According to World Cancer Report cervical cancer or
cancer of the uterine cervix is both the second-most common cause
of cancer and the fourth-most common cause of death from cancer in
women. It has also been reported that in the year 2012, an
estimated 528,000 cases has resulted 266,000 deaths worldwide. It
has been observed that the most common causative agent of cervical
cancer is by a virus called human papilloma virus, or HPV. The
progression of cervical carcinoma occurs from normal to low grade
squamous intra-epithelial lesion (LSIL) then to high grade squamous
intra-epithelial lesion (HSIL) to carcinoma in situ (CIS)
and finally to metastatic cancer (1).
Oncogenic human papillomaviruses (HPVs), mostly HPV types-16, 18,
31, 33 are related with invasive cervical cancer lesions. HPV has
its viral genome present in the 72-capsomere capsid. The genome is
divided into three regions which include-the long control region
(LCR) without coding potential; the region of early proteins (E
protein) (E1-E7), and the region of late proteins (L1 and L2)
(2) LCR, early region and late region
regulates DNA replication, oncogenesis, cellular transformation and
viral capsule formation respectively. There are over 200 different
varieties of HPV (3). The E proteins
of HPV are used to determine the risk factor involved in cervical
cancer. HPV-6, 11, 42, 43 and 44 are low risk types as they are
found in benign tumours; HPV-16, 18, 31, 33, 35, 51, 52 and 58 are
high risk types as they are found in both in benign tumors and
invasive cancers. The E6 and E7 proteins of the oncogenic HPV
interacts with cellular proliferation and apoptotic mechanisms at
well-defined targets, the primary targets are p53 and
retinoblastoma (Rb) tumour suppressor proteins, respectively
(4,5).
The E1 and E2 proteins mediate viral replication and act to recruit
cellular replication factors to viral replication origins (6).
A change in genomic DNA and chromatin resulting in
alteration of gene expression which is somatically heritable but
does not involve DNA sequence changes is defined as an epigenetic
phenomenon. A post-replicative DNA modification that occurs on the
5′-position of cytosine rings located in CpG dinucleotides is
termed as DNA methylation. It is the most studied epigenetic change
which influence gene expression (7).
The reasons for epigenetic changes may be a regular or natural
occurrence but can also be influenced by several factors including
age, the environment, lifestyle, and disease state. Recently it has
been established that in cervical cancer epigenetic changes play a
major role in its development (8–11). DNA
methylation may be useful as a marker for early detection or
predicting the risk of cancer precursors. The existence of HPV
alone is not sufficient to cause cervical cancer; presence of DNA
methylation in cervical cancer and precancerous lesions support a
role for this phenomenon in cervical cancer development (12). This is mainly because there is
obscurity in the molecular processes which is the main cause of
futility in HPV-mediated cervical cancer. During all the stages of
cervical cancer a numerous epigenetic alterations are observed
which include DNA hypomethylation, hypermethylation of tumour
suppressor genes and histone modifications (12). DNA methylation is done by an enzyme
known as DNA methyl transferase (DNMTs) which regulates the gene
expression in prokaryotes, eukaryotes and viruses (1). It is a reversible reaction catalyzed by
DNMTs which conserve the prototype of methylation during each
cellular division. The two de novo methyl transferases are DNMT3a
and DNMT3b. DNMTs add a methyl group onto carbon 5 of cytosine
residues adjacent to guanine residues (5′-CpG-3′), which mainly
occurs in CpG islands. CpG islands are generally found in the
promoter regions of protein-coding genes, and expression is
silenced upon their methylation. Non-coding genes, such as
microRNAs (miRNAs), are also susceptible to regulation by
methylation (13). The epigenome
could be targeted by environmental factors such as dietary methyl
donors. CpG methylation of Type 1 HPV was observed in early
transfection studies (14). A study
on cell transfection with HPV-16 in vitro methylated genomes
proved that DNA is repressed transcriptionally (15). Some of the genes regulated by
methylaion are also involved in cervical cancer which acts as
transcription factors, viz p53, UTF1, paired box 1 (PAX1)
and TWIST1 (16–19).
During early cancer development the DNA
modifications which are generally observed are global DNA
hypomethylation in repetitive regions and hypermethylation in CpG
island regions of tumour-suppressor gene promoters. Cervical cancer
is correlated with the silencing of tumour-suppressor genes through
DNA hypermethylation. However, silencing of tumour suppressor
miRNAs through hypermethylation of CpG islands in their promoter
regions has also been a reason for carcinogenesis (20,21). Wang
et al, in the year 2013 had reported about the role of DNMT1
in cervical cancer (22). From in
vitro and in vivo studies performed it was established
that low levels of serum folate and high expression of DNMT1
protein or mRNA were extensively related with cervical
carcinogenesis. Integration of high risk-HPV DNA like HPV-16 and
−18 into the host's genome is an essential step in cervical
carcinogenesis (23). The HPV-16, 18
DNA gets methylated after it integrates into the human genome
(24).
A miRNA is a small non-coding (containing about 22
nucleotides) RNA generally found in plants, animals and some
viruses. Its main purpose is RNA silencing and post-transcriptional
regulation of gene expression (25,26). In
cervical cancer high risk HPV (HR-HPV) plays a role in the
deregulation of miRNA gene methylation. In patients and cervical
cancer cell lines (HeLa, SiHa, CaSki and C33A) it has been observed
that silencing of tumour-suppressor miRNAs through aberrant
promoter methylation favours cervical carcinogenesis. It has been
proposed that HR-HPV can lead to modifications in the methylation
pattern of miRNA promoters. After cervical cancer cell lines were
subjected to treatment with hypomethylating agents, decreased
methylation levels were found for certain miRNAs, which resulted in
their increased expression and associated decreased expression of
their target genes It is possible that HR-HPV genotypes are
involved in the methylation processes of miRNAs in cervical cancer
(27–30).
Basal transcription machinery and accessibility of
transcription factors are regulated by post-translational
modification of histones bound to promoter regions which control
gene expression. Acetylation and methylation as well as
phosphorylation, ubiquitination, and sumoylation are different
types of modifications included (6).
The expression of tumour suppressor genes, oncogenes and growth
factors are regulated by many histone-modifying enzymes such as the
histone deacetylases (HDACs), sirtuins, and histone
acetyltransferases (HATs). Thus, dysregulation and mutation of
expression of histone modifiers leads to epigenetic control of gene
expression (31,32).
The most important oncogenes of HPV are E6 and E7
genes which encode oncoproteins that cause transformation of the
host cell. Transcription of E6 and E7 oncogenes has always been
observed in cervical carcinomas, being the first indication of a
main role of these viral genes in HPV mediated cervical cancer
(33). Oncogenic properties of E6 and
E7, as well as their effects on p53 and Rb protein (pRb), have
provided the general basis for further investigations of the role
of HPV in carcinogenesis in the HPV-infected cervix. The oncogenic
protein E6 and E7 of HPV initiate dysregulation of cellular
proliferation and apoptotic machinery by targeting tumour
suppressor proteins such as p53 and Rb (34,35).
Several studied reported that HPV interferes with
the cellular DNA methylation machinery; in order to disguise itself
or as part of its viral cycle (50–52).
During HPV 16 infection, cellular DNA undergoes epigenetic
alterations induced by the E6 and E7 oncoproteins (51,53,54). Hence
DNA methylation might serve as a defence mechanism of host cell to
silence viral DNA (50,55). HPV encoded E6 and E7 interacts with
p53 and E2F, inducing the expression of DNMT1 resulting in aberrant
methylation. In HPV mediated carcinogenesis, E6/E7 expression is
also regulated by E2 protein and other cellular transcription
factors (NF1, AP1, KRF1, Oct1, Sp1, and YY1). In HR-HPV E6
oncoproteins increase the expression and activity of DNMT1 by
degrading p53 (54). It has been
reported that knockdown of E6 is associated with an increase in p53
and a decrease in DNMT1 expression in cervical cancer cell lines
SiHa and CaSki (51,54). p53 binds to the Sp1 and
chromatin-remodeling proteins, and this resulting complex further
binds to the promoter region of DNMT1. This resulting complex
prevents Sp1 from activating the transcription of DNMT1 (56).
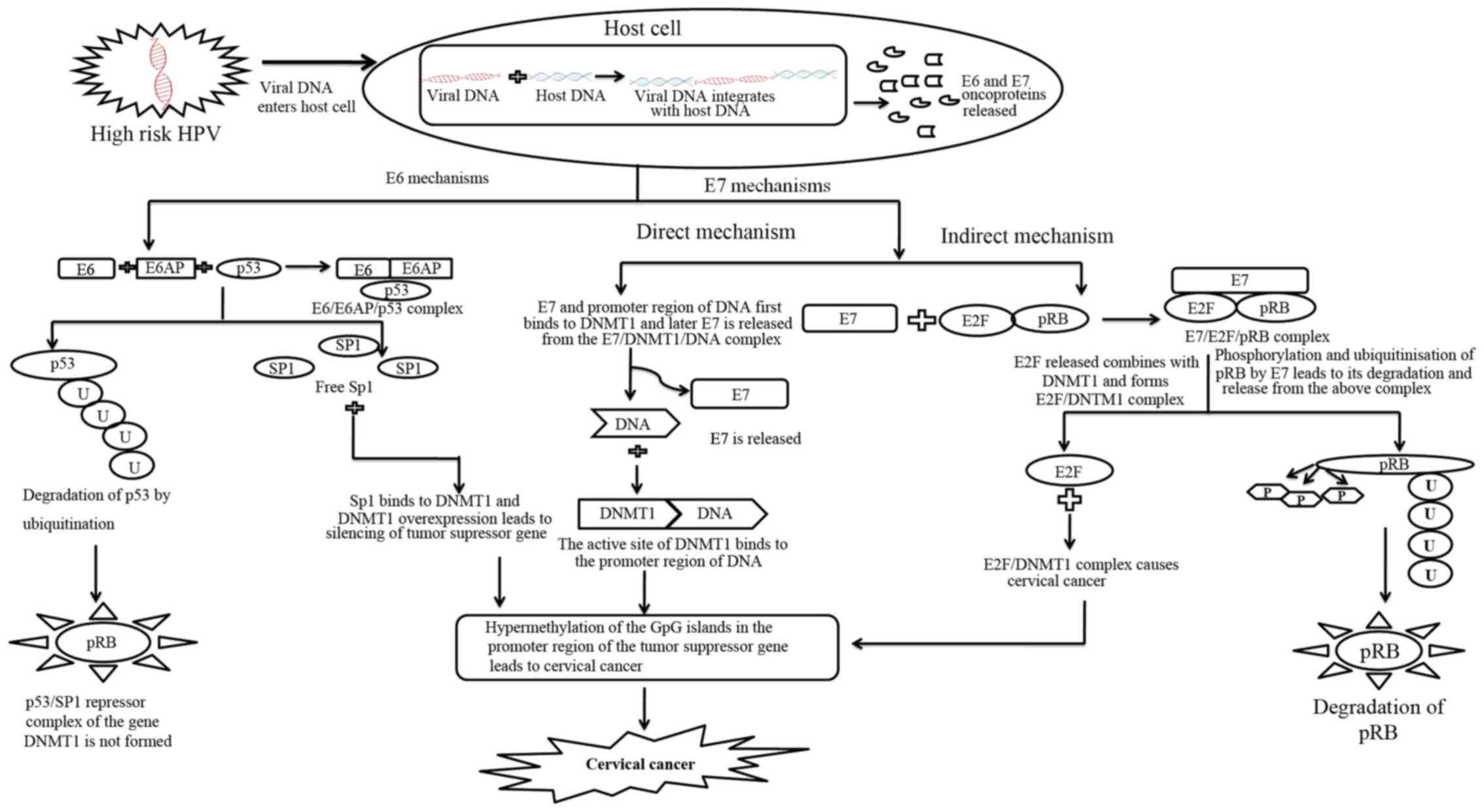
In addition DNMT1 expression is modulated by E7 in
two distinct ways. In the indirect mechanism E7 binds to pRb and
releases the transcription factor E2F. As E2F is present at the
transcription start site of DNMT1 the release of E2F activates the
promoter of DNMT1. In the direct mechanism E7 binds directly to
DNMT1 (53). This binding causes
conformational changes in DNMT1 exposing the active site and
binding of E7/DNMT1 to DNA. A stable DNMT1/DNA complex is formed
after E7 dissociates from the complex (51). This results to methylation of the
genome due to silencing of the tumour suppressor gene and hence
causes cervical cancer.
As stated earlier the oncoproteins of HPV E6 and E7
are responsible for the carcinogenicity of the virus. The promoter,
enhancer and silencer sequences in the HPV LCR controls the
expression of E6 and E7 (57).
Hypomethylation of the promoter region is considered to be an
important part of activation of gene in the early stage of cancer
which is also a very frequent affair. A study was conducted
recently to compare normal samples with cervical cancer samples
which reported that cervical cancer is related with progressive
hypomethylation (48). It has been
established by various studies that in cervical cancer both host
and HPV genome are hypomethylated. In cervical cancer not only the
HPV genome undergoes hypomethylation but Rap guanine nucleotide
exchange factor 1 (RAPGEF1), long interspersed elements (LINE1) and
Cancer Antigen Gene (CAGE) promoter are also hypomethylated
(58–60).
Like hypomethylation, hypermethylation is also an
early and frequent event in cervical cancer. It is mainly related
with the CpG island and associated with promoter region which leads
to gene silencing in cervical carcinogenesis (20). Badal et al, 2003 illustrated
that hypermethylation of HPV genome suppresses the abnormal growth
of normal tissue to malignant tissue where as hypomethylation
promotes carcinogenesis (61).
Research on epigenetic for the past few decades in cervical cancer
has established the fact that methylation of DNA can be used for
early recognition, analysis and maturity of unique curative
approaches along with its pathogenic role (62). Signalling pathways such as cell cycle,
cell to cell adhesion, Apoptotic pathway and cell signalling
pathways are effected by hypermethylation of HPV in cervical cancer
(20). Therefore, the epigenetic
pathways that are active in the host cell also cause modification
of the viral genome.
In both squamous cell carcinoma (SCC) and
adenocarcinoma, the frequently methylated genes are cell adhesion
molecule 1 (CADM1), cadherin 1 (CDH1), death-associated protein
kinase 1 (DAPK1), EPB41L3, FAM19A4, myelin and lymphocyte (MAL),
PAX1, PR domain containing 14 (PRDM14) and telomerase
reverse transcriptase (TERT) (63). Recently a wide profiling of
methylation in cervical intraepithelial neoplasia (CIN) lesions and
cervical cancer has been conducted which requires proper
justification and further studies. Tumour suppressive activities in
cervical cancer has been observed in these subset of
genes-CADM1, WNT signalling pathway inhibitor 3
(DKK3), MAL, secreted frizzled-related protein 2
(SFRP2) and C13ORF18 (also known as
KIAA0226L). Other than these findings methylation events and
their related biological relevance in cervical cancer remains
indefinable (64).
A combined mechanism involving both E6 and E7 acting
together on DNMT1 has been shown in Fig.
1. This mechanism is most likely to be occurring in vivo
when both E6 and E7 are over expressed. Thus, a combined
mechanistic action of both E6 and E7 results in hypermethylation of
many tumour suppressor genes leading to unregulated cell
proliferation. It might be possible that through indirect
mechanisms of E6/E7, there is an over expression of DNMT1, which is
bound directly by E7 to cause hypermethylation of the gene
promoters. Therefore, the indirect mechanism is likely to precede
the direct pathway.
For cell cycle functioning of cyclin dependent
kinases (CDKs) its proper expression is essential (65). In cervical carcinogenesis, CDK gene is
not included in HPV induced cervical cancer but the expression of
this gene is observed to be abnormal. P16INK4a is a tumour
suppressor protein which acts as an inhibitor of CDK4 and CDK6,
which initiate the phosphorylation of the Rb tumour suppressor
protein, Rb. E7 oncoprotein plays an important role in disrupting
the normal cell cycle when it binds to pRB and resulting in
activation of p16INK4a which causes cell death by arresting cells
in G1 phase. Thus, p16INK4a probable physiological role is in the
implementation of irreversible growth arrest termed cellular
senescence (66). The normal cell
cycle pathway is disrupted when the oncoprotein E7 of HPV binds to
pRB and disrupts its function. Hypermethylation results in
downregulation of E7 leading to activation of p16INK4A (1). In the year 2011, Huang et al
reported that hypermethylation was present in 17.6% of CIN I, 42.1%
of CIN II, 55.0% of CIN III, and 65.0% of invasive cancers
(67). Additionally, this can be used
as a biomarker to foresee the development of low grade dysplasia
and reappearance of high grade intraepithelial neoplasias (68). In HPV induced cervical cancer the
downregulation of Cyclin A1 expression is observed due to its
methylation. Other than these above mentioned proteins, fragile
histidine triad (FHIT) is another protein related to cell cycle
regulation and apoptosis. In HPV related cervical cancer this
protein (FHIT) is inactivated. Promoter hypermethylation is the
reason for silencing of the gene rather than genetic alterations
(69).
A transmembrane glycoprotein called E-cadherin acts
as a mediator for calcium-dependent interactions between adjacent
epithelial cells. In cervical cancer, previous data showed that the
expression of E-cadherin is reduced at both the mRNA and protein
levels. Loss of membranous E-cadherin expression was detected in in
three HPV mediated cervical cancer cell lines (HeLa, SiHa, C33A)
due to hypermethylation of CpG islands in its promoter region.
These data suggested that E-cadherin might be an important factor
in cervical cancer (70). Later it
was reported that 52.6% of gene expression of CDH1 gene is
downregulated due to promoter methylation in HPV positive cervical
cancer samples (71). Another study
conducted on a tumor suppressor gene TSLC1 gene and reported that
it is silenced in cervical cancer due to methylation of the
promoter region. Hypermethylation of TSLC1 results in
downregulation of its expression which was reported in 82 and 91%
of cervical cancer cell lines (11).
It was also stated that there was 89% epigenetic change in case of
invasive cervical cancers whereas only 26% in case of CIN III and
none in normal tissues (72). Cancer
invasion or metastasis could lead to loss of expression of CADM1
which is same as the TSLC1 gene which is associated with epithelial
cell adhesion. A study reported that HR-HPV is associated with the
frequency and density of methylation in cervical cancer. Expression
of CADM1 is reduced in the methylated regions of high grade CIN and
SCC (73). In another study related
to chemokine signalling axes stromal derived factor-1
(SDF-1)α/CXCR4, demonstrated its expression to be downregulated at
both transcript and protein level in cervical cancer cell line
(C33A a HPV positive cervical cancer cell line) and primary tumors
due to promoter hypermethylation (74).
Apoptosis or programmed cell death occurs in
multi-cellular organisms. It is a highly synchronized and
controlled process which is essential in an organism's lifecycle.
For cancer to develop in an organism the cells should be resistant
to apoptosis, chemotherapy and radiotherapy. Recognising and
identifying the genes which are related to apoptosis might help
scientist in early development of new therapeutic approaches to
cure cervical cancer.
Methylation of decoy receptor (DcR1 and DcR2) were
found in HPV related cervical cancer patients which resulted in
silencing and inhibition of apoptosis (75). Recent study showed that PRDM14 is
downregulated in cervical cancer HPV positive cell lines due to
promoter hypermethylation and its abnormal levels resulted in
apoptosis (76). In many tumours it
is observed that transcriptional silencing and inactivation of
tumour suppressor genes is due to the hypermethylation of CpG
islands. It was reported that p73 and p53 genes were
hypermethylated among the 50 CpG islands of p21, p27, p57, p53, p73
and Rb1 genes in HPV positive cervical cancer samples as compared
to control samples (17). DAPK is a
pro-apoptotic serine/threonine kinase which plays a major role in
metastasis and tumour pathogenesis (77). This gene is located in the chromosome
number 9q34.1. In cervical cancer this gene was reported to be
inactivated due to hypermethylation of the promoter region
(78,79).
The retinoic acid receptor β2 (RARβ2) belongs to the
nuclear superfamily and is a type of retinoic acid which is
regulated by tumour suppressor genes. Re-expression of this gene
was observed due to methylation of the promoter region in the
cervical cancer cell lines SiHa and HeLa when administered with
natural compounds curcumin and genistein (80). Thus, it can be stated that RARβ2 might
be used as a biomarker for epigenetic therapy by targeting natural
compounds and is related to HPV positive cervical cancer cell
lines. The RAS association domain family 1 (RASSF1) is a tumour
suppressor gene. When it is inactivated the gene could be
correlated with the hypermethylation of its CpG-island promoter
region. RASSF1 gene is mainly of two types-RASSF1A and RASSF1C,
which are transcribed from separate promoters (81). Cell death is activated during
transient over-expression when human scaffold protein CNK1, a
c-Raf1 binding protein necessary for Ras-induced Raf activation,
binds the human tumor suppressor RASSF1A and is stably reintroduced
into human tumor cell lines. RASSF1 gene is not only related to
cervical cancer but is related to different other cancer like colon
cancer. Adenomatous polyposis coli (APC) is a tumor suppressor
protein complex; cell overgrowth is prevented by regulating the
genes that encourage cell division by controlling beta-catenin
which in turn is controlled by APC. Over-expression of β-catenin
targets, including cyclin D, cMyc, ephrins, and caspases are caused
due to inactivation or loss of APC expression. Certain proteins
such as microtubules, the components for Wnt/Wg pathway-catenin and
axin, EB1 and IQGAP1 which are cytoskeletal regulators and the Rac
guanine-nucleotide-exchange factor (GEF) Asef1 have their binding
site in APC which is a multi-domain protein. Mutation cluster
region is the region where ~60% of cancer-linked APC mutations
occur. For the tumour suppressor activity of APC interaction with
β-catenin or microtubules is considered to be necessary (82). An in vitro study documented
that in HPV induced cervical carcinoma cell line HeLa 60% of APC
was hypermethylated as compared to 13% in SCCs cell line SiHa
(83). When cervical carcinoma cell
line HeLa was de-methylated with the drug hydralazine it resulted
in re-expression of methylation (84). Thus, methylation is a potential
epigenetic marker for cervical adenocarcinoma. From the above
mentioned studies it has been observed that epigenetic alterations
such as hypermethylation or de-methylation of the promoter region
of a gene can act as a marker to predict cervical cancer in the
early stage of its onset and cure it accordingly. In 30% of the
tumour in cervical cancer the genes have been silenced due to
methylation (85,86).
A molecule which secreted by a tumour or a specific
response of the body due to the presence of cancer or a process
which indicates the presence of cancer clinically is known as a
cancer biomarker. The types of biomarkers used to detect cancer
are- genetic, epigenetic, proteomic glycomic or imaging biomarker.
Recently to predict, evaluate and treat the outcomes of cancer,
biomarkers have been identified to conduct this process. The
biomarkers which show response to anticancer therapy in SCC are-
checkpoint with forkhead and ring finger (CHFR); Werner DNA
helicases (WRN); hypoxia inducible factor-1α subunit (HIF-1α),
which is associated with hypoxic response; epidermal growth factor
receptor (EGFR), which may be a molecular target; and
cyclooxygenase-2 (COX-2), which predicts radiation sensitivity
(87).
However, it is thought that viral gene transcription
is caused due to methylation of viral. The common methylating genes
for samples with invasive SCC and adenocarcinoma have the following
biomarkers-CADM1, CDH1, DAPK1, erythrocyte membrane protein band
4.1-like 3 (EPB41L3) family with sequence similarity 19, chemokine
(CC motif)-like, member A4 (FAM19A4) myelin and lymphocyte protein
(MAL), PAX1, PR domain-containing 14 (PRDM14), and TERT (63). In case of cervical cancer frequently
methylated genes can serve as molecular markers for the detection
of neoplastic cells in body fluids such as urine and plasma. It can
be served as non-invasive early diagnostic tool to detect cervical
cancer. Different stages of cervical carcinogenesis can be
represented by different methylation pattern of tumour suppressor
genes in serum as well as in tissue samples (88,89).
Single epigenetic biomarkers are not adequately sensitive to
predict oncogenic progression, thus panels of candidate genes can
improve sensitivity. The panel of gene mainly includes genes having
high CpG methylation rate such as DAPK1, CDH1, IGSF4, CALCA, which
were supplemented with genes of lower methylation rate such as
RARβ2, or APC (10,90,91). The
Table I shows different classes of
genes along with their nature of DNA methylation and how the gene
expression gets modulated giving rise to cervical cancer phenotype.
A combination of some of these genes could be useful biomarkers for
cervical cancer. Aberrant DNA methylation pattern in urine can be
compared with exfoliated cervical cells but sensitivity is lower
(92).
Recently there have been few studies in cervical
cancer which are related to histone modification and alteration.
Although analysis of this relation between cervical cancer and
histone modification is at its beginning stage, some records
regarding modification of the histones in cervical cancer samples
may help in the identification of this cancer. The molecular level
of diagnosis of chromatin modification has still not been fully
established.
It was reported that in cervical cancer the
expression levels of HDACs increases and due to continuous
hypomethylation of DNA Sequence, tumor cells lose monoacethylated
and trimethylated histone H4 form (93). In the same year it was also published
that silencing of a DNA protein O6-methylguanine DNA
methyltransferase (MGMT) which is important for genome stability
was caused due to histone modification in cervical cancer (94). Furthermore it was reported that
histone deacetylation caused repression of Wnt signalling pathway
antagonist dickkopf-related protein 1 (DKK-1) in cervical cancer
cell lines (95) by promoter CpG
hypermethylation and histone deacetylation. It is generally
observed that in HPV infected cervical cancer lesions and warts are
formed. These deformities are mainly related to lack of oxygen
present in tissues because of amplified expression of
pro-angiogenic factors through the activity of oxygen sensitive
HIF-1α. It was stated in another study that the transcriptional
activity of HIF-1 is enhanced by the oncoprotein E7 of HPV while
the suppressive action of p53 gene is neutralized by E6. Thus, they
concluded that E7 oncoprotein in HPV mediated cervical cancer can
activate the function of a key transcription factor mediating
hypoxic response by displacing histone deacetylases HDAC1, HDAC4,
and HDAC7 from HIF-1α (96). Histone
modification and DNA mehtylation caused silencing of proliferation
repressor protein osteo-protegerin (OPG) and RARβ2 in another study
(97,98).
The main reason reported for the cause of
progression in cervical cancer from CIN I to CIN II and CIN III has
been reported to be phosphorylated and acetylated forms of histone
H3 (99). For gene transcription,
balance between HDACs and HATs plays a very important function
which should be maintained in normal cells so that uncontrolled
cell death and proliferation is under control (100). But in case of HPV mediated cervical
cancer the E6 and E7 oncoprotein disrupts this balance between
HDACs and HATS and thus uncontrollable cell growth and cell
proliferation of cancer cell occurs.
Epigenetic therapy is a new and fast growing area of
medicines with the use of drugs to cure epigenetic defects. This
kind of therapy is considered to be very potential as it can easily
reverse the genetic defects and prevent diseases with the help of
medical involvement (101).
Methylation of the gene promoter is targeted for cancer therapy by
reactivation of tumour suppressor genes (102). It has been established through in
vitro and in vivo clinical models that inhibition of DNA
methylation exerts anti-tumour effects; restore gene expression and
inhibit hypermethylation. HDACs is a potential candidate for cancer
treatment. Several experimental models demonstrated that HDAC
inhibition leads to induced apoptosis, or growth arrest, tumour
cell differentiation and to sensitize cells to chemotherapy or
radiation therapy (103,104). The molecular interplay between DNA
hypermethylation and histone deacetylation for modulation of gene
expression and silencing of different cancer related gene predicts
synergy of gene expression as well as for anti-tumour activity
(105). Therefore, a combination of
DNA demethylating agent with HDAC inhibitor can be a potential
target for cancer treatment.
Two main classes of epigenetic drugs, methylation
inhibitors and HDACs inhibitors, are currently in clinical trials
for cancer treatment. These drugs target the epigenome in a
non-cell specific way. They act on most or all tumour types as
deacetylation machinery or as deregulators of methylation.
Hydralazine, a peripheral vasodilatator drug is a stable DNA
methylation inhibitor having negligible toxicity. This drug has
been widely used to treat hypertension. Hydralazine treatment can
induce demethylation of promoter sequences, reactivation of mRNA
transcription and protein synthesis of tumour suppressor genes in
in vivo as well as in vitro clinical models (106). A randomized phase III trial,
conducted by Coronel et al, compared chemotherapy alone with
epigenetic therapy with hydralazine and the deacylation agent
valproate plus cisplatin and topotecan for treatment of advanced
cervical cancer (107). The outcome
of the study suggested that patients treated with combination
therapy had longer progression free survival than did patients
treated with chemotherapy alone. Another widely used anti-epileptic
drug, Valproic acid (VPA) is an effective inhibitor of HDACs. VPA
has potent anti-tumour effects by modulating multiple cellular
pathways including cell cycle arrest, apoptosis, angiogenesis,
metastasis, differentiation and senescence in a variety of in
vitro and in vivo systems (108). It leads to growth inhibition in
HeLa, SiHa and CaSki cells of cervical epithelial origin. This
anti-tumour effect of VPA in cervical cancer is mediated by
hyperacetylation of p53 protein which protects it from degradation
by E6 (109). In addition, Apicidin,
a cyclic peptide HDAC downregulate DNMT1 gene expression in HeLa
cells (110). In a recent study
conducted by Chen et al, in the year 2015 observed there
were methylation changes in cervical cancer cell line S3 compared
to control cell line SiHa. When S3 cell line was treated with a
demethylating agent it reversed the resistance and restored the
sensitivity of the cell line to cisplatin, taxol and oxaliplatin
drugs. Hence, it was concluded that methylation is caused due to
resistance towards drug. This property of the cervical cancer cell
line could act as a biomarker and also a therapeutic target to
detect drug resistance in cervical cancer (111). Epigenetic therapy has certain
limitations due to the fact that both DNA methylation and HDAC
inhibitors may activate oncogenes because of non specific targeting
which can lead to tumour progression. Furthermore, once cured
epigenetic states may revert back to its previous state due to the
reversible nature of DNA (112).
More investigations are needed for specific targeting of the drugs
as well as to maintain the cured epigenetic state and prevent
reversal.
The crucial role of epigenetics in development of
cervical cancer is well established. Apart from the drugs that
target epigenetic machinery (HDAC & DNMT1), new approaches need
to be developed to target cancer cells specifically. Using
anti-HDAC inhibitors can have a bystander effect on normal, healthy
cells causing undue stress to them. Other than drugs,
liposome-based cell permeable peptides targeting E6/E7 could be
used. These peptides would bind to E6/E7 and prevent their
interaction with other cellular components including the epigenetic
machinery. Blocking E6/E7 would be the most effective approach as
this will have a cascading effect on all the cellular pathways that
are modulated by E6/E7. This approach requires more studies and
testing to optimize the amount of peptide to be delivered inside
the infected cell that would be sufficient to block E6/E7. These
peptides can be periodically injected at the site of cancer and the
patient can be monitored over time for cancer regression.
This review primarily focuses on modulation of DNA
methylation by HPV E6 and E7 oncoproteins in cervical cancer. There
are several reviews on DNA methylation and cancers but this review
focuses on the key mechanisms that are likely to operate during HPV
E6 and E7 mediated modulation of gene expression. The review
discusses a wide variety of genes involved in diverse cellular
functions that are affected in cervical cancer. The review also
discusses various implications in clinical applications of
epigenetic based therapy and what classes of drugs would be
suitable for epigenetic therapy. Cervical cancer remains one of the
lethal cancers of woman worldwide and thus, there is a continuous
need to identify non-invasive biomarkers for early detection,
prognosis, selection of the treatment, prediction of treatment
response, and development of novel therapeutic strategies.
Epigenetic based therapy is a new approach toward early prognosis
and hence treatment of this lethal cancer. Understanding the role
of these epigenetic alterations in cervical carcinogenesis will
contribute to developing effective strategies in detecting and
treating this disease.
Funding from DST-SERB, Govt. of India is duly
acknowledeged.
|
1
|
Bhat S, Kabekkodu SP, Noronha A and
Satyamoorthy K: Biological implications and therapeutic
significance of DNA methylation regulated genes in cervical cancer.
Biochimie. 121:298–311. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Panatto D, Amicizia D, Bragazzi NL,
Rizzitelli E, Tramalloni D, Valle I and Gasparini R: Chapter
Eight-Human Papillomavirus Vaccine: State of the Art and Future
Perspectives. Adv Protein Chemistry Structural Biol. 101:231–322.
2015. View Article : Google Scholar
|
|
3
|
IARC Monographs on the Evaluation of
Carcinogenic Risks to Humans: Pharmaceuticals: A Review of human
carcinogens. IARC monographs on the evaluation of carcinogenic
risks to humans. 100A:(IARC Monographs). 1–401. 2012.
|
|
4
|
D'Andrilli G: Advances in cervical cancer
and ongoing clinical trialsGynecological Cancers. Giordano A and
Macaluso M: Springer International Publishing; Cham: pp. 51–64.
2016
|
|
5
|
Zhao S: Specific Type Epigenetic Changes
in Cervical CancersCancer Epigenetics, Methods in Molecular Biology
(Methods and Protocols). 1238. Verma M: Humana Press; New York, NY:
pp. 733–749. 2015
|
|
6
|
Langsfeld E and Laimins LA: Human
papillomaviruses: Research priorities for the next decade. Trends
Cancer. 2:234–240. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hu H, Shu M, He L, Yu X, Liu X, Lu Y, Chen
Y, Miao X and Chen X: Epigenomic landscape of
5-hydroxymethylcytosine reveals its transcriptional regulation of
lncRNAs in colorectal cancer. Br J Cancer. 116:658–668. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sova P, Feng Q, Geiss G, Wood T, Strauss
R, Rudolf V, Lieber A and Kiviat N: Discovery of novel methylation
biomarkers in cervical carcinoma by global demethylation and
microarray analysis. Cancer Epidemiol Biomarkers Prev. 15:114–123.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Narayan G, Arias-Pulido H, Koul S, Vargas
H, Zhang FF, Villella J, Schneider A, Terry MB, Mansukhani M and
Murty VV: Frequent promoter methylation of CDH1, DAPK, RARB, and
HIC1 genes in carcinoma of cervix uteri: Its relationship to
clinical outcome. Mol Cancer. 2:242003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Feng Q, Balasubramanian A, Hawes SE, Toure
P, Sow PS, Dem A, Dembele B, Critchlow CW, Xi L, Lu H, et al:
Detection of hypermethylated genes in women with and without
cervical neoplasia. J Natl Cancer Inst. 97:273–282. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Steenbergen RD, Kramer D, Braakhuis BJ,
Stern PL, Verheijen RH, Meijer CJ and Snijders PJ: TSLC1 gene
silencing in cervical cancer cell lines and cervical neoplasia. J
Natl Cancer Inst. 96:294–305. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lu Q, Ma D and Zhao S: DNA methylation
changes in cervical cancers. Methods Mol Biol. 863:155–176. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Trang P, Weidhaas JB and Slack FJ:
MicroRNAs and cancerThe Molecular Basis of Human Cancer. Coleman WB
and Tsongalis GJ: 2nd. Springer; New York, New York, NY: pp.
277–286. 2017, View Article : Google Scholar
|
|
14
|
Leung TW, Liu SS, Leung RC, Chu MM, Cheung
AN and Ngan HY: HPV 16 E2 binding sites 1 and 2 become more
methylated than E2 binding site 4 during cervical carcinogenesis. J
Med Virol. 87:1022–1033. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Doeberitz MVK and Vinokurova S: Compounds
and methods associated with differential methylation of human
papilloma virus genomes in epithelial cells. US Patent 12/740,986.
Filed October 31, 2008; issued September 29. 2015.
|
|
16
|
Kan YY, Liou YL, Wang HJ, Chen CY, Sung
LC, Chang CF and Liao CI: PAX1 methylation as a potential biomarker
for cervical cancer screening. Int J Gynecol Cancer. 24:928–934.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jha AK, Nikbakht M, Jain V, Sehgal A,
Capalash N and Kaur J: Promoter hypermethylation of p73 and p53
genes in cervical cancer patients among north Indian population.
Mol Biol Rep. 39:9145–9157. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guenin S, Mouallif M, Deplus R, Lampe X,
Krusy N, Calonne E, Delbecque K, Kridelka F, Fuks F, Ennaji MM and
Delvenne P: Aberrant promoter methylation and expression of UTF1
during cervical carcinogenesis. PLoS One. 7:e427042012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Missaoui N, Hmissa S, Trabelsi A, Traoré
C, Mokni M, Dante R and Frappart L: Promoter hypermethylation of
CDH13, DAPK1 and TWIST1 genes in precancerous and cancerous lesions
of the uterine cervix. Pathol Res Pract. 207:37–42. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang HJ: Aberrant DNA methylation in
cervical carcinogenesis. Chin J Cancer. 32:42–48. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liz J and Esteller M: lncRNAs and
microRNAs with a role in cancer development. Biochim Biophys Acta.
1859:169–176. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang JT, Ding L, Jiang SW, Hao J, Zhao WM,
Zhou Q, Yang ZK and Zhang L: Folate deficiency and aberrant
expression of DNA methyltransferase 1 were associated with cervical
cancerization. Curr Pharm Des. 20:1639–1646. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Berdasco M and Esteller M: Aberrant
epigenetic landscape in cancer: How cellular identity goes awry.
Dev Cell. 19:698–711. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jackson R, Rosa BA, Lameiras S, Cuninghame
S, Bernard J, Floriano WB, Lambert PF, Nicolas A and Zehbe I:
Functional variants of human papillomavirus type 16 demonstrate
host genome integration and transcriptional alterations
corresponding to their unique cancer epidemiology. BMC Genomics.
17:8512016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wilting SM, van Boerdonk RA, Henken FE,
Meijer CJ, Diosdado B, Meijer GA, le Sage C, Agami R, Snijders PJ
and Steenbergen RD: Methylation-mediated silencing and tumour
suppressive function of hsa-miR-124 in cervical cancer. Mol Cancer.
9:1672010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Botezatu A, Goia-Rusanu CD, Iancu IV,
Huica I, Plesa A, Socolov D, Ungureanu C and Anton G: Quantitative
analysis of the relationship between microRNA-124a, −34b and −203
gene methylation and cervical oncogenesis. Mol Med Rep. 4:121–128.
2011.PubMed/NCBI
|
|
29
|
Yao T, Rao Q, Liu L, Zheng C, Xie Q, Liang
J and Lin Z: Exploration of tumor-suppressive microRNAs silenced by
DNA hypermethylation in cervical cancer. Virol J. 10:1752013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wilting SM, Verlaat W, Jaspers A, Makazaji
NA, Agami R, Meijer CJ, Snijders PJ and Steenbergen RD:
Methylation-mediated transcriptional repression of microRNAs during
cervical carcinogenesis. Epigenetics. 8:220–228. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Morgan MA and Shilatifard A: Chromatin
signatures of cancer. Genes Dev. 29:238–249. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Khan SA, Reddy D and Gupta S: Global
histone post-translational modifications and cancer: Biomarkers for
diagnosis, prognosis and treatment? World J Biol Chem. 6:333–345.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ghittoni R, Accardi R, Chiocca S and
Tommasino M: The role of human papillomaviruses in carcinogenesis.
Ecancermedicalscience. 9:5262015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ruttkay-Nedecky B, Jimenez AM Jimenez,
Nejdl L, Chudobova D, Gumulec J, Masarik M, Adam V and Kizek R:
Relevance of infection with human papillomavirus: The role of the
p53 tumor suppressor protein and E6/E7 zinc finger proteins
(Review). Int J Oncol. 43:1754–1762. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mesri EA, Feitelson MA and Munger K: Human
viral oncogenesis: A cancer hallmarks analysis. Cell Host Microbe.
15:266–282. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen AA, Gheit T, Franceschi S, Tommasino
M and Clifford GM; IARC HPV Variant Study Group, : Human
papillomavirus 18 genetic variation and cervical cancer risk
worldwide. J Virol. 89:10680–10687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Evans W, Filippova M, Aragon R, Filippov
V, Reeves ME and Duerksen-Hughes P: Abstract 1828: Proteomic
analysis of the effect of E6 star expression on cellular pathways
in HPV positive SiHa and HPV negative C33A cervical carcinoma
cells. Cancer Res. 75:1828. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Illiano E, Demurtas OC, Massa S, Di Bonito
P, Consalvi V, Chiaraluce R, Zanotto C, De Giuli Morghen C,
Radaelli A, Venuti A and Franconi R: Production of functional,
stable, unmutated recombinant human papillomavirus E6 oncoprotein:
Implications for HPV-tumor diagnosis and therapy. J Transl Med.
14:2242016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Duensing S and Münger K: The human
papillomavirus type 16 E6 and E7 oncoproteins independently induce
numerical and structural chromosome instability. Cancer Res.
62:7075–7082. 2002.PubMed/NCBI
|
|
40
|
Munger K, Baldwin A, Edwards KM, Hayakawa
H, Nguyen CL, Owens M, Grace M and Huh K: Mechanisms of human
papillomavirus-induced oncogenesis. J Virol. 78:11451–11460. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kruiswijk F, Labuschagne CF and Vousden
KH: p53 in survival, death and metabolic health: A lifeguard with a
licence to kill. Nat Rev Mol Cell Biol. 16:393–405. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hengstermann A, Linares LK, Ciechanover A,
Whitaker NJ and Scheffner M: Complete switch from Mdm2 to human
papillomavirus E6-mediated degradation of p53 in cervical cancer
cells. Proc Natl Acad Sci USA. 98:pp. 1218–1223. 2001; View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Mantovani F and Banks L: The human
papillomavirus E6 protein and its contribution to malignant
progression. Oncogene. 20:7874–7887. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Paek AL, Liu JC, Loewer A, Forrester WC
and Lahav G: Cell-to-cell variation in p53 dynamics leads to
fractional killing. Cell. 165:631–642. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yim EK and Park JS: The role of HPV E6 and
E7 oncoproteins in HPV-associated cervical carcinogenesis. Cancer
Res Treat. 37:319–324. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ganguly N and Parihar SP: Human papilloma
virus E6 and E7 oncoproteins as risk factors for tumorigenesis. J
Biosci. 34:113–123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tommasino M: The human papillomavirus
family and its role in carcinogenesis. Semin Cancer Biol. 26:13–21.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yin FF, Wang N, Bi XN, Yu X, Xu XH, Wang
YL, Zhao CQ, Luo B and Wang YK: Serine/threonine kinases 31(STK31)
may be a novel cellular target gene for the HPV16 oncogene E7 with
potential as a DNA hypomethylation biomarker in cervical cancer.
Virol J. 13:602016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dueñas-González A, Lizano M, Candelaria M,
Cetina L, Arce C and Cervera E: Epigenetics of cervical cancer. An
overview and therapeutic perspectives. Mol Cancer. 4:382005.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Whiteside MA, Siegel EM and Unger ER:
Human papillomavirus and molecular considerations for cancer risk.
Cancer. 113 Suppl 10:S2981–S2994. 2008. View Article : Google Scholar
|
|
51
|
Leonard SM, Wei W, Collins SI, Pereira M,
Diyaf A, Constandinou-Williams C, Young LS, Roberts S and Woodman
CB: Oncogenic human papillomavirus imposes an instructive pattern
of DNA methylation changes which parallel the natural history of
cervical HPV infection in young women. Carcinogenesis.
33:1286–1293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Jiang P and Yue Y: Human papillomavirus
oncoproteins and apoptosis (Review). Exp Ther Med. 7:3–7. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Burgers WA, Blanchon L, Pradhan S, de
Launoit Y, Kouzarides T and Fuks F: Viral oncoproteins target the
DNA methyltransferases. Oncogene. 26:1650–1655. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Au Yeung CL, Tsang WP, Tsang TY, Co NN,
Yau PL and Kwok TT: HPV-16 E6 upregulation of DNMT1 through
repression of tumor suppressor p53. Oncol Rep. 24:1599–1604.
2010.PubMed/NCBI
|
|
55
|
Richards KL, Zhang B, Baggerly KA, Colella
S, Lang JC, Schuller DE and Krahe R: Genome-wide hypomethylation in
head and neck cancer is more pronounced in HPV-negative tumors and
is associated with genomic instability. PLoS One. 4:e49412009.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lin RK, Wu CY, Chang JW, Juan LJ, Hsu HS,
Chen CY, Lu YY, Tang YA, Yang YC, Yang PC and Wang YC:
Dysregulation of p53/Sp1 control leads to DNA methyltransferase-1
overexpression in lung cancer. Cancer Res. 70:5807–5817. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Cai Q, Lv L, Shao Q, Li X and Dian A:
Human papillomavirus early proteins and apoptosis. Arch Gynecol
Obstet. 287:541–548. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Banzai C, Nishino K, Quan J, Yoshihara K,
Sekine M, Yahata T and Tanaka K: Gynecological Cancer Registry of
Niigata: Promoter methylation of DAPK1, FHIT, MGMT, and CDKN2A
genes in cervical carcinoma. Int J Clin Oncol. 19:127–132. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Samuelsson J, Alonso S, Ruiz-Larroya T,
Cheung TH, Wong YF and Perucho M: Frequent somatic demethylation of
RAPGEF1/C3G intronic sequences in gastrointestinal and
gynecological cancer. Int J Oncol. 38:1575–1577. 2011.PubMed/NCBI
|
|
60
|
Shuangshoti S, Hourpai N, Pumsuk U and
Mutirangura A: Line-1 hypomethylation in multistage carcinogenesis
of the uterine cervix. Asian Pac J Cancer Prev. 8:307–309.
2007.PubMed/NCBI
|
|
61
|
Badal V, Chuang LS, Tan EH, Badal S, Villa
LL, Wheeler CM, Li BF and Bernard HU: CpG methylation of human
papillomavirus type 16 DNA in cervical cancer cell lines and in
clinical specimens: Genomic hypomethylation correlates with
carcinogenic progression. J Virol. 77:6227–6234. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
How Kit A, Nielsen HM and Tost J: DNA
methylation based biomarkers: Practical considerations and
applications. Biochimie. 94:2314–2337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Mersakova S, Nachajova M, Szepe P,
Kasajova PS and Halasova E: DNA methylation and detection of
cervical cancer and precancerous lesions using molecular methods.
Tumor Biol. 37:23–27. 2016. View Article : Google Scholar
|
|
64
|
Steenbergen RD, Snijders PJ, Heideman DA
and Meijer CJ: Clinical implications of (epi)genetic changes in
HPV-induced cervical precancerous lesions. Nat Rev Cancer.
14:395–405. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Whittaker SR, Mallinger A, Workman P and
Clarke PA: Inhibitors of cyclin-dependent kinases as cancer
therapeutics. Pharmacol Ther. 173:83–105. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Sherr CJ and Bartek J: Cell cycle-targeted
cancer therapies S phase: The DNA synthesis phase of the cell
cycle. 1:Annual Review of Cancer Biology. 41–57. 2017. View Article : Google Scholar
|
|
67
|
Huang LW, Pan HS, Lin YH, Seow KM, Chen HJ
and Hwang JL: P16 methylation is an early event in cervical
carcinogenesis. Int J Gynecol Cancer. 21:452–456. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Terra AP, Murta EF, Maluf PJ, Caballero
OL, Brait M and Adad SJ: Aberrant promoter methylation can be
useful as a marker of recurrent disease in patients with cervical
intraepithelial neoplasia grade III. Tumori. 93:572–579.
2007.PubMed/NCBI
|
|
69
|
Aran D, Sabato S and Hellman A: DNA
methylation of distal regulatory sites characterizes dysregulation
of cancer genes. Genome Biol. 14:R212013. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Chen CL, Liu SS, Ip SM, Wong LC, Ng TY and
Ngan HY: E-cadherin expression is silenced by DNA methylation in
cervical cancer cell lines and tumours. Eur J Cancer. 39:517–523.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Jeong DH, Youm MY, Kim YN, Lee KB, Sung
MS, Yoon HK and Kim KT: Promoter methylation of p16, DAPK, CDH1,
and TIMP-3 genes in cervical cancer: Correlation with
clinicopathologic characteristics. Int J Gynecol Cancer.
16:1234–1240. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Shivapurkar N, Sherman ME, Stastny V,
Echebiri C, Rader JS, Nayar R, Bonfiglio TA, Gazdar AF and Wang SS:
Evaluation of candidate methylation markers to detect cervical
neoplasia. Gynecol Oncol. 107:549–553. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Overmeer RM, Henken FE, Snijders PJ,
Claassen-Kramer D, Berkhof J, Helmerhorst TJ, Heideman DA, Wilting
SM, Murakami Y, Ito A, et al: Association between dense CADM1
promoter methylation and reduced protein expression in high-grade
CIN and cervical SCC. J Pathol. 215:388–397. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Yadav SS, Prasad SB, Das M, Kumari S,
Pandey LK, Singh S, Pradhan S and Narayan G: Epigenetic silencing
of CXCR4 promotes loss of cell adhesion in cervical cancer. Biomed
Res Int. 2014:5814032014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Narayan G, Xie D, Ishdorj G, Scotto L,
Mansukhani M, Pothuri B, Wright JD, Kaufmann AM, Schneider A,
Arias-Pulido H and Murty VV: Epigenetic inactivation of TRAIL decoy
receptors at 8p12-21.3 commonly deleted region confers sensitivity
to Apo2L/trail-cisplatin combination therapy in cervical cancer.
Genes Chromosomes Cancer. 55:177–189. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Snellenberg S, Cillessen SA, Van Criekinge
W, Bosch L, Meijer CJ, Snijders PJ and Steenbergen RD:
Methylation-mediated repression of PRDM14 contributes to apoptosis
evasion in HPV-positive cancers. Carcinogenesis. 35:2611–2618.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Michie AM, McCaig AM, Nakagawa R and
Vukovic M: Death-associated protein kinase (DAPK) and signal
transduction: Regulation in cancer. FEBS J. 277:74–80. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Kim JH, Choi YD, Lee JS, Lee JH, Nam JH
and Choi C: Assessment of DNA methylation for the detection of
cervical neoplasia in liquid-based cytology specimens. Gynecol
Oncol. 116:99–104. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Yang N, Nijhuis ER, Volders HH, Eijsink
JJ, Lendvai A, Zhang B, Hollema H, Schuuring E, Wisman GB and van
der Zee AG: Gene promoter methylation patterns throughout the
process of cervical carcinogenesis. Cell Oncol. 32:131–143.
2010.PubMed/NCBI
|
|
80
|
Jha AK, Nikbakht M, Parashar G,
Shrivastava A, Capalash N and Kaur J: Reversal of hypermethylation
and reactivation of the RARβ2 gene by natural compounds in cervical
cancer cell lines. Folia Biol (Praha). 56:195–200. 2010.PubMed/NCBI
|
|
81
|
Rabizadeh S, Xavier RJ, Ishiguro K,
Bernabeortiz J, Lopez-Ilasaca M, Khokhlatchev A, Mollahan P,
Pfeifer GP, Avruch J and Seed B: The scaffold protein CNK1
interacts with the tumor suppressor RASSF1A and augments
RASSF1A-induced cell death. J Biol Chem. 279:29247–29254. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Aoki K and Taketo MM: Adenomatous
polyposis coli (APC): A multi-functional tumor suppressor gene. J
Cell Sci. 120:3327–3335. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Dong SM, Kim HS, Rha SH and Sidransky D:
Promoter hypermethylation of multiple genes in carcinoma of the
uterine cervix. Clin Cancer Res. 7:1982–1986. 2001.PubMed/NCBI
|
|
84
|
Song Y and Zhang C: Hydralazine inhibits
human cervical cancer cell growth in vitro in association with APC
demethylation and re-expression. Cancer Chemother Pharmacol.
63:605–613. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Baer C, Claus R and Plass C: Genome-wide
epigenetic regulation of miRNAs in cancer. Cancer Res. 73:473–477.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Jiménez-Wences H, Peralta-Zaragoza O and
Fernández-Tilapa G: Human papilloma virus, DNA methylation and
microRNA expression in cervical cancer (Review). Oncol Rep.
31:2467–2476. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Iida M, Banno K, Yanokura M, Nakamura K,
Adachi M, Nogami Y, Umene K, Masuda K, Kisu I, Iwata T, et al:
Candidate biomarkers for cervical cancer treatment: Potential for
clinical practice (Review). Mol Clin Oncol. 2:647–655. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Dakubo GD: Methylated DNA as cancer
biomarkers in circulationCancer Biomarkers in Body Fluids. 1st.
Springer International Publishing; Cham: pp. 103–123. 2016,
View Article : Google Scholar
|
|
89
|
Huang RL, Su PH, Liao YP, Wu TI, Hsu YT,
Lin WY, Wang HC, Weng YC, Ou YC, Huang TH and Lai HC: Integrated
Epigenomics analysis reveals a DNA methylation panel for
endometrial cancer detection using cervical scrapings. Clin Cancer
Res. 23:263–272. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Kahn SL, Ronnett BM, Gravitt PE and
Gustafson KS: Quantitative methylation-specific PCR for the
detection of aberrant DNA methylation in liquid-based Pap tests.
Cancer. 114:57–64. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Wisman GB, Nijhuis ER, Hoque MO,
Reesink-Peters N, Koning AJ, Volders HH, Buikema HJ, Boezen HM,
Hollema H, Schuuring E, et al: Assessment of gene promoter
hypermethylation for detection of cervical neoplasia. Int J Cancer.
119:1908–1914. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Feng Q, Hawes SE, Stern JE, Dem A, Sow PS,
Dembele B, Toure P, Sova P, Laird PW and Kiviat NB: Promoter
hypermethylation of tumor suppressor genes in urine from patients
with cervical neoplasia. Cancer Epidemiol Biomarkers Prev.
16:1178–1184. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Huang BH, Laban M, Leung CH, Lee L, Lee
CK, Salto-Tellez M, Raju GC and Hooi SC: Inhibition of histone
deacetylase 2 increases apoptosis and p21Cip1/WAF1 expression,
independent of histone deacetylase. Cell Death Differ. 12:395–404.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Danam RP, Howell SR, Brent TP and Harris
LC: Epigenetic regulation of O6-methylguanine-DNA methyltransferase
gene expression by histone acetylation and methyl-CpG binding
proteins. Mol Cancer Ther. 4:61–69. 2005.PubMed/NCBI
|
|
95
|
Lee J, Yoon YS and Chung JH: Epigenetic
silencing of the WNT antagonist DICKKOPF-1 in cervical cancer cell
lines. Gynecol Oncol. 109:270–274. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Bodily JM, Mehta KP and Laimins LA: Human
papillomavirus E7 enhances hypoxia-inducible factor 1-mediated
transcription by inhibiting binding of histone deacetylases. Cancer
Res. 71:1187–1195. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Lu TY, Kao CF, Lin CT, Huang DY, Chiu CY,
Huang YS and Wu HC: DNA methylation and histone modification
regulate silencing of OPG during tumor progression. J Cell Biochem.
108:315–325. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Zhang Z, Joh K, Yatsuki H, Zhao W, Soejima
H, Higashimoto K, Noguchi M, Yokoyama M, Iwasaka T and Mukai T:
Retinoic acid receptor beta 2 is epigenetically silenced either by
DNA methylation or repressive histone modifications at the promoter
in cervical cancer cells. Cancer Lett. 247:318–327. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Seligson DB, Horvath S, Shi T, Yu H, Tze
S, Grunstein M and Kurdistani SK: Global histone modification
patterns predict risk of prostate cancer recurrence. Nature.
435:1262–1266. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Plesa A, Iancu IV, Botezatu A, Huica I,
Stoian M and Anton G: The involvement of epigenetic mechanisms in
HPV-induced cervical cancer. Human Papillomavirus – Research in a
Global Perspective. Rajkumar R: 9InTech. 2016. View Article : Google Scholar
|
|
101
|
Easwaran H, Tsai HC and Baylin SB: Cancer
Epigenetics: Tumor heterogeneity, plasticity of stem-like states,
and drug resistance. Mol Cell. 54:716–727. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
de la Cruz-Hernández E, Pérez-Cárdenas E,
Contreras-Paredes A, Cantú D, Mohar A, Lizano M and Dueñas-González
A: The effects of DNA methylation and histone deacetylase
inhibitors on human papillomavirus early gene expression in
cervical cancer, an in vitro and clinical study. Virol J. 4:182007.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Sui X, Chen R, Wang Z, Huang Z, Kong N,
Zhang M, Han W, Lou F, Yang J, Zhang Q, et al: Autophagy and
chemotherapy resistance: A promising therapeutic target for cancer
treatment. Cell Death Dis. 4:e8382013. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Hassan M, Watari H, AbuAlmaaty A, Ohba Y
and Sakuragi N: Apoptosis and molecular targeting therapy in
cancer. Biomed Res Int. 2014:1508452014. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Esteller M: Epigenetics in cancer. N Engl
J Med. 358:1148–1159. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Fang J, Zhang H and Jin S: Epigenetics and
cervical cancer: From pathogenesis to therapy. Tumor Biol.
35:5083–5093. 2014. View Article : Google Scholar
|
|
107
|
Coronel J, Cetina L, Pacheco I,
Trejo-Becerril C, González-Fierro A, de la Cruz-Hernandez E,
Perez-Cardenas E, Taja-Chayeb L, Arias-Bofill D, Candelaria M, et
al: A double-blind, placebo-controlled, randomized phase III trial
of chemotherapy plus epigenetic therapy with hydralazine valproate
for advanced cervical cancer. Preliminary results. Med Oncol. 28
Suppl 1:S540–S546. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Zambrano P, Segura-Pacheco B,
Perez-Cardenas E, Cetina L, Revilla-Vazquez A, Taja-Chayeb L,
Chavez-Blanco A, Angeles E, Cabrera G, Sandoval K, et al: A phase I
study of hydralazine to demethylate and reactivate the expression
of tumor suppressor genes. BMC Cancer. 5:442005. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
You JS, Kang JK, Lee EK, Lee JC, Lee SH,
Jeon YJ, Koh DH, Ahn SH, Seo DW, Lee HY, et al: Histone deacetylase
inhibitor apicidin downregulates DNA methyltransferase 1 expression
and induces repressive histone modifications via recruitment of
corepressor complex to promoter region in human cervix cancer
cells. Oncogene. 27:1376–1386. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Kelly WK, O'Connor OA, Krug LM, Chiao JH,
Heaney M, Curley T, MacGregore-Cortelli B, Tong W, Secrist JP,
Schwartz L, et al: Phase I study of an oral histone deacetylase
inhibitor, suberoylanilide hydroxamic acid, in patients with
advanced cancer. J Clin Oncol. 23:3923–3931. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Chen CC, Lee KD, Pai MY, Chu PY, Hsu CC,
Chiu CC, Chen LT, Chang JY, Hsiao SH and Leu YW: Changes in DNA
methylation are associated with the development of drug resistance
in cervical cancer cells. Cancer Cell Int. 15:982015. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Sato N, Maitra A, Fukushima N, van Heek
NT, Matsubayashi H, Iacobuzio-Donahue CA, Rosty C and Goggins M:
Frequent hypomethylation of multiple genes overexpressed in
pancreatic ductal adenocarcinoma. Cancer Res. 63:4158–1166.
2003.PubMed/NCBI
|
|
113
|
Virmani AK, Muller C, Rathi A,
Zoechbauer-Mueller S, Mathis M and Gazdar AF: Aberrant methylation
during cervical carcinogenesis. Clin Cancer Res. 7:584–589.
2001.PubMed/NCBI
|
|
114
|
Kim YT and Zhao M: Aberrant cell cycle
regulation in cervical carcinoma. Yonsei Med J. 46:597–613. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Ohtani N, Yamakoshi K, Takahashi A and
Hara E: The p16INK4a-RB pathway: Molecular link between cellular
senescence and tumor suppression. J Med Invest. 51:146–153. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Ki KD, Lee SK, Tong SY, Lee JM, Song DH
and Chi SG: Role of 5′-CpG island hypermethylation of the FHIT gene
in cervical carcinoma. J Gynecol Oncol. 19:117–122. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Özören N and El-Deiry WS: Cell surface
death receptor signaling in normal and cancer cells. Semin Cancer
Biol. 13:135–147. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Jung S, Yi L, Jeong D, Kim J, An S, Oh TJ,
Kim CH, Kim CJ, Yang Y, Kim KI, et al: The role of ADCYAP1,
adenylate cyclase activating polypeptide 1, as a methylation
biomarker for the early detection of cervical cancer. Oncol Rep.
25:245–252. 2011.PubMed/NCBI
|
|
119
|
Huang RL, Chang CC, Su PH, Chen YC, Liao
YP, Wang HC, Yo YT, Chao TK, Huang HC, Lin CY, et al: Methylomic
analysis identifies frequent DNA methylation of zinc finger protein
582 (ZNF582) in cervical neoplasms. PLoS One. 7:e410602012.
View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Ivanova T, Vinokurova S, Petrenko A,
Eshilev E, Solovyova N, Kisseljov F and Kisseljova N: Frequent
hypermethylation of 5′flanking region of TIMP-2 gene in cervical
cancer. Int J Cancer. 108:882–886. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Widschwendter A, Müller HM, Fiegl H,
Ivarsson L, Wiedemair A, Müller-Holzner E, Goebel G, Marth C and
Widschwendter M: DNA methylation in serum and tumors of cervical
cancer patients. Clin Cancer Res. 10:565–571. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Zhai Y, Bommer GT, Feng Y, Wiese AB,
Fearon ER and Cho KR: Loss of estrogen receptor 1 enhances cervical
cancer invasion. Am J Pathol. 177:884–895. 2010. View Article : Google Scholar : PubMed/NCBI
|