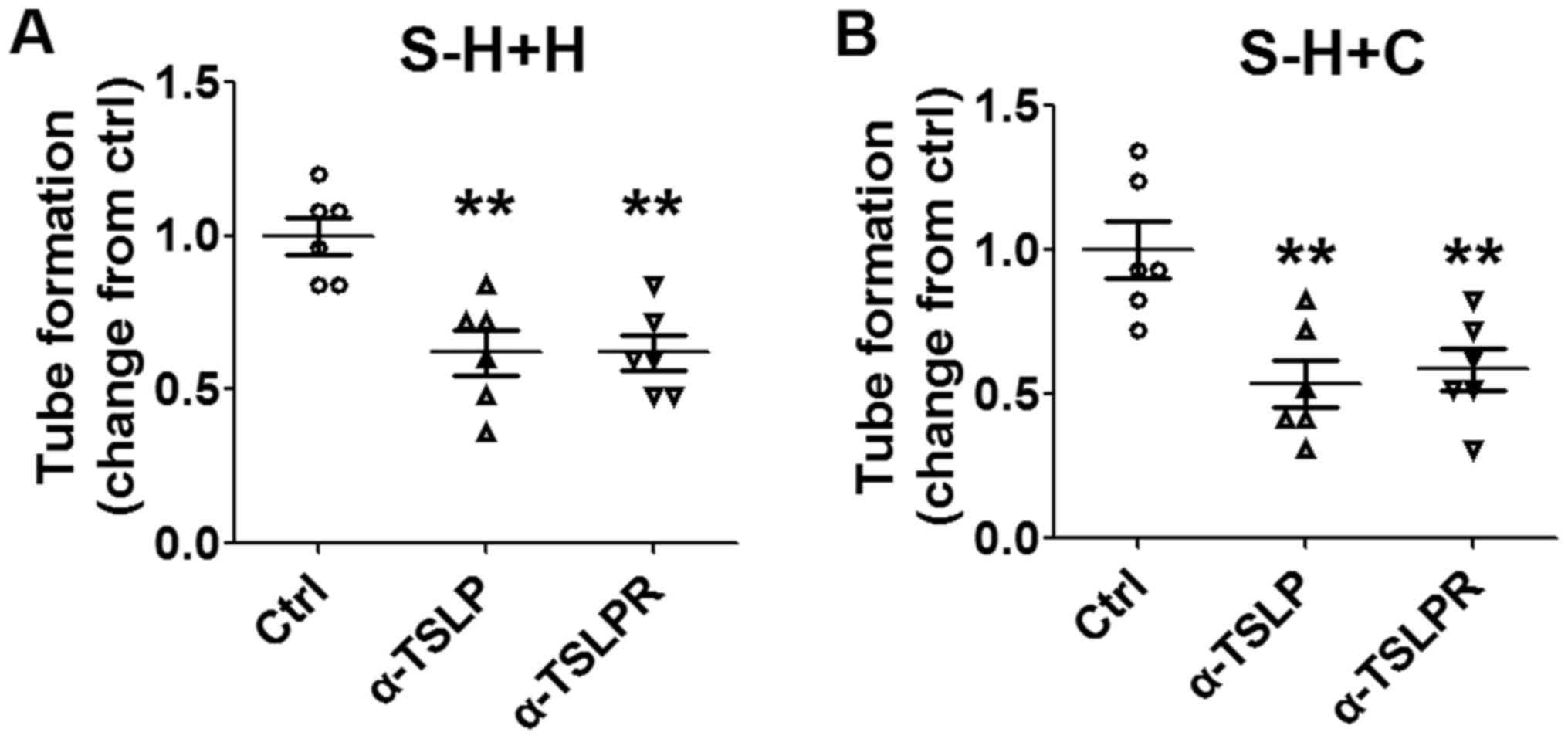 | Figure 5.Stimulatory effect of cervical cancer
cells and EOS on angiogenesis is dependent on TSLP. Following
pre-treatment with α-TSLP (5 ug/ml) or α-TSLPR (5 ug/ml) for 48 h,
supernatants were selected from the co-culture system of HL-60E
with (A) HeLa or (B) CaSki cells. Subsequently, HUVECs were
stimulated with these supernatants and angiogenesis was analyzed
using a tube formation assay. The data are expressed as the mean ±
standard error of the mean. **P<0.01 vs. control, using one-way
analysis of variance. EOS, eosinophils; Ctrl, control; S-H+H,
supernatant of co-culture of HL-60E with HeLa cells; S-H+C,
supernatant of co-culture of HL-60E with CaSki cells; EOS,
eosinophils; TSLP, thymic stromal lymphopoietin; α-TSLP, anti-human
TSLP neutralizing antibody; α-TSLPR, anti-human TSLP receptor
neutralizing antibody; HUVECs, human umbilical vein endothelial
cells. |
Discussion
EOS infiltration is a common host response of solid
tumors. However, there is no change observed in the number and
activation state of peripheral blood EOS (20–22). Ayhan
et al (23) demonstrated that
between 25 and 100% of cervical carcinoma tissues contained EOS,
and between 2 and 26% of cervical tumor microenvironments exhibited
a significant proportion of EOS infiltration (23). EOS express numerous types of surface
functional molecules, including pattern-recognition receptors,
siglec-lectin receptors, adhesion molecules, Toll-like receptors,
and receptors for cytokines and chemokines (20,24). The
expression of these molecules are required for functions in
cytotoxic activity via secretory granule proteins, including a
matrix composed of eosinophil cationic protein, major basic protein
1 and 2, eosinophil-derived neurotoxin, and eosinophil peroxidase.
Three cytokines, IL-3, IL-5 and granulocyte macrophage
colony-stimulating factor (GM-CSF), are required for the regulation
of EOS development. EOS may be recruited via eosinophil chemokines
eotaxin-1 (CCL11), eotaxin-2 and eotaxin-3 (24,25). In
the majority of types of solid tumor, EOS tissue infiltration is
located in the tumor necrosis area (21). Our previous study revealed that EOS
infiltration of the lesion site increased with the progression of
cervical cancer (19). TSLP of
cervical cancer cells induced by hypoxia was identified to be
involved in the recruitment of EOS by stimulating the secretion of
chemokine (C-C motif) ligand 17 (19). Previous studies have demonstrated an
improved prognosis with tumor-associated tissue eosinophilia
(TATE), due to the tumoricidal effects of EOS via degranulation in
the local cancer lesions (26,27).
However, other studies have suggested that TATE was a poor
prognostic indicator in distinct types of solid tumor, including
oral squamous cell carcinoma and cervical carcinoma (19,28). Thus,
the underlying molecular mechanism of EOS in cancer remains
unknown. Previously, we have demonstrated that abnormal increased
TSLP in cancer lesions is an important regulator in the progression
of cervical cancer, via recruiting and enabling tumor-associated
EOS to promote the growth of cervical cancer cells (19).
Blood vessels may serve as a promoter for cancer
growth and metastasis by transporting oxygen and nutrients, and
removing metabolites (29).
Furthermore, in order to metastasize, cancer cells must invade the
tumor-associated neovasculature to obtain access to a distant site
in the body (30). Angiogenesis, the
formation of new blood vessels from existing ones, is an essential
process in physiological and pathological conditions (31,32).
Numerous cytokines, including VEGF, regulate the functions of
vascular endothelial cells (32–34). Our
previous study demonstrated that cervical cancer cells stimulate
angiogenesis of vascular endothelial cells by secreting TSLP
(18).
A previous study revealed that the recruitment of
EOS to the tumor site may promote angiogenesis and this effect may
be induced by VEGF (35). In
addition, EOS releases angiogenic factors, including FGF, IL-6,
IL-8, GM-CSF, platelet derived factor and transforming growth
factor β (36). A previous in
vivo study demonstrated that EOS-derived CCL11 induced an
angiogenic response (37). EOS and
tumor associated macrophages accumulated in the hypoxic and/or
necrotic areas of mouse subcutaneous B16F10 melanoma, and this
process was associated with EOS degranulation (22). These particles of EOS degranulation
contain VEGF and other angiogenic factors, which may promote tumor
angiogenesis in the hypoxic areas (38,39). In
addition, EOS promotes angiogenesis in local cancer lesion by
releasing a large number of vasoactive leukotrienes (40). Therefore, in the present study, it was
hypothesized that TSLP produced by cervical cancer cells may
indirectly regulate angiogenesis by influencing EOS function. The
results of the present study demonstrated that rhTSLP treatment
significantly upregulated the level of VEGF, but not the FGF level
in HL-60E cells in vitro. EOS of peripheral blood could
produce IL-8. HL-60E cells did not secrete IL-8. However, HL-60E
may promote IL-8 secretion from HeLa or CasKi cells. Following the
co-culture of HL-60E cells with HeLa or CasKi cells, the secretion
level of IL-8 and VEGF was significantly increased, and this effect
was upregulated and inhibited by rhTSLP, and TSLP signal
inhibitors, respectively. The results of the present study
suggested that TSLP results in an increased level of angiogenic
factors through promoting the interaction between EOS and cervical
cancer cells.
Subsequent analysis revealed that cervical cancer
cells (HeLa or CasKi) and EOS promoted the angiogenesis of HUVECs
in vitro, in particular following the co-culture of cervical
cancer cells and EOS. However, these effects were inhibited by
suppressing TSLP or TSLPR by α-TSLP or α-TSLPR, respectively, which
supported our hypothesis.
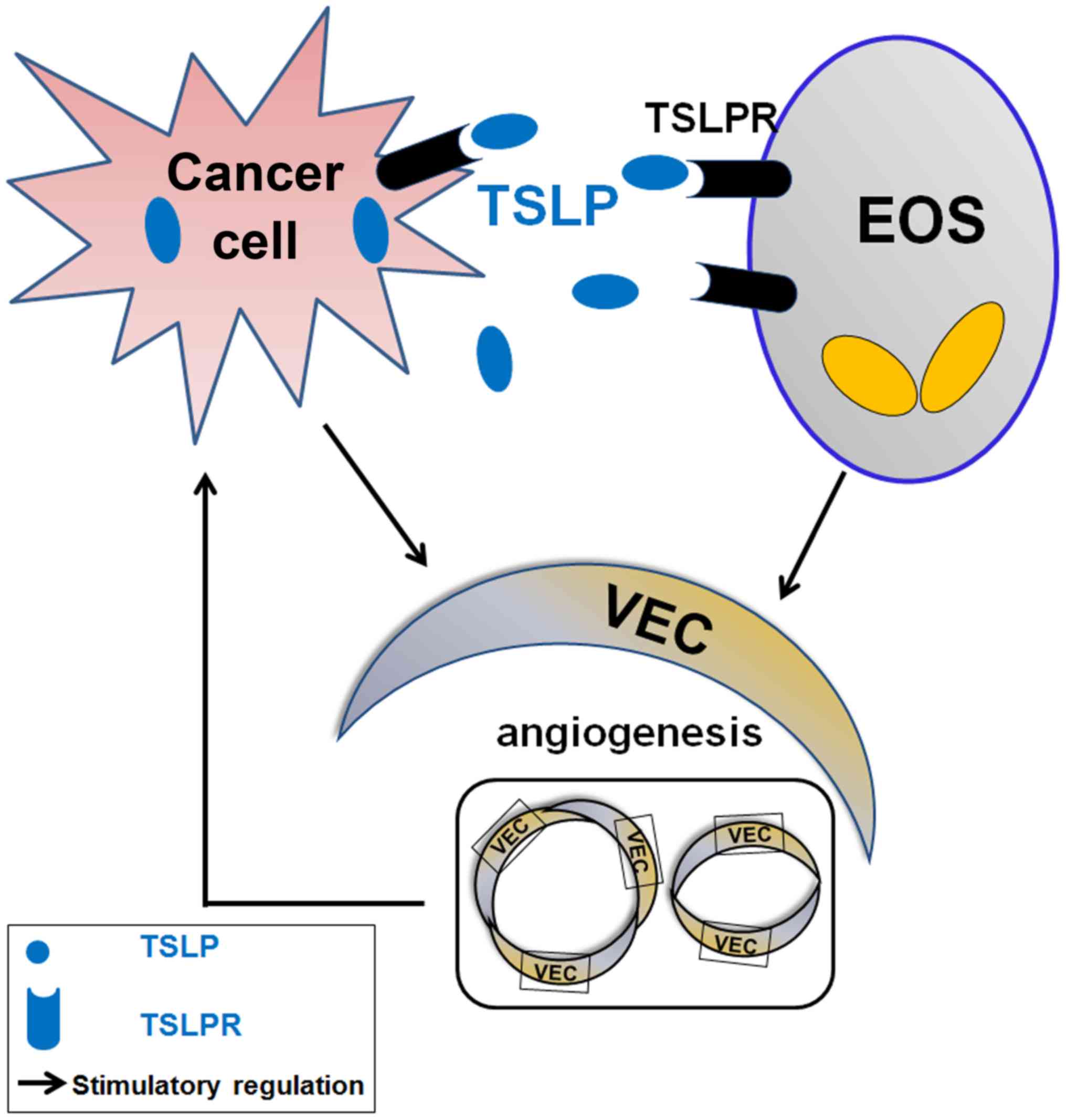
The present study demonstrated that TSLP derived
from cervical cancer cells promotes the secretion of VEGF of EOS by
binding TSLPR and increases the levels of angiogenic factors (IL-8
and VEGF) in the co-culture system by stimulating the interaction
between cervical cancer cells and EOS (Fig. 6). The effects of TSLP on cervical
cells and EOS may result in the increase of tumor angiogenesis, and
contribute to the development and progression of cervical cancer.
The results of the present study further the understanding of the
biological function of EOS in cancer progression. However, the
molecular exact mechanism underlying the effects of TSLP on the
level of angiogenic factors secreted from cervical cancer cells and
EOS remains unknown, and requires further investigation.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 91542108, 81471513
and 31600735), the Shanghai Rising-Star Program (grant no.
16QA1400800), the Development Fund of Shanghai Talents (grant no.
201557), the Program for Zhuoxue of Fudan University (grant no.
1326) and the Shanghai Natural Science Foundation (grant no.
17ZR1403200).
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
de Freitas AC, Gurgel AP, Chagas BS,
Coimbra EC and do Amaral CM: Susceptibility to cervical cancer: An
overview. Gynecol Oncol. 126:304–311. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wani K and Nair CK: Genetic alterations in
cervical cancer. Indian J Exp Biol. 41:789–796. 2003.PubMed/NCBI
|
|
4
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jiang J, Yan M, Mehta JL and Hu C:
Angiogenesis is a link between atherosclerosis and tumorigenesis:
Role of LOX-1. Cardiovasc Drugs Ther. 25:461–468. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
De Palma M, Biziato D and Petrova TV:
Microenvironmental regulation of tumour angiogenesis. Nat Rev
Cancer. 17:457–474. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nyberg P, Salo T and Kalluri R: Tumor
microenvironment and angiogenesis. Front Biosci. 13:6537–6553.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Folkman J and Shing Y: Angiogenesis. J
Biol Chem. 267:10931–10934. 1992.PubMed/NCBI
|
|
9
|
Chung AS and Ferrara N: Developmental and
pathological angiogenesis. Annu Rev Cell Dev Biol. 27:563–584.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fujimoto J, Sakaguchi H, Aoki I and Tamaya
T: Clinical implications of expression of interleukin 8 related to
angiogenesis in uterine cervical cancers. Cancer Res. 60:2632–2635.
2000.PubMed/NCBI
|
|
11
|
López-Ocejo O, Viloria-Petit A,
Bequet-Romero M, Mukhopadhyay D, Rak J and Kerbel RS: Oncogenes and
tumor angiogenesis: The HPV-16 E6 oncoprotein activates the
vascular endothelial growth factor (VEGF) gene promoter in a p53
independent manner. Oncogene. 19:4611–4620. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu YJ, Soumelis V, Watanabe N, Ito T,
Wang YH, Malefyt Rde W, Omori M, Zhou B and Ziegler SF: TSLP: An
epithelial cell cytokine that regulates T cell differentiation by
conditioning dendritic cell maturation. Annu Rev Immunol.
25:193–219. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sokol CL, Barton GM, Farr AG and Medzhitov
R: A mechanism for the initiation of allergen-induced T helper type
2 responses. Nat Immunol. 9:310–318. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rochman Y and Leonard WJ: Thymic stromal
lymphopoietin: A new cytokine in asthma. Curr Opin Pharmacol.
8:249–254. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu YJ: Thymic stromal lymphopoietin and
OX40 ligand pathway in the initiation of dendritic cell-mediated
allergic inflammation. J Allergy Clin Immunol. 120:238–246. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Park LS, Martin U, Garka K, Gliniak B, Di
Santo JP, Muller W, Largaespada DA, Copeland NG, Jenkins NA, Farr
AG, et al: Cloning of the murine thymic stromal lymphopoietin
(TSLP) receptor: Formation of a functional heteromeric complex
requires interleukin 7 receptor. J Exp Med. 192:659–670. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Reche PA, Soumelis V, Gorman DM, Clifford
T, Liu Mr, Travis M, Zurawski SM, Johnston J, Liu YJ, Spits H, et
al: Human thymic stromal lymphopoietin preferentially stimulates
myeloid cells. J Immunol. 167:336–343. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xie F, Meng YH, Liu LB, Chang KK, Li H, Li
MQ and Li DJ: Cervical carcinoma cells stimulate the angiogenesis
through TSLP promoting growth and activation of vascular
endothelial cells. Am J Reprod Immunol. 70:69–79. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xie F, Liu LB, Shang WQ, Chang KK, Meng
YH, Mei J, Yu JJ, Li DJ and Li MQ: The infiltration and functional
regulation of eosinophils induced by TSLP promote the proliferation
of cervical cancer cell. Cancer Lett. 364:106–117. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Davis BP and Rothenberg ME: Eosinophils
and cancer. Cancer Immunol Res. 2:1–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Facciabene A, Peng X, Hagemann IS, Balint
K, Barchetti A, Wang LP, Gimotty PA, Gilks CB, Lal P, Zhang L and
Coukos G: Tumour hypoxia promotes tolerance and angiogenesis via
CCL28 and T(reg) cells. Nature. 475:226–230. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cormier SA, Taranova AG, Bedient C, Nguyen
T, Protheroe C, Pero R, Dimina D, Ochkur SI, O'Neill K, Colbert D,
et al: Pivotal advance: Eosinophil infiltration of solid tumors is
an early and persistent inflammatory host response. J Leukoc Biol.
79:1131–1139. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ayhan A, Altintaş A, Tuncer ZS, Küçükali T
and Yüce K: Prognostic value of mitotic activity, eosinophilic and
inflammatory reaction in stage I cancer of the uterine cervix. Eur
J Surg Oncol. 18:264–266. 1992.PubMed/NCBI
|
|
24
|
Lotfi R, Lee JJ and Lotze MT: Eosinophilic
granulocytes and damage-associated molecular pattern molecules
(DAMPs): Role in the inflammatory response within tumors. J
Immunother. 30:16–28. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Stone KD, Prussin C and Metcalfe DD: IgE,
mast cells, basophils, and eosinophils. J Allergy Clin Immunol. 125
2 Suppl 2:S73–S80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pretlow TP, Keith EF, Cryar AK, Bartolucci
AA, Pitts AM, Pretlow TG II, Kimball PM and Boohaker EA: Eosinophil
infiltration of human colonic carcinomas as a prognostic indicator.
Cancer Res. 43:2997–3000. 1983.PubMed/NCBI
|
|
27
|
Caruso RA, Parisi A, Quattrocchi E,
Scardigno M, Branca G, Parisi C, Lucianò R, Paparo D and Fedele F:
Ultrastructural descriptions of heterotypic aggregation between
eosinophils and tumor cells in human gastric carcinomas.
Ultrastruct Pathol. 35:145–149. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Horiuchi K, Mishima K, Ohsawa M, Sugimura
M and Aozasa K: Prognostic factors for well-differentiated squamous
cell carcinoma in the oral cavity with emphasis on
immunohistochemical evaluation. J Surg Oncol. 53:92–96. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hanahan D and Folkman J: Patterns and
emerging mechanisms of the angiogenic switch during tumorigenesis.
Cell. 86:353–364. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Carmeliet P: Angiogenesis in life, disease
and medicine. Nature. 438:932–936. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Folkman J and Kalluri R: Cancer without
disease. Nature. 427:7872004. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ruhrberg C: Growing and shaping the
vascular tree: Multiple roles for VEGF. Bioessays. 25:1052–1060.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sulpice E, Ding S, Muscatelli-Groux B,
Bergé M, Han ZC, Plouet J, Tobelem G and Merkulova-Rainon T:
Cross-talk between the VEGF-A and HGF signalling pathways in
endothelial cells. Biol Cell. 101:525–539. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cross MJ and Claesson-Welsh L: FGF and
VEGF function in angiogenesis: Signalling pathways, biological
responses and therapeutic inhibition. Trends Pharmacol Sci.
22:201–207. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Puxeddu I, Alian A, Piliponsky AM, Ribatti
D, Panet A and Levi-Schaffer F: Human peripheral blood eosinophils
induce angiogenesis. Int J Biochem Cell Biol. 37:628–636. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Munitz A and Levi-Schaffer F: Eosinophils:
‘New’ roles for ‘old’ cells. Allergy. 59:268–275. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Salcedo R, Young HA, Ponce ML, Ward JM,
Kleinman HK, Murphy WJ and Oppenheim JJ: Eotaxin (CCL11) induces in
vivo angiogenic responses by human CCR3+ endothelial
cells. J Immunol. 166:7571–7578. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Efraim Nissim Ben AH, Eliashar R and
Levi-Schaffer F: Hypoxia modulates human eosinophil function. Clin
Mol Allergy. 8:102010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Porter LM, Cowburn AS, Farahi N, Deighton
J, Farrow SN, Fiddler CA, Juss JK, Condliffe AM and Chilvers ER:
Hypoxia causes IL-8 secretion, Charcot Leyden crystal formation,
and suppression of corticosteroid-induced apoptosis in human
eosinophils. Clin Exp Allergy. 47:770–784. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Temkin V, Aingorn H, Puxeddu I, Goldshmidt
O, Zcharia E, Gleich GJ, Vlodavsky I and Levi-Schaffer F:
Eosinophil major basic protein: First identified natural
heparanase-inhibiting protein. J Allergy Clin Immunol. 113:703–709.
2004. View Article : Google Scholar : PubMed/NCBI
|