Introduction
Nasopharyngeal carcinoma (NPC) is an epithelial
malignancy of the head and neck with the highest incidence rate in
southern China, with a high prevalence of 20–50 cases per 100,000
individuals (1). Several factors are
involved in the progression and development of NPC, including
Epstein-Barr virus (EBV) infection, genetic components,
environmental factors and interactions between these factors
(2). Although the application of
intensity-modulated radiotherapy and chemoradiotherapy has
increased the 5-year overall survival rate to approximately 70%,
the prognosis remains poor because of distant metastasis and
recurrence (3,4). Thus, it is critical to understand the
molecular mechanisms that underlie the progression of NPC in order
to promote the development of novel therapeutic strategy.
MicroRNAs (miRNAs) are small non-coding RNA
sequences, 19–25 nucleotides long, which negatively regulate gene
expression by targeting the 3-untranslated region (3′-UTR) of their
target mRNA transcripts at the post-transcriptional level (5,6).
Increasing evidence showed that miRNAs regulate a wide range of
cell functions, including proliferation, migration, differentiation
and apoptosis (5). Emerging evidence
indicates that miRNAs are abnormally expressed in human disease,
especially cancer, and they function as either tumor suppressors or
oncogenes in the progression of tumors (7,8). Multiple
miRNAs have been found to be dysregulated in NPC, including miR-124
(9), miR-29c (10), miR-34c (11) and miR-16 (12). These findings indicated that the
abnormally expressed miRNAs may contribute to the progression and
development of NPC. miR-93, derived from a paralogue (miR-106b-25),
is dysregulated in several cancer types, such as epithelial ovarian
carcinoma (13), ischemic heart
disease (14) and bone cancer pain
mouse model (15). The evidence
indicates that miR-93 plays a key role in cancer progression.
CDKN1A, also known as p21, is a cyclin-dependent
kinase inhibitor that inhibits the complexes of CDK2 and CDK1, and
regulates the cell cycle progression at G1 and S phase (16). Its expression is tightly controlled by
the tumor suppressor protein p53, through which CDKN1A mediates the
p53-dependent cell cycle G1 phase arrest in response to a variety
of stress stimuli (17). p21 can also
interact with proliferating cell nuclear antigen (PCNA) and plays a
regulatory role in DNA damage repair and S-phase DNA replication
(18).
In the present study, we demonstrated that miR-93
was upregulated in NPC tissues and cells. Re-expression of miR-93
promoted cell growth in vitro. CDKN1A was downregulated by
miR-93 by directly targeting its 3′-UTR. The newly identified
miR-93/CDKN1A axis may provide new evidence to understand the
molecular mechanism of NPC and present a new target for the
treatment of NPC.
Materials and methods
Tissue specimens and cell lines
A collection of 23 freshly frozen NPC biopsy
specimens and 13 normal nasopharyngeal epithelial tissue specimens
were obtained from the Yidu Central Hospital. The specimens were
confirmed by histopathological examination. None of the patients
with NPC had received chemotherapy or radiotherapy before biopsy.
Written informed consent was obtained from the patients. This study
was approved by the Ethics Review Committee of the Yidu Central
Hospital (Shandong, China).
The human NPC cell line SUNE-1 and the human
immortalized nasopharyngeal epithelial cell line NP69 were used in
this study. All cells were cultured in a humidified atmosphere at
37°C with 5% CO2.
Oligonucleotide and plasmid
transfection
Cells were cultured on 6-well plates 24 h prior to
transfection. The oligonucleotide was transfected into NPC cells
using Lipofectamine™ 2000 reagent (Invitrogen, Carlsbad, CA, USA).
The transfected cells were incubated at 37°C in complete medium and
harvested at the indicated time-points.
Quantitative real-time PCR
qRT-PCR was used to detect miR-93 and CDKN1A
expression levels. Total RNA was extracted from cells or tissues by
using TRIzol reagent (Invitrogen) according to the manufacturers
instruction. qRT-PCR was performed using agents from Takara Bio
(Dalian, China) with the Stratagene Mx3000P real-time PCR system
(Agilent Technologies, Santa Clara, CA, USA). For miR-93 detection,
U6 was used as an internal control. For CDKN1A detection, GAPDH was
used as an endogenous control. The PCR primers for CDKN1A were as
follows: 5-GTGGGGTTATCTCTGTGTTAGGG-3 and 5-CCCTGT
CCATAGCCTCTACTGC-3. The primers for GAPDH were as follows:
5-CTCCTCCTGTTCGACAGTCAGC-3 and 5-CCCAATACGACCAAATCCGTT-3. The
relative expression was calculated with the 2−∆∆CT. All
reactions were performed in triplicate.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
MTT assay was used to detect the proliferation of
NPC cells. After transfection, cells were plated in 96-well plates
at 1,000 cells/well and cultured for 1–5 days. Cell viability was
examined once daily. On the indicated days, the MTT dye were added
to the well and incubated for 4 h at 37°C. Then the medium were
removed and dimethyl sulfoxide (DMSO) at 150 µl/well (Sigma Corp.,
Ronkonkoma, NY, USA) was added to dissolve the formazan crystals.
The absorbance at 490 nm was measured on a spectrophotometer
(Perkin-Elmer, Waltham, MA, USA).
Luciferase reporter assay
293 cells were used for luciferase reporter assay.
3′-UTR of CDKN1A sequences were cloned from mRNA and inserted into
the downstream of luciferase expression gene in psiCHECK-2 plasmids
(psi-CDKN1A-3′-UTR).
Cells were co-transfected with miR-93 mimic or
control, and psi-CDKN1A-3′-UTR-WT or psi-CDKN1A-3′-UTR-MUT using
Lipofectamine 2000 (Invitrogen). The mutation (MUT) of 3′-UTR was
generated using QuikChange Multi Site-Directed Mutagenesis kit
(Agilent Technologies). After 24-h incubation, cells were collected
and assayed for luciferase activity using the Dual-Luciferase
reporter assay system (Promega Bio Systems, Sunnyvale, CA, USA).
The Renilla luciferase activity was used as internal
control.
Western blotting
Cells were cultutred, lysed and processed for
western blotting according to the standard protocols. The primary
antibodies include anti-CDKN1 (1:1,000; Cell Signaling Technology,
Danvers, MA, USA), anti-GAPDH (1:1,000; Bioworld Technology, Inc.,
Nanjing, China). Then the membrane was incubated with appropriate
secondary antibody at room temperature for 1 h. Protein bands were
detected using enhanced chemiluminescence system (GE Healthcare,
Chicago, IL, USA).
Statistical analysis
All statistical analyses were performed using SPSS
16.0 software (SPSS, Inc., Chicago, IL, USA). All data are
presented as mean ± SD. Differences between groups were assessed
using the Students t-test or Tukey's post hoc test after one-way
analysis of variance in SPSS. Differences were considered
significant at P<0.05.
Results
miR-93 is upregulated in NPC tissues
and cell lines
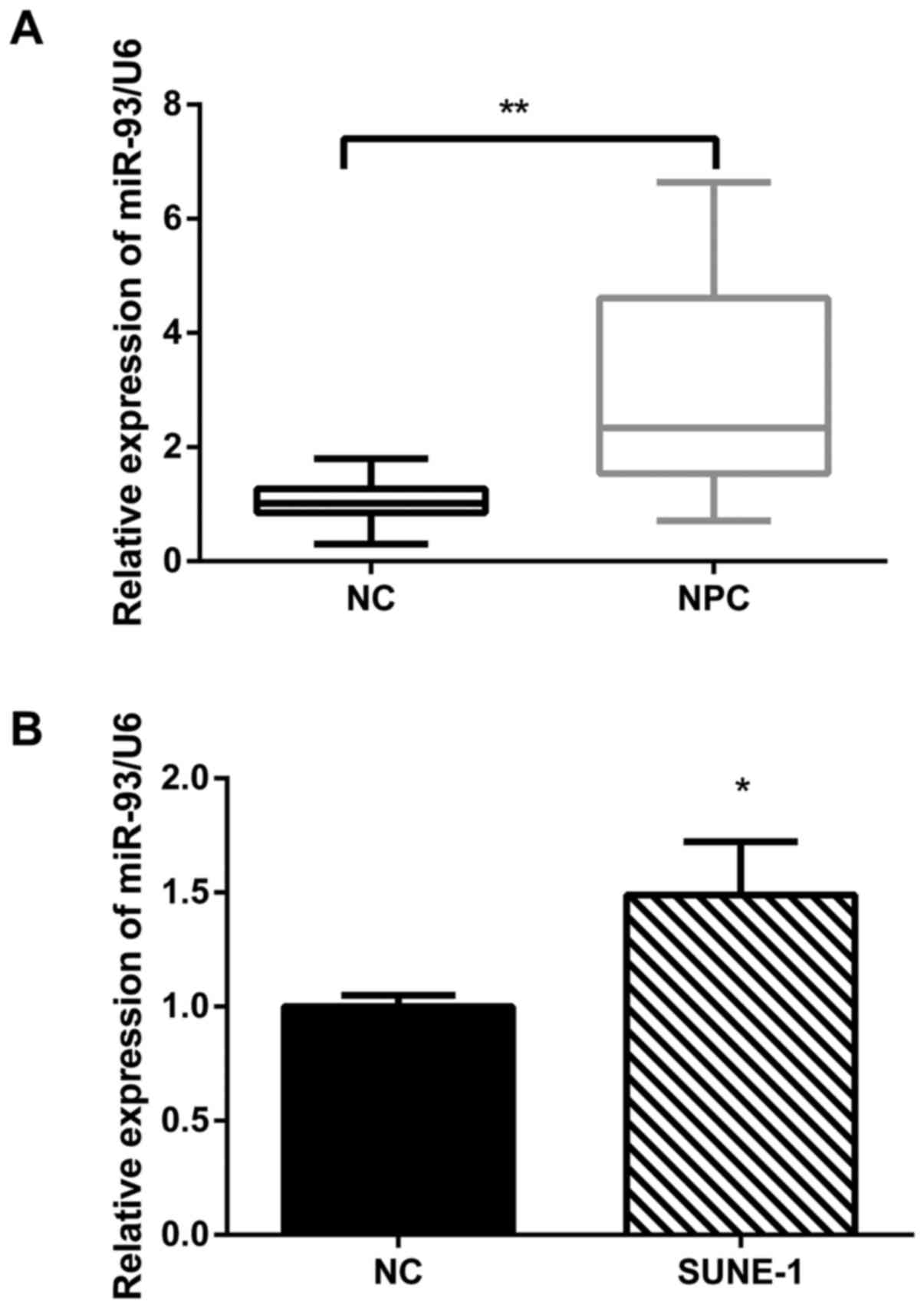
To study the expression level of miR-93 in NPC, 23
freshly frozen NPC biopsy specimens and 13 normal nasopharyngeal
epithelial tissue specimens were analyzed by the qRT-PCR. The
expression of miR-93 was increased by ~2-fold in 23 NPC tissues
compared with 13 normal tissues (Fig.
1A). Furthermore, we detected the expression of miR-93 in
SUNE-1 NPC cell lines. The qRT-PCR results revealed that miR-93 was
significantly upregulated in SUNE-1 cell line compared with the
normal nasopharyngeal epithelial cell line NP69 (Fig. 1B). Taken together, miR-93 was
aberrantly upregulated in NPC tissues and cell lines.
miR-93 promotes NPC cell proliferation
in vitro
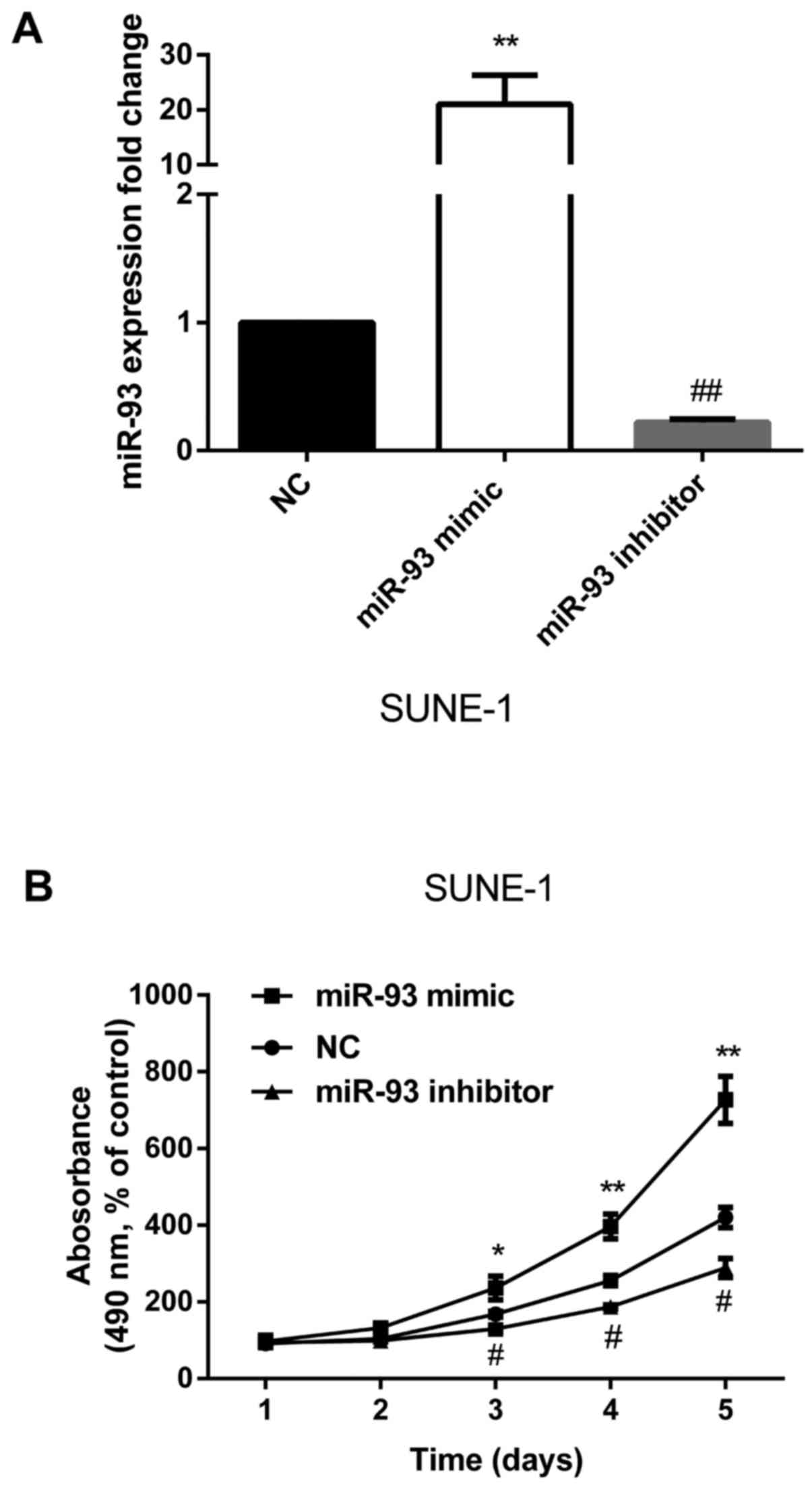
To further understand the biological functions of
miR-93 in the development of NPC, we re-expressed miR-93 or
inhibited miR-93 expression separately. The successful
re-expression or inhibition of miR-93 was confirmed by qRT-PCR
(Fig. 2A). MTT assay showed that
re-expression of miR-93 significantly promoted cell proliferation
and miR-93 inhibitor significantly decreased cell proliferation
compared with control (Fig. 2B).
Collectively, these results indicate that miR-93 enhances the
proliferation of NPC.
CDKN1A is a direct target of
miR-93
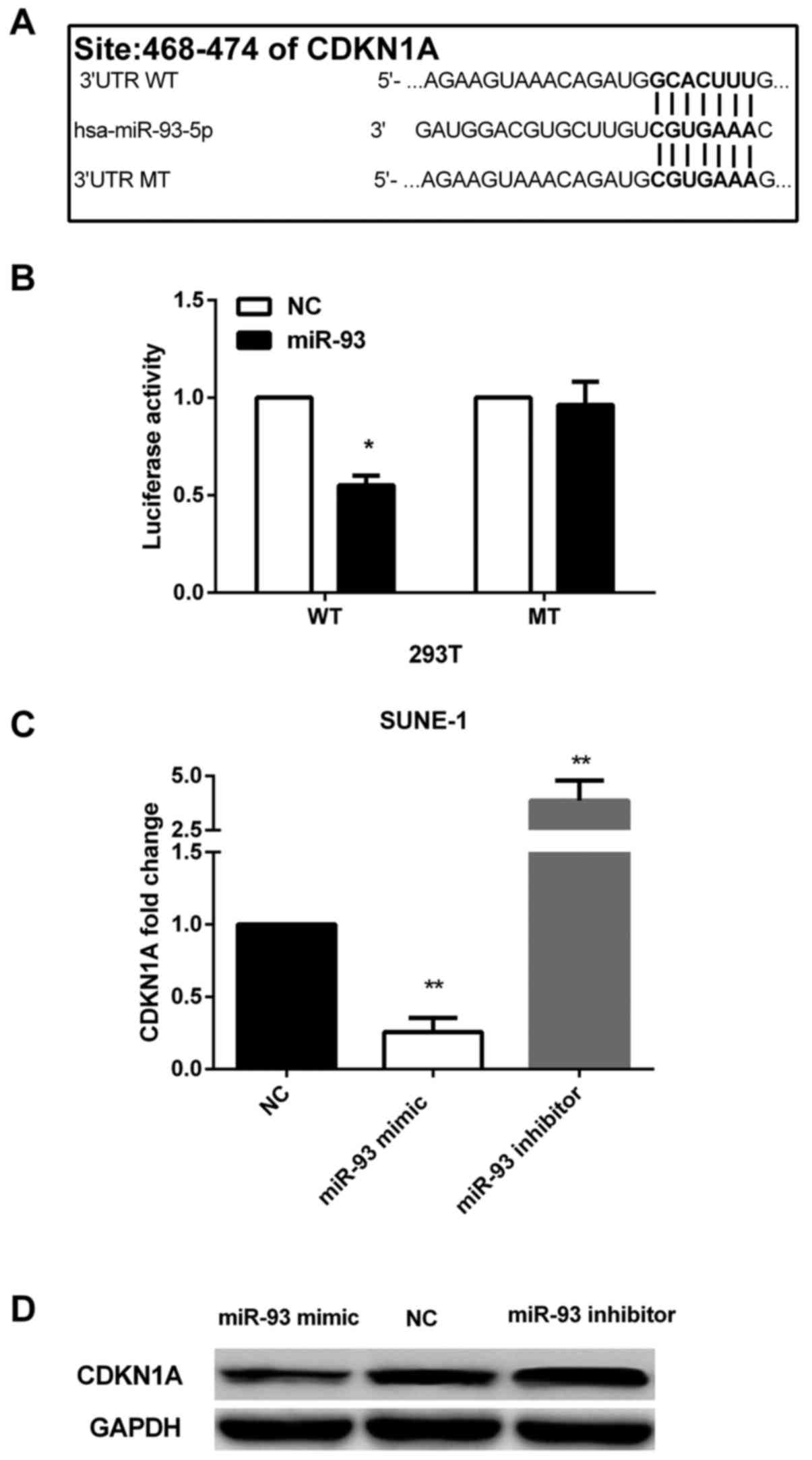
To explore the downstream of miR-93, we performed
bioinformatics analysis by using two online algorithms, TargetScan
(http://www.targetscan.org/) and miRanda
(http://www.microrna.org/microrna/home.do) to predict
its putative mRNA targets. CDKN1A was identified as a direct
target, and a highly conserved putative binding site was found at
468–474 bp of CDKN1A 3′-UTR (Fig.
3A). Thus, we further performed Dual-Luciferase reporter assay
to confirm the prediction. In cells co-transfected with
psi-CDKN1A-3UTR-WT and miR-93 mimic, luciferase activity was
dramatically decreased compared with cells co-transfected with
miR-control and psi-CDKN1A-3UTR-WT. However, the inhibition of
luciferase activity was abolished when cells were co-transfected
with miR-93 mimic and psi-CDKN1A-3UTR-MT (Fig. 3B).
Besides, re-expression of miR-93 in SUNE-1 cells
could reduce the expression of CDKN1A at both miRNA and protein
levels and silencing of miR-93 could increase endogenous CDKN1A at
both mRNA and protein levels in SUNE-1 cells (Fig. 3C and D). The evidence indicated that
miR-93 downregulated CDKN1A by directly targeting its 3′-UTR.
The expression of miR-93 is inversely
correlated with the expression of CDKN1A
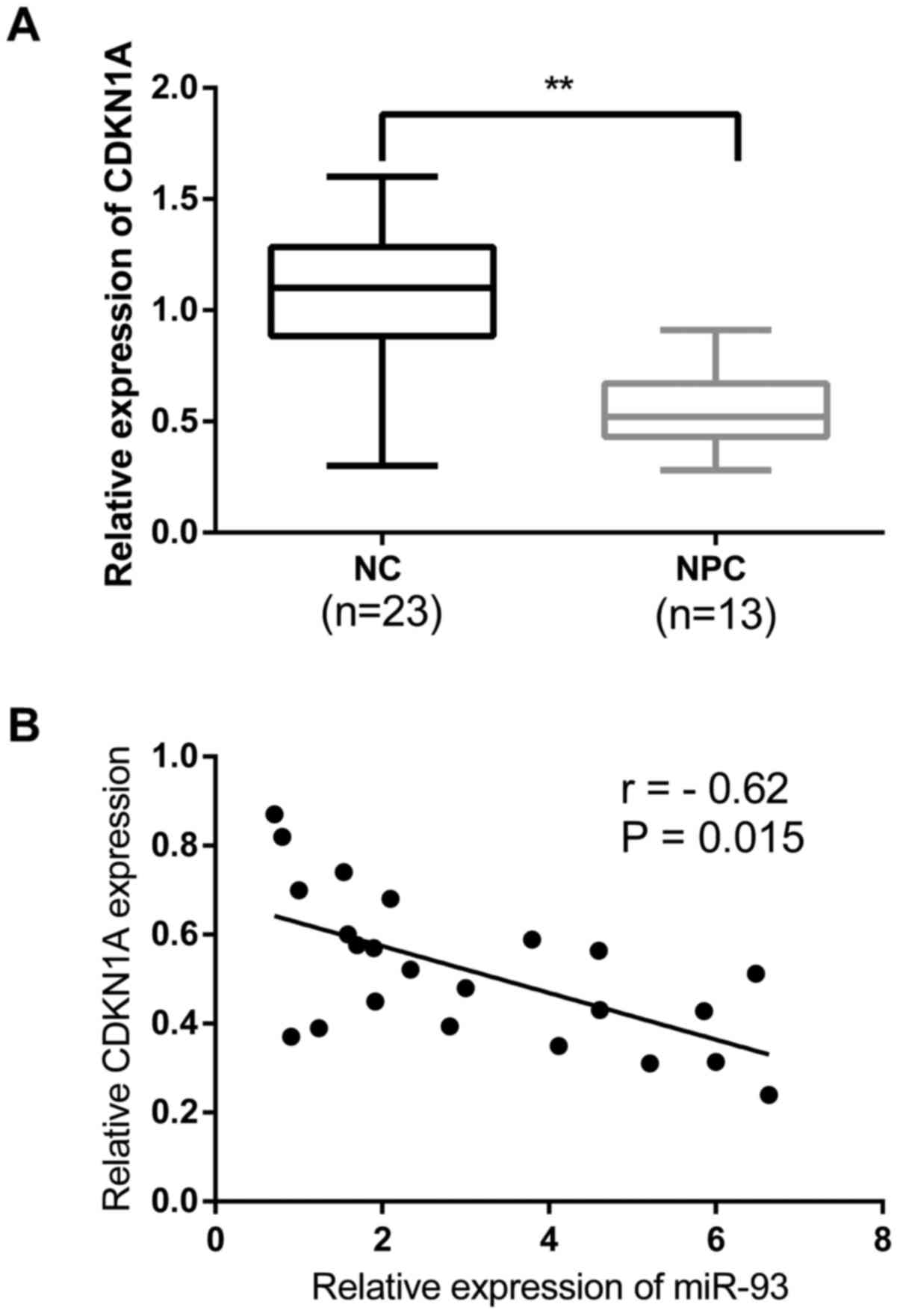
To further investigate the expression of CDKN1A, 23
freshly frozen NPC tissues and 13 normal nasopharyngeal epithelial
tissue specimens were detected to analyze the clinicopathological
significance. The relative expression of CDKN1A in NPC tissues were
significantly decreased compared with that in normal tissues as
assayed by qRT-PCR (Fig. 4A).
Furthermore, we detected the relationship between the endogenous
expression levels of CDKN1A and miR-93 in the same NPC tissues. The
results showed a significant inverse correlation between the
expression of miR-93 and CDKN1A in NPC tissues (r=−0.62; P<0.05)
(Fig. 4B).
Discussion
Dysregulation of miRNAs has been reported in many
types of cancer. Increasing evidence indicate that miRNAs function
as oncogenes or tumor suppressors. Although the tumorigenesis of
NPC is a multistage process relying on the control of the gene
expression, several dysregulated miRNA expression profiles and
dysregulated genes of NPC have been identified in some studies
(19–21). Several miRNAs are identified
upregulated, including miR-206, miR-99a, miR-30a and some are
down-regulated, such as let-7i, miR-7 and miR-222 (21). The capability to regulate
proliferation, invasion and migration is considered as an important
determinant in the process of tumor progression. In the present
study, we demonstrated that the expression of miR-93 was
significantly increased in NPC cell lines and tumor tissues. The
evidence indicates that miR-93 was upregulated in NPC, which is
consistent with the results obtained from bone cancer (15), myocardial ischemia/reperfusion (I/R)
injury (14) and polycystic ovarian
syndrome (22). The enhanced
proliferation of NPC cell lines indicated that miR-93 was an
oncogene in NPC. To date, the ectopic expression of miR-93 is shown
as a frequent epigenetic tumor promoter in various types of cancer
(23–25). Furthermore, aberrant expression of
miR-93 has been demonstrated to regulate tumor migration and
invasion (26). The evidence
indicates that miR-93 exerts pivotal biological and pathological
functions, which may contribute to the development and progression
of NPC.
miRNAs exert their function by regulating their
target genes by binding to the 3′-UTR. Several studies have
identified the target genes of miR-93, including RAB11 family
interacting protein 1 (27), PTEN
(14), PDCD4 (24), Sirt7 and Tbx3 (28). In the present study, we identified
CDKN1A as a direct target of miR-93 by bioinformatics analysis and
luciferase activity report, which is consistent with the findings
in polycystic ovarian syndrome (22).
Re-expression of miR-93 significantly reduced CDKN1A expression and
silencing of miR-93 increased CDKN1A expression. Furthermore, we
demonstrated that the expression of miR-93 was inversely correlated
with the expression of CDKN1A protein. Recently, it has been
reported that CDKN1A is directly regulated by multiple miRNAs,
including miR-663 (29), miR-106b
(30), miR-221 and miR-222 (31). In NPC, p21, functioning as a critical
cell cycle regulator, is regulated by miR-663 to promote the G1/S
transition (29). miR-106b family
members contribute to tumor cell proliferation partially by
regulating cell cycle progression via p21/CDKN1A and by modulating
checkpoint functions (30). Abnormal
expression of miR-221 and miR-222 promotes cancer growth by
inhibiting the expression of p27Kip1 (31). Thus, emerging evidence presents that
miRNAs induce the aberrant G1/S transition, promote cell
proliferation and tumorigenicity in cancer by targeting CDKN1A.
In conclusion, this study found that miR-93 was
upregulated in NPC cell lines and tissues. Re-expression of miR-93
promotes NPC proliferation, functioning as an oncogene. CDKN1A was
identified as a direct target of miR-93. It is suggested that
miR-93/CDKN1A axis may present a new target for the treatment of
NPC.
References
|
1
|
Jia WH, Luo XY, Feng BJ, Ruan HL, Bei JX,
Liu WS, Qin HD, Feng QS, Chen LZ, Yao SY, et al: Traditional
Cantonese diet and nasopharyngeal carcinoma risk: A large-scale
case-control study in Guangdong, China. BMC Cancer. 10:4462010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xu FH, Xiong D, Xu YF, Cao SM, Xue WQ, Qin
HD, Liu WS, Cao JY, Zhang Y, Feng QS, et al: An epidemiological and
molecular study of the relationship between smoking, risk of
nasopharyngeal carcinoma, and Epstein-Barr virus activation. J Natl
Cancer Inst. 104:1396–1410. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yoshizaki T, Ito M, Murono S, Wakisaka N,
Kondo S and Endo K: Current understanding and management of
nasopharyngeal carcinoma. Auris Nasus Larynx. 39:137–144. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lai SZ, Li WF, Chen L, Luo W, Chen YY, Liu
LZ, Sun Y, Lin AH, Liu MZ and Ma J: How does intensity-modulated
radiotherapy versus conventional two-dimensional radiotherapy
influence the treatment results in nasopharyngeal carcinoma
patients? Int J Radiat Oncol Biol Phys. 80:661–668. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bandres E, Agirre X, Ramirez N, Zarate R
and Garcia-Foncillas J: MicroRNAs as cancer players: Potential
clinical and biological effects. DNA Cell Biol. 26:273–282. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Valencia-Sanchez MA, Liu J, Hannon GJ and
Parker R: Control of translation and mRNA degradation by miRNAs and
siRNAs. Genes Dev. 20:515–524. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Peng XH, Huang HR, Lu J, Liu X, Zhao FP,
Zhang B, Lin SX, Wang L, Chen HH, Xu X, et al: MiR-124 suppresses
tumor growth and metastasis by targeting Foxq1 in nasopharyngeal
carcinoma. Mol Cancer. 13:1862014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu N, Tang LL, Sun Y, Cui RX, Wang HY,
Huang BJ, He QM, Jiang W and Ma J: MiR-29c suppresses invasion and
metastasis by targeting TIAM1 in nasopharyngeal carcinoma. Cancer
Lett. 329:181–188. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rietzschel N, Pierik AJ, Bill E, Lill R
and Mühlenhoff U: The basic leucine zipper stress response
regulator Yap5 senses high-iron conditions by coordination of
[2Fe-2S] clusters. Mol Cell Biol. 35:370–378. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
He Q, Ren X, Chen J, Li Y, Tang X, Wen X,
Yang X, Zhang J, Wang Y, Ma J, et al: miR-16 targets fibroblast
growth factor 2 to inhibit NPC cell proliferation and invasion via
PI3K/AKT and MAPK signaling pathways. Oncotarget. 7:3047–3058.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen X, Chen S, Xiu YL, Sun KX, Zong ZH
and Zhao Y: RhoC is a major target of microRNA-93-5P in epithelial
ovarian carcinoma tumorigenesis and progression. Mol Cancer.
14:312015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ke ZP, Xu P, Shi Y and Gao AM: MicroRNA-93
inhibits ischemia-reperfusion induced cardiomyocyte apoptosis by
targeting PTEN. Oncotarget. 7:28796–28805. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xiao WF, Li YS, Lou W, Cai T, Zhang S, Hu
XY, Zhang XW and Luo W: MicroRNA-93-5p may participate in the
formation of morphine tolerance in bone cancer pain mouse model by
targeting Smad5. Oncotarget. 7:52104–52114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gartel AL and Radhakrishnan SK: Lost in
transcription: p21 repression, mechanisms, and consequences. Cancer
Res. 65:3980–3985. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rodriguez R and Meuth M: Chk1 and p21
cooperate to prevent apoptosis during DNA replication fork stress.
Mol Biol Cell. 17:402–412. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xiong Y, Zhang H and Beach D: D type
cyclins associate with multiple protein kinases and the DNA
replication and repair factor PCNA. Cell. 71:505–514. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ho SL, Chan HM, Ha AW, Wong RN and Li HW:
Direct quantification of circulating miRNAs in different stages of
nasopharyngeal cancerous serum samples in single molecule level
with total internal reflection fluorescence microscopy. Anal Chem.
86:9880–9886. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bruce JP, Yip K, Bratman SV, Ito E and Liu
FF: Nasopharyngeal Cancer: Molecular Landscape. J Clin Oncol.
33:3346–3355. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang HY, Yan LX, Shao Q, Fu S, Zhang ZC,
Ye W, Zeng YX and Shao JY: Profiling plasma microRNA in
nasopharyngeal carcinoma with deep sequencing. Clin Chem.
60:773–782. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang L, Huang J, Li L, Chen Y, Chen X,
Zhao X and Yang D: MicroRNA-93 promotes ovarian granulosa cells
proliferation through targeting CDKN1A in polycystic ovarian
syndrome. J Clin Endocrinol Metab. 100:E729–E738. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu J, Wang H, Wang Y, Li Z, Pan Y, Liu Q,
Yang M and Wang J: Repression of the miR-93-enhanced sensitivity of
bladder carcinoma to chemotherapy involves the regulation of LASS2.
Onco Targets Ther. 9:1813–1822. 2016.PubMed/NCBI
|
|
24
|
Liang H, Wang F, Chu D, Zhang W, Liao Z,
Fu Z, Yan X, Zhu H, Guo W, Zhang Y, et al: miR-93 functions as an
oncomiR for the downregulation of PDCD4 in gastric carcinoma. Sci
Rep. 6:237722016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qu MH, Han C, Srivastava AK, Cui T, Zou N,
Gao ZQ and Wang QE: miR-93 promotes TGF-β-induced
epithelial-to-mesenchymal transition through downregulation of
NEDD4L in lung cancer cells. Tumour Biol. 37:5645–5651. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu YF, Mao YP, Li YQ, Ren XY, He QM, Tang
XR, Sun Y, Liu N and Ma J: MicroRNA-93 promotes cell growth and
invasion in nasopharyngeal carcinoma by targeting disabled
homolog-2. Cancer Lett. 363:146–155. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang J, Wang F, Xu J, Wang X, Ye F and
Xie X: Micro ribonucleic acid-93 promotes oncogenesis of cervical
cancer by targeting RAB11 family interacting protein 1. J Obstet
Gynaecol Res. 42:1168–1179. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cioffi M, Vallespinos-Serrano M, Trabulo
SM, Fernandez-Marcos PJ, Firment AN, Vazquez BN, Vieira CR, Mulero
F, Camara JA, Cronin UP, et al: MiR-93 controls adiposity via
inhibition of Sirt7 and Tbx3. Cell R. 12:1594–1605. 2015.
View Article : Google Scholar
|
|
29
|
Yi C, Wang Q, Wang L, Huang Y, Li L, Liu
L, Zhou X, Xie G, Kang T, Wang H, et al: MiR-663, a microRNA
targeting p21WAF1/CIP1, promotes the proliferation and
tumorigenesis of nasopharyngeal carcinoma. Oncogene. 31:4421–4433.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ivanovska I, Ball AS, Diaz RL, Magnus JF,
Kibukawa M, Schelter JM, Kobayashi SV, Lim L, Burchard J, Jackson
AL, et al: MicroRNAs in the miR-106b family regulate p21/CDKN1A and
promote cell cycle progression. Mol Cell Biol. 28:2167–2174. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
le Sage C, Nagel R, Egan DA, Schrier M,
Mesman E, Mangiola A, Anile C, Maira G, Mercatelli N, Ciafrè SA, et
al: Regulation of the p27Kip1 tumor suppressor by miR-221 and
miR-222 promotes cancer cell proliferation. EMBO J. 26:3699–3708.
2007. View Article : Google Scholar : PubMed/NCBI
|