Introduction
Cervical cancer is the third most prevalent
gynecological malignancy and the second leading cause of
cancer-associated mortality among females worldwide, with an
estimated 530,000 mortalities per year (1). It is the result of a multistep process
involving the transformation of normal cervical epithelium to
pre-neoplastic cervical intraepithelial neoplasia, which is
subsequently transformed to cervical cancer (2). High-risk human papillomavirus (HPV)
infection performs an important role in cervical cancer
progression, whereas HPV infection alone is insufficient for tumor
development (3). The molecular
mechanisms underlying the development of cervical cancer have not
been fully elucidated. Therefore, it is necessary to further
explore the molecular mechanisms underlying cervical carcinogenesis
to facilitate the development of novel and effective therapeutic
targets for cervical cancer treatment.
MicroRNAs (miRNAs/miRs) represent a class of small
non-coding regulatory RNA molecules (19–25 nucleotides), which can
downregulate gene expression by binding to a complementary sequence
in the 3′untranslated region (3′UTR) region of target mRNAs
(4,5).
Functionally, miRNAs are involved in the regulation of numerous
critical biological processes, including cell proliferation,
differentiation, migration and invasion (6). Accumulating evidence has indicated that
dysregulation of miRNAs occurs in various tumor types, and that
they function as oncogenes or tumor suppressors according to the
nature of the target (7).
Additionally, many studies have demonstrated that the deregulation
of miRNAs is associated with cancer initiation and progression
(8). Clinically, miRNAs can be used
as potential biomarkers for the diagnosis and prognosis assessment
of patient with cancer, and possibly as therapeutic targets
(9).
Currently, more than several hundred, unique mature
human miRNAs are known, a number of which are involved in
tumorigenesis. Among them, miR-204 was reported as a tumor
suppressor and is frequently downregulated in gastric cancer,
breast cancer, renal cell carcinoma, non-small cell lung cancer
(NSCLC) and glioma (10–14). Overexpression of miR-204 in cancer
cells inhibits migration and proliferation, and promotes apoptosis
(15–18). However, to the best of our knowledge,
there are no studies regarding the association between miR-204 and
cervical cancer. Therefore, the present study was performed to
determine whether miR-204 serves a role in cervical
carcinogenesis.
In the present study, it was revealed that miR-204
is significantly decreased in primary tumor tissues, and that
reduced miR-204 was associated with lymph node status and poor
overall survival. In vitro analysis demonstrated that
overexpression of miR-204 inhibits cell migration and invasion in
cervical cancer cells. Furthermore, transcription factor 12 (TCF12)
was identified as a direct target of miR-204, and to be involved in
the miR-204 regulated cell motility alteration. Thus, results of
the present study, for the first time, provided novel clues
regarding the role of miR-204 as a metastasis-associated gene by
regulating TCF12 in cervical cancer.
Materials and methods
Clinical samples and cell lines
Fresh cervical cancer and matched adjacent
non-cancerous tissues were collected from 58 patients who underwent
surgery between April 2009 and January 2011 at the Department of
Obstetrics and Gynecology, Huzhou Central Hospital (Huzhou, China).
No patients received preoperative radiotherapy, chemotherapy or
other treatment history. The corresponding non-cancerous tissues
were obtained 4 cm beyond the boundary of cervical cancer tissues.
Clinicopathological variables, including age, tumor size,
differentiation and International Federation of Gynecology and
Obstetrics (FIGO) stage (19) were
collected prospectively. All the histological diagnoses for tumor
tissues and normal tissues were examined and confirmed by two
independent pathologists who were blinded to clinical outcomes of
the patients. The fresh tissue specimens were immediately frozen in
liquid nitrogen until use. The study protocol was approved by the
institutional review board of Huzhou Central Hospital, and the
study was conducted according to the principles of the Declaration
of Helsinki. Written informed consent was obtained from all
patients.
Four human cervical cancer cell lines, HeLa, C33A,
SiHa and CaSki, and an immortalized HPV-negative skin keratinocyte
cell line, HaCaT, were purchased from the American Type Culture
Collection (Manassas, VA, USA) and cultured in RPMI 1640
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Invitrogen; Thermo
Fisher Scientific, Inc.), 100 IU/ml of penicillin and 100 lg/ml of
streptomycin. All cells were incubated in a humidified incubator at
37°C and 5% CO2.
miRNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total miRNA was extracted from cell lines and
tissues using the miRNeasy Mini kit (Qiagen, Inc., Valencia, CA,
USA) according to the manufacturer's instructions. An RT-qPCR assay
was conducted to detect the expression levels of miR-204 in cell
lines and tissues. Briefly, 10 µg total RNA was subjected to RT.
The cDNA was used for the amplification of mature miR-204, and the
endogenous control U6, by PCR. The PCR conditions were as follows:
Initial denaturation at 95°C for 3 min, followed by 45 cycles of
95°C for 15 sec, 62°C for 1 min and 72°C for 1 min. RT-qPCR was
performed using SYBR® Green PCR Master Mix (Applied
Biosystems; Thermo Fisher Scientific, Inc.) on an ABI 7000 HT
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The primers used were as follows: miR-204 forward,
5′-GTCCCTGTGTCATCCT-3′ and reverse, 5′-CAGTGCAGGGTCCGAGGTAT-3′; U6
forward, 5′-CTCTCTGCGGCAGCACA-3′ and reverse,
5′-AACGCTGTACGAATGTGAGT-3′. The quantity of miR-204 was normalized
to U6, and calculated using the 2−ΔΔCq method (20). All RT-qPCR reactions were performed in
triplicate.
Plasmid construction, oligonucleotide
synthesis and transfection
The miR-204 mimic (sequence:
5′-UUCCCUUUGUCAUCCUAUGCCU-3′) and the mimic negative control
(mim-NC, sequence: 5′-UUUGUACUACACAAAAGUACUG-3′) were purchased
from Shanghai GenePharma Co., Ltd. (Shanghai, China). HeLa and C33A
cells were transfected with 400 µM mimic or 400 µM mim-NC using
Lipofectamine® 2000 Transfection Reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). A total of 24 h post-transfection,
cells were used for in vitro migration and invasion assays.
TCF12 cDNA without its 3′UTR was inserted into a pcDNA3.1(+) vector
(Invitrogen; Thermo Fisher Scientific, Inc.) to generate the
recombinant vector.
Migration assay
A wound-healing assay was conducted to detect cell
migration. Cells (5×105 cells/well) were seeded onto
6-well plates and grown to 80% confluency in RPMI 1640 medium
supplemented with 10% FBS (Invitrogen; Thermo Fisher Scientific,
Inc.). The cells were then transfected with miR-204 mimics, and the
miRNA mimic control (mim-NC) was used as the control group. After
24 h, a wound was created in the confluent cell monolayer using a
200-ml pipette tip. The cells were washed with RPMI 1640 medium
three times and then incubated at 37°C for 24 h. The mean distance
of the migrated cells was determined at 24 h.
Invasion assay
For the invasion assay, cells were serum-starved for
6 h in Dulbecco's modified Eagle's medium (DMEM; Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA) containing 0.1% FBS
(Invitrogen; Thermo Fisher Scientific, Inc.). Serum-starved cells
were trypsinized and resuspended in DMEM containing 0.1% FBS,
following which cells (4×104) were added to the upper
chambers of Transwell assay plate; each well was coated with 30
mg/cm2 Matrigel (Sigma-Aldrich; Merck KGaA). After 24 h
at 37°C, cells remaining on the upper membrane surface were removed
by careful wiping with a cotton swab, the migrated cells were fixed
in 95% ethanol at room temperature for 30 min and then stained with
0.2% crystal violet solution (Sigma-Aldrich; Merck KGaA) for 30 min
at room temperature. Invasive cells adhering to the undersurface of
the filter were then counted using an inverted microscope
(×100).
Target prediction
Three publically available databases [including
TargetScan, www.targetscan.org/vert_71 (21), PicTar, www.pictar.org (22) and
miRanda, www.microrna.org (23)] were used to predict the candidate
targets of miR-204.
Luciferase reporter assay
To construct the pMIR-TCF12-3′UTR plasmid that
contained the potential binding sites of TCF12 3′UTR downstream of
the firefly luciferase gene, a fragment was amplified and inserted
into the pMIR-REPORT Luciferase Vector (Ambion; Thermo Fisher
Scientific, Inc.). Amplification conditions were as follows: 94°C
for 5 min, 40 cycles of 60°C for 20 sec and 72°C for 20 sec, and
Taq DNA Polymerase (Thermo Fisher Scientific, Inc.) was used in
this procedure. The primers were used as follows: forward,
5′-AAGCAACTGGTCAACACTTCC-3′, and reverse,
5′-CGGTGAGTGGAGAGATCGAG-3′. HeLa and C33A cells were used to
measure luciferase activity. Cells were co-transfected with 100 ng
luciferase plasmid and 50 ng Renilla plasmid (Ambion; Thermo
Fisher Scientific, Inc.), along with 600 ng miR-204 mimic or NC-mim
using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc.). Mutations in the miR-204-binding site of TCF12 3′-UTR were
introduced by the QuikChange Site-Directed Mutagenesis kit
(Stratagene; Agilent Technologies, Inc., Santa Clara, CA, USA)
according to manufacturer's protocol. Luciferase activity was
measured 48 h post-transfection using the Dual-Luciferase Reporter
Assay System (Promega Corporation, Madison, WI, USA). Firefly
luciferase activity was then normalized to the corresponding
Renilla luciferase activity. Data are presented as the mean value
obtain from triplicate experiments.
Western blot analysis
Proteins were extracted from the cells using the
protein extraction reagent radioimmunoprecipitation assay (RIPA)
buffer (Beyotime Institute of Biotechnology, Haimen, China)
supplemented with protease inhibitors (cat no. 8553; Cell Signaling
Technology, Inc., Danvers, MA, USA), and quantified by a
bicinchoninic acid protein assay (Beyotime Institute of
Biotechnology). Subsequently, 50 µg protein extractions were
separated via 12% SDS-PAGE, transferred to nitrocellulose membranes
(Whatman® Ptotran®; GE Healthcare, Chicago,
IL, USA) and incubated with specific primary antibodies, as
follows: TCF12 Rabbit mAb (cat no. 11825; dilution, 1:1,000; Cell
Signaling Technology, Inc.) and GAPDH antibody (cat no. sc-47724;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) overnight at 4°C.
The membranes were then washed and probed with horseradish
peroxidase (HRP)-conjugated secondary antibodies for GAPDH (mouse
IgG binding protein-HRP; cat no. sc-516102; 1:5,000; Santa Cruz
Biotechnology, Inc.) and for TCF12 (Mouse Anti-rabbit IgG; cat no.
5127; 1:5,000; Cell Signaling Technology, Inc.) at room temperature
for 30 min. The signals were detected using the Chemiluminescent
ECL Detection system (Merck KGaA), and analysed using ImageJ 2×
version 2.1.4.7 software (Rawak Software, Inc., Dresden,
Germany).
Statistical analysis
Statistical analysis was performed using SPSS 18.0
software (SPSS, Inc., Chicago, IL, USA) and Prism 5.0 (GraphPad
Software, Inc., La Jolla, CA, USA). The expression of miR-204 in
tissues were presented as the mean ± the standard deviation. The
difference in expression levels between paired samples was analyzed
by a paired t-test. Comparisons between groups were performed using
the Mann-Whitney U test for continuous variables. The Kaplan-Meier
method was used for survival analysis, and differences in survival
curves were compared using the log-rank test. All tests were
two-sided, and P<0.05 was considered to indicate a statistically
significant difference.
Results
miR-204 is downregulated in cervical
cancer tissues
To determine the expression level of miR-204 in
cervical cancer cells, RT-qPCR was performed in four cancer cells
(HeLa, C33A, SiHa and CaSki), and HaCaT cells as the normal
control. The cervical cancer cells exhibited significantly lower
expression of miR-204 compared with HaCaT (data not shown). The
level of miR-204 in tissue specimens from patients with cervical
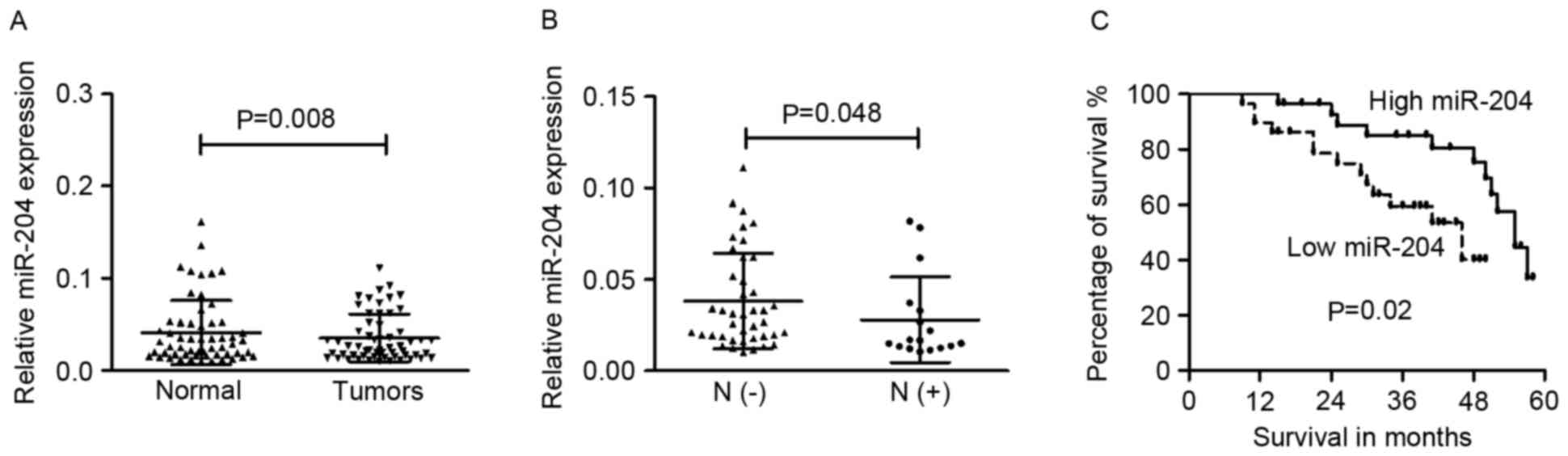
cancer was then examined. The expression of miR-204 in the primary
tumor tissues (0.035±0.025) was significantly lower than that in
the adjacent non-cancerous tissues (0.041±0.034; Fig. 1A; P=0.008).
Low expression of miR-204 is
associated with poor prognosis in patients with cervical
cancer
The associations between miR-204 expression levels
and different clinicopathological characteristics were tested. It
was revealed that miR-204 expression was significantly decreased in
the tumor tissues of patients with lymph node metastasis
(0.028±0.023) compared with those without lymph node metastasis
(0.038±0.026; P=0.048; Fig. 1B). The
downregulated miR-204 had no association with other
clinicopathological characteristics of cervical cancer cases. It
was speculated that downregulated miR-204 may be involved in the
regulation of metastasis in the development of cervical cancer.
The median level of miR-204 was used as a cut-off
value to divide 58 cases into two groups (low- or high- group). The
association between miR-204 and the overall survival of cervical
cancer patients was determined via Kaplan-Meier analysis and the
log-rank test. As shown in Fig. 1C,
patients with cervical cancer and low miR-204 expression had poorer
overall survival rates compared with those with high miR-204
expression (P=0.02).
miR-204 inhibits cell migration and
invasion in vitro
Considering the association between miR-204 levels
and lymph node metastasis, the present study then focused on the
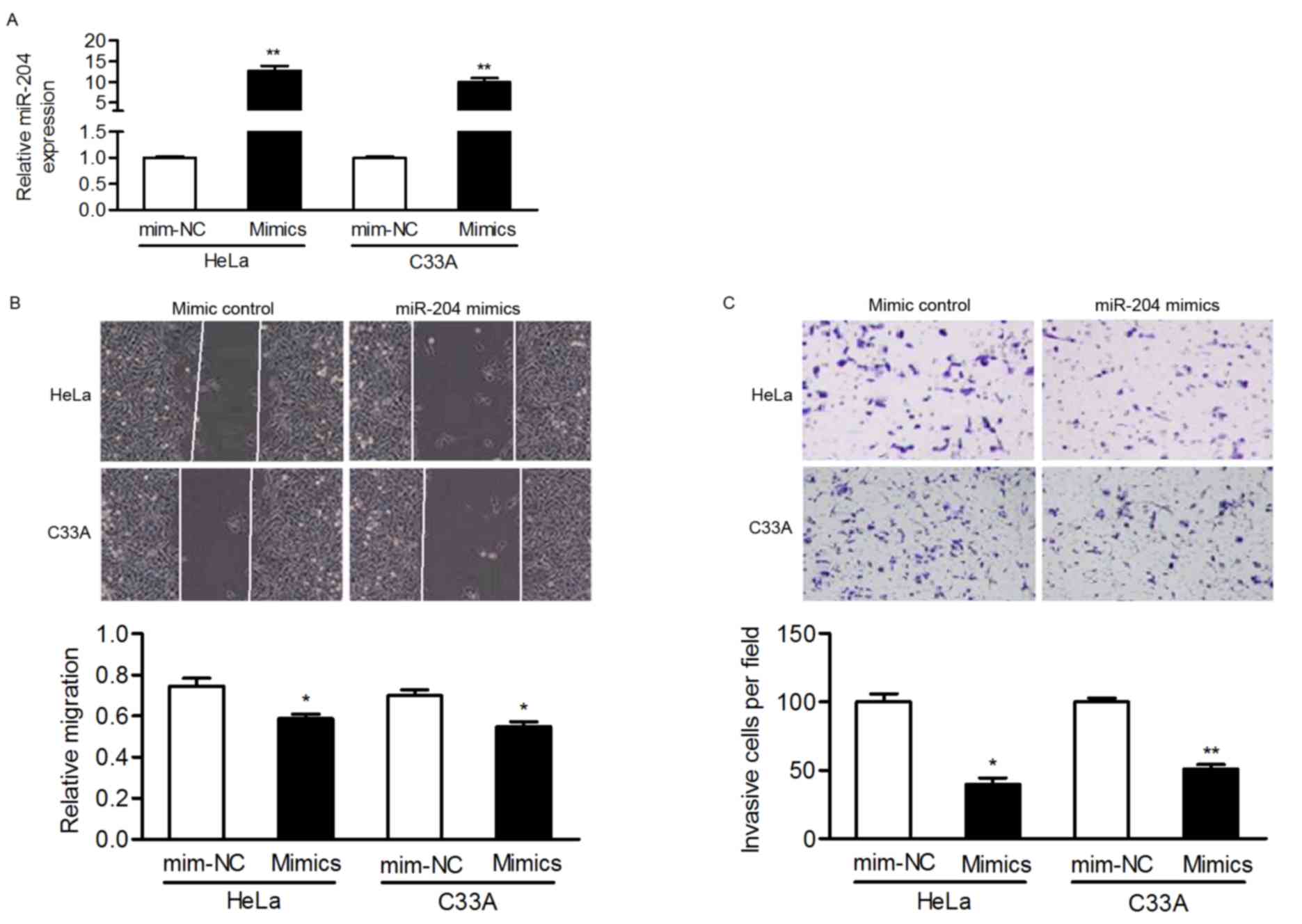
effect of miR-204 on cell motility. HeLa and C33A cells, selected
as they express relatively low levels of miR-204, were transfected
with miR-204 mimics and it was confirmed that the expression of
miR-204 was upregulated in transfected cells (Fig. 2A).
Wound-healing and Matrigel Transwell assays were
performed to verify whether miR-204 inhibited the migration and
invasion of cervical cancer cells. In HeLa and C33A cells,
overexpressing miR-204 inhibited the migratory ability of cells in
comparison to cells transfected with the mim-NC (Fig. 2B). Furthermore, the invasive capacity
of cervical cancer cells was markedly suppressed in
miR-204-transfected cells (Fig. 2C).
These results indicated that miR-204 may be a metastasis-associated
gene and involved in cervical cancer pathogenesis.
miR-204 directly inhibits TCF12
expression by targeting its 3′UTR
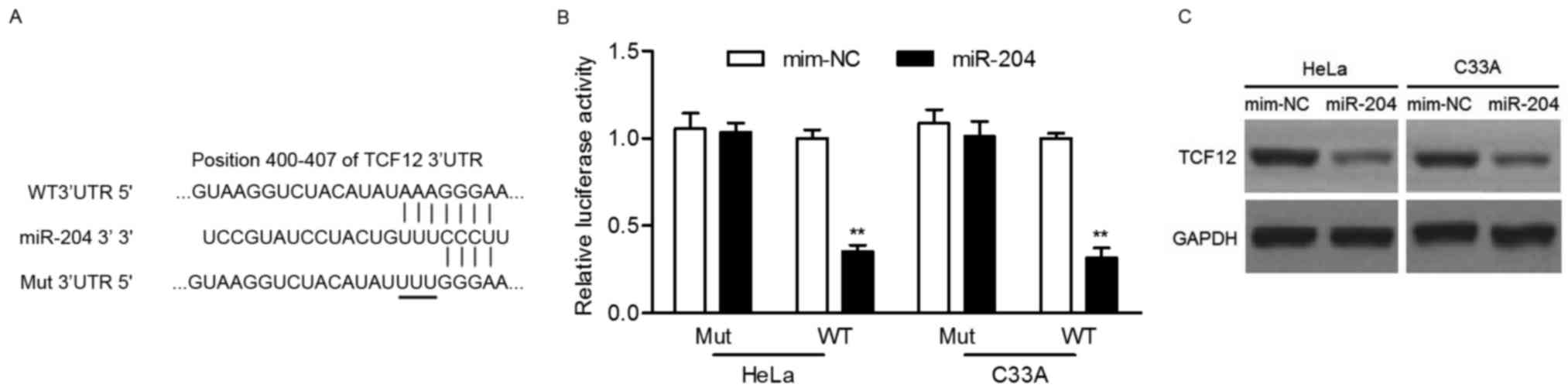
To explore the molecular mechanism by which miR-204
inhibits cervical cancer cells, potential targets of miR-204 were
predicted using the PicTar, TargetScan and miRanda tools. Among the
predicted candidate targets, the present study focused on TCF12 and
considered the involvement of TCF12 in tumor metastasis (24,25). As
presented in Fig. 3A, the 3′-UTR of
TCF12 possesses a potential binding site of miR-204. To determine
if miR-204 could directly target the 3′UTR of TCF12, luciferase
activity assay was performed. Luciferase reporter assays revealed
that the upregulation of miR-204 significantly decreased the
relative luciferase activity of the 3′UTR of TCF12 in HeLa or C33A
cells, but had no effect on the mutant 3′UTR of TCF12 (Fig. 3B). In addition, western blot analysis
revealed that overexpression of miR-204 noticeably suppressed the
protein expression of TCF12 (Fig.
3C). Taken together, these results demonstrated that miR-204
suppresses TCF12 expression by directly targeting its 3′UTR.
miR-204 inhibits the malignant
phenotypes of cervical cancer cells by regulating TCF12
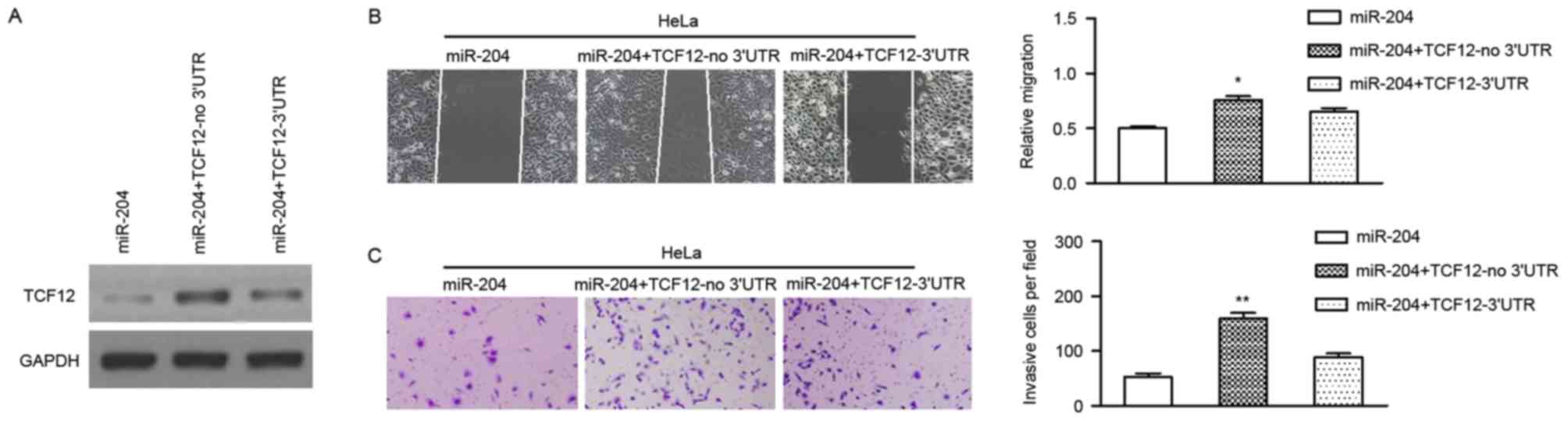
As miR-204 regulates the expression of TCF12
post-transcriptionally, and miR-204 inhibits the migration and
invasion of cervical cancer cells, it was postulated that these
effects were attributed to TCF12. To verify this hypothesis, a
vector was constructed using cDNA that lacked the 3′UTR of TCF12,
and the vector was transiently co-transfected with miR-204 mimics
into cervical cancer cells. It was revealed that the transfection
of TCF12 (no 3′UTR) could effectively reverse the expression of
TCF12 in HeLa and C33A cells (Fig.
4A). Subsequently, it was revealed that the restoration of
TCF12 could significantly attenuate the inhibitory role of miR-204
in the migration and invasion of cervical cancer cells (Fig. 4B and C).
Discussion
Exploring the underlying molecular mechanisms of
carcinogenesis is essential for early diagnosis and effective
therapy. Furthermore, one miRNA is able to regulate the expression
of multiple genes, since it can bind to its mRNA targets either as
an imperfect or a perfect complement. It is, therefore, not
surprising that miRNAs are involved in the regulation of all major
tumor-associated biological processes, including differentiation,
proliferation, migration and apoptosis. The present study
demonstrated that miR-204 expression was downregulated in cervical
cancer tissues. The results of Kaplan-Meier analysis revealed that
the downregulation of miR-204 was associated with a poor prognosis.
It was demonstrated that the overexpression of miR-204 suppressed
migration and invasion in cervical cancer cells. Furthermore, TCF12
was also identified as a novel and direct target of miR-204.
miR-204, located on chromosome 9q21.12, has
attracted attention due to its involvement in numerous tumor types.
Previous studies have documented that miR-204 expression was
frequently downregulated in gastric cancer, breast cancer, renal
cell carcinoma, glioma, prostate cancer and NSCLC (10–14). In
accordance with these studies, the present data demonstrated the
decreased expression of miR-204 in primary cervical cancer tissues
compared with adjacent normal tissues, indicating the potential
tumor suppressor role of miR-204 in cervical cancer
progression.
In addition, the prognostic value of the expression
of miR-204 in patients with cervical cancer was analyzed. It was
revealed that low expression of miR-204 was associated with lymph
node metastasis and poor overall survival. Consistent with the
present data, Bao et al (26)
reported that lower miR-204-5p expression was associated with
advanced FIGO stages, and possibly a lower rate of survival in
patients with endometrial carcinoma. A low level of miR-204 was
significantly associated with a more aggressive and poor prognostic
phenotype of nasopharyngeal carcinoma (27). Low miR-204 expression was also
significantly associated with higher tumor-node-metastasis stage
and poorer overall survival in breast cancer cases (11). These results indicated that miR-204
expression may be a promising prognostic marker for patients with
cancer.
Furthermore, miR-204 expression was associated with
chemotherapeutic resistance of patients with breast cancer
(11). Enhanced miR-204 expression
significantly increased sensitivity to cisplatin and etoposide
in vitro (28). Therefore,
additional studies regarding the association between expression of
miR-204 and anti-carcinogen, such as cisplatin and etoposide, may
provide new insights into chemotherapy of cervical cancer.
miR-204 acts as a tumor suppressor in various types
of tumors by regulating its target genes and is involved in several
tumor-associated processes. Wu et al (17) indicated that miR-204 could regulate
the proliferation and invasion of retinoblastoma cells by directly
targeting the cyclin D2 and matrix metalloproteinase-9 genes.
Downregulation of miR-204 and the consequent overexpression of SRY
sex-determining region-Y box 4 promoted epithelial-mesenchymal
transition (EMT) and enhanced the invasion and proliferation of
gastric cancer cells in vitro (29). Loss of miR-204 promotes to cancer cell
migration and invasion, while restoration of miR-204 inhibited EMT
by targeting the EMT inducer, including slug and sirtuin 1
(30,31). Shi et al (13) indicated that miR-204 inhibited NSCLC
tumor migration and invasion by directly targeting and
downregulating NUAK family kinase 1 expression. By contrast,
overexpression of miR-204 suppressed papillary thyroid carcinoma
cell proliferation, induced cell cycle arrest and apoptosis and
inhibited insulin-like growth factor binding protein 5 mRNA
(32). In the present study, it was
observed that the overexpression of miR-204 suppressed cell
migration and invasion ability. The present study expanded the
knowledge of miR-204 in cervical cancer and demonstrated that
miR-204 acts as a metastasis-associated gene in cervical cancer.
More importantly, it was revealed that miR-204 directly regulates
TCF12 expression by targeting its 3′UTR. Previous studies reported
that TCF12 functions as a transcriptional repressor of E-cadherin
and that its overexpression was significantly associated with the
occurrence of colorectal cancer metastasis (24,25). Thus,
it was proposed that miR-204/TCF12 axis may serve important roles
in the migration and invasion of cervical cancer.
In conclusion, miR-204 was significantly
downregulated in cervical cancer tissues and ectopic miR-204
inhibited cell motility. TCF12 was a direct target of miR-204 and
upregulated TCF12 impaired the inhibitory effect of miR-204 on cell
migration, indicating that miR-204 functions as a
metastasis-associated gene, possibly by targeting TCF12 in cervical
cancer. Thus, these results demonstrated that miR-204 performs
important roles in the migration and invasion of cervical cancer by
targeting TCF12.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Waggoner SE: Cervical cancer. Lancet.
361:2217–2225. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hildesheim A and Wang SS: Host and viral
genetics and risk of cervical cancer: A review. Virus Res.
89:229–240. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Engels BM and Hutvagner G: Principles and
effects of microRNA-mediated posttranscriptional gene regulation.
Oncogene. 25:6163–6169. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Calin GA and Croce CM: MicroRNA-cancer
connection: The beginning of a new tale. Cancer Res. 66:7390–7394.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McManus MT: MicroRNAs and cancer. Semin
Cancer Biol. 13:253–258. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Davidson B, Tropé CG and Reich R: The
clinical and diagnostic role of microRNAs in ovarian carcinoma.
Gynecol Oncol. 133:640–646. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Seven M, Karatas OF, Duz MB and Ozen M:
The role of miRNAs in cancer: From pathogenesis to therapeutic
implications. Future Oncol. 10:1027–1048. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang B, Yin Y, Hu Y, Zhang J, Bian Z,
Song M, Hua D and Huang Z: MicroRNA-204-5p inhibits gastric cancer
cell proliferation by downregulating USP47 and RAB22A. Med Oncol.
32:3312015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li W, Jin X, Zhang Q, Zhang G, Deng X and
Ma L: Decreased expression of miR-204 is associated with poor
prognosis in patients with breast cancer. Int J Clin Exp Pathol.
7:3287–3292. 2014.PubMed/NCBI
|
|
12
|
Munari E, Marchionni L, Chitre A, Hayashi
M, Martignoni G, Brunelli M, Gobbo S, Argani P, Allaf M, Hoque MO
and Netto GJ: Clear cell papillary renal cell carcinoma: micro-RNA
expression profiling and comparison with clear cell renal cell
carcinoma and papillary renal cell carcinoma. Hum Pathol.
45:1130–1138. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shi L, Zhang B, Sun X, Lu S, Liu Z, Liu Y,
Li H, Wang L, Wang X and Zhao C: MiR-204 inhibits human NSCLC
metastasis through suppression of NUAK1. Br J Cancer.
111:2316–2327. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ying Z, Li Y, Wu J, Zhu X, Yang Y, Tian H,
Li W, Hu B, Cheng SY and Li M: Loss of miR-204 expression enhances
glioma migration and stem cell-like phenotype. Cancer Res.
73:990–999. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Imam JS, Plyler JR, Bansal H, Prajapati S,
Bansal S, Rebeles J, Chen HI, Chang YF, Panneerdoss S, Zoghi B, et
al: Genomic loss of tumor suppressor miRNA-204 promotes cancer cell
migration and invasion by activating AKT/mTOR/Rac1 signaling and
actin reorganization. PLoS One. 7:e523972012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yin Y, Zhang B, Wang W, Fei B, Quan C,
Zhang J, Song M, Bian Z, Wang Q, Ni S, et al: miR-204-5p inhibits
proliferation and invasion and enhances chemotherapeutic
sensitivity of colorectal cancer cells by downregulating RAB22A.
Clin Cancer Res. 20:6187–6199. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu X, Zeng Y, Wu S, Zhong J, Wang Y and Xu
J: MiR-204, down-regulated in retinoblastoma, regulates
proliferation and invasion of human retinoblastoma cells by
targeting CyclinD2 and MMP-9. FEBS Lett. 589:645–650. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li G, Luna C, Qiu J, Epstein DL and
Gonzalez P: Role of miR-204 in the regulation of apoptosis,
endoplasmic reticulum stress response, and inflammation in human
trabecular meshwork cells. Invest Ophthalmol Vis Sci. 52:2999–3007.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pecorelli S: Revised FIGO staging for
carcinoma of the vulva, cervix, and endometrium. Int J Gynaecol
Obstet. 105:103–104. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:2015. View Article : Google Scholar
|
|
22
|
Krek A, Grün D, Poy MN, Wolf R, Rosenberg
L, Epstein EJ, MacMenamin P, da Piedade I, Gunsalus KC, Stoffel M
and Rajewsky N: Combinatorial microRNA target predictions. Nat
Genet. 37:495–500. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
John B, Enright AJ, Aravin A, Tuschl T,
Sander C and Marks DS: Human MicroRNA targets. PLoS Biol.
2:e3632004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee CC, Chen WS, Chen CC, Chen LL, Lin YS,
Fan CS and Huang TS: TCF12 protein functions as transcriptional
repressor of E-cadherin, and its overexpression is correlated with
metastasis of colorectal cancer. J Biol Chem. 287:2798–2809. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen WS, Chen CC, Chen LL, Lee CC and
Huang TS: Secreted heat shock protein 90α (HSP90α) induces nuclear
factor-κB-mediated TCF12 protein expression to down-regulate
E-cadherin and to enhance colorectal cancer cell migration and
invasion. J Biol Chem. 288:9001–9010. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bao W, Wang HH, Tian FJ, He XY, Qiu MT,
Wang JY, Zhang HJ, Wang LH and Wan XP: A TrkB-STAT3-miR-204-5p
regulatory circuitry controls proliferation and invasion of
endometrial carcinoma cells. Mol Cancer. 12:1552013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ma L, Deng X, Wu M, Zhang G and Huang J:
Down-regulation of miRNA-204 by LMP-1 enhances CDC42 activity and
facilitates invasion of EBV-associated nasopharyngeal carcinoma
cells. FEBS Lett. 588:1562–1570. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ryan J, Tivnan A, Fay J, Bryan K, Meehan
M, Creevey L, Lynch J, Bray IM, O'Meara A, Tracey L, et al:
MicroRNA-204 increases sensitivity of neuroblastoma cells to
cisplatin and is associated with a favourable clinical outcome. Br
J Cancer. 107:967–976. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou X, Li L, Su J and Zhang G: Decreased
miR-204 in H. pylori-associated gastric cancer promotes cancer cell
proliferation and invasion by targeting SOX4. PLoS One.
9:e1014572014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Qiu YH, Wei YP, Shen NJ, Wang ZC, Kan T,
Yu WL, Yi B and Zhang YJ: miR-204 inhibits epithelial to
mesenchymal transition by targeting slug in intrahepatic
cholangiocarcinoma cells. Cell Physiol Biochem. 32:1331–1341. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang L, Wang X and Chen P: MiR-204 down
regulates SIRT1 and reverts SIRT1-induced epithelial mesenchymal
transition, anoikis resistance and invasion in gastric cancer
cells. BMC Cancer. 13:2902013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu L, Wang J, Li X, Ma J, Shi C, Zhu H,
Xi Q, Zhang J, Zhao X and Gu M: miR-204-5p suppresses cell
proliferation by inhibiting IGFBP5 in papillary thyroid carcinoma.
Biochem Biophys Res Commun. 457:621–626. 2015. View Article : Google Scholar : PubMed/NCBI
|