Introduction
Breast cancer accounts for 25% of all cancer cases
in women and is the leading cause of cancer-associated mortality
worldwide, contributing to 15% of all cancer-associated mortalities
(1). The interaction between tumor
cells and their surrounding microenvironment, composed of the
stroma, blood vessels, infiltrating immune cells and host of
associated tissue cells, serves a pivotal function in influencing
tumor establishment and progression. Numerous studies have
recognized the critical function of immune cells, particularly
tumor-associated macrophages (TAMs), in promoting primary tumor
growth, metastatic progression, a poor overall survival and
therapeutic resistance (2–6). TAMs are derived from circulating
monocytes or resident tissue macrophages and are usually found
within or in close proximity to tumor masses (7,8). TAMs are
recruited to tumors by macrophage-attracting chemokines and
cytokines, which are usually secreted by tumor cells (9,10).
Following infiltration, TAMs secrete a host of growth factors,
including epidermal growth factor (EGF) and transforming growth
factor (TGF)-β, as well as pro-angiogenic factors, including
vascular endothelial growth factor (VEGF)A, platelet-derived growth
factor and fibroblast growth factor, that work to modulate tumor
growth (11,12).
The Cbp/p300-interacting transactivator with
Glu/Asp-rich carboxy-terminal domain-2 (CITED2) is a non-DNA
binding transcriptional co-regulator and modulates gene
transcription by directly or indirectly interacting with
transcription factors and co-factors, thereby influencing their
ability to promote or repress gene activation (13–18).
During embryonic development, CITED2 serves a critical function in
the development of the liver, lungs, heart and neural tube, as
deleting CITED2 is associated with embryonic lethality
(19–21). Furthermore, it has been reported that
CITED2 serves a role in promoting skin, colon, lung and breast
cancer (22–27). Beyond potentially facilitating breast
cancer metastasis (25), it has been
suggested that CITED2 may promote primary tumor growth (24). This effect may be mediated, at least
partly, by modulating tumor vasculature via regulation of VEGFA;
however, the mechanisms by which CITED2 regulates tumor growth
remain to be fully elucidated.
Using two murine xenograft models of human breast
cancer, one in which tumor growth was sensitive to CITED2 silencing
(MDA-MB-231) and one that was insensitive (MDA-MB-468), the present
study demonstrated that the reduced primary tumor growth observed
in response to silencing CITED2 is associated with the attenuation
of macrophage recruitment, as determined by in vivo and
in vitro assays. Following CITED2 silencing, expression of
the macrophage chemoattractant C-C motif chemokine ligand (CCL)20
was decreased. Correspondingly, the present study provided evidence
that CITED2 localizes to the CCL20 promoter. Finally, supporting a
function for this chemokine, neutralizing CCL20 in tumor
cell-conditioned media inhibited macrophage recruitment, as
determined by a Transwell migration assay. Collectively, these
results suggest that CITED2 modulates macrophage recruitment in
breast tumors, potentially via the regulation of tumor
cell-secreted CCL20.
Materials and methods
Cell lines and transfection
The human breast cancer cell lines, MDA-MB-231 and
MDA-MB-468, were obtained from the American Type Culture Collection
(ATCC; Manassas, VA, USA) and were cultured as previously described
(25). These cell lines were
authenticated by the ATCC using DNA profiling and cytogenetic
analysis and used for experiments within 6 months from the time of
resuscitation. Scramble and short hairpin (sh)CITED2-expressing
MDA-MB-231 and MDA-MB-468 cells were generated following a
previously described protocol (24,25), using
the lentiviral shRNA expression vector pLKO.1-puro (Addgene plasmid
8453; Addgene, Inc., Cambridge, MA, USA) containing a shRNA
sequence specific for scramble or CITED2, as previously described
(15,28).
Macrophages were obtained from murine bone marrow.
Tibiae and femora from one 8–12-week-old C57BL/6 healthy female
mouse (Jackson Laboratory, Bar Harbor, ME, USA) were aseptically
dissected in cold PBS (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 2% penicillin/streptomycin (Gibco;
Thermo Fisher Scientific, Inc.). The bone marrow was flushed with
α-minimum essential medium (α-MEM; Gibco; Thermo Fisher Scientific,
Inc.) containing 10% fetal bovine serum (FBS; Atlanta Biologicals,
Flowery Branch, GA, USA) and 1% penicillin/streptomycin (Gibco;
Thermo Fisher Scientific, Inc.), centrifuged at 500 × g for 5 min
at 4°C and re-suspended in fresh media. Cells were permitted to
attach in a 100-mm culture dish overnight at 37°C in a 5%
CO2 incubator with 5 ng/ml macrophage colony stimulating
factor (M-CSF; Promega Corporation, Madison, WI, USA). Non-adherent
marrow cells were collected and strained through a 70-µm nylon mesh
cell strainer (BD Biosciences, Franklin Lakes, NJ, USA) and
cultured (6×106 cells) in a 100-mm petri dish for 72 h
in the presence of 50 ng/ml M-CSF. Cells were detached using 4 ml
cold Versene (Invitrogen; Thermo Fisher Scientific, Inc.) for 10
min, resuspended in fresh α-MEM, 4 fields of view were counted
under a light microscope using a hematocytometer at ×40
magnification and the desired cell number was plated for in
vitro Transwell migration assays.
Orthotopic xenograft models of human
breast cancer
The orthotopic xenograft tumor models used in the
present study have been previously described (24). Briefly, 1.5×106 scramble-
or shCITED2-expressing tumor cells (n=5 per group) in 0.1 ml of
Hanks' balanced salt solution (Gibco; Thermo Fisher Scientific,
Inc.) were injected once bilaterally into the third mammary fat pad
of five-week-old athymic nude female mice (Taconic, Hudson, NY,
USA). Mice were anesthetized by intraperitoneal injection of 0.25
ml ketamine hydrochloride (100 mg/ml; Hospira; Pfizer, Inc., New
York, NY, USA) prepared in xylazine solution (2 mg/ml; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) prior to tumor inoculation.
With the onset of tumor growth, tumor volume was measured
periodically as described previously (24). Once the tumor volume in the scramble
group reached the maximum permitted size of 2 cm3 in any
dimension, animals from both the scramble and shCITED2 group were
euthanized and the tumor tissues were surgically isolated from the
orthotopic site. All animal experiments were conducted in
accordance with the National Research Council's ‘Guide to the Care
and Use of Laboratory Animals’ (29).
Mice of mean weight equaling 15–17 g were housed in ventilated
racks supplied with HEPA filtered, tempered (~70°F) and humidified
(~50%) air and exhausted direct to the outside through the
interstitial space above. Mice were provided 24 h access to food
and water. Cages were supplied with reverse osmosis filtered
hyperchlorinated water via an in-cage automatic watering system.
Light is controlled by central timer providing a 14 h light/10 h
dark cycle. The Johns Hopkins Animal Care and Use Committee
approved the use of animals in the present study (animal welfare
assurance no. A3272-01, protocol no. MO16M362).
Immunohistochemistry
Tumor tissues surgically removed from orthotopic
sites of euthanized mice were fixed in 10% formalin (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) for 24 h at room temperature,
paraffin-embedded and cut into 10-mm sections. For
immunohistochemical analysis, these tissues sections were
deparaffinized in xylene (Thermo Fisher Scientific, Inc.),
rehydrated using a graded series of ethanol (Pharmco-AAPER;
Greenfield Global, Shelbyville, KT, USA) and washed in PBS (Gibco;
Thermo Fisher Scientific, Inc.). Sections were immersed in antigen
retrieval solution (Dako; Agilent Technologies, Inc., Santa Clara,
CA, USA) and heated at 95°C in a steamer for 20 min. Cooled
sections were washed with PBS and endogenous peroxidase activity
was quenched by immersion in 3% hydrogen peroxide (Thermo Fisher
Scientific, Inc.) for 12 min. Sections were blocked with
protein-blocking solution (Dako; Agilent Technologies, Inc.) for 20
min at room temperature. Subsequently, sections were incubated with
rat anti-mouse F4/80 antibody (1:300; cat. no. MCA497RT; Bio-Rad
Laboratories, Inc., Hercules, CA, USA) at room temperature for 1 h.
PBS-washed sections were sequentially incubated for 30 min at room
temperature with an avidin-bound rabbit anti-rat IgG antibody (cat.
no. BA-4000) and streptavidin-horseradish peroxidase (Vector
Laboratories, Inc., Burlingame, CA, USA). Proteins were visualized
by adding the chromogen 3,3-diaminobenzamindine (GE Healthcare
Dharmacon, Inc., Lafayette, CO, USA) at room temperature for 8 min
and counterstained with hematoxylin Gill No. 3 (Sigma-Aldrich;
Merck KGaA) at room temperature for 5 sec.
Two representative images from each tumor tissue
(n=4 per experimental group) were obtained by light microscopy
(×20, magnification) and the number of macrophages/area was
calculated using ImageJ image analysis software (version 1.51k;
National Institutes of Health, Bethesda, MD, USA).
Migration assay
The migration of murine macrophages in response to
tumor cell-conditioned media was assessed using an in vitro
Transwell chamber migration assay. Briefly, macrophages
(2.0×105) isolated from murine bone marrow, were loaded
into the upper chamber of a 5.0-µm porous membrane Transwell insert
(Corning Incorporated, Corning, NY, USA) in a 24-well plate in aMEM
(Gibco; Thermo Fisher Scientific, Inc.) containing FBS (0% for 6 h
and 0.5% for 20 h). The lower chamber was loaded with 72 h old
conditioned media collected from scramble or shCITED2-expressing
tumor cells composed of the same media but containing 10% FBS. The
medium in the upper and lower chambers was maintained in the
presence of 10 ng/ml M-CSF (Thermo Fisher Scientific, Inc.). At the
end of each time point, the 5.0-µm porous membrane Transwell
inserts were removed, fixed in 100% methanol at room temperature
for 15 min, washed in PBS and stained with 2% crystal violet for 20
min at room temperature. For each well, two representative images
(magnification, ×400) were obtained under a light microscope and
the number of macrophages that migrated into the lower chamber, as
indicated by the purple color, was counted. Data are representative
of 3 independent replicates per experimental condition.
For CCL20 neutralization, mouse anti-human CCL20
antibody (cat. no. MAB360; dilution, 1:165; R&D Systems, Inc.,
Minneapolis, MN, USA) or non-specific mouse immunoglobulin G (IgG;
cat. no. MAB002; dilution, 1:165; R&D Systems, Inc.) was added
to the conditioned medium of wild-type MDA-MB-231 cells. The
conditioned media was collected 72 h later and used in the
macrophage migration experiment as aforementioned. For each well, 3
representative images (magnification, ×400) were obtained and the
number of migratory macrophages was counted under a light
microscope. Data are representative of three independent replicates
per experimental condition.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from the cell lines (three biological
replicates per experimental condition) and tumor tissues (6 tumors
per experimental group for MDA-MB-231; 4 tumors per experimental
group for MDA-MB-468) were obtained as previously described
(24). Amplification of 36B4 served
as the internal control in the RT-qPCR reaction (25). Relative expression between samples was
calculated using the comparative 2−ΔΔCq method (30). The primers used in the present study
were: CCL2, sense, 5′-CAGCCAGATGCAATCAATGCC-3′ and antisense,
5′-TGGAATCCTGAACCCACTTCT-3′; CCL3, sense,
5′-AGTTCTCTGCATCACTTGCTG-3′ and antisense,
5′-CGGCTTCGCTTGGTTAGGAA-3′; CCL4, sense,
5′-CTGTGCTGATCCCAGTGAATC-3′ and antisense,
5′-TCAGTTCAGTTCCAGGTCATACA-3′; CCL5, sense,
5′-CCAGCAGTCGTCTTTGTCAC-3′ and antisense,
5′-CTCTGGGTTGGCACACACTT-3′; CCL7, sense, 5′-CCCTAAGCAGAGGCTGGAGA-3′
and antisense, 5′-TGGGTTTTCTTGTCCAGGTG-3′; CCL8, sense,
5′-TGGAGAGCTACACAAGAATCACC-3′ and antisense,
5′-TGGTCCAGATGCTTCATGGAA-3′; CCL13, sense,
5′-CTCAACGTCCCATCTACTTGC-3′ and antisense,
5′-TCTTCAGGGTGTGAGCTTTCC-3′; CCL20, sense,
5′-TGCTGTACCAAGAGTTTGCTC-3′ and antisense,
5′-CGCACACAGACAACTTTTTCTTT-3′; interleukin (IL)-4, sense,
5′-CCAACTGCTTCCCCCTCTG-3′ and antisense,
5′-TCTGTTACGGTCAACTCGGTG-3′; IL-10, sense,
5′-GACTTTAAGGGTTACCTGGGTTG-3′ and antisense,
5′-TCACATGCGCCTTGATGTCTG-3′; IL-13, sense,
5′-CCTCATGGCGCTTTTGTTGAC-3′ and antisense,
5′-TCTGGTTCTGGGTGATGTTGA-3′; IL-34, sense,
5′-CCTGGCTGCGCTATCTTGG-3′ and antisense,
5′-AGTGTTTCATGTACTGAAGTCGG-3′; C-X3-C motif chemokine ligand
(CX3CL)1, sense, 5′-ACCACGGTGTGACGAAATG-3′ and antisense,
5′-TGTTGATAGTGGATGAGCAAAGC-3′. The qPCR parameters were as follows:
1 cycle (95°C for 3 min), 40 cycles (95°C for 30 sec, 61.9°C for 30
sec and 72°C for 45 sec) and final extension (72°C for 2 min).
ELISA
The expression of the chemokines CCL20 and CCL5 in
the whole cell lysates of MDA-MB-231 and MDA-MB-468 orthotopic
tumors and conditioned media of scramble and shCITED2-expressing
MDA-MB-231 and MDA-MB-468 cells, were analyzed using a DuoSet ELISA
kit (cat. no. DY360 for CCL20 and cat. no. DY278 for CCL5; R&D
Systems, Inc.), according to the manufacturer's protocol. The data
are representative of three biological replicates per experimental
condition.
Chromatin immunoprecipitation
(ChIP)
ChIP was performed on nuclear cell lysates using
anti-mouse CITED2 (cat. no. AF5005; dilution, 1:40; R&D
Systems, Inc.) and anti-mouse IgG (cat. no. 515-005-003; dilution,
1:100; Jackson ImmunoResearch Europe, Ltd., Newmarket, UK)
antibodies, according to the manufacturer's protocol of the
SimpleChIP Enzymatic Chromatin IP kit (Cell Signaling Technology,
Inc., Danvers, MA, USA). The promoter primer sequences used for
human CCL20 were: Sense, 5′-AGAAAATATTGGGAATGTACACAG-3′ and
antisense, 5′-CTTCGCACCTTCCCAATATG-3′. Data are representative of
two independent experiments performed in triplicate per
experimental condition.
Statistical analysis
Differences in the number of macrophages
infiltrating the orthotopic tumor, the number of macrophages that
migrated in vitro at each time point and the expression of
CCL20 and CCL5 in the orthotopic tumors and cell culture were
compared using an unpaired Student's t-test.
Differences in the mRNA expression of the different
macrophage-recruiting factors in shCITED2-expressing tumors
relative to that of the scramble-expressing tumors normalized to
1.0; the mRNA expression of the different macrophage recruiting
factors in shCITED2-expressing cells relative to that of
scramble-expressing cells normalized to 1.0 and the fold enrichment
between CITED2 antibody relative to IgG antibody normalized to 1.0
were compared using a one sample t-test.
Differences in the migration of macrophages in the
presence and absence of anti-CCL20 neutralizing antibody and
non-specific IgG were compared using one-way analysis of variance
followed by Tukey's multiple comparison test.
Data are expressed as the mean ± standard deviation.
The graphs and statistical analysis were generated using GraphPad
Prism 7.0 (GraphPad Software Inc., La Jolla, CA, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
Silencing CITED2 in MDA-MB-231 but not
MDA-MB-468 cells attenuates macrophage recruitment
To determine the effect of CITED2 on the recruitment
of TAMs, the present study evaluated the effects of silencing
CITED2 on macrophage presence in two murine orthotopic xenograft
models of human breast cancer using the MDA-MB-231 and MDA-MB-468
cell lines. In previous studies, CITED2 expression was silenced in
cell lines following stable infection with a lentiviral expression
vector (24,25) containing either shRNA specific for
CITED2 (shCITED2) or scrambled shRNA (15,28).
Following implantation in the murine mammary fat pad, the
proliferation of MDA-MB-231 cells was significantly inhibited by
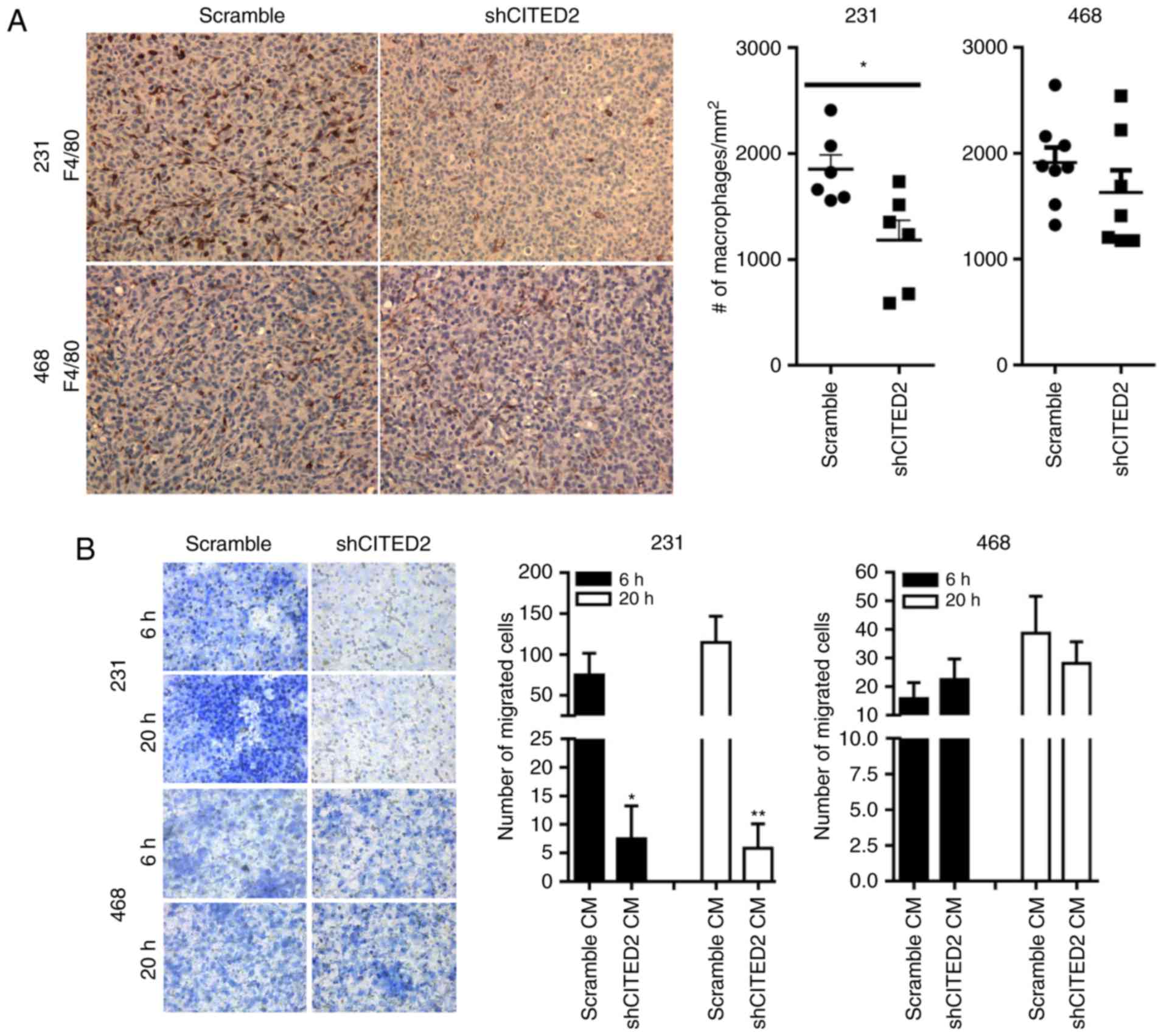
silencing CITED2, while that of MDA-MB-468 was unaffected (24). In the present study, paraffin-embedded
sections of tumor tissues were obtained from each of these models
and immunohistochemical staining for the macrophage marker F4/80
was performed. While CITED2 silencing in MDA-MB-231 xenograft
tumors significantly (P<0.05) decreased the number of
macrophages present relative to the scramble control, it did not
affect the number of macrophages in MDA-MB-468 xenograft tumors
(Fig. 1A), concordant with its effect
on tumor growth in these cell lines.
A key mechanism regulating the presence of
macrophages in the tumor microenvironment is the production of
chemoattractants by tumor cells (11,31). To
determine whether the decreased macrophage numbers observed
following CITED2 silencing in MDA-MB-231 xenograft tumors was due
to altered production of tumor-derived chemotactic factors, the
present study evaluated the ability of conditioned media obtained
from breast cancer cells to induce chemotactic recruitment of
macrophages by performing an in vitro Transwell migration
assay. Consistent with the decreased number of macrophages in
shCITED2-expressing MDA-MB-231 xenograft tumors, significantly
fewer macrophages were recruited in response to conditioned media
from shCITED2-expressing MDA-MB-231 cells compared with conditioned
media from scramble-expressing cells (Fig. 1B). By contrast, adding conditioned
media obtained from shCITED2-expressing MDA-MB-468 cells did not
affect macrophage recruitment (Fig.
1B), which corresponds with the lack of effect of CITED2
silencing on macrophage presence in MDA-MB-468-derived xenograft
tumors. Taken together, these results indicate that CITED2
silencing attenuates macrophage recruitment by MDA-MB-231 cells and
xenograft tumors, concordant with its effect on tumor growth,
potentially by influencing cancer cell production of macrophage
chemoattractants.
CITED2 silencing in MDA-MB-231 cells
decreases expression of the macrophage chemoattractant CCL20
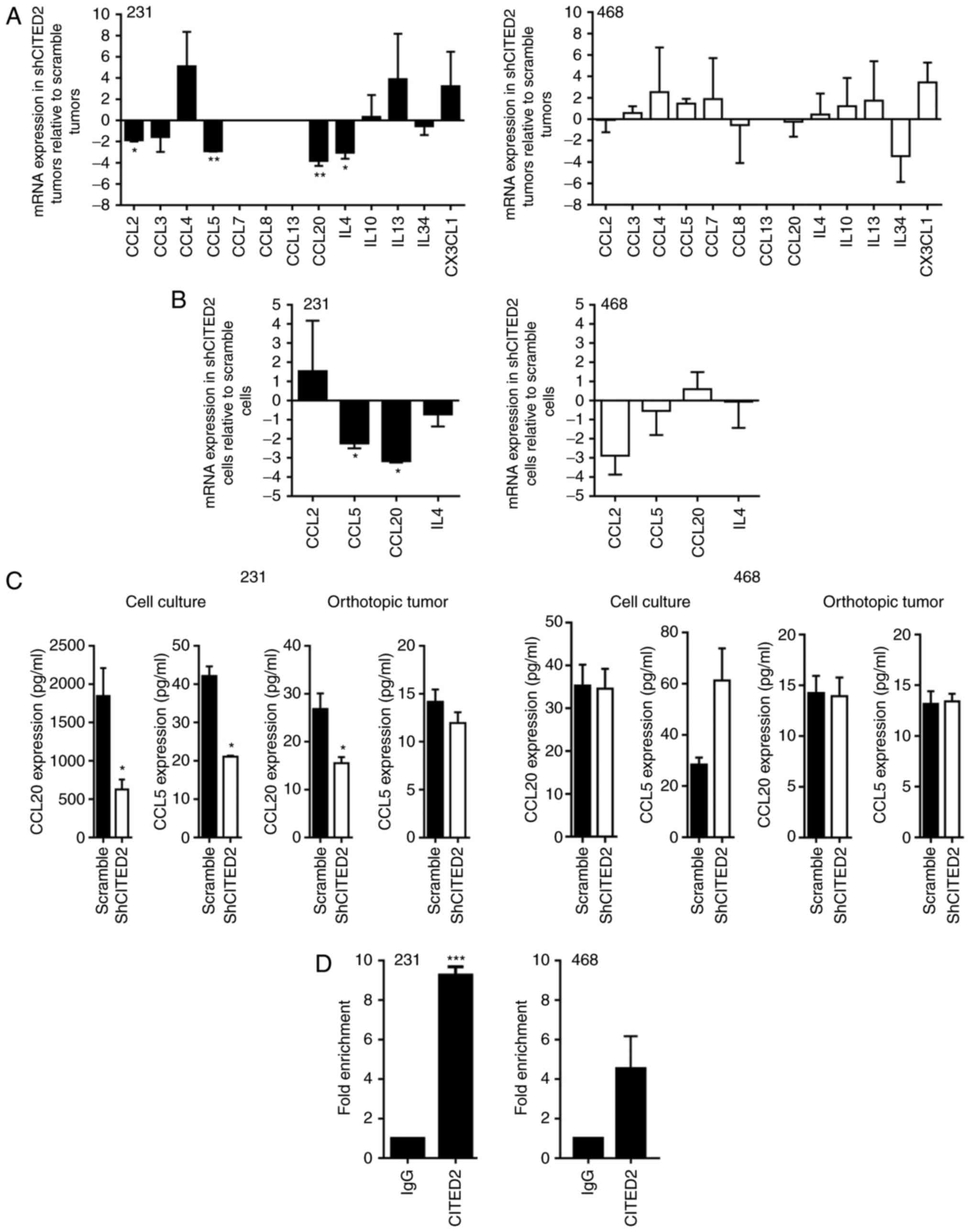
To identify the tumor-secreted factors potentially
associated with CITED2 effects on TAM presence, the present study
assessed the expression of a panel of common macrophage
chemoattractants, including CCL2, CCL3, CCL4, CCL5, CCL7, CCL8,
CCL13, CCL20, IL-4, IL-10, IL-13, IL-34 and CX3CL1. Levels of CCL2,
CCL5, CCL20 and IL-4 mRNA were significantly decreased in
shCITED2-expressing MDA-MB-231 xenograft tumors compared with
scramble shRNA-expressing tumors, while levels of CCL3, CCL4,
IL-10, IL-13, IL-34 and CX3CL1 mRNA were not significantly altered
and CCL7, CCL8 and CCL13 were undetectable (Fig. 2A). Further examining the expression of
CCL2, CCL5, CCL20 and IL-4 in MDA-MB-231 cells cultured in
vitro, wherein CITED2 silencing inhibited macrophage
recruitment, the present study identified that CCL5 and CCL20 mRNA
levels were significantly decreased in shCITED2-expressing cells
compared with scramble shRNA-expressing cells, whereas CCL2 and
IL-4 mRNA levels did not differ significantly (Fig. 2B). Subsequently examining the
expression of CCL5 and CCL20 at the protein level revealed that,
although CCL5 protein expression was significantly attenuated in
MDA-MB-231 cell culture following CITED2 silencing, its expression
in xenograft tumors was unaffected (Fig.
2C), rendering its contribution to macrophage recruitment
unclear. By contrast, CITED2 silencing significantly attenuated
CCL20 protein expression in MDA-MB-231 cells in culture and
established xenograft tumors (Fig.
2C). Furthermore, ChIP analysis revealed that CITED2 localizes
to the CCL20 promoter, which explains the effect of CITED2
silencing on CCL20 expression (Fig.
2D). Corresponding with the lack of effect on macrophage
recruitment by MDA-MB-468 cells and xenograft tumors, CITED2
silencing did not affect the expression of CCL20 or any other
chemoattractants tested, either in orthotopic tumors or cell
culture (Fig. 2A-C). Furthermore,
consistent with the lack of effect on CCL20 expression, CITED2 did
not reveal significant enrichment at the CCL20 promoter in
MDA-MB-468 cells (Fig. 2D).
Collectively, these results indicate that CITED2 regulates the
expression of the macrophage chemoattractant CCL20 in MDA-MB-231
cells, which means that CCL20 may be a potential mediator of the
effects of CITED2 silencing on macrophage recruitment.
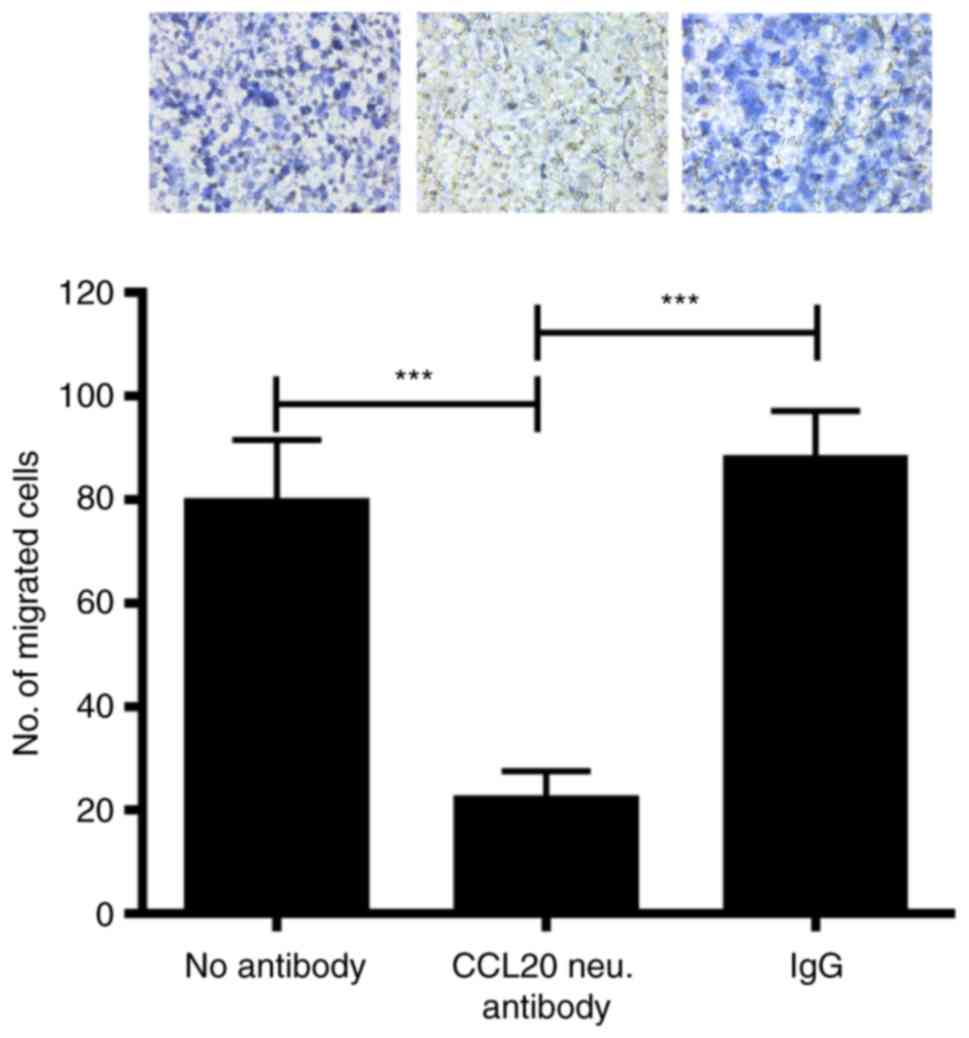
Neutralizing CCL20 in MDA-MB-231
cell-conditioned media inhibits macrophage recruitment
CCL20 expression was significantly decreased in
cultured MDA-MB-231 cells and xenograft tumors following CITED2
silencing (Fig. 2C), corresponding
with the decreased macrophage recruitment observed in xenograft
tumors and cells in culture (Fig. 1);
however, the functional contribution of CCL20 to this phenotype
remained unknown. To determine whether cancer cell-secreted CCL20
serves a function in the ability of MDA-MB-231 cells to recruit
macrophages, CCL20 expression was inhibited in the conditioned
media of these cells and its effect on macrophage chemotaxis was
assessed by a Transwell migration assay. Notably, treating
MDA-MB-231 cell conditioned media with CCL20-neutralizing antibody
significantly attenuated the migration of macrophages compared with
non-specific IgG and ‘no antibody’ controls (Fig. 3). The results of the present study
suggest that MDA-MB-231-secreted CCL20 directly promotes macrophage
recruitment.
Discussion
Previous studies have indicated that TAMs contribute
to the pathogenesis of breast cancer (2–6). Improving
understanding of the tumor-secreted paracrine signaling factors
associated with macrophage recruitment and the mechanism by which
these factors are regulated may aid in the development of novel
therapeutic strategies to treat breast cancer. The present study
demonstrated that CITED2 regulates macrophage infiltration in
MDA-MB-231-derived orthotopic xenograft tumors in vivo and
macrophage recruitment in vitro. It may do so by decreasing
cancer cell production of the macrophage chemoattractant CCL20 via
modulation of CCL20 gene transcription. To the best of our
knowledge, these results provide the first evidence that CITED2 is
associated with the regulation of macrophages in breast cancer and
the regulation of CCL20 expression.
It has been reported that CITED2 regulates breast
tumor growth, at least partly by influencing the tumor vasculature
through the regulation of VEGFA (24). Another previous study demonstrated
that macrophage infiltrates may affect the tumor vasculature by
promoting angiogenesis via the secretion of various pro-angiogenic
factors, including VEGFA (32). In
one such example, inhibiting colony stimulating factor 1, a
cytokine critically associated with the development and survival of
macrophages, decreased macrophage infiltration and angiogenesis in
human mammary tumor xenografts in mice (33). The results of these studies suggest
that the effects of CITED2 on tumor vasculature may be mediated not
only by its ability to directly regulate the production of VEGFA by
tumor cells, but also indirectly via macrophage recruitment. In
addition, studies in pancreatic cancer have demonstrated that
overexpressing or neutralizing CCL20 in cancer cells promote or
inhibit the growth of xenograft pancreatic tumors, respectively
(34). This effect was attributed to
the effect of CCL20 on tumor cell proliferation. Since macrophages
may produce tumor growth factors, such as EGF (35), in addition to pro-angiogenic factors,
it is possible that macrophages recruited by tumor-produced CCL20
may directly influence the growth of breast tumors, in addition to
affecting angiogenesis.
The results of ChIP performed in the present study
indicated that CITED2 was significantly enriched at the CCL20 gene
promoter in MDA-MB-231 but not MDA-MB-468 cells. As a non-DNA
binding transcriptional co-regulator, CITED2 recruitment to gene
promoters is influenced by the transcription factors/co-factors
that interact with CITED2 (15).
Therefore, the lack of significant CITED2 enrichment at the CCL20
promoter in MDA-MB-468 cells may be caused by alterations in the
transcriptional machinery associated with the regulation of CCL20
expression in this cell line. Supporting this, a similar phenomenon
was observed regarding TGF-β in MDA-MB-468 cells, wherein the
transcriptional co-factor SMAD family member (SMAD)4, which
complexes with SMAD2/3 is deleted (36), rendering this cell line unresponsive
to canonical TGF-β signaling. Furthermore, it has previously been
demonstrated that this inherent difference in TGF-β-associated
transcriptional machinery between MDA-MB-231 and MDA-MB-468 cells
impacts the ability of CITED2, a SMAD2/3 co-factor (15), to regulate TGF-β-induced factors,
including VEGFA (24). This may
explain the different effect silencing CITED2 had on xenograft
tumor growth in these cell lines in the present study. Future
studies investigating the mechanism of CCL20 gene regulation may
provide further insight into the differential regulation of CCL20
by CITED2 in MDA-MB-231 and MDA-MB-468 cells and potentially reveal
novel targets for inhibiting the production of CCL20.
In conclusion, the present study demonstrates that
silencing the transcriptional co-activator CITED2 in breast cancer
attenuates macrophage infiltration. The effect of CITED2 on
macrophage recruitment is likely mediated by its regulation of
tumor-produced CCL20. These findings present a novel mechanism by
which CITED2, at least in part, regulates primary tumor growth.
Acknowledgements
The present study was supported by The National
Cancer Institute of the National Institutes of Health (grant no.
R01CA157687).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
De Palma M and Lewis CE: Cancer:
Macrophages limit chemotherapy. Nature. 472:303–304. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nielsen SR and Schmid MC: Macrophages as
key drivers of cancer progression and metastasis. Mediators
Inflamm. 2017:96247602017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pollard JW: Tumour-educated macrophages
promote tumour progression and metastasis. Nat Rev Cancer. 4:71–78.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ruffell B and Coussens LM: Macrophages and
therapeutic resistance in cancer. Cancer Cell. 27:462–472. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang QW, Liu L, Gong CY, Shi HS, Zeng YH,
Wang XZ, Zhao YW and Wei YQ: Prognostic significance of
tumor-associated macrophages in solid tumor: A meta-analysis of the
literature. PloS One. 7:e509462012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Birbrair A, Zhang T, Wang ZM, Messi ML,
Olson JD, Mintz A and Delbono O: Type-2 pericytes participate in
normal and tumoral angiogenesis. Am J Physiol Cell Physiol.
307:C25–C38. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shih JY, Yuan A, Jeremy JW and Yang PC:
Tumor-associated macrophage: Its role in cancer invasion and
metastasis. J Cancer Mol. 101–106. 2006.
|
|
9
|
Frankenberger C, Rabe D, Bainer R,
Sankarasharma D, Chada K, Krausz T, Gilad Y, Becker L and Rosner
MR: Metastasis suppressors regulate the tumor microenvironment by
blocking recruitment of prometastatic tumor-associated macrophages.
Cancer Res. 75:4063–4073. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qian BZ, Li J, Zhang H, Kitamura T, Zhang
J, Campion LR, Kaiser EA, Snyder LA and Pollard JW: CCL2 recruits
inflammatory monocytes to facilitate breast-tumour metastasis.
Nature. 475:222–225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Green CE, Liu T, Montel V, Hsiao G, Lester
RD, Subramaniam S, Gonias SL and Klemke RL: Chemoattractant
signaling between tumor cells and macrophages regulates cancer cell
migration, metastasis and neovascularization. PloS One.
4:e67132009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wyckoff J, Wang W, Lin EY, Wang Y, Pixley
F, Stanley ER, Graf T, Pollard JW, Segall J and Condeelis J: A
paracrine loop between tumor cells and macrophages is required for
tumor cell migration in mammary tumors. Cancer Res. 64:7022–7029.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bhattacharya S, Michels CL, Leung MK,
Arany ZP, Kung AL and Livingston DM: Functional role of p35srj, a
novel p300/CBP binding protein, during transactivation by HIF-1.
Genes Dev. 13:64–75. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Braganca J, Eloranta JJ, Bamforth SD,
Ibbitt JC, Hurst HC and Bhattacharya S: Physical and functional
interactions among AP-2 transcription factors, p300/CREB-binding
protein, and CITED2. J Biol Chem. 278:16021–16029. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chou YT, Wang H, Chen Y, Danielpour D and
Yang YC: Cited2 modulates TGF-beta-mediated upregulation of MMP9.
Oncogene. 25:5547–5560. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Glenn DJ and Maurer RA: MRG1 binds to the
LIM domain of Lhx2 and may function as a coactivator to stimulate
glycoprotein hormone alpha-subunit gene expression. J Biol Chem.
274:36159–36167. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lau WM, Doucet M, Huang D, Weber KL and
Kominsky SL: CITED2 modulates estrogen receptor transcriptional
activity in breast cancer cells. Biochem Biophys Res Commun.
437:261–266. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tien ES, Davis JW and Vanden Heuvel JP:
Identification of the CREB-binding protein/p300-interacting protein
CITED2 as a peroxisome proliferator-activated receptor alpha
coregulator. J Biol Chem. 279:24053–24063. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qu X, Lam E, Doughman YQ, Chen Y, Chou YT,
Lam M, Turakhia M, Dunwoodie SL, Watanabe M, Xu B, et al: Cited2, a
coactivator of HNF4alpha, is essential for liver development. EMBO
J. 26:4445–4456. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu B, Qu X, Gu S, Doughman YQ, Watanabe M,
Dunwoodie SL and Yang YC: Cited2 is required for fetal lung
maturation. Dev Biol. 317:95–105. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yin Z, Haynie J, Yang X, Han B,
Kiatchoosakun S, Restivo J, Yuan S, Prabhakar NR, Herrup K, Conlon
RA, et al: The essential role of Cited2, a negative regulator for
HIF-1alpha, in heart development and neurulation. Proc Natl Acad
Sci USA. 99:pp. 10488–10493. 2002; View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bai L and Merchant JL: A role for CITED2,
a CBP/p300 interacting protein, in colon cancer cell invasion. FEBS
Lett. 581:5904–5910. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chou YT, Hsieh CH, Chiou SH, et al: CITED2
functions as a molecular switch of cytokine-induced proliferation
and quiescence. Cell Death Differ. 19:2015–2028. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jayaraman S, Doucet M and Kominsky SL:
Down-regulation of CITED2 attenuates breast tumor growth, vessel
formation and TGF-β-induced expression of VEGFA. Oncotarget.
8:6169–6178. 2017.PubMed/NCBI
|
|
25
|
Jayaraman S, Doucet M, Lau WM and Kominsky
SL: CITED2 modulates breast cancer metastatic ability through
effects on IKKα. Mol Cancer Res. 14:730–739. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lau WM, Weber KL, Doucet M, Chou YT, Brady
K, Kowalski J, Tsai HL, Yang J and Kominsky SL: Identification of
prospective factors promoting osteotropism in breast cancer: A
potential role for CITED2. Int J Cancer. 126:876–884.
2010.PubMed/NCBI
|
|
27
|
Sun HB, Zhu YX, Yin T, Sledge G and Yang
YC: MRG1, the product of a melanocyte-specific gene related gene,
is a cytokine-inducible transcription factor with transformation
activity. Proc Natl Acad Sci USA. 95:pp. 13555–13560. 1998;
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chou YT and Yang YC: Post-transcriptional
control of Cited2 by transforming growth factor beta. Regulation
via Smads and Cited2 coding region. J Biol Chem. 281:18451–18462.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . Guide for the Care and Use of Laboratory Animals. 8th.
Washington (DC): National Academies Press (US); 2011, ISBN-13.
ISBN: 978-0-309-15400-0ISBN-10. ISBN: 0-309-15400-6PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantiative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Baay M, Brouwer A, Pauwels P, Peeters M
and Lardon F: Tumor cells and tumor-associated macrophages:
Secreted proteins as potential targets for therapy. Clin Dev
Immunol. 2011:5651872011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bingle L, Lewis CE, Corke KP, Reed MW and
Brown NJ: Macrophages promote angiogenesis in human breast tumour
spheroids in vivo. Br J Cancer. 94:101–107. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Aharinejad S, Paulus P, Sioud M, Hofmann
M, Zins K, Schäfer R, Stanley ER and Abraham D: Colony-stimulating
factor-1 blockade by antisense oligonucleotides and small
interfering RNAs suppresses growth of human mammary tumor
xenografts in mice. Cancer Res. 64:5378–5384. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Beider K, Abraham M, Begin M, Wald H,
Weiss ID, Wald O, Pikarsky E, Abramovitch R, Zeira E, Galun E, et
al: Interaction between CXCR4 and CCL20 pathways regulates tumor
growth. PloS One. 4:e51252009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Goswami S, Sahai E, Wyckoff JB, Cammer M,
Cox D, Pixley FJ, Stanley ER, Segall JE and Condeelis JS:
Macrophages promote the invasion of breast carcinoma cells via a
colony-stimulating factor-1/epidermal growth factor paracrine loop.
Cancer Res. 65:5278–5283. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schutte M, Hruban RH, Hedrick L, Cho KR,
Nadasdy GM, Weinstein CL, Bova GS, Isaacs WB, Cairns P, Nawroz H,
et al: DPC4 gene in various tumor types. Cancer Res. 56:2527–2530.
1996.PubMed/NCBI
|