Introduction
Cancer is a malignant disease associated with
abnormal proliferation, aberrant differentiation and metastasis
(1). A high proportion of patients
with cancer present with varying degrees of metastasis at the point
of diagnosis. Although efforts have been made to identify
strategies to treat various types of cancer, numerous types
continue to be significant contributors to mortality and high
healthcare expenditures in populations (2,3).
Therefore, identifying early diagnostic or prognostic biomarkers,
and exploring effective treatment strategies to improve survival
times in different types of cancer, is urgent.
Tumorigenesis is associated with the functional
inhibition or loss of tumor suppressor genes and the overexpression
of oncogenes (4–6). With the continuous improvement of
high-throughput genome-wide and RNA-sequencing technologies, more
non-coding (nc)RNAs associated with human cancer progression have
been discovered (7,8). A threshold of 200 nucleotides divides
regulatory ncRNAs into short ncRNAs (sncRNAs) and long ncRNAs
(lncRNAs) (9,10). sncRNAs include microRNAs (miRNAs),
endogenous short interfering RNAs (endo-siRNAs) and
Piwi-interacting RNAs (piRNAs). lncRNAs are composed of >200
nucleotides, and include multiple well-studied ncRNAs, such as
metastasis-associated lung adenocarcinoma transcript 1 (non-protein
coding), hepatocellular carcinoma upregulated lncRNA and HOX
transcript antisense RNA. Circular (circ)RNAs, a subset of ncRNAs
that are generally composed of >200 nucleotides (with a minority
<200 nucleotides), have also been grouped into lncRNAs (11).
circRNAs have become another focus in the field of
ncRNA research (12). Previous
studies have indicated that certain circRNAs are endogenous,
stable, abundant and conserved in eukaryotic cells (13–16).
Differing from the linear RNAs, which feature 5′ caps and 3′ tails
as their termini, all known circRNAs possess covalently closed loop
structures, may occur in any genomic region, including gene-bearing
regions and intergenic regions, and range in length from a few
hundred to thousands of nucleotides (11,15,17).
Previously, certain circRNAs have been identified as potential
biomarkers for aging in Drosophila (18), which suggested the potential of
circRNAs as biomarkers for the diagnosis and treatment of cancer.
With regard to the function of circRNAs, previous studies have
indicated that circRNAs may serve as miRNA sponges and RNA-binding
protein (RBP) sponges and could regulate miRNA-mediated signaling
or RBP-associated transcription, and influence the function of
certain types of cell (11,13,14,16,19).
The present study reviews circRNAs in cancer,
summarizes the vital functions of circRNAs in different types of
cancer, highlights the competing endogenous (ce)RNA function of
circRNAs in regulating downstream gene expression, and provides
evidence for the application of circRNAs in the future diagnosis
and treatment of numerous types of cancer.
Landscape of circRNAs
In recent years, circRNAs have emerged as a novel
class of functional molecules among lncRNAs (8,10,15,20).
Differing from the linear RNAs with the typical termini consisting
of 5′ caps and 3′ tails, all known circRNAs display covalently
closed loop structures, without any 5′ to 3′ polarity or
polyadenylation at the 3′ ends (21,22),
leading to mass escape of circRNAs from general transcriptomic
polyadenylated RNA profiling (23,24).
However, RNA sequencing technology and deep bioinformatics analyses
have previously revealed numerous novel circRNAs in different
diseases and cell lines (11,12,15,18,25–27).
Origins of circRNAs
circRNAs may be found in serum, saliva and the
exosomes of mammalian cells. Using a genome-wide approach, circRNAs
were first detected in excised exons or introns (28) and were generally classified into three
subtypes: Exonic circRNAs (ecircRNAs) (29), circular intronic RNAs (ciRNAs)
(30), and exon-intron circRNAs
(EIciRNAs) (31). Cytoplasmic
ecircRNAs account for >90% of total circRNAs (11,13,21),
whereas nuclear circRNAs are mainly ciRNAs and EIciRNAs (30,31), and
exosomal circRNAs (exo-circRNAs), another type of circRNA, are
distributed in the exosomes of human serum and in multiple human
cancer cell lines (32).
To date, circRNAs have mainly been reported to form
via back-splicing (22), which is
characterized by the covalent binding of a 5′ splicing acceptor to
a 3′ splicing donor (15). Jeck et
al (13) proposed two models to
assess the production of circRNAs; model one is referred to as
‘direct back-splicing’, while model two is termed ‘exon skipping’
or ‘lariat intermediate’. Generally, alternative circularization is
generated from different numbers of exons (13,22,25,33),
with or without an internal intron (14,22,31).
Nevertheless, it remains unclear whether alternative
circularization occurs co-transcriptionally or
post-transcriptionally, and what factors affect
circularization.
Characteristics of circRNAs
circRNAs exhibit specific characteristics distinct
from linear RNAs. Firstly, circRNAs have particular covalently
closed loop structures, rather than the typical 5′ caps and 3′
tails at the termini of linear RNAs (21,22), which
render circRNAs more stable compared with their linear counterparts
and more resistant to degradation by various endogenous RNA
exonucleases (34,35). Additionally, certain circRNAs are
widespread and more abundant compared with their linear
counterparts (13,36). A previous study revealed that certain
circRNAs were expressed in a cell type-specific manner; for
example, hsa_circRNA_21 was only detected in cluster of
differentiation (CD)19+ leukocytes and not in
CD34+ leukocytes or neutrophils (11). The majority of circRNAs are
evolutionarily conserved among different species (13,14,37), with
the exception of intergenic or intronic circRNAs (30).
Association between circRNAs and multiple
types of cancer
Since the function of one circRNA molecule was first
proposed (11), the potential of
circRNAs has invoked widespread interest (38). Numerous circRNAs are differentially
expressed between cancerous and normal tissues, including in
colorectal cancer (CRC), gastric cancer (GC), hepatocellular
carcinoma (HCC), breast cancer, and bladder urothelial
carcinoma.
One example is homeodomain-interacting protein
kinase (HIPK)3 circRNA (circ-HIPK3), which is derived from exon 2
of the HIPK3 gene, and demonstrates significant differential
expression between cancerous and normal tissues (39).
Yang et al (40) revealed that the expression of forkhead
box O3 (FOXO3), one of the members of the forkhead family of
transcription factors, may be regulated by FOXO3 circRNA
(circ-FOXO3) and FOXO3 pseudogene (FOXO3P). The aberrant expression
of FOXO3 mRNA, circ-FOXO3 and FOXO3P may synergistically affect
tumor growth and survival (40).
Additionally, Du et al (41,42)
demonstrated that circ-FOXO3 may retard cell cycle progression and
induce cell apoptosis.
Li et al (32)
reported that abundant exo-circRNAs were markedly enriched in the
exosomes derived from multiple cancer cell lines, including lung,
breast, colon, cervical and stomach cancer, and the detected
exo-circRNAs derived from human serum further suggested their
potential as novel diagnostic or prognostic markers for multiple
types of cancer. Although the function of circRNAs in tumorigenesis
is limited, studies explored in the present review have indicated
that certain circRNAs may be associated with the development of
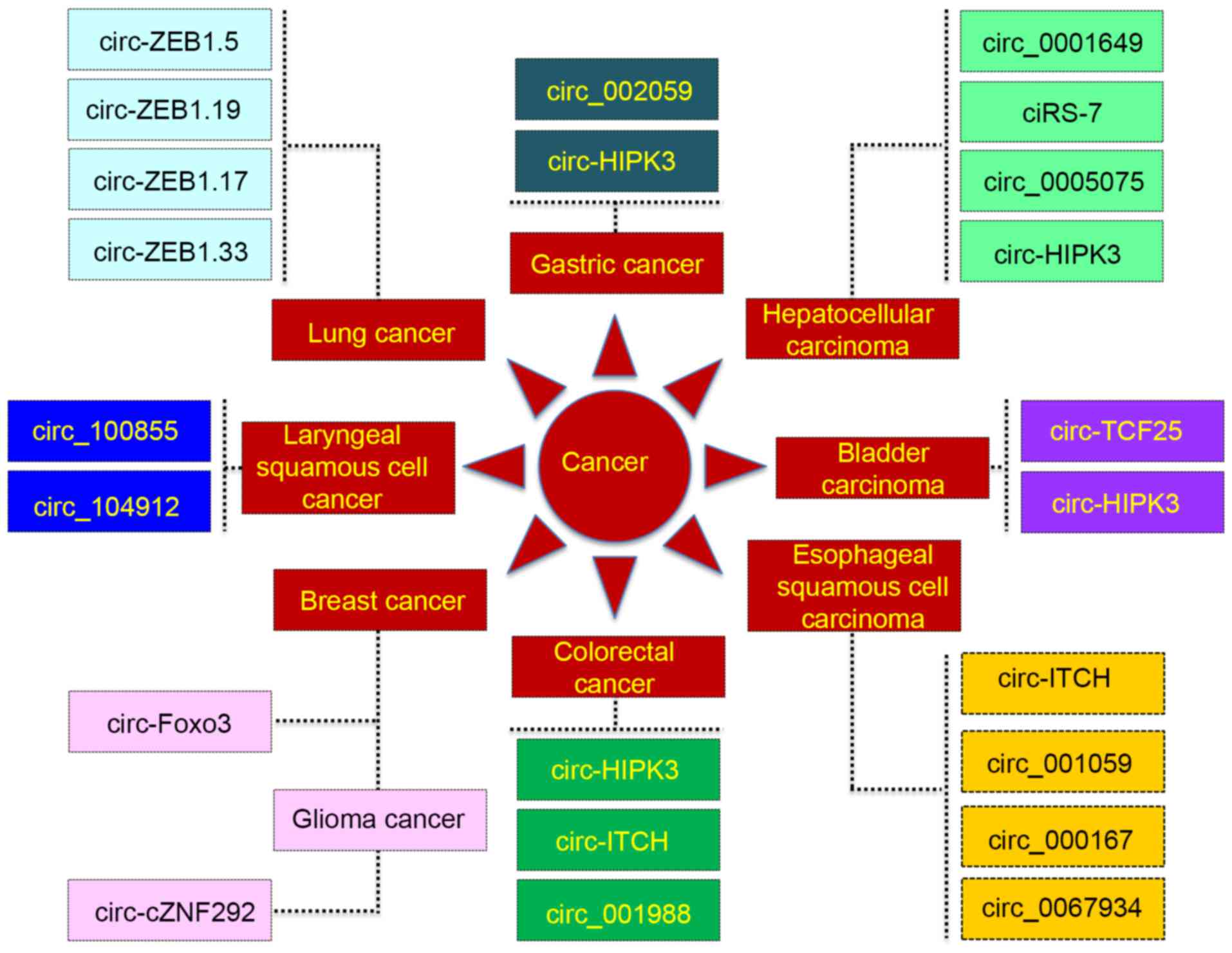
various types of cancer (Fig. 1).
circRNAs and CRC
In a previous study on CRC, 39 differentially
expressed circRNAs were identified between CRC and matched normal
colonic mucosa tissues (43). In
addition, the ratio of circRNAs to linear RNA counterparts was
decreased in CRC tissues compared with that in normal colon
tissues, and even further decreased in CRC cell lines compared with
that in normal colon tissues. Furthermore, there was a negative
correlation between this ratio and the proliferation index, which
provided the basis to explore their functions and mechanisms in
CRC. In addition, the authors hypothesized that their findings were
due to the dysfunction of back-splicing or dysregulation of
oncogenic miRNAs, resulting in the degradation and downregulation
of circRNAs in CRC (43).
Similarly, itchy E3 ubiquitin protein ligase (ITCH)
circRNA (circ-ITCH) expression was revealed to be significantly
downregulated in CRC tissues compared with that in peritumoral
tissues, and circ-ITCH could upregulate the expression of its
linear counterpart, ITCH, and thereafter affect the Wnt/β-catenin
signaling pathway in CRC (44).
Additionally, decreased expression of hsa_circ_001988 was
identified in CRC, and the expression of hsa_circ_001988 was
significantly correlated with differentiation and perineural
invasion of CRC cells (45). Dou
et al (46) also revealed that
numerous circRNAs are downregulated in KRAS proto-oncogene mutant
colon cancer cells and may be transferred to exosomes, and that
circRNAs were more abundant in exosomes compared with cells.
circRNAs and HCC
Qin et al (47)
revealed that hsa_circ_0001649 expression was significantly
decreased in HCC and may represent a novel biomarker for HCC. Xu
et al (48) demonstrated that,
although the expression of CDR1 antisense RNA (CDR1-AS; also known
as ciRS-7) was comparable between HCC and matched non-tumor
tissues, the increased ciRS-7 expression in HCC tissues was
significantly correlated with hepatic microvascular invasion and
therefore associated with the deterioration of HCC. Furthermore,
circRNA profiling revealed that hsa_circ_0005075, a circRNA
biomarker, exhibited a significant difference in expression between
HCC and normal tissues, and a pathological correlation analysis
indicated that hsa_circ_0005075 was associated with the development
of HCC (49).
circRNAs and esophageal squamous cell
carcinoma (ESCC)
Li et al (50)
demonstrated that the expression of circ-ITCH was downregulated in
ESCC. Su et al (51) also
performed expression profiling of circRNAs in radioresistant ESCC
cells, and revealed that circRNA_001059 and circRNA_000167 were
associated with the development of radiation resistance. In
addition, Xia et al (52)
demonstrated that, hsa_circ_0067934 expression was significantly
upregulated in ESCC and could promote the proliferation of ESCC
cells.
circRNAs and lung cancer
Expression patterns in CircNet revealed that
expression of the circRNAs circ-zinc finger E-box binding homeobox
(ZEB)1.5, circ-ZEB1.19, circ-ZEB1.17 and circ-ZEB1.33 were
downregulated in lung cancer specimens compared with that in normal
lung tissues (53). Another study
demonstrated that the expression of circ-ITCH was significantly
decreased in lung cancer tissue compared with normal lung tissue
(54).
circRNAs and other types of
cancer
By microarray analysis, Qu et al (55) identified that numerous circRNAs were
dysregulated in pancreatic ductal adenocarcinoma (PDAC), and these
results implied their vital functions in the progression of
PDAC.
Hsa_circ_002059, a representative circRNA, was
downregulated in GC and its differential expression was
significantly correlated with distal metastasis and
tumor-node-metastasis stage (56),
suggesting it could potentially serve as a diagnostic biomarker for
GC.
Zhong et al (57) screened differential circRNA expression
profiles in bladder carcinoma and revealed that the expression of
family with sequence similarity 169 member A circRNA and tripartite
motif-containing 24 circRNA were significantly downregulated in
bladder carcinoma, while the expression levels of transcription
factor (TCF)25 circRNA (circ-TCF25), zinc finger RNA-binding
protein circRNA, protein tyrosine kinase 2 circRNA and BC048201
circRNA were significantly increased in carcinoma tissues. This
suggested that circ-TCF25 could represent a novel biomarker for
bladder cancer.
Ahmed et al (58) revealed that altered expression
patterns of circRNAs were present in primary and metastatic sites
of epithelial ovarian carcinoma. In addition, circRNA expression
was inversely associated with linear RNA expression for numerous
cancer-associated signaling pathways; the majority of these
signaling pathways, such as the transforming growth factor β1
signaling pathway, were typically activated by linear RNA (mRNA)
expression in metastases compared to primary site of origin, but
repressed by circRNA expression (58).
Xuan et al (59) demonstrated that hsa_circ_100855
expression was significantly increased in laryngeal squamous cell
cancer tissues compared with that in matched adjacent
non-neoplastic tissues, and increased hsa_circ_100855 expression
was revealed in patients with T3-4 stage disease, neck nodal
metastasis, or advanced clinical stage. However, hsa_circ_104912
expression exhibited the reverse trend compared with
hsa_circ_100855 (59).
In breast cancer, circRNA expression profiling
results revealed that normal-adjacent tissues in patients with the
estrogen receptor (ER)+ subtype were associated with an
increased number of circRNAs compared with that in breast cancer
tissues, and the number of circRNAs in normal-adjacent samples of
ER+ subtype was inversely correlated with the
risk-of-relapse proliferation score for proliferation-associated
genes (60). Yang et al
(61) revealed that zinc finger
protein 292 circRNA could suppress human glioma tube formation via
the Wnt/β-catenin signaling pathway and may potentially be applied
as a therapeutic target and biomarker in glioma.
However, the association between certain circRNA and
tumorigenesis is unclear and supported by sufficient evidence.
Further studies will provide a more extensive awareness of circRNAs
in different types of cancer, including their properties, functions
and mechanisms.
Function of circRNAs as competing endogenous
RNAs in cancer
Currently, studies on the function and mechanism of
circRNAs are few. Since circRNAs may be considered alternative
splicing isoforms, they may regulate alternative splicing in
certain diseases, including cancer (30,32).
Certain circRNAs, including sirtuin (SIRT)7 circRNA and
minichromosome maintenance complex component (MCM)5 circRNA, may
enhance the expression of their parental mRNAs, while the parental
linear mRNAs may be associated with tumorigenesis, functioning as a
tumor suppressor (e.g. SIRT7) or promoter (e.g. MCM5) (30,32,62).
circRNAs mainly function as ceRNAs in cancer by competing with
other RNAs to bind miRNAs and RBPs (Fig.
2A-C).
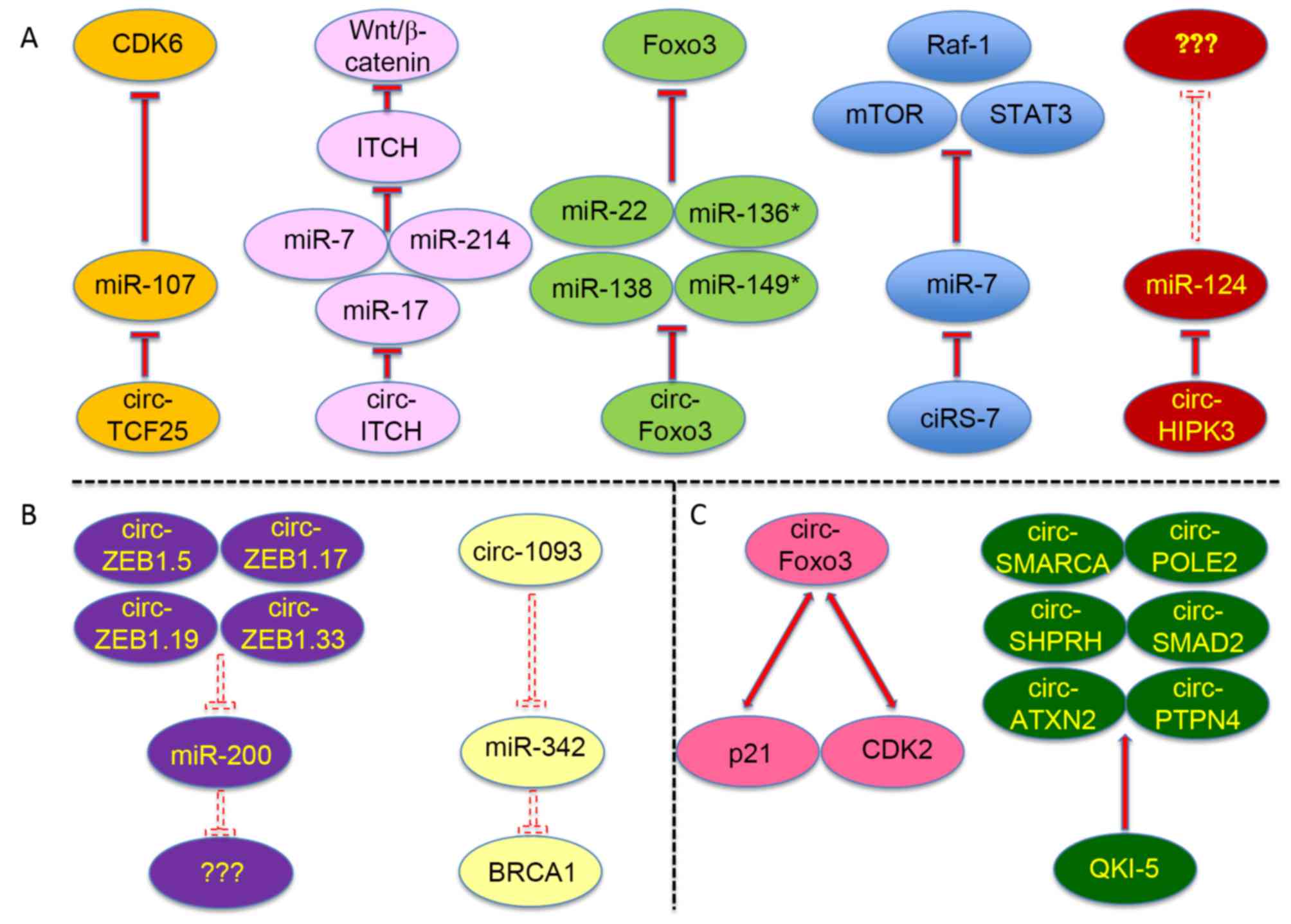 | Figure 2.Circular RNAs function as competing
endogenous RNAs in multiple types of cancer. The specific microRNAs
and RNA-binding proteins sponged by certain circular RNAs are
shown, as well as the protein targets of certain sponged miRNAs.
(A) Specific and validated microRNAs sponged by certain circular
RNAs, as well as the protein targets of certain sponged miRNAs. (B)
Predicted microRNAs sponged by certain circular RNAs. (C) Validated
and specific RNA-binding proteins sponged by certain circular RNAs.
The dashed inhibition arrows represent predicted but not validated
inhibitory effects, and the solid inhibition arrows represent that
the inhibitory effects have been experimentally validated. ‘???’
denotes that the targeted genes of the microRNAs were not addressed
in the corresponding reference. circ, circular RNA; miR, microRNA;
CDK, cyclin-dependent kinase; TCF25, transcription factor 25; ITCH,
itchy E3 ubiquitin protein ligase; Foxo3, forkhead box O3; Raf-1,
Raf-1 proto-oncogene; mTOR, mechanistic target of rapamycin; STAT3,
signal transducer and activator of transcription 3; HIPK3,
homeodomain-interacting protein kinase 3; ZEB, zinc finger E-box
binding homeobox; SMARCA, SWI/SNF-related, matrix-associated,
actin-dependent regulator of chromatin, subfamily A; POLE2, DNA
polymerase ε2, accessory subunit; SHPRH, SNF2 histone linker PHD
RING helicase; SMAD2, SMAD family member 2; ATXN2, ataxin 2; PTPN4,
protein tyrosine phosphatase, non-receptor type 4; QKI-5, Quaking
5. |
circRNAs as miRNA sponges in
cancer
miRNAs, a large class of sncRNAs, serve important
functions in multiple types of cancer by repressing downstream
cancer-associated mRNAs (63). Due to
the complementarity between circRNAs and targeted genes, circRNAs
may bind miRNAs and competitively suppress the function of targeted
miRNAs, thereby permitting circRNAs to regulate the progression of
cancer by sequestering specific miRNAs associated with
proliferation, differentiation, metastasis and carcinogenesis.
Human ciRS-7/CDR1-AS is a typical example of a
circRNA that functions as an miRNA sponge, and comprises as many as
74 miR-7-binding sites (11,19,62). miR-7
may serve as a tumor suppressor, and is downregulated in various
types of cancer, including GC (64),
HCC (65) and CRC (66). miR-7 is involved in numerous
cancer-associated signaling pathways by downregulating the
expression of crucial oncogenic factors, including Raf-1
proto-oncogene (67), mechanistic
target of rapamycin (65) and signal
transducer and activator of transcription 3 (68). Therefore, ciRS-7 could regulate cancer
progression through miR-7-mediated downstream signaling pathways.
However, other reports have revealed a tumor-promoting function of
miR-7 in CRC (69), human papilloma
virus-positive HeLa cells (70) and
A549 cells (71). miR-7 is coupled to
ciRS-7 through multiple miRNA response elements in ciRS-7, and
ciRS-7 may either promote cancer progression or suppress
carcinogenesis depending on the expression of miRNA targets
(11,19,62).
Li et al (50)
demonstrated that circ-ITCH may serve a tumor-suppressive function
in ESCC and serve as a sponge for miR-7, miR-17 and miR-214,
resulting in increased ITCH expression, thereby promoting
ubiquitin-mediated degradation of disheveled segment polarity
protein 2, and inhibiting the Wnt/β-catenin signaling pathway. This
mechanism for circ-ITCH was also verified in lung cancer (54). Similarly, Yang et al (40) revealed that circ-FOXO3 may also
function as a sponge of miRNAs to increase FOXO3 translation and
thereby suppress tumor growth, cancer cell proliferation and
survival. In addition, Zhong et al (57) demonstrated in bladder cancer that the
overexpression of circ-TCF25 could decrease the expression of
miR-103a-3p and miR-107, increase the expression of
cyclin-dependent kinase (CDK)6 and promote the proliferation and
migration of cancer cells in vitro and in vivo. Zheng
et al (39) also revealed that
circ-HIPK3 sponged 9 miRNAs and directly bound miR-124 and
inhibited its activity.
Although multiple circRNAs have been demonstrated to
regulate cancer progression in vitro or in vivo
(53,17,72),
certain predicted circRNAs remain to be validated. For example, Liu
et al (53) proposed that
circ-ZEB1.5, circ-ZEB1.19, circ-ZEB1.17 and circ-ZEB1.33 possess
the potential to sponge miR-200. Zhang et al (17) revealed that circRNA_1093, with a short
sequence of 108 bp, has 4 potential miR-342 binding sites, while
miR-342 has been reported to regulate the expression of BRCA1 in
breast cancer (72). These circRNAs
require further study to confirm their function and molecular
mechanisms. Certainly, the circRNAs that function as miRNA sponges
provide a novel direction for diagnosing and treating multiple
types of cancer and the circRNA-miRNA-gene regulatory networks in
cancer require further study.
circRNAs as RBP sponges in cancer
circRNAs have also been associated with another
important function: Acting as RBP sponges to regulate downstream
gene transcription (12). Multiple
studies have indicated that certain circRNAs may be associated with
RBPs, such as argonaute (AGO) proteins (11,19),
Quaking (QKI) (73), RNA polymerase
II (30), mannose-binding lectin 2
(74) and eukaryotic translation
initiation factor 4A3 (75). These
circRNAs can serve as carriers to store, sort or deliver RBPs to
particular subcellular locations, or regulate the function of RBPs
by functioning as ceRNAs (11,76).
Certain RBPs serve key regulatory functions in cell proliferation,
differentiation, apoptosis and migration (77,78). Their
dysregulated expression may affect cancer development (79,80). For
example, AGO proteins may regulate tumorigenesis via
miRNA-dependent or independent signaling pathways and AGO2 was
shown to be associated with cancer progression by interacting with
RISC-loading complex subunit TARBP2 and endoribonuclease Dicer
(81).
As an RBP, QKI-5 has been characterized as a novel
tumor suppressor in multiple types of cancer. For example, QKI-5
has been demonstrated to inhibit the proliferation of lung cancer
cells (82), and the proliferation,
cell cycle progression and invasion of prostate cancer cells
(83). Furthermore, QKI-5 was shown
to regulate the formation of circRNAs, and the upregulation of the
majority of circRNAs during the epithelial-mesenchymal transition
(EMT) suggests they may serve a crucial function in EMT (73). Since EMT serves an important function
in initiating metastasis (84,85),
further study is required to elucidate the function of QKI-mediated
circRNAs in cancer. In addition, Du et al (41) also revealed that circ-FOXO3 could
decelerate cell cycle progression by forming ternary complexes with
p21 and CDK2 proteins.
These studies suggest that RBPs and circRNAs could
interact reciprocally to affect cancer progression.
Prospects and challenges
Increasing numbers of circRNAs are being identified,
and certain circRNAs have been revealed to be abundant in clinical
blood samples (86), saliva (35) and exosomes (32). Considering their covalently closed
loop structures, stability and cell/tissue type-specific manner
(21,22,34,35,11,13,14,37),
circRNAs are stable, sensitive, specific and are easily detectable.
Detecting circRNAs early in samples of bodily fluids may render
circRNAs promising diagnostic biomarkers for different types of
cancer.
Currently, the concept of ceRNAs as regulators is
novel (87,88) and controversial (89). However, the stability and other
particular properties of circRNAs may confer ceRNA functions with
additional advantages for effective binding with miRNA/RBP,
exhibiting long-term effects. Given the important functions of
circRNAs as miRNA and RBP sponges, the functional potential of
circRNAs could be developed for cancer therapy. For example, an
artificial circRNA (90,91) functioning as a ‘super-sponge’ or
silencing the circRNA in cancer cells could regulate the expression
of miRNAs and other RNAs or RBPs and increase the activities of
suppressor genes or decrease the activities of oncogenes. Although
synthetic miRNA inhibitors for treating human diseases have already
been clinically trialed in phase 2a (92), small RNA-based drugs, including those
based on miRNAs, circRNAs and piRNAs, are associated with different
challenges due to the lack of targeted delivery strategies in
cancer therapy (93). However,
delivery systems mediated by nano materials or microbodies
(including exosomes) are providing promising treatment directions
(32).
Presently, multiple circRNAs are undergoing initial
exploration and the molecular mechanisms underlying the functions
of circRNAs in cancer progression remain to be fully understood.
The strategies for assessing circRNAs remain limited and have
developmental potential (13,15). High-throughput RNA sequencing
technology combined with bioinformatics analysis may be applied to
predict circRNAs, and multiple circRNA-associated databases have
been developed for public use, including circRNAFinder (18), Circ2Traits (94), find_circ (11), CIRCexplorer (22), CIRI (95), starBase (96), CircNet (53) and circBase (97). Identifying more cancer-associated
circRNAs and evaluating their underlying mechanisms may assist
healthcare professionals in clinically applying circRNAs in cancer
diagnosis and therapy.
Acknowledgements
The present study was supported by the International
Cooperation Key Project of the National Natural Science Foundation
of China (grant no. 81520108031), the National Natural Science
Foundation of China (grant nos. 81303102, 81473478, 81473628,
81573749 and 81573805), the Key Project of Shanghai Municipal
Science and Technology Commission (grant no. 16401970500), the
Three Year Plan of Action for Innovation of Traditional Chinese
Medicine in Shanghai (grant no. ZY3-CCCX-3-3012), the Program of
Shanghai Academic Research Leader (grant no. 16XD1403600) and the
Program for Shanghai Outstanding Medical Academic Leader.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dušek L, Mužík J, Malúšková D, Májek O,
Pavlík T, Koptíková J, Gregor J, Brabec P and Abrahámová J:
Epidemiology of screening-targeted cancers according to new data of
the Czech National Cancer Registry. Klin Onkol. 27 Suppl 2:S19–S39.
2014. View Article : Google Scholar
|
|
3
|
Rosenberg AR, Kroon L, Chen L, Li CI and
Jones B: Insurance status and risk of cancer mortality among
adolescents and young adults. Cancer. 121:1279–1286. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
van der Weyden L, Arends MJ, Rust AG,
Poulogiannis G, McIntyre RE and Adams DJ: Increased tumorigenesis
associated with loss of the tumor suppressor gene Cadm1. Mol
Cancer. 11:292012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chan XH, Nama S, Gopal F, Rizk P, Ramasamy
S, Sundaram G, Ow GS, Ivshina AV, Tanavde V, Haybaeck J, et al:
Targeting glioma stem cells by functional inhibition of a
prosurvival oncomiR-138 in malignant gliomas. Cell Rep. 2:591–602.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shroff EH, Eberlin LS, Dang VM, Gouw AM,
Gabay M, Adam SJ, Bellovin DI, Tran PT, Philbrick WM, Garcia-Ocana
A, et al: MYC oncogene overexpression drives renal cell carcinoma
in a mouse model through glutamine metabolism. Proc Natl Acad Sci
USA. 112:pp. 6539–6544. 2015; View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hashim A, Rizzo F, Marchese G, Ravo M,
Tarallo R, Nassa G, Giurato G, Santamaria G, Cordella A, Cantarella
C and Weisz A: RNA sequencing identifies specific PIWI-interacting
small non-coding RNA expression patterns in breast cancer.
Oncotarget. 5:9901–9910. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun QL, Zhao CP, Wang TY, Hao XB, Wang XY,
Zhang X and Li YC: Expression profile analysis of long non-coding
RNA associated with vincristine resistance in colon cancer cells by
next-generation sequencing. Gene. 572:79–86. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Brosnan CA and Voinnet O: The long and the
short of noncoding RNAs. Curr Opin Cell Biol. 21:416–425. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gibb EA, Brown CJ and Lam WL: The
functional role of long non-coding RNA in human carcinomas. Mol
Cancer. 10:382011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qu S, Yang X, Li X, Wang J, Gao Y, Shang
R, Sun W, Dou K and Li H: Circular RNA: A new star of noncoding
RNAs. Cancer Lett. 365:141–148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jeck WR, Sorrentino JA, Wang K, Slevin MK,
Burd CE, Liu J, Marzluff WF and Sharpless NE: Circular RNAs are
abundant, conserved, and associated with ALU repeats. RNA.
19:141–157. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Salzman J, Chen RE, Olsen MN, Wang PL and
Brown PO: Cell-type specific features of circular RNA expression.
PLoS Genet. 9:e10037772013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guo JU, Agarwal V, Guo H and Bartel DP:
Expanded identification and characterization of mammalian circular
RNAs. Genome Biol. 15:4092014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rybak-Wolf A, Stottmeister C, Glažar P,
Jens M, Pino N, Giusti S, Hanan M, Behm M, Bartok O, Ashwal-Fluss
R, et al: Circular RNAs in the mammalian brain are highly abundant,
conserved, and dynamically expressed. Mol Cell. 58:870–885. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang C, Wu H, Wang Y, Zhao Y, Fang X,
Chen C and Chen H: Expression patterns of circular RNAs from
primary kinase transcripts in the mammary glands of lactating rats.
J Breast Cancer. 18:235–241. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Westholm JO, Miura P, Olson S, Shenker S,
Joseph B, Sanfilippo P, Celniker SE, Graveley BR and Lai EC:
Genome-wide analysis of drosophila circular RNAs reveals their
structural and sequence properties and age-dependent neural
accumulation. Cell Rep. 9:1966–1980. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen LL and Yang L: Regulation of circRNA
biogenesis. RNA Biol. 12:381–388. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jeck WR and Sharpless NE: Detecting and
characterizing circular RNAs. Nat Biotechnol. 32:453–461. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang XO, Wang HB, Zhang Y, Lu X, Chen LL
and Yang L: Complementary sequence-mediated exon circularization.
Cell. 159:134–147. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Graveley BR: Molecular biology: Power
sequencing. Nature. 453:1197–1198. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wilhelm BT, Marguerat S, Watt S, Schubert
F, Wood V, Goodhead I, Penkett CJ, Rogers J and Bähler J: Dynamic
repertoire of a eukaryotic transcriptome surveyed at
single-nucleotide resolution. Nature. 453:1239–1243. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Salzman J, Gawad C, Wang PL, Lacayo N and
Brown PO: Circular RNAs are the predominant transcript isoform from
hundreds of human genes in diverse cell types. PLoS One.
7:e307332012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ivanov A, Memczak S, Wyler E, Torti F,
Porath HT, Orejuela MR, Piechotta M, Levanon EY, Landthaler M,
Dieterich C and Rajewsky N: Analysis of intron sequences reveals
hallmarks of circular RNA biogenesis in animals. Cell Rep.
10:170–177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Danan M, Schwartz S, Edelheit S and Sorek
R: Transcriptome-wide discovery of circular RNAs in Archaea.
Nucleic Acids Res. 40:3131–3142. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang L, Duff MO, Graveley BR, Carmichael
GG and Chen LL: Genomewide characterization of non-polyadenylated
RNAs. Genome Biol. 12:R162011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kelly S, Greenman C, Cook PR and
Papantonis A: Exon skipping is correlated with exon
circularization. J Mol Biol. 427:2414–2417. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin
QF, Xing YH, Zhu S, Yang L and Chen LL: Circular intronic long
noncoding RNAs. Mol Cell. 51:792–806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Z, Huang C, Bao C, Chen L, Lin M, Wang
X, Zhong G, Yu B, Hu W, Dai L, et al: Exon-intron circular RNAs
regulate transcription in the nucleus. Nat Struct Mol Biol.
22:256–264. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J,
Chen D, Gu J, He X and Huang S: Circular RNA is enriched and stable
in exosomes: A promising biomarker for cancer diagnosis. Cell Res.
25:981–984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Burd CE, Jeck WR, Liu Y, Sanoff HK, Wang Z
and Sharpless NE: Expression of linear and novel circular forms of
an INK4/ARF-associated non-coding RNA correlates with
atherosclerosis risk. PLoS Genet. 6:e10012332010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Suzuki H and Tsukahara T: A view of
pre-mRNA splicing from RNase R resistant RNAs. Int J Mol Sci.
15:9331–9342. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bahn JH, Zhang Q, Li F, Chan TM, Lin X,
Kim Y, Wong DT and Xiao X: The landscape of microRNA,
Piwi-interacting RNA, and circular RNA in human saliva. Clin Chem.
61:221–230. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lasda E and Parker R: Circular RNAs:
Diversity of form and function. RNA. 20:1829–1842. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang PL, Bao Y, Yee MC, Barrett SP, Hogan
GJ, Olsen MN, Dinneny JR, Brown PO and Salzman J: Circular RNA is
expressed across the eukaryotic tree of life. PLoS One.
9:e908592014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Perkel JM: Assume nothing: The tale of
circular RNA. Biotechniques. 55:55–57. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen
B, Luo Y, Lyu D, Li Y, Shi G, et al: Circular RNA profiling reveals
an abundant circHIPK3 that regulates cell growth by sponging
multiple miRNAs. Nat Commun. 7:112152016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang W, Du WW, Li X, Yee AJ and Yang BB:
Foxo3 activity promoted by non-coding effects of circular RNA and
Foxo3 pseudogene in the inhibition of tumor growth and
angiogenesis. Oncogene. 35:3919–3931. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Du WW, Yang W, Liu E, Yang Z, Dhaliwal P
and Yang BB: Foxo3 circular RNA retards cell cycle progression via
forming ternary complexes with p21 and CDK2. Nucleic Acids Res.
44:2846–2858. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Du WW, Fang L, Yang W, Wu N, Awan FM, Yang
Z and Yang BB: Induction of tumor apoptosis through a circular RNA
enhancing Foxo3 activity. Cell Death Differ. 24:357–370. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Bachmayr-Heyda A, Reiner AT, Auer K,
Sukhbaatar N, Aust S, Bachleitner-Hofmann T, Mesteri I, Grunt TW,
Zeillinger R and Pils D: Correlation of circular RNA abundance with
proliferation-Exemplified with colorectal and ovarian cancer,
idiopathic lung fibrosis, and normal human tissues. Sci Rep.
5:80572015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Huang G, Zhu H, Shi Y, Wu W, Cai H and
Chen X: cir-ITCH plays an inhibitory role in colorectal cancer by
regulating the Wnt/β-catenin pathway. PLoS One. 10:e01312252015.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang X, Zhang Y, Huang L, Zhang J, Pan F,
Li B, Yan Y, Jia B, Liu H, Li S and Zheng W: Decreased expression
of hsa_circ_001988 in colorectal cancer and its clinical
significances. Int J Clin Exp Pathol. 8:16020–16025.
2015.PubMed/NCBI
|
|
46
|
Dou Y, Cha DJ, Franklin JL, Higginbotham
JN, Jeppesen DK, Weaver AM, Prasad N, Levy S, Coffey RJ, Patton JG
and Zhang B: Circular RNAs are down-regulated in KRAS mutant colon
cancer cells and can be transferred to exosomes. Sci Rep.
6:379822016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Qin M, Liu G, Huo X, Tao X, Sun X, Ge Z,
Yang J, Fan J, Liu L and Qin W: Hsa_circ_0001649: A circular RNA
and potential novel biomarker for hepatocellular carcinoma. Cancer
Biomark. 16:161–169. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xu L, Zhang M, Zheng X, Yi P, Lan C and Xu
M: The circular RNA ciRS-7 (Cdr1as) acts as a risk factor of
hepatic microvascular invasion in hepatocellular carcinoma. J
Cancer Res Clin Oncol. 143:17–27. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shang X, Li G, Liu H, Li T, Liu J, Zhao Q
and Wang C: Comprehensive circular RNA profiling reveals that
hsa_circ_0005075, a new circular RNA biomarker, is involved in
hepatocellular crcinoma development. Medicine (Baltimore).
95:e38112016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li F, Zhang L, Li W, Deng J, Zheng J, An
M, Lu J and Zhou Y: Circular RNA ITCH has inhibitory effect on ESCC
by supupressing the Wnt/β-catenin pathway. Oncotarget. 6:6001–6013.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Su H, Lin F, Deng X, Shen L, Fang Y, Fei
Z, Zhao L, Zhang X, Pan H, Xie D, et al: Profiling and
bioinformatics analyses reveal differential circular RNA expression
in radioresistant esophageal cancer cells. J Transl Med.
14:2252016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xia W, Qiu M, Chen R, Wang S, Leng X, Wang
J, Xu Y, Hu J, Dong G, Xu PL and Yin R: Circular RNA
has_circ_0067934 is upregulated in esophageal squamous cell
carcinoma and promoted proliferation. Sci Rep. 6:355762016.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liu YC, Li JR, Sun CH, Andrews E, Chao RF,
Lin FM, Weng SL, Hsu SD, Huang CC, Cheng C, et al: CircNet: A
database of circular RNAs derived from transcriptome sequencing
data. Nucleic Acids Res. 44:D209–D215. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wan L, Zhang L, Fan K, Cheng ZX, Sun QC
and Wang JJ: Circular RNA-ITCH suppresses lung cancer proliferation
via inhibiting the Wnt/β-catenin pathway. Biomed Res Int.
2016:15794902016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Qu S, Song W, Yang X, Wang J, Zhang R,
Zhang Z, Zhang H and Li H: Microarray expression profile of
circular RNAs in human pancreatic ductal adenocarcinoma. Genom
Data. 5:385–387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Li P, Chen S, Chen H, Mo X, Li T, Shao Y,
Xiao B and Guo J: Using circular RNA as a novel type of biomarker
in the screening of gastric cancer. Clin Chim Acta. 444:132–136.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhong Z, Lv M and Chen J: Screening
differential circular RNA expression profiles reveals the
regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in
bladder carcinoma. Sci Rep. 6:309192016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ahmed I, Karedath T, Andrews SS, Al-Azwani
IK, Mohamoud YA, Querleu D, Rafii A and Malek JA: Altered
expression pattern of circular RNAs in primary and metastatic sites
of epithelial ovarian carcinoma. Oncotarget. 7:36366–36381. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Xuan L, Qu L, Zhou H, Wang P, Yu H, Wu T,
Wang X, Li Q, Tian L, Liu M and Sun Y: Circular RNA: A novel
biomarker for progressive laryngeal cancer. Am J Transl Res.
8:932–939. 2016.PubMed/NCBI
|
|
60
|
Nair AA, Niu N, Tang X, Thompson KJ, Wang
L, Kocher JP, Subramanian S and Kalari KR: Circular RNAs and their
associations with breast cancer subtypes. Oncotarget.
7:80967–80979. 2016.PubMed/NCBI
|
|
61
|
Yang P, Qiu Z, Jiang Y, Dong L, Yang W, Gu
C, Li G and Zhu Y: Silencing of cZNF292 circular RNA suppresses
human glioma tube formation via the Wnt/β-catenin signaling
pathway. Oncotarget. 7:63449–63455. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Chen I, Chen CY and Chuang TJ: Biogenesis,
identification, and function of exonic circular RNAs. Wiley
Interdiscip Rev RNA. 6:563–579. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Romero-Cordoba SL, Salido-Guadarrama I,
Rodriguez-Dorantes M and Hidalgo-Miranda A: miRNA biogenesis:
Biological impact in the development of cancer. Cancer Biol Ther.
15:1444–1455. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kong D, Piao YS, Yamashita S, Oshima H,
Oguma K, Fushida S, Fujimura T, Minamoto T, Seno H, Yamada Y, et
al: Inflammation-induced repression of tumor suppressor miR-7 in
gastric tumor cells. Oncogene. 31:3949–3960. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Fang Y, Xue JL, Shen Q, Chen J and Tian L:
MicroRNA-7 inhibits tumor growth and metastasis by targeting the
phosphoinositide 3-kinase/Akt pathway in hepatocellular carcinoma.
Hepatology. 55:1852–1862. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhang N, Li X, Wu C, Dong Y, Cai M, Mok M,
Wang H, Chen J, Ng SS, Chen M, et al: microRNA-7 is a novel
inhibitor of YY1 contributing to colorectal tumorigenesis.
Oncogene. 32:5078–5088. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Webster RJ, Giles KM, Price KJ, Zhang PM,
Mattick JS and Leedman PJ: Regulation of epidermal growth factor
receptor signaling in human cancer cells by microRNA-7. J Biol
Chem. 284:5731–5741. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhang H, Cai K, Wang J, Wang X, Cheng K,
Shi F, Jiang L, Zhang Y and Dou J: MiR-7, inhibited indirectly by
lincRNA HOTAIR, directly inhibits SETDB1 and reverses the EMT of
breast cancer stem cells by downregulating the STAT3 pathway. Stem
Cells. 32:2858–2868. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Nakagawa Y, Akao Y, Taniguchi K, Kamatani
A, Tahara T, Kamano T, Nakano N, Komura N, Ikuno H, Ohmori T, et
al: Relationship between expression of onco-related miRNAs and the
endoscopic appearance of colorectal tumors. Int J Mol Sci.
16:1526–1543. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Honegger A, Schilling D, Bastian S,
Sponagel J, Kuryshev V, Sültmann H, Scheffner M, Hoppe-Seyler K and
Hoppe-Seyler F: Dependence of intracellular and exosomal microRNAs
on Viral E6/E7 oncogene expression in HPV-positive tumor cells.
PLoS Pathog. 11:e10047122015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Meza-Sosa KF, Pérez-García EI,
Camacho-Concha N, López-Gutiérrez O, Pedraza-Alva G and
Pérez-Martínez L: MiR-7 promotes epithelial cell transformation by
targeting the tumor suppressor KLF4. PLoS One. 9:e1039872014.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Crippa E, Lusa L, De Cecco L, Marchesi E,
Calin GA, Radice P, Manoukian S, Peissel B, Daidone MG, Gariboldi M
and Pierotti MA: miR-342 regulates BRCA1 expression through
modulation of ID4 in breast cancer. PLoS One. 9:e870392014.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Conn SJ, Pillman KA, Toubia J, Conn VM,
Salmanidis M, Phillips CA, Roslan S, Schreiber AW, Gregory PA and
Goodall GJ: The RNA binding protein quaking regulates formation of
circRNAs. Cell. 160:1125–1134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Ashwal-Fluss R, Meyer M, Pamudurti NR,
Ivanov A, Bartok O, Hanan M, Evantal N, Memczak S, Rajewsky N and
Kadener S: circRNA biogenesis competes with pre-mRNA splicing. Mol
Cell. 56:55–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Dudekula DB, Panda AC, Grammatikakis I, De
S, Abdelmohsen K and Gorospe M: CircInteractome: A web tool for
exploring circular RNAs and their interacting proteins and
microRNAs. RNA Biol. 13:34–42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hentze MW and Preiss T: Circular RNAs:
Splicing's enigma variations. EMBO J. 32:923–925. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Glisovic T, Bachorik JL, Yong J and
Dreyfuss G: RNA-binding proteins and post-transcriptional gene
regulation. FEBS Lett. 582:1977–1986. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Abdelmohsen K, Kuwano Y, Kim HH and
Gorospe M: Posttranscriptional gene regulation by RNA-binding
proteins during oxidative stress: Implications for cellular
senescence. Biol Chem. 389:243–255. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Kim MY, Hur J and Jeong S: Emerging roles
of RNA and RNA-binding protein network in cancer cells. BMB Rep.
42:125–130. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Calabretta S and Richard S: Emerging roles
of disordered sequences in RNA-binding proteins. Trends Biochem
Sci. 40:662–672. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Ye Z, Jin H and Qian Q: Argonaute 2: A
novel rising star in cancer research. J Cancer. 6:877–882. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zong FY, Fu X, Wei WJ, Luo YG, Heiner M,
Cao LJ, Fang Z, Fang R, Lu D, Ji H and Hui J: The RNA-binding
protein QKI suppresses cancer-associated aberrant splicing. PLoS
Genet. 10:e10042892014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Zhao Y, Zhang G, Wei M, Lu X, Fu H, Feng
F, Wang S, Lu W, Wu N, Lu Z and Yuan J: The tumor suppressing
effects of QKI-5 in prostate cancer: A novel diagnostic and
prognostic protein. Cancer Biol Ther. 15:108–118. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Rokavec M, Öner MG, Li H, Jackstadt R,
Jiang L, Lodygin D, Kaller M, Horst D, Ziegler PK, Schwitalla S, et
al: IL-6R/STAT3/miR-34a feedback loop promotes EMT-mediated
colorectal cancer invasion and metastasis. J Clin Invest.
124:1853–1867. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Gujral TS, Chan M, Peshkin L, Sorger PK,
Kirschner MW and MacBeath G: A noncanonical Frizzled2 pathway
regulates epithelial-mesenchymal transition and metastasis. Cell.
159:844–856. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Memczak S, Papavasileiou P, Peters O and
Rajewsky N: Identification and characterization of circular RNAs as
a new class of putative biomarkers in human blood. PLoS One.
10:e01412142015. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Ebert MS and Sharp PA: Emerging roles for
natural microRNA sponges. Curr Biol. 20:R858–R861. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Cheng DL, Xiang YY, Ji LJ and Lu XJ:
Competing endogenous RNA interplay in cancer: Mechanism,
methodology, and perspectives. Tumour Biol. 36:479–488. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Denzler R, Agarwal V, Stefano J, Bartel DP
and Stoffel M: Assessing the ceRNA hypothesis with quantitative
measurements of miRNA and target abundance. Mol Cell. 5:766–776.
2014. View Article : Google Scholar
|
|
90
|
Errichelli L, Modigliani S Dini, Laneve P,
Colantoni A, Legnini I, Capauto D, Rosa A, De Santis R, Scarfò R,
Peruzzi G, et al: FUS affects circular RNA expression in murine
embryonic stem cell-derived motor neurons. Nat Commun. 8:147412017.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Tatomer DC, Liang D and Wilusz JE:
Inducible expression of eukaryotic circular RNAs from plasmids.
Methods Mol Biol. 1648:143–154. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Janssen HL, Reesink HW, Lawitz EJ, Zeuzem
S, Rodriguez-Torres M, Patel K, van der Meer AJ, Patick AK, Chen A,
Zhou Y, et al: Treatment of HCV infection by targeting microRNA. N
Engl J Med. 368:1685–1694. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Gong Z, Yang J, Li J, Yang L, Le Y, Wang S
and Lin HK: Novel insights into the role of microRNA in lung cancer
resistance to treatment and targeted therapy. Curr Cancer Drug
Targets. 14:241–258. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Ghosal S, Das S, Sen R, Basak P and
Chakrabarti J: Circ2Traits: A comprehensive database for circular
RNA potentially associated with disease and traits. Front Genet.
4:2832013. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Gao Y, Wang J and Zhao F: CIRI: An
efficient and unbiased algorithm for de novo circular RNA
identification. Genome Biol. 16:42015. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42(Database Issue): D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Glažar P, Papavasileiou P and Rajewsky N:
circBase: A database for circular RNAs. RNA. 20:1666–1670. 2014.
View Article : Google Scholar : PubMed/NCBI
|