Human leukocyte antigen-G (HLA-G), a non-classical
major histocompatibility complex class I (MHC-I) antigen, has
well-recognized tolerogenic properties (1). HLA-G has been detected under
physiological conditions in fetal tissues, adult immune-privileged
organs and cells of the hematopoietic lineage (1), as well as under pathological conditions
in cancer, viral infections, inflammatory diseases, autoimmune
diseases and transplantation (2).
HLA-G was first detected at the maternal-fetal
interface in the trophoblast, possibly performing a role in the
modulation of the maternal immune system during pregnancy (3). The expression of HLA-G in fetal tissue
successfully suppressed the local immune response in the placenta,
therefore inducing maternal-fetal tolerance, preventing the fetus
from being recognized as a non-self tissue and protecting it from
lysis mediated by natural killer (NK) cells (4). Similarly, HLA-G expression in tumors can
also protect cancer cells from NK cell and cytotoxic T lymphocyte
(CTL)-mediated destruction (5). In
this context, HLA-G expression may be a mechanism used by cancer
cells to escape host immune surveillance.
Differing from classical HLA, the HLA-G gene
presents limited coding region variability (6). Variation at the 5′ upstream regulatory
region (5′URR) and 3′ untranslated region (3′UTR) has been observed
most frequently (6). Several lines of
evidence have indicated that HLA-G polymorphisms affect the
expression level of HLA-G, the production of different isoforms and
the pattern of alternative splicing (7,8).
Polymorphisms in HLA-G have been studied in pathophysiological
conditions, revealing that HLA-G polymorphisms are associated with
numerous types of disorders, including pre-eclampsia (9), recurrent spontaneous abortion (10), asthma (11), systemic lupus erythematosus (12) and pemphigus vulgaris (13). In addition, several isolated segments
of the HLA-G gene have been studied in different tumor types and
were identified to be associated with tumor progression (14).
The present review discusses HLA-G expression and
its gene polymorphisms in tumors, and analyzes its underlying
mechanisms in promoting tumor development. Furthermore, the
possible clinical applications of HLA-G (or sHLA-G) as a diagnostic
and prognostic biomarker for cancer, and as a potential therapeutic
target for cancer biotherapy are considered.
In healthy individuals, HLA-G was first detected in
extra-villous cytotrophoblast at the maternal-fetal interface,
being regarded as a molecule that modulates the maternal immune
system during pregnancy (3). It is
also expressed by amnion epithelial cells, endothelial cells of
fetal blood vessels in the placenta, erythroblasts, macrophages,
antigen-presenting cells and dendritic cells (DCs), as well as by
immune-privileged organs of adults, including the thymus, cornea
and pancreatic islets (22).
Additionally, ectopic HLA-G expression can be induced in various
pathological conditions, including cancer, viral infections,
inflammatory diseases, viral infections, autoimmune diseases and
transplantation (1).
Considered as immunomodulatory molecules, HLA-G
antigens can induce immunological tolerance by inhibiting certain
immune-competent cells (22). The
immune-suppressive function of HLA-G antigens may be mediated by
binding membrane-bound and sHLA-G to their specific inhibitory
receptors (23). Regarding these
receptors, HLA-G antigens preferentially act as ligands for
immunoglobulin-like transcript 2 (ILT2) and immunoglobulin-like
transcript 4 (ILT4) (23). ILT2 is
expressed by B cells/T cells/NK cells/monocytes/DCs, and ILT4 is
presented on monocytes/macrophages/DCs (24,25). In
addition, HLA-G antigens were also reported to be ligands for the
killer cell immunoglobulin-like receptor [KIR2DL4/p49 (CD158d)],
which is expressed on NK cells (26).
The direct interactions between HLA-G proteins and their specific
inhibitory receptors promote the maintenance of tolerance at
different stages of the immune response, consisting of
differentiation, proliferation, cytolysis and cytokine secretion
(27). It was also demonstrated that
HLA-G could modulate the differentiation of DCs by interacting with
ILT4, which requires the interleukin (IL)-6-signal transducer and
activator of transcription 3 (STAT3) signaling pathway and results
in the recruitment of Src homology region 2 domain-containing
phosphatase (SHP)-1 and SHP-2 protein tyrosine phosphatases
(28). Furthermore, the cell cycle of
human-activated T cells can be suppressed by activating SHP-2
phosphatase and inhibiting the mechanistic target of rapamycin
pathway mediated by HLA-G (29).
Notably, HLA-G may upregulate the inhibitory receptors in immune
cells, which may precede an immune response and be involved in
immune escape mechanisms by increasing their activation thresholds
(30).
In addition to these direct effects, HLA-G can
implement its indirect immune-inhibitory function through the
expression of non-classical HLA class I molecule HLA-E (16,31). HLA-E
can directly bind the peptides derived from HLA-G, and the
HLA-E/peptide complexes can interact with the inhibitory receptor
CD94/NKG2A, which is predominantly expressed on NK cells (31). Thus, the effect of HLA-E on immune
cells is of great importance for the inhibition of NK and T cell
reactivity (31). Indeed,
HLA-G-mediated immune tolerance may be extended to induce
regulatory T cells (Tregs), a subpopulation of T cells
possessing abilities to modulate the immune system, abrogate the
autoimmune reaction and maintain the tolerance-to-self antigens
(32). For instance, in the presence
of HLA-G, CD4+ and CD8+ T cells may lose
their capability to respond to antigenic stimulation and be
differentiated into Tregs (32).
Since the ectopic expression of HLA-G in tumor cells
was first described in melanoma by Paul et al (5) in 1998, a high frequency of HLA-G
expression and plasma sHLA-G levels have been observed successively
in different solid tumor types, as well as in hematological
malignancies, including melanoma (33,34),
breast cancer (35–44), lung cancer (45–49),
hepatocellular carcinoma (50–53),
colorectal cancer (49,54–58),
gastric cancer (49,59–61),
esophageal carcinoma (49,62–65),
nasopharyngeal carcinoma (66),
laryngeal lesions (67), bladder
transitional cell carcinoma (68),
renal cell carcinoma (69–72), cervical cancer (73), thyroid carcinoma (74,75),
neuroblastoma (76,77), glioblastoma (78–81) and
myeloid leukemia (82). HLA-G
expression in various cancer types is listed in Table I, and data regarding plasma sHLA-G
levels detected by ELISA are presented in Table II. As shown in Table II, in the majority of cancer types,
serum sHLA-G levels were significantly higher in patients with
cancer compared with in the healthy controls. Notably, sHLA-G
levels were similar between patients with bladder transitional cell
carcinoma (TCC) and healthy controls (68).
However, the associations between increased HLA-G
expression and the clinical parameters of patients with cancer
remain conflicting. Although there was a significant association
between increased HLA-G expression levels and certain clinical
parameters, including advanced disease stage, poor histological
grade, higher tumor grade, presence of metastasis, shorter survival
time, greater tumor size, tumor recurrence or tumor invasion in
many types of tumors (27), the
association was not detected in other tumors, including bladder TCC
(68) and acute myeloid leukemia
(82). Although several studies have
demonstrated that high serum sHLA-G levels were associated with
aggressive behavior in cancer, histological type,
tumor-node-metastasis stage or a shorter survival time of patients
suffering from breast cancer, papillary thyroid carcinoma (PTC) and
lung cancer among others, no clear associations were identified
between the plasma sHLA-G levels and the clinicopathological
parameters in colorectal cancer, hepatocellular carcinoma,
esophageal carcinoma, renal cell carcinoma and gastric cancer
(38,43,45,83).
Notably, numerous factors may affect sHLA-G expression (65,82). The
upregulated plasma IL-10 level in primary esophageal squamous cell
carcinoma was determined to be associated with high sHLA-G levels
(65). In addition, sHLA-G levels
have been established as associated with the deficiency of anterior
myelodysplasia along with higher leukocytosis in acute myeloid
leukemia (82). Notably,
contradictions exist in the same type of tumor between different
studies. For instance, Kren et al (70) confirmed that HLA-G is upregulated in
renal cell carcinoma tissues and that it is associated with a worse
prognosis. By contrast, Reimers et al (84) reported that weak HLA-G expression was
associated with a poor prognosis and a significantly worse
survival. These contradictions may possibly be attributed to
differing ethnicities in the patient cohorts, varied criteria of
patient selection, the methods used and factitious surgical
errors.
Compared with classical HLA, the HLA-G gene presents
limited coding region variability (6). However, variations at the 5′URR and at
the 3′UTR have been observed in numerous previous studies (6,85–87). Recently, a number of studies indicated
that HLA-G polymorphisms are associated with HLA-G expression
(41), cancer susceptibility
(88–91) and cancer development (86).
Although it remains a controversial issue as to
where HLA-G transcription begins, the polymorphisms at the 5′URR
and haplotypes at the 3′UTR are considered to affect HLA-G
expression (7). In healthy Brazilian
and French individuals, the 14-bp del/del genotype exhibited higher
sHLA-G levels compared with the 14-bp ins/ins genotype (85). Additionally, in patients with invasive
ductal carcinoma, 14-bp ins/del polymorphisms may induce a higher
expression of the HLA-G molecule compared with 14-bp ins/ins and
14-bp del/del polymorphisms (41).
The differential expression levels were attributed to the fact that
polymorphic sites at the 5′URR coincided with the known
transcription factor binding sites, and that the polymorphic sites
at 3′UTR affect mRNA stability and the binding of specific
microRNAs (85).
It has previously been demonstrated that 14-bp
polymorphisms may be a genetic risk factor for susceptibility to
several types of cancer (87,88,91). A
study in Australia and New Zealand identified an association
between heterozygote carriers of the HLA-G 14-bp ins/del
polymorphism and decreased risk of neuroblastoma (88). Another study demonstrated that the
14-bp del allele may promote human papillomavirus infection and the
14-bp del/+3142C haplotype may be involved in invasive cervical
cancer (ICC) development (91). In
addition, in Brazilian patients who smoke, the polymorphic 14-bp
ins/ins genotype and 14-bp ins/+3142 G haplotypes were associated
with an increased risk of high-grade squamous intraepithelial
lesion (HSIL) and a higher risk of developing ICC, while the 14-bp
del/del genotype and 14-bp del/+3142G haplotypes were associated
with a lower risk of HSIL and cervical cancer (86). By contrast, no significant differences
in the +14/−14 bp polymorphism frequencies were observed between
patients with PTC and the healthy control group. However, the
association between 14-bp polymorphisms and cancer susceptibility
is discrepant in distinct ethnic populations. The presence of the
HLA-G 3′UTR 14-bp sequence was observed to be associated with a
reduced risk of breast cancer susceptibility in HLA-G-expressing
tissues in Southeastern Iranian (92)
and Korean patients (43). However,
the association between HLA-G 3′UTR 14-bp sequence and breast
cancer susceptibility was not observed in Brazilian patients with
breast cancer (93). In addition, the
14-bp del/del allele was revealed to increase hepatocellular
carcinoma (HCC) susceptibility in Brazilian (94) and Chinese (87) patients, but the variation was not
associated with HLA-G expression and susceptibility to HCC in
Korean patients (95).
Other polymorphisms of HLA-G have also been
investigated. In Canadian populations with cervical cancer, the
homozygous genotype form of the HLA-G*01:01:02, -G*01:06 and
-G*3′UTR 14-bp insertions were identified to be associated with a
significantly increased risk for invasive cancer, compared with the
wild-type heterozygous form of the HLA-G*01:01:01 allele (89). Evaluating the effect of HLA-G
polymorphism occurrence on nasopharyngeal carcinoma (NPC)
susceptibility in Tunisian patients, researchers observed that the
lle110 allele was associated with fewer lymph node metastases and
an increased number of patients with higher tumor stages, and the
occurrence of codon 130C deletion reduced the disease-free and
overall survival rates of patients with NPC (90). To date, and to the best of our
knowledge, HLA-G polymorphisms have not been investigated in a
number of tumor types, including lung cancer, gastric cancer,
colorectal cancer, melanoma, renal cell carcinoma and
glioblastoma.
Therefore, HLA-G polymorphisms may be risk factors
and predictive markers for several types of cancer. However, the
conflicting results regarding the association between HLA-G
polymorphisms and cancer susceptibility indicate that additional
studies consisting of populations from different ethnicities and
larger sample sizes are required.
Since HLA-G expression was observed in various types
of cancer, the roles of HLA-G in promoting cancer progression were
investigated in a number of studies (17,96–98).
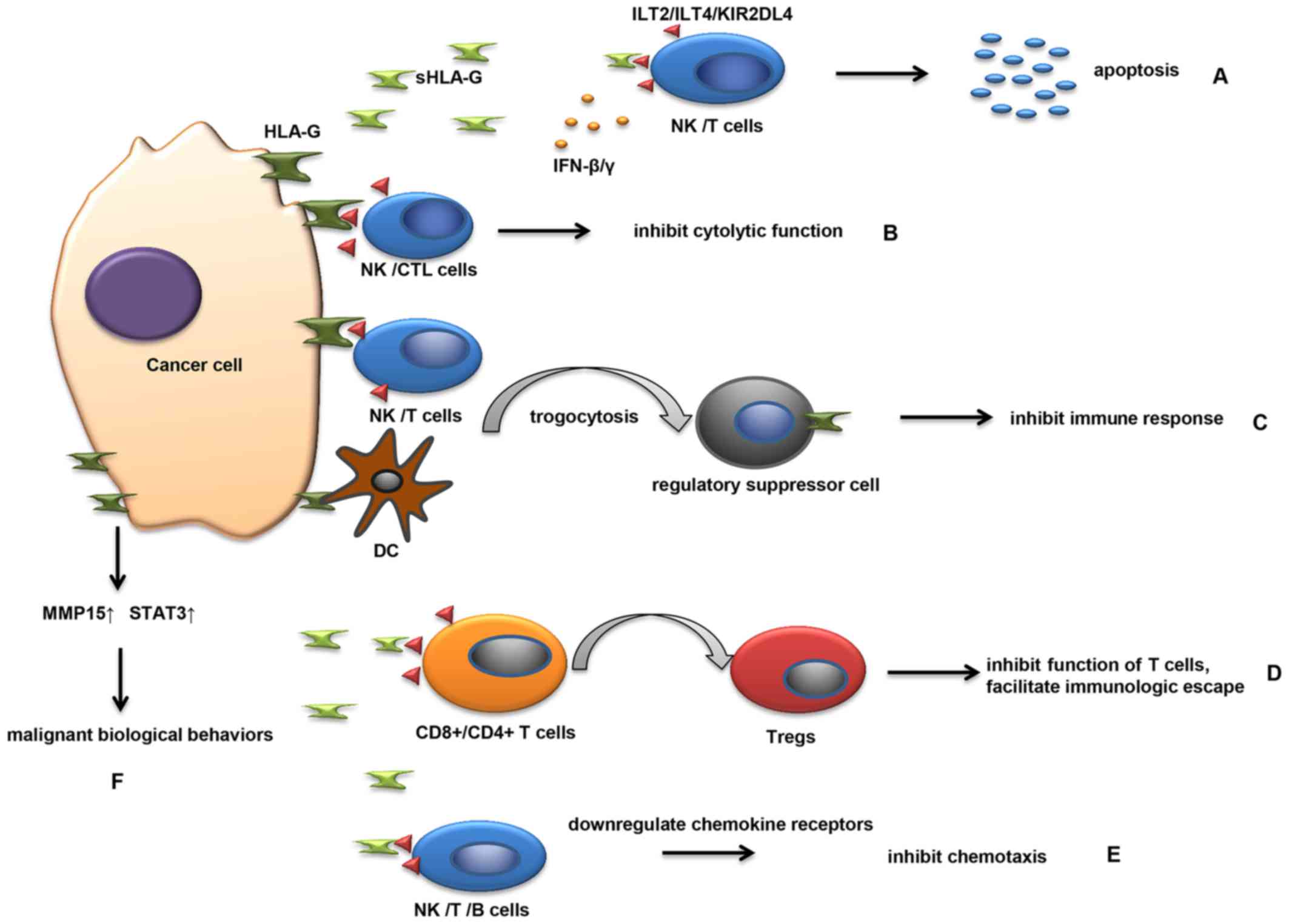
Studies revealed that HLA-G and sHLA-G have multiple mechanisms of
involvement in the development of malignancies (Fig. 1).
Available evidence has demonstrated that HLA-G may
be actively involved in the immune escape mechanism of tumor cells
(17). Cancer immunoediting is an
important host protection process (97). In this process, the immune
surveillance can be divided into three essential steps, including
an elimination phase, equilibrium phase and escape phase (97), and HLA-G is involved in each of these
three phases (2). Several
adaptive/innate immunizing molecules and cells are involved in
inhibiting the elimination of tumor cells (98). As mentioned earlier, HLA-G could bind
to its special receptors on immune effector cells, thereby blocking
the proliferation and lytic function of uterine and peripheral NK
cells in a physiological context (4).
Similarly, sHLA-G released by cancer cells can bind to the
receptors on NK cells and T cells, leading to the apoptosis of
immune cells (Fig. 1A). HLA-G
expression in tumors such as hepatocellular carcinoma, glioma,
melanoma, renal, ovarian and lung carcinoma can also protect cancer
cells from NK and CTL-mediated destruction (99,100)
(Fig. 1B). In addition, plasmatic
sHLA-G molecules secreted by melanoma M8 cells functionally and
potently inhibit NK cell cytotoxicity by weakening the lytic
granules polarization toward the target cells (101). The formation of HLA-G and its
soluble counterpart, HLA-G5, can be increased by numerous
pro-inflammatory cytokines from the tumor microenvironment,
including interferon (IFN)-β and IFN-γ, which enhance the
protection of tumor cells from NK cell-mediated cytolysis (62). HLA-G can affect the equilibrium phase
by allowing cancer cell persistence, resulting in the selection of
tumor cells with reduced immunogenicity (102). Tumor development primarily occurs in
the evasion phase (103); in this
phase, tumor cells tend to express only HLA-G on the cell surface,
and not molecules essential to immune recognition, leading to the
rapid growth of tumor cells and the creation of a hypoxic
microenvironment. Furthermore, the hypoxic microenvironment and
cytokines derived by tumor-like transforming growth factor-β and
IL-10 in this phase can upregulate HLA-G expression (104). Estrogenic G-protein-coupled estrogen
receptor-1 signaling can trigger HLA-G expression by inhibiting
miR148a in breast cancer cells (105). These mechanisms modulated by HLA-G
are considered to facilitate the escape of tumor cells from the
antigen-specific immune response and to create a microenvironment
suitable for tumor cells (98).
Notably, HLA-G can be transferred from one
HLA-G-expressing cell to another, a phenomenon defined as
trogocytosis (106) (Fig. 1C). In this manner, cancer cells
transfer HLA-G-containing membrane patches to activated NK cells
(106). NK cells receiving the HLA-G
temporarily behave as regulatory suppressor cells, and acquire the
ability to inhibit immune responses such as prohibiting the
proliferation of immune cells and inhibiting the cytotoxic effector
of neighboring NK cells (106,107). T
cells and DCs may also possess regulatory properties subsequent to
acquiring HLA-G antigens from malignant plasma cells, providing
another novel mechanism for escaping effective immune surveillance
(108). Similarly, the
HLA-G-negative tumor cells in the vicinity of HLA-G-positive cells
can be protected in this way (1).
Therefore, this phenomenon may be an important mechanism underlying
immune inhibition in physical conditions as well as in pathological
conditions.
During tumor development, immune cells are able to
infiltrate tumors and participate in tumor formation and
progression (22,98). Tregs are one type of
tumor-infiltrating lymphocyte, which serve an important role in
immunologic escape of tumor cells. High tumor-infiltrating
Tregs are associated with the poor prognosis of human
malignant tumors (98).
Tregs can be induced by HLA-G not only under
physiological conditions but also pathological conditions (22). Several studies have elucidated the
association between HLA-G and Tregs in malignant
diseases (38,60,61). It
has been observed that the proportion of
CD4+CD25+FoxP3+ Tregs
was markedly increased in the plasma of breast cancer patients when
compared with that in the plasma of healthy controls, and the
increase was strongly associated with sHLA-G levels (38). In gastric cancer, a significant
positive association between HLA-G expression and the presence of
tumor-infiltrating Tregs was observed in tumor tissues
(60). When gastric cancer cells were
co-cultured with human peripheral blood mononuclear cells in
vitro, HLA-G overexpression in gastric cancer cells
significantly enhanced the number of Tregs (60). In addition, a negative correlation
between HLA-G expression and the number of CD8+ T
lymphocytes was observed in gastric tumor tissues using
immunohistochemistry (61). This may
be attributed to CD8+ T lymphocyte differentiation into
Tregs, induced by HLA-G (Fig.
1D). Taken together, these results indicate that HLA-G may be
associated with tumor progression and may be involved in tumor
evasion by inducing the production of Tregs.
Another mechanism by which sHLA-G promotes cancer
development is to inhibit the chemokine receptors expressed on
NK/T/B cells (Fig. 1E), which can
regulate the migration and recruitment of these cells by binding
with chemotactic cytokines (109).
Several studies have demonstrated that sHLA-G reduces the
expression of CCR2/CXCR3/CXCR5 on T cells, downregulates
CCR2/CXCR3/CX3CR1/CXCR5 expression on NK cells and inhibits
CXCR4/CXCR5 expression on B cells by interacting with ILT2
(109–111). Following the loss of chemotaxic
ability, NK/T/B cells cannot migrate to pathological tissues and
consequently cannot regulate a series of immunological responses
(109).
As aforementioned, HLA-G expression levels in tumor
tissue samples and high levels of sHLA-G in plasma samples, all
obtained from patients with various types of cancer, have
previously been detected, and high expression was determined to be
significantly associated with high histological grade, lymph node
metastasis, advanced clinical stage and a poor prognosis (96). Therefore, the application of HLA-G as
a diagnostic and prognostic biomarker of cancer has been
proposed.
Studies have demonstrated that HLA-G expression was
higher in malignant lesions compared with that in benign
hyperplasias (73,75). In thyroid tissues, the percentage of
cell staining for HLA-G antigen in PTC, follicular thyroid
carcinomas and follicular adenomas (FA) was significantly higher
than in colloid goiter and histologically normal thyroid glands
(75). In cervical cancer lesions,
HLA-G expression was increased in patients with invasive cervical
cancer compared with that in patients with CIN III, and HLA-G
expression was more frequently observed in cancer lesions from
patients with a higher FIGO stage of cancer (73). In liver tissues, HLA-G expression was
identified in the primary sites of HCC, but not detected in benign
lesions represented by liver cirrhosis (LC) (52). ELISA assay also showed that the plasma
sHLA-G level in patients with HCC was higher than in LC (52). These results indicated that HLA-G and
sHLA-G may be involved in tumorigenesis and tumor development,
which provides the basis for their purpose as indicators of early
diagnosis.
HLA-G expression may also be a prognostic predictor
of carcinoma following curative resection (50). For instance, high expression levels of
HLA-G in hepatocellular carcinoma are independently associated with
the shortening of overall survival times and an increase in tumor
recurrence (50). In addition,
patients with high HLA-G levels and a high
Tregs:CD8+ ratio exhibited ≥3 times increased
risk of tumor relapse and mortality, compared with those without
(50). Therefore, the combination of
HLA-G expression and the Tregs:CD8+ ratio
served as an improved prognosticator (50). In colorectal cancer, patients with
HLA-G expression in tissue have a significantly shorter survival
time compared with those patients with HLA-G negative, indicating
that HLA-G may be an independent prognostic factor for colorectal
cancer (55).
In addition, HLA-G expression in tissues and sHLA-G
concentrations in body fluids were useful for the prediction and
diagnosis of cancer patients with various subtypes (44,116).
When sHLA-G plasma level was detected in 120 patients with breast
cancer, Provatopoulou et al (44) found that sHLA-G levels were closely
associated with histological type: sHLA-G expression in patients
with mixed type co-existing ductal and lobular breast lesions was
significantly higher than patients with pure ductal carcinoma or
pure lobular neoplasia. Furthermore, the expression of HLA-G was
significantly higher in non-luminal subtypes of invasive ductal
breast carcinoma compared with in luminal subtypes (116).
Therefore, HLA-G/sHLA-G could be used as biomarkers
to diagnose cancer in the early stages, to predict the prognosis of
cancer patients and be used as subsidiary indicators to distinguish
various cancer subtypes of cancer. Additionally, as aforementioned,
HLA-G polymorphisms may be useful in the diagnosis of cancer.
In accordance with the important role of HLA-G in
promoting tumor evolution and progression, targeting HLA-G has been
deemed to be a novel innovative therapeutic strategy in cancer
(99,114). In human HCC cell lines, when HLA-G
expression is diminished by applying the vectors containing small
interfering RNA specifically targeting the HLA-G gene, a
significant increase in NK cell-mediated lysis occurs, which
prevents tumor progression (99). In
xenotumor mouse models, blocking HLA-G using a specific antibody
successfully inhibits the development of the tumor (114). Additional studies and clinical
trials are required to demonstrate the value of HLA-G in targeted
cancer therapy.
However, a noteworthy phenomenon to consider is that
several therapeutics may induce HLA-G-negative tumors to express
HLA-G, and thus contribute to cancer recurrence (34,81). For
example, a multicentric study has demonstrated that high expression
levels of HLA-G in glioblastoma may be induced by combined
5-aza-2′-deoxycytidine and interferon-γ treatments in vitro
(81). Melanoma patients undergoing
INF-α immunotherapy exhibited significantly enhanced increases in
the serum sHLA-G level (34).
Furthermore, it was demonstrated that HLA-G1 expression may
modulate the radiosensitivity of human tumoral cell lines: HLA-G1
expressing cells had a higher radiosensitivity in human melanoma M8
and human erythroleukemia K562 cell lines (117). Thus, identification of HLA-G status
may be conducive to improved selection of cancer patients who could
benefit from more tailored immunological therapy or neoadjuvant
biological therapy.
In conclusion, HLA-G is a potent immune-inhibitory
molecule in healthy individuals and in pathological organisms.
Although the frequency of HLA-G/sHLA-G expression and its
association with clinical parameters varies between different
cancer types, and even between different studies of the same tumor
type, HLA-G expression in tumors has been considered to be
detrimental. However, studies with a larger number of populations
from different ethnicities are required.
Considering the ectopic expression of HLA-G and its
gene polymorphisms in tumors, it is promising that membrane-bound
and sHLA-G could be a potential diagnostic biomarker to identify
tumors and to monitor disease stage. Since HLA-G has been
demonstrated to be an important molecule in tumor immune escape and
cancer development, blockade of HLA-G expression or elimination of
HLA-G-expressing cancer cells may be important to the efficacy of
anticancer therapies. Thus far, several molecular inhibitors have
demonstrated their ability to specifically target the HLA-G gene;
however, additional studies involving higher animals and clinical
trials involving humans are required in order for these inhibitors
targeting HLA-G to be used clinically.
The present study was financially supported by the
National Natural Science Foundation of China (grant no.
81372334).
|
1
|
Carosella ED, Favier B, Rouas-Freiss N,
Moreau P and Lemaoult J: Beyond the increasing complexity of the
immunomodulatory HLA-G molecule. Blood. 111:4862–4870. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Amiot L, Ferrone S, Grosse-Wilde H and
Seliger B: Biology of HLA-G in cancer: A candidate molecule for
therapeutic intervention? Cell Mol Life Sci. 68:417–431. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jiang F, Zhao H, Wang L, Guo X, Wang X,
Yin G, Hu Y, Li Y and Yao Y: Role of HLA-G1 in trophoblast cell
proliferation, adhesion and invasion. Biochem Biophys Res Commun.
458:154–160. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rouas-Freiss N, Kirszenbaum M, Dausset J
and Carosella ED: Fetomaternal tolerance: Role of HLA-G molecule in
the protection of the fetus against maternal natural killer
activity. C R Acad Sci III. 320:385–392. 1997.(In French).
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Paul P, Rouas-Freiss N, Khalil-Daher I,
Moreau P, Riteau B, Le Gal FA, Avril MF, Dausset J, Guillet JG and
Carosella ED: HLA-G expression in melanoma: A way for tumor cells
to escape from immunosurveillance. Proc Natl Acad Sci USA. 95:pp.
4510–4515. 1998; View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Castelli EC, Mendes-Junior CT,
Veiga-Castelli LC, Roger M, Moreau P and Donadi EA: A comprehensive
study of polymorphic sites along the HLA-G gene: Implication for
gene regulation and evolution. Mol Biol Evol. 28:3069–3086. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rousseau P, Le Discorde M, Mouillot G,
Marcou C, Carosella ED and Moreau P: The 14 bp deletion-insertion
polymorphism in the 3′ UT region of the HLA-G gene influences HLA-G
mRNA stability. Hum Immunol. 64:1005–1010. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Castelli EC, Mendes-Junior CT, Deghaide
NH, de Albuquerque RS, Muniz YC, Simões RT, Carosella ED, Moreau P
and Donadi EA: The genetic structure of 3′untranslated region of
the HLA-G gene: Polymorphisms and haplotypes. Genes Immun.
11:134–141. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hylenius S, Andersen AM, Melbye M and
Hviid TV: Association between HLA-G genotype and risk of
pre-eclampsia: A case-control study using family triads. Mol Hum
Reprod. 10:237–246. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pandey MK, Rani R and Agrawal S: An update
in recurrent spontaneous abortion. Arch Gynecol Obstet. 272:95–108.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tahan F and Patiroglu T: Plasma soluble
human leukocyte antigen G levels in asthmatic children. Int Arch
Allergy Immunol. 141:213–216. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Consiglio CR, Veit TD, Monticielo OA,
Mucenic T, Xavier RM, Brenol JC and Chies JA: Association of the
HLA-G gene +3142C>G polymorphism with systemic lupus
erythematosus. Tissue Antigens. 77:540–545. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gazit E, Slomov Y, Goldberg I, Brenner S
and Loewenthal R: HLA-G is associated with pemphigus vulgaris in
Jewish patients. Hum Immunol. 65:39–46. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dias FC, Castelli EC, Collares CV, Moreau
P and Donadi EA: The Role of HLA-G molecule and HLA-G gene
polymorphisms in tumors, viral hepatitis, and parasitic diseases.
Front Immunol. 6:92015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Koller BH, Geraghty DE, DeMars R, Duvick
L, Rich SS and Orr HT: Chromosomal organization of the human major
histocompatibility complex class I gene family. J Exp Med.
169:469–480. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Morandi F and Pistoia V: Interactions
between HLA-G and HLA-E in Physiological and Pathological
Conditions. Front Immunol. 5:3942014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Carosella ED, Rouas-Freiss N, Tronik-Le
Roux D, Moreau P and LeMaoult J: HLA-G: An immune checkpoint
molecule. Adv Immunol. 127:33–1444. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Carosella ED, Moreau P, Le Maoult J, Le
Discorde M, Dausset J and Rouas-Freiss N: HLA-G molecules: From
maternal-fetal tolerance to tissue acceptance. Adv Immunol.
81:199–252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fujii T, Ishitani A and Geraghty DE: A
soluble form of the HLA-G antigen is encoded by a messenger
ribonucleic acid containing intron 4. J Immunol. 153:5516–5524.
1994.PubMed/NCBI
|
|
20
|
Park GM, Lee S, Park B, Kim E, Shin J, Cho
K and Ahn K: Soluble HLA-G generated by proteolytic shedding
inhibits NK-mediated cell lysis. Biochem Biophys Res Commun.
313:606–611. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ishitani A and Geraghty DE: Alternative
splicing of HLA-G transcripts yields proteins with primary
structures resembling both class I and class II antigens. Proc Natl
Acad Sci USA. 89:pp. 3947–3951. 1992; View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Curigliano G, Criscitiello C, Gelao L and
Goldhirsch A: Molecular pathways: Human leukocyte antigen G
(HLA-G). Clin Cancer Res. 19:5564–5571. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gao GF, Willcox BE, Wyer JR, Boulter JM,
O'Callaghan CA, Maenaka K, Stuart DI, Jones EY, van der Merwe PA,
Bell JI and Jakobsen BK: Classical and nonclassical class I major
histocompatibility complex molecules exhibit subtle conformational
differences that affect binding to CD8alphaalpha. J Biol Chem.
275:15232–15238. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Colonna M, Samaridis J, Cella M, Angman L,
Allen RL, O'Callaghan CA, Dunbar R, Ogg GS, Cerundolo V and Rolink
A: Human myelomonocytic cells express an inhibitory receptor for
classical and nonclassical MHC class I molecules. J Immunol.
160:3096–3100. 1998.PubMed/NCBI
|
|
25
|
Borges L and Cosman D: LIRs/ILTs/MIRs,
inhibitory and stimulatory Ig-superfamily receptors expressed in
myeloid and lymphoid cells. Cytokine Growth Factor Rev. 11:209–217.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yan WH and Fan LA: Residues Met76 and
Gln79 in HLA-G alpha1 domain involve in KIR2DL4 recognition. Cell
Res. 15:176–182. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
González A, Rebmann V, LeMaoult J, Horn
PA, Carosella ED and Alegre E: The immunosuppressive molecule HLA-G
and its clinical implications. Crit Rev Clin Lab Sci. 49:63–84.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liang S, Ristich V, Arase H, Dausset J,
Carosella ED and Horuzsko A: Modulation of dendritic cell
differentiation by HLA-G and ILT4 requires the IL-6-STAT3 signaling
pathway. Proc Natl Acad Sci USA. 105:pp. 8357–8362. 2008;
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ketroussi F, Giuliani M, Bahri R, Azzarone
B, Charpentier B and Durrbach A: Lymphocyte cell-cycle inhibition
by HLA-G is mediated by phosphatase SHP-2 and acts on the mTOR
pathway. PLoS One. 6:e227762011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
LeMaoult J, Zafaranloo K, Le Danff C and
Carosella ED: HLA-G up-regulates ILT2, ILT3, ILT4, and KIR2DL4 in
antigen presenting cells, NK, cells and T cells. FASEB J.
19:662–664. 2005.PubMed/NCBI
|
|
31
|
Gooden MJ and van Hall T: Infiltrating
CTLs are bothered by HLA-E on tumors. Oncoimmunology. 1:92–93.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
LeMaoult J, Krawice-Radanne I, Dausset J
and Carosella ED: HLA-G1-expressing antigen-presenting cells induce
immunosuppressive CD4+ T cells. Proc Natl Acad Sci USA. 101:pp.
7064–7069. 2004; View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ibrahim EC, Aractingi S, Allory Y, Borrini
F, Dupuy A, Duvillard P, Carosella ED, Avril MF and Paul P:
Analysis of HLA antigen expression in benign and malignant
melanocytic lesions reveals that upregulation of HLA-G expression
correlates with malignant transformation, high inflammatory
infiltration and HLA-A1 genotype. Int J Cancer. 108:243–250. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ugurel S, Rebmann V, Ferrone S, Tilgen W,
Grosse-Wilde H and Reinhold U: Soluble human leukocyte antigen-G
serum level is elevated in melanoma patients and is further
increased by interferon-alpha immunotherapy. Cancer. 92:369–376.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
da Silva GB, Silva TG, Duarte RA, Neto NL,
Carrara HH, Donadi EA, Gonçalves MA, Soares EG and Soares CP:
Expression of the Classical and Nonclassical HLA molecules in
breast cancer. Int J Breast Cancer. 2013:2504352013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Elliott RL, Jiang XP, Phillips JT, Barnett
BG and Head JF: Human leukocyte antigen G expression in breast
cancer: Role in immunosuppression. Cancer Biother Radiopharm.
26:153–157. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lefebvre S, Antoine M, Uzan S, McMaster M,
Dausset J, Carosella ED and Paul P: Specific activation of the
non-classical class I histocompatibility HLA-G antigen and
expression of the ILT2 inhibitory receptor in human breast cancer.
J Pathol. 196:266–274. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen HX, Lin A, Shen CJ, Zhen R, Chen BG,
Zhang X, Cao FL, Zhang JG and Yan WH: Upregulation of human
leukocyte antigen-G expression and its clinical significance in
ductal breast cancer. Hum Immunol. 71:892–898. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
He X, Dong DD, Yie SM, Yang H, Cao M, Ye
SR, Li K, Liu J and Chen J: HLA-G expression in human breast
cancer: Implications for diagnosis and prognosis, and effect on
allocytotoxic lymphocyte response after hormone treatment in vitro.
Ann Surg Oncol. 17:1459–1469. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
de Kruijf EM, Sajet A, van Nes JG, Natanov
R, Putter H, Smit VT, Liefers GJ, van den Elsen PJ, van de Velde CJ
and Kuppen PJ: HLA-E and HLA-G expression in classical HLA class
I-negative tumors is of prognostic value for clinical outcome of
early breast cancer patients. J Immunol. 185:7452–7459. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ramos CS, Gonçalves AS, Marinho LC, Gomes
Avelino MA, Saddi VA, Lopes AC, Simões RT and Wastowski IJ:
Analysis of HLA-G gene polymorphism and protein expression in
invasive breast ductal carcinoma. Hum Immunol. 75:667–672. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sayed D, Badr G, Maximous D, Mikhail NN,
Abu-Tarboush F and Alhazza IM: HLA-G and its relation to
proliferation index in detection and monitoring breast cancer
patients. Tissue Antigens. 75:40–47. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jeong S, Park S, Park BW, Park Y, Kwon OJ
and Kim HS: Human leukocyte antigen-G (HLA-G) polymorphism and
expression in breast cancer patients. PLoS One. 9:e982842014.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Provatopoulou X, Kalogera E, Sagkriotis A,
Zagouri F, Nonni A, Zografos GC and Gounaris A: Soluble human
leukocyte antigen-G expression in patients with ductal and lobular
breast malignancy. Anticancer Res. 32:1021–1026. 2012.PubMed/NCBI
|
|
45
|
Urosevic M, Kurrer MO, Kamarashev J,
Mueller B, Weder W, Burg G, Stahel RA, Dummer R and Trojan A: Human
leukocyte antigen G up-regulation in lung cancer associates with
high-grade histology, human leukocyte antigen class I loss and
interleukin-10 production. Am J Pathol. 159:817–824. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yie SM, Yang H, Ye SR, Li K, Dong DD and
Lin XM: Expression of human leucocyte antigen G (HLA-G) is
associated with prognosis in non-small cell lung cancer. Lung
Cancer. 58:267–274. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lin A, Zhu CC, Chen HX, Chen BF, Zhang X,
Zhang JG, Wang Q, Zhou WJ, Hu W, Yang HH, et al: Clinical relevance
and functional implications for human leucocyte antigen-g
expression in non-small-cell lung cancer. J Cell Mol Med.
14:2318–2329. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Schütt P, Schütt B, Switala M, Bauer S,
Stamatis G, Opalka B, Eberhardt W, Schuler M, Horn PA and Rebmann
V: Prognostic relevance of soluble human leukocyte antigen-G and
total human leukocyte antigen class I molecules in lung cancer
patients. Hum Immunol. 71:489–495. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cao M, Yie SM, Liu J, Ye SR, Xia D and Gao
E: Plasma soluble HLA-G is a potential biomarker for diagnosis of
colorectal, gastric, esophageal and lung cancer. Tissue Antigens.
78:120–128. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cai MY, Xu YF, Qiu SJ, Ju MJ, Gao Q, Li
YW, Zhang BH, Zhou J and Fan J: Human leukocyte antigen-G protein
expression is an unfavorable prognostic predictor of hepatocellular
carcinoma following curative resection. Clin Cancer Res.
15:4686–4693. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lin A, Chen HX, Zhu CC, Zhang X, Xu HH,
Zhang JG, Wang Q, Zhou WJ and Yan WH: Aberrant human leucocyte
antigen-G expression and its clinical relevance in hepatocellular
carcinoma. J Cell Mol Med. 14:2162–2171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang Y, Ye Z, Meng XQ and Zheng SS:
Expression of HLA-G in patients with hepatocellular carcinoma.
Hepatobiliary Pancreat Dis Int. 10:158–163. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Park Y, Park Y, Lim HS, Kim YS, Hong DJ
and Kim HS: Soluble human leukocyte antigen-G expression in
hepatitis B virus infection and hepatocellular carcinoma. Tissue
Antigens. 79:97–103. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Fukushima Y, Oshika Y, Nakamura M,
Tokunaga T, Hatanaka H, Abe Y, Yamazaki H, Kijima H, Ueyama Y and
Tamaoki N: Increased expression of human histocompatibility
leukocyte antigen-G in colorectal cancer cells. Int J Mol Med.
2:349–351. 1998.PubMed/NCBI
|
|
55
|
Ye SR, Yang H, Li K, Dong DD, Lin XM and
Yie SM: Human leukocyte antigen G expression: As a significant
prognostic indicator for patients with colorectal cancer. Mod
Pathol. 20:375–383. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zeestraten EC, Reimers MS, Saadatmand S,
Goossens-Beumer IJ, Dekker JW, Liefers GJ, van den Elsen PJ, van de
Velde CJ and Kuppen PJ: Combined analysis of HLA class I, HLA-E and
HLA-G predicts prognosis in colon cancer patients. Br J Cancer.
110:459–468. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Guo ZY, Lv YG, Wang L, Shi SJ, Yang F,
Zheng GX, Wen WH and Yang AG: Predictive value of HLA-G and HLA-E
in the prognosis of colorectal cancer patients. Cell Immunol.
293:10–16. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhu CB, Wang CX, Zhang X, Zhang J and Li
W: Serum sHLA-G levels: A useful indicator in distinguishing
colorectal cancer from benign colorectal diseases. Int J Cancer.
128:617–622. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yie SM, Yang H, Ye SR, Li K, Dong DD and
Lin XM: Expression of human leukocyte antigen G (HLA-G) correlates
with poor prognosis in gastric carcinoma. Ann Surg Oncol.
14:2721–2729. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Du L, Xiao X, Wang C, Zhang X, Zheng N,
Wang L, Zhang X, Li W, Wang S and Dong Z: Human leukocyte antigen-G
is closely associated with tumor immune escape in gastric cancer by
increasing local regulatory T cells. Cancer Sci. 102:1272–1280.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Tuncel T, Karagoz B, Haholu A, Ozgun A,
Emirzeoglu L, Bilgi O and Kandemir EG: Immunoregulatory function of
HLA-G in gastric cancer. Asian Pac J Cancer Prev. 14:7681–7684.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yie SM, Yang H, Ye SR, Li K, Dong DD and
Lin XM: Expression of HLA-G is associated with prognosis in
esophageal squamous cell carcinoma. Am J Clin Pathol.
128:1002–1009. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Lin A, Zhang X, Zhou WJ, Ruan YY, Xu DP,
Wang Q and Yan WH: Human leukocyte antigen-G expression is
associated with a poor prognosis in patients with esophageal
squamous cell carcinoma. Int J Cancer. 129:1382–1390. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hu J, Li L, Liu Y, Chen Y, Liu C, Liang W,
Zhao J, Zou H, Cui X, Qi Y, et al: Overexpression of HLA-G Is
positively associated with Kazakh esophageal squamous cell
carcinoma in Xinjiang, China. Viral Immunol. 26:180–184. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zheng J, Xu C, Chu D, Zhang X, Li J, Ji G,
Hong L, Feng Q, Li X, Wu G, et al: Human leukocyte antigen G is
associated with esophageal squamous cell carcinoma progression and
poor prognosis. Immunol Lett. 161:13–19. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Cai MB, Han HQ, Bei JX, Liu CC, Lei JJ,
Cui Q, Feng QS, Wang HY, Zhang JX, Liang Y, et al: Expression of
human leukocyte antigen G is associated with prognosis in
nasopharyngeal carcinoma. Int J Biol Sci. 8:891–900. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Silva TG, Crispim JC, Miranda FA, Hassumi
MK, de Mello JM, Simões RT, Souto F, Soares EG, Donadi EA and
Soares CP: Expression of the nonclassical HLA-G and HLA-E molecules
in laryngeal lesions as biomarkers of tumor invasiveness. Histol
Histopathol. 26:1487–1497. 2011.PubMed/NCBI
|
|
68
|
Gan LH, Huang LF, Zhang X, Lin A, Xu DP,
Wang Q, Wang TJ and Yan WH: Tumor-specific upregulation of human
leukocyte antigen-G expression in bladder transitional cell
carcinoma. Hum Immunol. 71:899–904. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Ibrahim EC, Guerra N, Lacombe MJ, Angevin
E, Chouaib S, Carosella ED, Caignard A and Paul P: Tumor-specific
up-regulation of the nonclassical class I HLA-G antigen expression
in renal carcinoma. Cancer Res. 61:6838–6845. 2001.PubMed/NCBI
|
|
70
|
Kren L, Valkovsky I, Dolezel J, Capak I,
Pacik D, Poprach A, Lakomy R, Redova M, Fabian P, Krenova Z and
Slaby O: HLA-G and HLA-E specific mRNAs connote opposite prognostic
significance in renal cell carcinoma. Diagn Pathol. 7:582012.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Ibrahim EC, Allory Y, Commo F, Gattegno B,
Callard P and Paul P: Altered pattern of major histocompatibility
complex expression in renal carcinoma: Tumor-specific expression of
the nonclassical human leukocyte antigen-G molecule is restricted
to clear cell carcinoma while up-regulation of other major
histocompatibility complex antigens is primarily distributed in all
subtypes of renal carcinoma. Am J Pathol. 162:501–508. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Li BL, Lin A, Zhang XJ, Zhang X, Zhang JG,
Wang Q, Zhou WJ, Chen HX, Wang TJ and Yan WH: Characterization of
HLA-G expression in renal cell carcinoma. Tissue Antigens.
74:213–221. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Li XJ, Zhang X, Lin A, Ruan YY and Yan WH:
Human leukocyte antigen-G (HLA-G) expression in cervical cancer
lesions is associated with disease progression. Hum Immunol.
73:946–949. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Nunes LM, Ayres FM, Francescantonio IC,
Saddi VA, Avelino MA, Alencar Rde C, Silva RC, Meneghini AJ and
Wastowski IJ: Association between the HLA-G molecule and lymph node
metastasis in papillary thyroid cancer. Hum Immunol. 74:447–451.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
de Figueiredo Feitosa NL, Crispim JC,
Zanetti BR, Magalhães PK, Soares CP, Soares EG, Neder L, Donadi EA
and Maciel LM: HLA-G is differentially expressed in thyroid
tissues. Thyroid. 24:585–592. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Morandi F, Levreri I, Bocca P, Galleni B,
Raffaghello L, Ferrone S, Prigione I and Pistoia V: Human
neuroblastoma cells trigger an immunosuppressive program in
monocytes by stimulating soluble HLA-G release. Cancer Res.
67:6433–6441. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Morandi F, Scaruffi P, Gallo F, Stigliani
S, Moretti S, Bonassi S, Gambini C, Mazzocco K, Fardin P, Haupt R,
et al: Bone marrow-infiltrating human neuroblastoma cells express
high levels of calprotectin and HLA-G proteins. PLoS One.
7:e299222012. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Wiendl H, Mitsdoerffer M, Hofmeister V,
Wischhusen J, Bornemann A, Meyermann R, Weiss EH, Melms A and
Weller M: A functional role of HLA-G expression in human gliomas:
An alternative strategy of immune escape. J Immunol. 168:4772–4780.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Kren L, Muckova K, Lzicarova E, Sova M,
Vybihal V, Svoboda T, Fadrus P, Smrcka M, Slaby O, Lakomy R, et al:
Production of immune-modulatory nonclassical molecules HLA-G and
HLA-E by tumor infiltrating ameboid microglia/macrophages in
glioblastomas: A role in innate immunity? J Neuroimmunol.
220:131–135. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Kren L, Slaby O, Muckova K, Lzicarova E,
Sova M, Vybihal V, Svoboda T, Fadrus P, Lakomy R, Vanhara P, et al:
Expression of immune-modulatory molecules HLA-G and HLA-E by tumor
cells in glioblastomas: An unexpected prognostic significance?
Neuropathology. 31:129–134. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wastowski IJ, Simões RT, Yaghi L, Donadi
EA, Pancoto JT, Poras I, Lechapt-Zalcman E, Bernaudin M, Valable S,
Carlotti CG Jr, et al: Human leukocyte antigen-G is frequently
expressed in glioblastoma and may be induced in vitro by combined
5-aza-2′-deoxycytidine and interferon-γ treatments: Results from a
multicentric study. Am J Pathol. 182:540–552. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Gros F, Sebti Y, de Guibert S, Branger B,
Bernard M, Fauchet R and Amiot L: Soluble HLA-G molecules increase
during acute leukemia, especially in subtypes affecting monocytic
and lymphoid lineages. Neoplasia. 8:223–230. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Dardano A, Rizzo R, Polini A, Stignani M,
Tognini S, Pasqualetti G, Ursino S, Colato C, Ferdeghini M,
Baricordi OR and Monzani F: Soluble human leukocyte antigen-g and
its insertion/deletion polymorphism in papillary thyroid carcinoma:
Novel potential biomarkers of disease? J Clin Endocrinol Metab.
97:4080–4086. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Reimers MS, Engels CC, Putter H, Morreau
H, Liefers GJ, van de Velde CJ and Kuppen PJ; Prognostic value of
HLA class I, HLA-E, HLA-G and Tregs in rectal cancer, : A
retrospective cohort study. Bmc Cancer. 14:4862014. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Martelli-Palomino G, Pancotto JA, Muniz
YC, Mendes-Junior CT, Castelli EC, Massaro JD, Krawice-Radanne I,
Poras I, Rebmann V, Carosella ED, et al: Polymorphic sites at the
3′ untranslated region of the HLA-G gene are associated with
differential hla-g soluble levels in the Brazilian and French
population. PLoS One. 8:e717422013. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Silva ID, Muniz YC, Sousa MC, Silva KR,
Castelli EC, Filho JC, Osta AP, Lima MI and Simões RT: HLA-G 3′UTR
polymorphisms in high grade and invasive cervico-vaginal cancer.
Hum Immunol. 74:452–458. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Jiang Y, Chen S, Jia S, Zhu Z, Gao X, Dong
D and Gao Y: Association of HLA-G 3′ UTR 14-bp insertion/deletion
polymorphism with hepatocellular carcinoma susceptibility in a
Chinese population. Dna Cell Biol. 30:1027–1032. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Lau DT, Norris MD, Marshall GM, Haber M
and Ashton LJ: HLA-G polymorphisms, genetic susceptibility, and
clinical outcome in childhood neuroblastoma. Tissue Antigens.
78:421–427. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Ferguson R, Ramanakumar AV, Koushik A,
Coutlée F, Franco E and Roger M; Biomarkers of Cervical Cancer Risk
Study Team, : Human leukocyte antigen G polymorphism is associated
with an increased risk of invasive cancer of the uterine cervix.
Int J Cancer. 131:E312–E319. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Ghandri N, Gabbouj S, Farhat K, Bouaouina
N, Abdelaziz H, Nouri A, Chouchane L and Hassen E: Association of
HLA-G polymorphisms with nasopharyngeal carcinoma risk and clinical
outcome. Hum Immunol. 72:150–158. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Xu HH, Shi WW, Lin A and Yan WH: HLA-G 3′
untranslated region polymorphisms influence the susceptibility for
human papillomavirus infection. Tissue Antigens. 84:216–222. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Eskandari-Nasab E, Hashemi M, Hasani SS,
Omrani M, Taheri M and Mashhadi MA: Association between HLA-G 3′UTR
14-bp ins/del polymorphism and susceptibility to breast cancer.
Cancer Biomark. 13:253–259. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Rolfsen GB, Castelli EC, Donadi EA, Duarte
RA and Soares CP: HLA-G polymorphism and breast cancer. Int J
Immunogenet. 41:143–148. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Teixeira AC, Mendes-Junior CT, Souza FF,
Marano LA, Deghaide NH, Ferreira SC, Mente ED, Sankarankutty AK,
Elias-Junior J, Castro-e-Silva O, et al: The 14bp-deletion allele
in the HLA-G gene confers susceptibility to the development of
hepatocellular carcinoma in the Brazilian population. Tissue
Antigens. 81:408–413. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Kim SK, Chung JH, Jeon JW, Park JJ, Cha
JM, Joo KR, Lee JI and Shin HP: Association between HLA-G 14-bp
insertion/deletion polymorphism and hepatocellular carcinoma in
Korean patients with chronic hepatitis B viral infection.
Hepatogastroenterology. 60:796–798. 2013.PubMed/NCBI
|
|
96
|
Yan WH: HLA-G expression in cancers:
Potential role in diagnosis, prognosis and therapy. Endocr Metab
Immune Disord Drug Targets. 11:76–89. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Teng MW, Galon J, Fridman WH and Smyth MJ:
From mice to humans: Developments in cancer immunoediting. J Clin
Invest. 125:3338–3346. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Kochan G, Escors D, Breckpot K and
Guerrero-Setas D: Role of non-classical MHC class I molecules in
cancer immunosuppression. Oncoimmunology. 2:e264912013. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Zeng XC, Zhang T, Huang DH, Wang GY, Chen
W, Li H, Zhang J, Fang TL, Zhang Q and Chen GH: RNA interfering
targeting human leukocyte antigen-G enhanced immune surveillance
mediated by the natural killer cells on hepatocellular carcinoma.
Ann Clin Lab Sci. 43:135–144. 2013.PubMed/NCBI
|
|
100
|
Lin A, Xu HH, Xu DP, Zhang X, Wang Q and
Yan WH: Multiple steps of HLA-G in ovarian carcinoma metastasis:
Alter NK cytotoxicity and induce matrix metalloproteinase-15
(MMP-15) expression. Hum Immunol. 74:439–446. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Lesport E, Baudhuin J, LeMaoult J, Sousa
S, Doliger C, Carosella ED and Favier B: Human melanoma cell
secreting human leukocyte antigen-G5 inhibit natural killer cell
cytotoxicity by impairing lytic granules polarization toward target
cell. Hum Immunol. 70:1000–1005. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Moreau P, Mouillot G, Rousseau P, Marcou
C, Dausset J and Carosella ED: HLA-G gene repression is reversed by
demethylation. Proc Natl Acad Sci USA. 100:pp. 1191–1196. 2003;
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Zidi I, Guillard C, Marcou C,
Krawice-Radanne I, Sangrouber D, Rouas-Freiss N, Carosella ED and
Moreau P: Increase in HLA-G1 proteolytic shedding by tumor cells: A
regulatory pathway controlled by NF-kappaB inducers. Cell Mol Life
Sci. 63:2669–2681. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Rodríguez JA, Galeano L, Palacios DM,
Gómez C, Serrano ML, Bravo MM and Combita AL: Altered HLA class I
and HLA-G expression is associated with IL-10 expression in
patients with cervical cancer. Pathobiology. 79:72–83. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Tao S, He H, Chen Q and Yue W: GPER
mediated estradiol reduces miR-148a to promote HLA-G expression in
breast cancer. Biochem Biophys Res Commun. 451:74–78. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Caumartin J, Favier B, Daouya M, Guillard
C, Moreau P, Carosella ED and LeMaoult J: Trogocytosis-based
generation of suppressive NK cells. Embo J. 26:1423–1433. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
LeMaoult J, Caumartin J, Daouya M, Favier
B, Le Rond S, Gonzalez A and Carosella ED: Immune regulation by
pretenders: Cell-to-cell transfers of HLA-G make effector T cells
act as regulatory cells. Blood. 109:2040–2048. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Brown R, Kabani K, Favaloro J, Yang S, Ho
PJ, Gibson J, Fromm P, Suen H, Woodland N, Nassif N, et al: CD86+
or HLA-G+ can be transferred via trogocytosis from myeloma cells to
T cells and are associated with poor prognosis. Blood.
120:2055–2063. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Morandi F, Ferretti E, Bocca P, Prigione
I, Raffaghello L and Pistoia V: A novel mechanism of soluble HLA-G
mediated immune modulation: Downregulation of T cell chemokine
receptor expression and impairment of chemotaxis. PLoS One.
5:e117632010. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Morandi F, Ferretti E, Castriconi R,
Dondero A, Petretto A, Bottino C and Pistoia V: Soluble HLA-G
dampens CD94/NKG2A expression and function and differentially
modulates chemotaxis and cytokine and chemokine secretion in
CD56bright and CD56dim NK cells. Blood. 118:5840–5850. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Naji A, Menier C, Morandi F, Agaugué S,
Maki G, Ferretti E, Bruel S, Pistoia V, Carosella ED and
Rouas-Freiss N: Binding of HLA-G to ITIM-bearing Ig-like transcript
2 receptor suppresses B cell responses. J Immunol. 192:1536–1546.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Liu X, Gu W and Li X: HLA-G regulates the
invasive properties of JEG-3 choriocarcinoma cells by controlling
STAT3 activation. Placenta. 34:1044–1052. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Liang S, Baibakov B and Horuzsko A: HLA-G
inhibits the functions of murine dendritic cells via the PIR-B
immune inhibitory receptor. Eur J Immunol. 32:2418–2426. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Agaugué S, Carosella ED and Rouas-Freiss
N: Role of HLA-G in tumor escape through expansion of
myeloid-derived suppressor cells and cytokinic balance in favor of
Th2 versus Th1/Th17. Blood. 117:7021–7031. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Loumagne L, Baudhuin J, Favier B,
Montespan F, Carosella ED and Rouas-Freiss N: In vivo evidence that
secretion of HLA-G by immunogenic tumor cells allows their evasion
from immunosurveillance. Int J Cancer. 135:2107–2117. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Dong DD, Yie SM, Li K, Li F, Xu Y, Xu G,
Song L and Yang H: Importance of HLA-G expression and tumor
infiltrating lymphocytes in molecular subtypes of breast cancer.
Hum Immunol. 73:998–1004. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Gallegos CE, Michelin S, Trasci SB, Lobos
EA, Dubner D and Carosella ED: HLA-G1 increases the
radiosensitivity of human tumoral cells. Cell Immunol. 287:106–111.
2014. View Article : Google Scholar : PubMed/NCBI
|