Introduction
Hepatocellular carcinoma (HCC) is the fifth most
common malignant tumor and the third leading cause of
cancer-associated mortality worldwide (1,2). There are
a variety of HCC treatment modalities, and chemotherapy is an
important and irreplaceable therapeutic strategy for the majority
of patients with HCC (3). Cisplatin
is a common chemotherapeutic agent used for HCC treatment. However,
cancer cells may develop multidrug resistance (MDR) against
chemotherapeutic agents, including cisplatin, which leads to the
failure of chemotherapy (4). There
are a number of mechanisms involved in MDR. One form of MDR is
caused by the overexpression of ATP-binding cassette sub-family B
member 1 (ABCB1), which pumps anticancer agents out of cells,
contributing to a reduced intracellular drug concentration and
cytotoxicity (5,6). Inhibition of ABCB1 expression is able to
reduce the resistance of HCC cells to anticancer agents (6). Therefore, identification of the
mechanism of action and identification of novel targets for
inhibiting ABCB1 may aid the reversal of MDR in HCC.
Long non-coding RNAs (lncRNAs) are
non-protein-coding transcripts >200 nucleotides. Within the past
10–15 years, lncRNAs have gained the widespread attention of
researchers, as they are frequently dysregulated in a variety of
human diseases, including cancer, and have critical roles in
regulating various biological processes (7,8). HOX
transcript antisense RNA (HOTAIR), is located within the homeobox C
(HOXC) gene cluster on chromosome 12, which interacts with polycomb
repressive complex (PRC2) to enhance the trimethylation of lysine
27 of histone H3 (H3K27), leading to the repression of multiple
genes (8,9). HOTAIR was initially identified to be
upregulated, and to promote invasiveness and metastasis in breast
cancer (8). In addition, HOTAIR was
demonstrated to promote proliferation and reduce apoptosis in
pancreatic cancer, colorectal cancer, and HCC (10–12).
Overexpression of HOTAIR was positively associated with poor
prognosis, tumor progression and recurrence in HCC (13,14). A
recent study reported that HOTAIR also promoted the malignant
growth of human liver cancer stem cells (15). A number of studies implicated that
HOTAIR has important roles in the tumorigenesis of HCC (12–15).
However, the association between HOTAIR and HCC MDR remains
unclear.
The present study investigated the potential role of
HOTAIR in the regulation of MDR in HCC. HOTAIR small interfering
RNA (siRNA) was used to silence HOTAIR expression in human Huh7 HCC
cells and then the chemosensitivity of Huh7 cells to cisplatin was
examined. Knockdown of HOTAIR resulted in decreased cell
proliferation and increased chemosensitivity to cisplatin.
Furthermore, the increased chemosensitivity of Huh7 cells to
cisplatin was associated with decreased expression levels of ABCB1
mRNA and protein when HOTAIR was knocked down. In addition,
inhibition of signal transducer and activator of transcription 3
(STAT3) reduced HOTAIR-mediated ABCB1 expression. These results
indicate that HOTAIR may have a particular important role in
regulation of the chemosensitivity of human HCC cells. The findings
of the present study provide a novel insight into the mechanism of
HOTAIR in regulating MDR of HCC.
Materials and methods
Cell line and culture
The Human Huh7 HCC cell line was purchased from the
American Type Culture Collection (ATCC, Manassas, VA, USA). Cells
were maintained in RPMI-1,640 medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (FBS) (Gibco; Thermo Fisher Scientific, Inc.) at 37°C
in a humid atmosphere with 5% CO2. The culture medium
was changed every 24 h. AG490 (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) was used as an inhibitor of STAT3 (16), at the final concentrations of 25 µM
for 1 h at room temperature prior to siRNA transfection.
HOTAIR siRNA transfection
Knockdown of human HOTAIR (National Center for
Biotechnology Information gene ID, 100,124,700) was achieved using
transfection with siRNAs. The siRNA sequences were as follows:
HOTAIR, 5′-UAACAAGACCAGAGAGCUGUU-3′; negative control (NC),
5′-TTCTCCGAACGTGTCACGT-3′. The siRNA were synthesized by GenePharma
(GenePharma Co, Ltd, Shanghai, China). Cells were transfected with
100 nM siRNA duplexes using Lipofectamine™ 2000
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
following manufacturer's protocols. The mock transfected cells were
transfected with Lipofectamine 2000 without siRNA (17). Following transfection, cells were
incubated in a humidified chamber supplemented with 5%
CO2 at 37°C. At 48 h after transfection, cells were
harvested for RT-qPCR and Western blotting assay.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was performed to determine expression of
lncRNA HOTAIR (12), and mRNA
expression of ABCB1 (16). Briefly,
at 48 h after transfection, cells were harvested in TRIzol Reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's protocols. Total RNA was reverse transcribed into
cDNA using PrimeScript™ RT Reagent kit (Takara Biotechnology Co.,
Ltd., Dalian, China). RT-qPCR amplification reactions were
performed with SYBR® Premix Ex Taq™ II (Takara
Biotechnology Co., Ltd.). RT-qPCR was performed on CFX96 Real-Time
PCR Detection system (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). Thermocycling conditions were as follows: Initial
denaturation at 95°C for 5 min, followed by 40 cycles of
denaturation at 95°C for 20 sec, annealing at 58°C for 30 sec and
elongation at 72°C for 30 sec. All fold changes were calculated
using the comparative Cq (2−ΔΔCq) method using GAPDH for
normalization (18). Primer sequences
used in this study were as follows: ABCB1 forward,
5′-CTTCAGGGTTTCACATTTGGC-3′ and reverse,
5′-GGTAGTCAATGCTCCAGTGG-3′; GAPDH forward,
5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse,
5′-TGGTGAAGACGCCAGTGGA-3′.
Western blotting assay
At 48 h after transfection, cells and supernatant of
each transfection group were collected. The cells were lysed with
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Nantong, China) in the presence of
protease/phosphatase inhibitor cocktail (Cell Signaling Technology,
Inc., Danvers, MA, USA) at 4°C for 30 min and the lysate was
centrifuged at 12,000 × g, 4°C for 20 min. The concentration of
protein was measured using a Bicinchoninic Acid Protein Assay kit
(Thermo Fisher Scientific, Inc.). Equal amounts (30 µg per lane) of
extracted protein samples were separated by 10% SDS-PAGE and
transferred onto polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA). Following blocking with 5% non-fat
milk for 1 h at room temperature, the membranes were incubated with
the following primary antibodies at 4°C under gentle agitation
overnight: ABCB1 (1:1,000; ab155421; Abcam, Cambridge, UK), pSTAT3
(1:1,000; #9145; Cell Signaling Technology, Inc.), STAT3 (1:1,000;
#9,139; Cell Signaling Technology, Inc.) and GAPDH (1:5,000;
sc-47,724; Santa Cruz Biotechnology, Inc., Dallas, TX, USA).
Following washing with TBST, membranes were incubated with
horseradish peroxidase-conjugated secondary antibody (1:5,000;
#1,662,408; Bio-Rad Laboratories, Inc.) for 1 h at room
temperature. The Supersignal® West Pico Chemiluminescent
substrate (Thermo Fisher Scientific, Inc.) was used to visualize
protein bands on X-ray films, and ImageJ software version 1.5
(National Institutes of Health, Bethesda, MD, USA) gel analysis was
used to quantify the protein bands from western blotting films.
Cell migration assay
Cell migration was tested using a Transwell chamber
assay. In total, 1.0×105 cells were seeded on an 8-µm
pore size Transwell (Merck KGaA) in serum-free RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc.), while RPMI-1,640 medium
(Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) was placed in the lower well. After
incubation at 37°C for 48 h, non-migrating cells in the upper
chamber surface were removed, migration cells were stained with the
Three Step Stain Set kit (Thermo Fisher Scientific, Inc.) for 30
min at room temperature. The number of migrated cells was counted
in 5 randomly selected fields using a light microscope at a
magnification of ×200.
Cell viability assay
Cell viability assay was performed by using the Cell
Counting kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan) method. Briefly, cells were seeded in 96-well
plates (Corning Incorporated, Corning, NY, USA). Following
overnight culture, Huh7 cells were transfected with siHOTAIR or
control siRNA for 24 h, then exposed to cisplatin (Selleck
Chemicals, Houston, TX, USA) at different final concentrations
(0.1, 0.25, 0.5, 1.0, 2.5, 5 µmol/l) for 48 h in a CO2
incubator, after which the viability was accessed. The CCK-8 assay
was used to detect the chemosensitivity of cells according to the
manufacturer's protocol. The absorbance at 450 nm was measured
using a microplate reader. A total of six replicate wells were used
for each group.
Statistical analysis
Results are presented as mean of three independent
experiments ± standard deviation. Statistical analyses were
performed using one-way analysis of variance and LSD t-test to
compare between multiple groups and Student's t-test to compare
between two groups using GraphPad Prism (version 4.0; GraphPad
Software Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
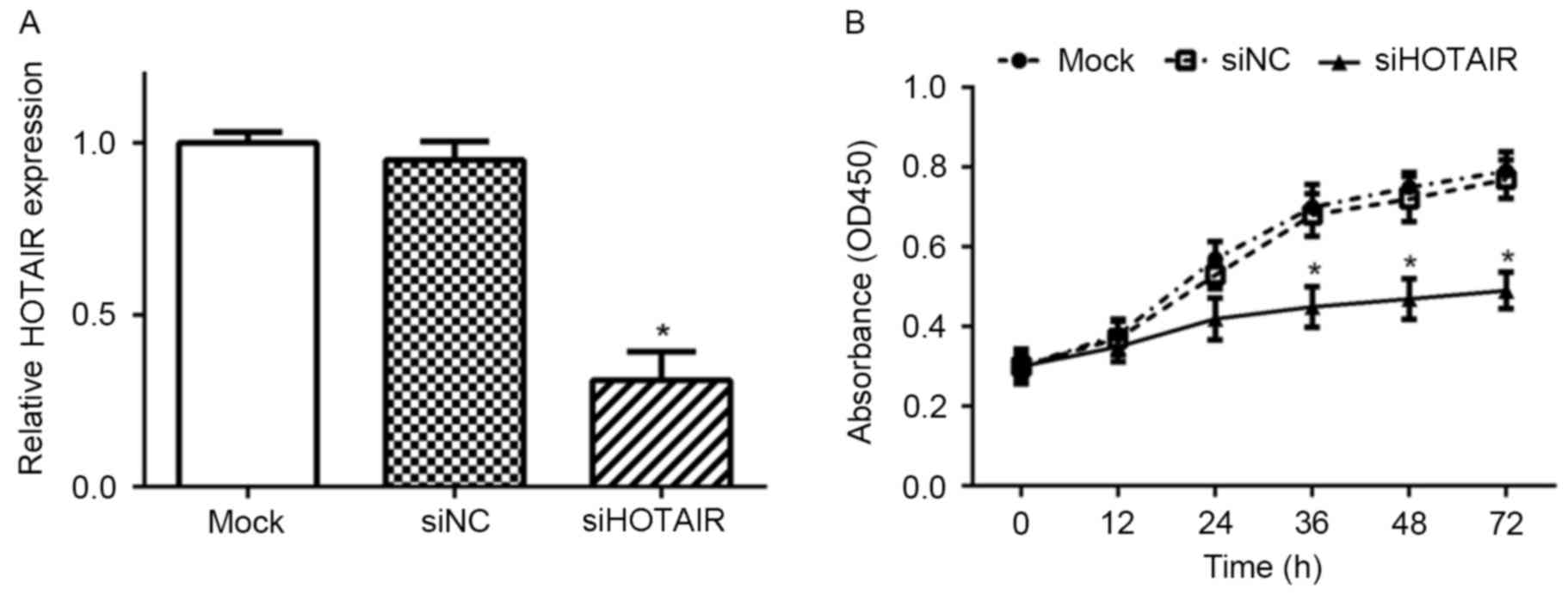
Knockdown of HOTAIR by specific
siRNA
To investigate the role of HOTAIR in HCC cells,
specific HOTAIR siRNA (siHOTAIR) was transfected into the Huh7 cell
line using Lipofectamine 2000. As presented in Fig. 1, transfection of Huh7 cells with
siHOTAIR resulted in significant knockdown of HOTAIR expression.
Furtheremore, NC siRNA (siNC) had no significant impact on HOTAIR
expression level compared with mock cells.
Knockdown of HOTAIR inhibits the
proliferation of Huh7 cells
To examine the role of HOTAIR on prolferation of
Huh7 cells, the effect of siHOTAIR on cell growth was assessed. As
presented in Fig. 1, the
proliferative capacity of Huh7 cells transfected with siHOTAIR was
significantly decreased, as compared with siNC and mock cells
(P<0.05).
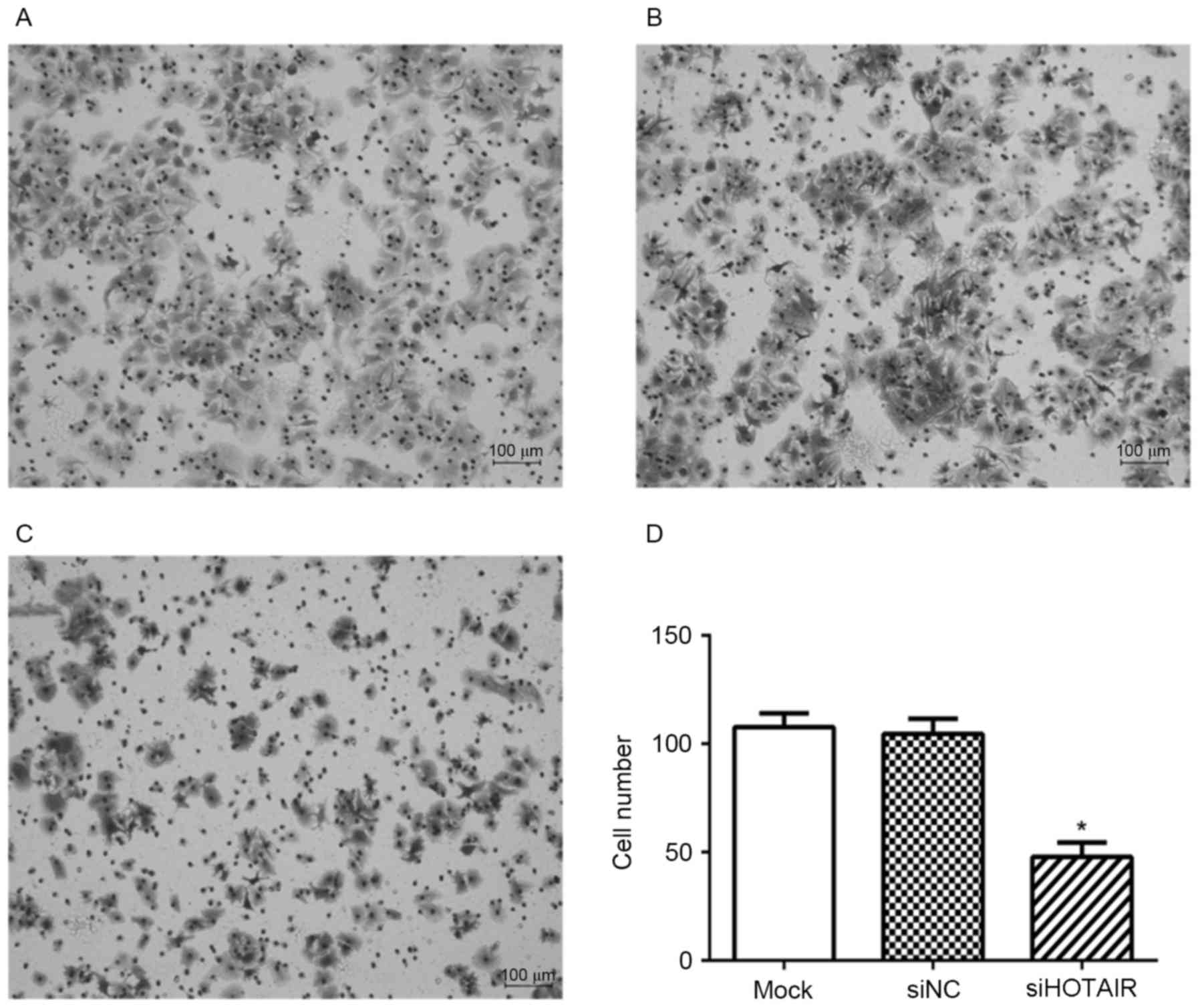
Knockdown of HOTAIR inhibits the
migration of Huh7 cells
The results of Transwell cell migration assay are
presented in Fig. 2. Knockdown of
HOTAIR significantly inhibited the migration ability of Huh7 cells
with 47.83±6.55 cells invaded, whereas 107.7±6.32 and 104.6±6.93
migrating cells were observed in mock, and siNC cells, respectively
(P<0.05, Fig. 2).
Knockdown of HOTAIR sensitizes Huh7
cells to cisplatin
To evaluate the effect of HOTAIR on chemosensitivity
of Huh7 cells, the viability of Huh7 cells transfected with
siHOTAIR and then exposed to cisplatin were assessed by CCK-8
assay. As presented in Fig. 3, Huh7
cells transfected with siHOTAIR were more sensitive to cisplatin
than siNC and mock cells, indicating that knockdown of HOTAIR
enhanced the sensitivity of Huh7 cells to cisplatin.
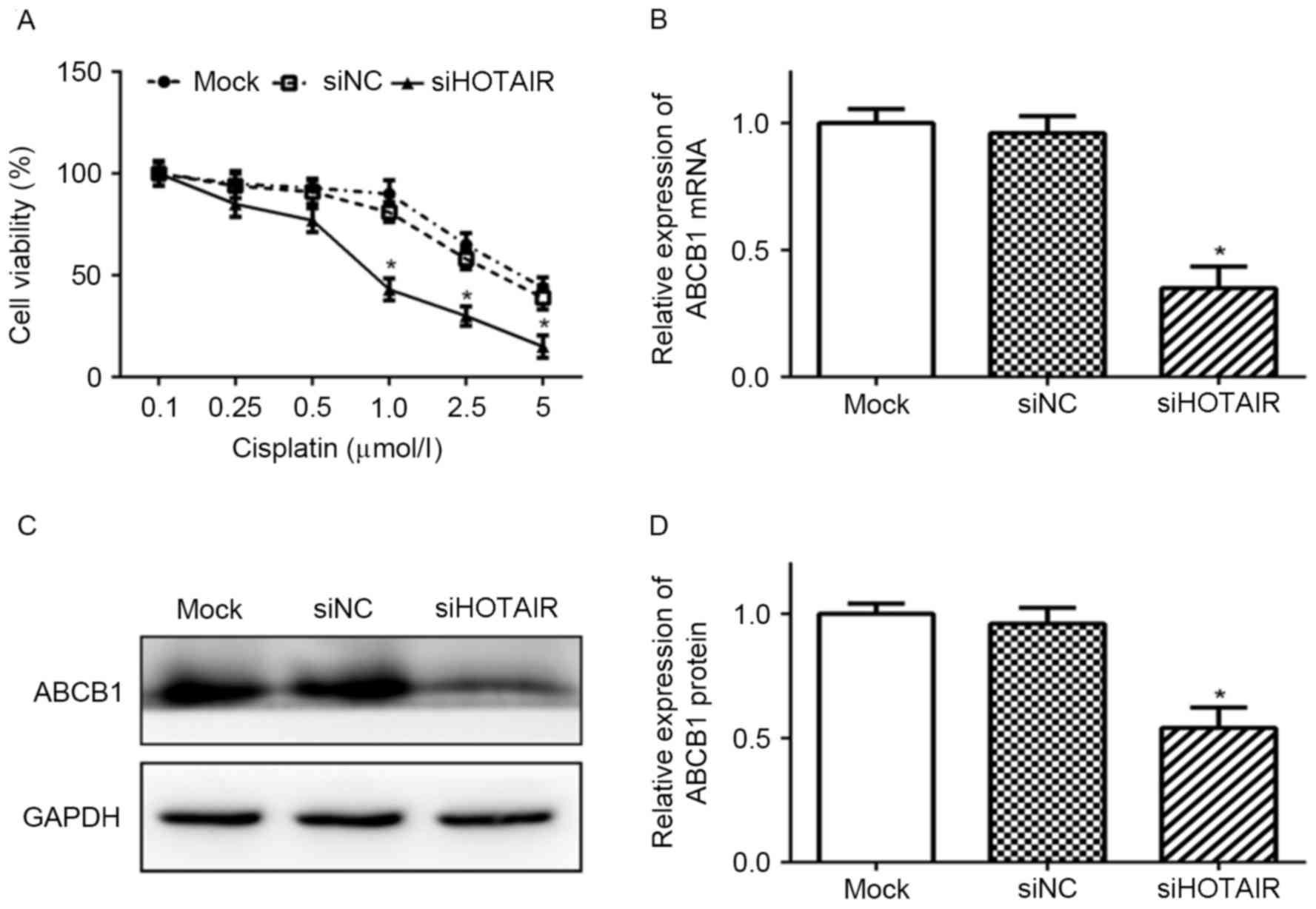 | Figure 3.Knockdown of HOTAIR sensitized Huh7
cells to cisplatin and reduced ABCB1 expression. (A) Huh7 cells
transfected with siHOTAIR or siNC were treated with cisplatin (0.1,
0.25, 0.5, 1.0, 2.5 or 5.0 µmol/l) for 48 h. Cell viability was
assessed by Cell Counting kit-8 assay (*P<0.05 vs. siNC).
Expression levels of ABCB1 (B) mRNA and (C, D) protein in Huh7
cells transfected with siHOTAIR or siNC, or mock cells was detected
by reverse transcription-quantitative polymerase chain reaction and
western blot analysis, respectively (*P<0.05 vs. siNC).
siHOTAIR, small interfering RNA targeted at HOX transcript
antisense RNA; NC, negative control; ABCB1, ATP binding cassette
subfamily B member 1. |
Knockdown of HOTAIR reduces the
expression of ABCB1 in Huh7 cells
To evaluate the effect of HOTAIR on cisplatin
sensitivity of Huh7 cells further, the expression of ABCB1, an
important mediator of MDR, was detected. As presented in Fig. 3, ABCB1 expression was positively
associated with HOTAIR expression. Compared with siNC and mock
cells, the expression levels of ABCB1 mRNA and protein were
significantly lower in siHOTAIR cells (P<0.05).
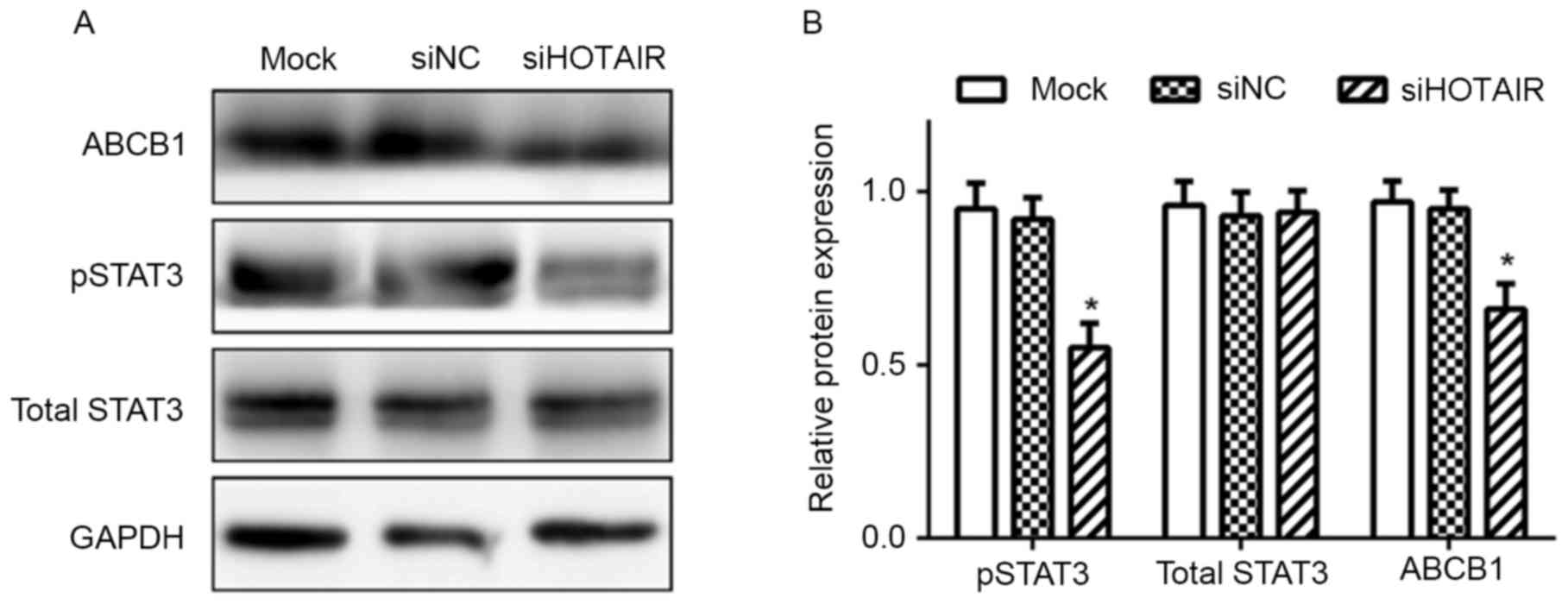
Knockdown of HOTAIR inhibits the
expression of phosphorylated (p)-STAT3 in Huh7 cells
Since STAT3 has been reported to be involved in the
regulation of ABCB1 expression (19),
whether HOTAIR regulated ABCB1 expression via the STAT3 signaling
was investigated. As demonstrated in Fig.
4, levels of p-STAT3 was significantly decreased in siHOTAIR
cells (P<0.05); in addition, the expression of total STAT3 was
not significantly affected in siHOTAIR cells, compared with siNC
and mock cells.
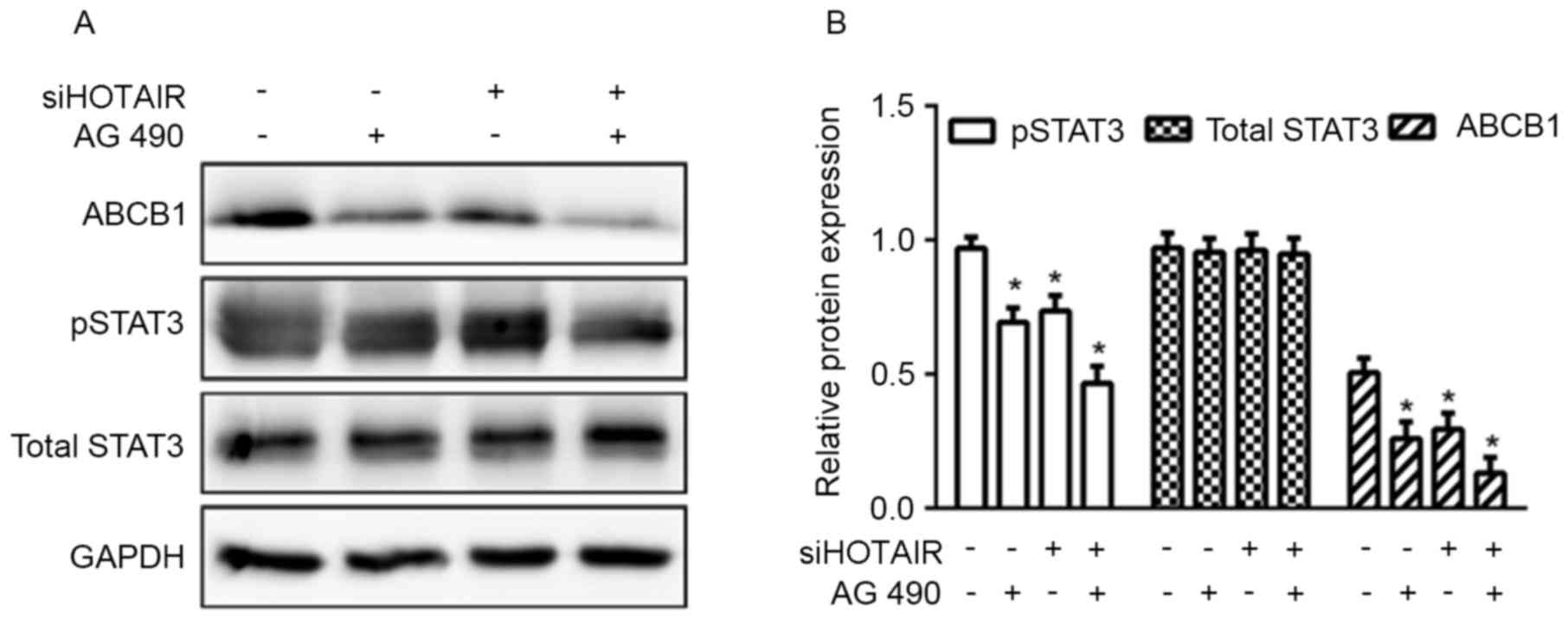
Inhibition of STAT3 reduces
HOTAIR-mediated ABCB1 expression
To further investigate the effect of STAT3 signaling
on HOTAIR-mediated ABCB1 expression, Huh7 cells were treated with
AG490, a STAT3 inhibitor (16), for 1
h prior to siRNA transfection. As presented in Fig. 5, levels of ABCB1 and pSTAT3 were
significantly reduced in the AG490-treated group (transfected with
either with siHOTAIR or siNC), but the suppresion was more evident
in the siHOTAIR group. Overall, the data indicated that HOTAIR
mediated ABCB1 expression and chemoresistance to cisplatin through
the STAT3 signaling pathway in HCC.
Discussion
MDR of cancer cells is one of primary reasons for
the failure of HCC chemotherapy and its suppression may increase
the efficacy of chemotherapy. A previous study confirmed that
inhibition of homeobox transcription factor NANOG reduced ABCB1
expression, leading to the enhanced chemosensitivity to doxorubicin
(17). The present study demonstrated
the role of the lncRNA HOTAIR in mediating MDR of HCC cells by
regulating ABCB1 expression, and revealed that the knockdown of
HOTAIR increased chemosensitivity to cisplatin by suppressing
STAT3/ABCB1 signaling.
lncRNAs have been identified as novel mediators of
critical biological activities in hepatocarcinogenesis (7). lncRNA HOTAIR has been reported to be
aberrantly upregulated in several types of cancer, including HCC,
and has roles in regulating the proliferation, apoptosis,
invasiveness and metastasis of HCC (12,13). The
data of the present study validated that the knockdown of HOTAIR
inhibited cell proliferation and migration of Huh7 cells, which was
consistent with previous studies (12,20).
However, the association between HOTAIR and HCC MDR has rarely been
reported. The present study identified that the knockdown of HOTAIR
increased the chemosensitivity of Huh7 cells to cisplatin by
reducing ABCB1 expression, indicating the role of HOTAIR in
mediating MDR in HCC. These findings revealed that the knockdown of
HOTAIR combined with cisplatin chemotherapy may serve as a
potential therapeutic strategy to reverse MDR for HCC treatment.
However, the mechanism by which HOTAIR regulates ABCB1 expression
is largely unknown. It has been confirmed that HOTAIR recruits PRC2
to regulate chromosome occupancy by enhancer of zeste 2 polycomb
repressive complex 2 subunit (EZH2; a subset of PRC2), which leads
to H3K27 trimethylation and the repression of multiple genes
(7). It has been reported that
inhibition of EZH2 may decrease the expression of ABCB1 as well as
ABC subfamily C member 1 and ABC subfamily G member 2 (junior blood
group) mRNA, and protein expression levels (21,22).
Together with these findings, the present data indicated that
HOTAIR may regulate ABCB1 expression in an EZH2-dependent manner.
HOTAIR represses target gene expression, including Wnt inhibitory
factor 1, by promoting histone H3K27 methylation in the promoter
region via EZH2, and then activation of Wnt/β-catenin signaling
(23). However, data from the present
and previous studies revealed that HOTAIR-knockdown or EZH2
inhibition may decrease ABCB1 expression (21,22),
indicating that HOTAIR regulated ABCB1 expression not via the
EZH2-mediated H3K27 methylation of the ABCB1 promoter, but possibly
through another mechanism.
Although the mechanism for transcriptional
regulation of ABCB1 is not fully understood, a number of
transcription factors, including Ras GTPase, SP1 transcription
factor, nuclear factor-κB and STAT3 have been reported to be
involved. STAT3, a transcription factor that participates in
numerous cytokine signaling, has been demonstrated to be
constitutively activated by tyrosine phosphorylation in HCC
(24). Activation of STAT3 is
associated with resistance of tumor cells to chemotherapeutic
agents (25), and the downregulation
of STAT3 is able to overcome chemoresistance (16). The present study identified that STAT3
activation had a mechanistic role in HOTAIR-mediated ABCB1
expression and MDR of HCC. Knockdown of HOTAIR reduced levels of
p-STAT3 and increased chemosensitivity to cisplatin. Inhibition of
STAT3 phosphorylation reduced HOTAIR-mediated ABCB1 expression in
the present study. Furthermore, it has been reported that pSTAT3
directly mediates ABCB1 transcription, and regulates multidrug
efflux in breast and ovarian cancer cells (19). Thus, the data from the present study
may explain the association between HOTAIR and MDR of HCC, and also
revealed the possible underlying mechanism that HOTAIR mediated
chemoresistance to cisplatin in HCC through activating the
STAT3/ABCB1 signaling pathway. In addition, the findings of the
present study added HOTAIR to the increasing list of upstream
pathway leading to STAT3 activation, including interleukin-6,
leukemia inhibitory factor, cytoplasmic tyrosine-protein kinase BMX
and various receptor tyrosine kinases (23,25).
However, the molecular mechanism by which HOTAIR regulates STAT3
activation remains unclear. It has been reported that HOTAIR in
laryngeal squamous cell cancer promoted the methylation of
phosphatase and tensin homolog (PTEN), a negative regulator for
phosphoinositide-3 kinase (PI3K)/RAC serine/threonine protein
kinase (AKT) signaling pathway (26).
HOTAIR-mediated PTEN methylation resulted in downregulation of PTEN
mRNA and protein, leading to the activation of PI3K/AKT signaling.
There are a number of negative regulators for STAT3 pathway,
including suppressor of cytokine signaling 3 (SOCS3) (27); thus, further studies are required to
confirm whether HOTAIR is able to regulate STAT3 activation through
SOCS3 methylation.
In conclusion, the present study identified that
HOTAIR-knockdown decreased cellular proliferation and cisplatin
resistance of human Huh7 HCC cells. In addition, HOTAIR-knockdown
resulted in a decrease in STAT3 activity and ABCB1 expression, and
increased chemosensitivity to cisplatin. Furthermore, inhibition of
STAT3 phosphorylation reduced HOTAIR-mediated ABCB1 expression and
chemoresistance. The findings of the present study indicate that
HOTAIR may have a role in mediating MDR by regulating STAT3/ABCB1
signaling, and identify HOTAIR as a potential novel therapeutic
target to reverse MDR in HCC.
Acknowledgements
The present study was supported by The National
Natural Science Foundation of China (grant no. 81000889), The
Science and Technology Planning Project of Guangdong Province,
China (grant no. 2014A020212094), The Natural Science Foundation of
Guangdong Province, China (grant no. 2016A030313218), The Project
of Sun Yat-Sen Memorial Hospital (grant no. YXQH201704), The Key
Laboratory of Malignant Tumor Molecular Mechanism and Translational
Medicine of Guangzhou Bureau of Science and Information Technology
[grant no. (2013) 163] and The Key Laboratory of Malignant Tumor
Gene Regulation and Target Therapy of Guangdong Higher Education
Institutes (grant no. KLB09001).
References
|
1
|
Okuda K: Hepatocellular carcinoma. J
Hepatol. 32 Suppl 1:S225–S237. 2000. View Article : Google Scholar
|
|
2
|
Chiba T, Iwama A and Yokosuka O: Cancer
stem cells in hepatocellular carcinoma: Therapeutic implications
based on stem cell biology. Hepatol Res. 46:50–57. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Warmann S, Hunger M, Teichmann B, Flemming
P, Gratz KF and Fuchs J: The role of the MDR1 gene in the
development of multidrug resistance in human hepatoblastoma:
Clinical course and in vivo model. Cancer. 95:1795–1801. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhuo L, Liu J, Wang B, Gao M and Huang A:
Differential miRNA expression profiles in hepatocellular carcinoma
cells and drug-resistant sublines. Oncol Rep. 29:555–562. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Roninson IB, Chin JE, Choi KG, Gros P,
Housman DE, Fojo A, Shen DW, Gottesman MM and Pastan I: Isolation
of human mdr DNA sequences amplified in multidrug-resistant KB
carcinoma cells. Proc Natl Acad Sci USA. 83:pp. 4538–4542. 1986,
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ye CG, Yeung JH, Huang GL, Cui P, Wang J,
Zou Y, Zhang XN, He ZW and Cho CH: Increased glutathione and
mitogen-activated protein kinase phosphorylation are involved in
the induction of doxorubicin resistance in hepatocellular carcinoma
cells. Hepatol Res. 43:289–299. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rinn JL, Kertesz M, Wang JK, Squazzo SL,
Xu X, Brugmann SA, Goodnough LH, Helms JA, Farnham PJ, Segal E and
Chang HY: Functional demarcation of active and silent chromatin
domains in human HOX loci by noncoding RNAs. Cell. 129:1311–1323.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim K, Jutooru I, Chadalapaka G, Frank J,
Burghardt R, Kim S and Safe S: HOTAIR is a negative prognostic
factor and exhibits pro-oncogenic activity in pancreatic cancer.
Oncogene. 32:1616–1625. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kogo R, Shimamura T, Mimori K, Kawahara K,
Imoto S, Sudo T, Tanaka F, Shibata K, Suzuki A, Komune S, et al:
Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin
modification and is associated with poor prognosis in colorectal
cancers. Cancer Res. 71:6320–6326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fu WM, Zhu X, Wang WM, Lu YF, Hu BG, Wang
H, Liang WC, Wang SS, Ko CH, Waye MM, et al: HOTAIR mediates
hepatocarcinogenesis through suppressing miRNA-218 expression and
activating P14 and P16 signaling. J Hepatol. 63:886–895. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ishibashi M, Kogo R, Shibata K, Sawada G,
Takahashi Y, Kurashige J, Akiyoshi S, Sasaki S, Iwaya T, Sudo T, et
al: Clinical significance of the expression of long non-coding RNA
HOTAIR in primary hepatocellular carcinoma. Oncol Rep. 29:946–950.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gao JZ, Li J, DU JL and Li XL: Long
non-coding RNA HOTAIR is a marker for hepatocellular carcinoma
progression and tumor recurrence. Oncol Lett. 11:1791–1798.
2016.PubMed/NCBI
|
|
15
|
Li H, An J, Wu M, Zheng Q, Gui X, Li T, Pu
H and Lu D: LncRNA HOTAIR promotes human liver cancer stem cell
malignant growth through downregulation of SETD2. Oncotarget.
6:27847–27864. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Real PJ, Sierra A, De Juan A, Segovia JC,
Lopez-Vega JM and Fernandez-Luna JL: Resistance to chemotherapy via
Stat3-dependent overexpression of Bcl-2 in metastatic breast cancer
cells. Oncogene. 21:7611–7618. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou JJ, Deng XG, He XY, Zhou Y, Yu M, Gao
WC, Zeng B, Zhou QB, Li ZH and Chen RF: Knockdown of NANOG enhances
chemosensitivity of liver cancer cells to doxorubicin by reducing
MDR1 expression. Int J Oncol. 44:2034–2040. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bourguignon LY, Peyrollier K, Xia W and
Gilad E: Hyaluronan-CD44 interaction activates stem cell marker
Nanog, Stat-3-mediated MDR1 gene expression and ankyrin-regulated
multidrug efflux in breast and ovarian tumor cells. J Biol Chem.
283:17635–17651. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu Y, Zhang L, Zhang L, Wang Y, Li H, Ren
X, Wei F, Yu W, Liu T and Wang X: Long non-coding RNA HOTAIR
promotes tumor cell invasion and metastasis by recruiting EZH2 and
repressing E-cadherin in oral squamous cell carcinoma. Int J Oncol.
46:2586–2594. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Y, Liu G, Lin C, Liao G and Tang B:
Silencing the EZH2 gene by RNA interference reverses the drug
resistance of human hepatic multidrug-resistant cancer cells to
5-Fu. Life Sci. 92:896–902. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fan TY, Wang H, Xiang P, Liu YW, Li HZ,
Lei BX, Yu M and Qi ST: Inhibition of EZH2 reverses
chemotherapeutic drug TMZ chemosensitivity in glioblastoma. Int J
Clin Exp Pathol. 7:6662–6670. 2014.PubMed/NCBI
|
|
23
|
Ge XS, Ma HJ, Zheng XH, Ruan HL, Liao XY,
Xue WQ, Chen YB, Zhang Y and Jia WH: HOTAIR, a prognostic factor in
esophageal squamous cell carcinoma, inhibits WIF-1 expression and
activates Wnt pathway. Cancer Sci. 104:1675–1682. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang L, Yang Z, Ma A, Qu Y, Xia S, Xu D,
Ge C, Qiu B, Xia Q, Li J and Liu Y: Growth arrest and DNA damage
45G down-regulation contributes to Janus kinase/signal transducer
and activator of transcription 3 activation and cellular senescence
evasion in hepatocellular carcinoma. Hepatology. 59:178–189. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bhardwaj A, Sethi G, Vadhan-Raj S,
Bueso-Ramos C, Takada Y, Gaur U, Nair AS, Shishodia S and Aggarwal
BB: Resveratrol inhibits proliferation, induces apoptosis and
overcomes chemoresistance through down-regulation of STAT3 and
nuclear factor-kappaB-regulated antiapoptotic and cell survival
gene products in human multiple myeloma cells. Blood.
109:2293–2302. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li D, Feng J, Wu T, Wang Y, Sun Y, Ren J
and Liu M: Long intergenic noncoding RNA HOTAIR is overexpressed
and regulates PTEN methylation in laryngeal squamous cell
carcinoma. Am J Pathol. 182:64–70. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ogata H, Chinen T, Yoshida T, Kinjyo I,
Takaesu G, Shiraishi H, Iida M, Kobayashi T and Yoshimura A: Loss
of SOCS3 in the liver promotes fibrosis by enhancing STAT3-mediated
TGF-beta1 production. Oncogene. 25:2520–2530. 2006. View Article : Google Scholar : PubMed/NCBI
|