Introduction
Single nucleotide polymorphisms (SNPs) are important
in determining the pathogenesis of various diseases, identifying
effective therapeutic targets and screening individualized
chemotherapy programs. A number of in-depth studies have explored
SNP variations in coding genes and the influence of this on the
incidence and development of human cancer (1,2). However,
research investigating SNPs in non-coding micro (mi)RNA genes and
their effects on breast cancer progression remain in the early
stages (3).
MiRNAs are involved in a number of important
biological processes, from embryonic development to the evolution
of various diseases. These processes are often accompanied by the
abnormal expression of various miRNAs and their target genes, with
previous studies having demonstrated that miRNA expression is
dysregulated in cancer, and experimental data having indicated that
cancer phenotypes may be modified by targeting miRNA expression
(4). It has been demonstrated that
SNPs arising in miRNA coding gene sequences affect the synthesis of
miRNA, whereas SNPs in miRNA target gene sequences affect the
interaction between miRNA and its target genes, thus affecting the
regulatory function of miRNAs (5).
The polymorphism of miRNA (miR)-323b (4) is associated with hepatitis B virus
replication, whereas the variation of rs4846049 in the
3′-untranslated region (UTR) of 5,10-methylenetetrahydrofolate
reductase affects the binding of miRNA to increase the risk of
coronary heart disease (6). Similar
studies have been conducted in other types of cancer, including
gastric and lung cancer (7,8).
SNP variation in miRNA coding genes is associated
with the onset and progression of breast cancer. It has been
demonstrated that variation at the miR-27a rs895819 site may affect
miR-27a expression: Compared with carriers of A-alleles, G-allele
carriers have a significantly reduced risk of developing breast
cancer (P=0.032) (9). Binding sites
from miR-515-5p exist in the 3′-UTR of the insulin-like growth
factor 1 receptor gene; variation in the rs28674628 site in this
region affects the binding of miR-515-5p to the gene, thus increase
the risk of breast cancer (10). SNP
variations of miRNAs, including miR-192-a2 and miR-423 are also
associated with breast cancer risk (11,12).
The present study aimed to provide a theoretical
basis for the exploration of SNPs in miRNA coding genes to be
applied in clinical practice as molecular markers. This would
entail the SNP screening of miRNA coding genes associated with
breast cancer metastasis. The association between SNP sites in
miRNA coding genes and the incidence and development of breast
cancer in clinical specimens was also investigated in the current
study.
Materials and methods
Cells
The highly malignant human breast cancer cell lines
BT549, HS578T, MDA-MB-231 and SUM159PT, and the less malignant
human breast cancer cell lines BT474, MCF7, MDA-MB-468, SK-BR-3,
T47D and ZR-75-1, as well as the immortalized normal human breast
cell lines MCF10A and MCF12A were acquired as gifts from Professor
Shi Huidong, Cancer Research Center of Medical College of Georgia
(Augusta, GA, USA). The human lung cancer cell lines 95-D and A549,
human stomach cancer cell lines AGS and MGC-803, and the human
hepatocarcinoma cell lines SMMC-7721 were purchased from the Cell
Bank of the Chinese Academy of Sciences (Shanghai, China). Tissue
chips of human breast diseases were acquired from Alenabio (Xi'an,
China; cat. no. BR723). The chips included 83 cases of breast
cancer and 35 cases of breast lobular hyperplasia in a dual-chip
matrix. The tissue samples were surgically resected during surgery
and fixed for 24 h with 10% neutral-buffered formaldehyde, with a
dot diameter of 1.1 mm and a thickness of 4 µm. A total of 60 human
breast surgical resection samples were obtained from the Department
of Pathology of Jinhua Hospital of Zhejiang University (Jinhua,
China) between March 2010 to August 2012. All patients were female
with a mean age of 62 years (range, 32–79 years). Written informed
consent was obtained from all patients for the use of their
samples.
Analysis of expression profile chip of
miRNA
Total RNA from cell lines was extracted using the
RNeasy kit (Qiagen, Inc., Valencia, CA, USA) and RNase-Free DNase
Set (Qiagen, Inc.), according to the manufacturer's protocols.
Expression profiles of extracted RNA from all cell lines were
analyzed using a GeneChip miRNA 2.0 array (Affymetrix; Thermo
Fisher Scientific, Inc.). A FlashTag Biotin RNA Labeling kit
(Genisphere, Hatfield, PA, USA) was used to perform biotin labeling
of 1 µg miRNA. Chip hybridization, elution, image acquisition and
data standardization were conducted according to the manufacturer's
protocol (Fluidics Protocol FS450_0003; Affymetrix; Thermo Fisher
Scientific, Inc.). Partek Genomic Suite v.6.2 software (Partek
Inc., St. Loius, MI, USA) was used to analyze the chip data.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
First-strand complementary DNA (cDNA) was
synthesized with oligo-dT or random hexamers as primers, using the
SuperScript First-Strand Synthesis System (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
An equal volume mixture of cDNA products (50 ng) were used as
templates for PCR amplification. QPCR reactions were performed in a
25 µl volume with iQ™ SYBR Green Supermix (Bio-Rad Laboratories,
Inc., Hercules, CA, USA), and 200 nM each of forward and reverse
primers using an iCyler iQ instrument and iQ software (Version 2.0;
Bio-Rad Laboratories, Inc.). Each sample was analyzed in
triplicate. QPCR cycling conditions included an initial
denaturation step of 1 min at 95°C, followed by 40 cycles
consisting of 30 sec at 95°C, 5 sec at 95°C and 30 sec at 60°C.
Mean quantification cycle (Cq) values from the triplicate qPCRs for
a gene of interest (GOI) were normalized against the average Cq
values for GAPDH from the same cDNA sample (13). The following primers were used:
miR-205 forward, 5′-ACAGGCTGAGGTTGACATGC-3′; and reverse,
5′-GAGTTACTCTTGCTGCTGCTG-3′; GAPDH forward,
5′-TGCACCACCAACTGCTTAGC-3′; and reverse,
5′-GGCATGGACTGTGGTCATGAG-3′.
Situ hybridization
Highly malignant human breast cancer cell lines
BT549, MDA-MB-231 and low malignant human breast cancer cell lines
MCF7 and T47D were used in the subsequent experiment. Cells were
grown on coverslips for 2–3 days, and the coverslips were washed
with PBS at 37°C, 3 times. Cells were fixed with 4% neutral
formalin for 5 min, and incubated with operating fluid of
proteinase K (0.25 mg/ml in DEPC H2O-HCl; Sigma Aldrich; Merck
KGaA), at 37°C for dissociation after 15 min, followed by situ
hybridization operation. Breast cancer and breast benign lesions
(fibroadenoma, breast hyperplasia), sectioned at 5-µm onto tissue
slides were deparaffinized, treated with 0.2 N HCl, incubated in 1
M sodium thiocyanate and immersed in protease solution (Vysis Inc.,
Downers Grove, IL, USA) for 10 min at 37°C. The tissues were then
fixed with 10% formalin for 10 min, denatured for 5 min at 72°C and
sequentially incubated in 75, 90 and 100% ethanol. The tissues were
then treated with proteinase K for 6 min at 37°C, followed by their
dehydration and hybridization. The hybridization mixture contained
the labeled oligonucleotide probe, 50% formamide, 10 mmol/l
Tris-HCl, 1 mmol/l vanadyl-ribonucleoside complex (94740; Sigma
Aldrich; Merck KGaA), 1 mmol/l CTAB (855820; pH 7.0; Sigma Aldrich;
Merck KGaA), 0.15 mol/l NaCl, 1 mmol/l EDTA (pH 7.0), 1X Denhardt's
mixture and 10% dextran sulfate. Subsequent to hybridization, the
slides were washed 3 times, for 30 min each time, in 0.1 mol/l
Tris-buffered saline (TBS) at room temperature. The slides were
then treated with TBS [100 mmol/l Tris (pH 7.5) and 150 mmol/l
NaCl] containing a 1% blocking reagent (Roche Diagnostics,
Shanghai, China) and 0.03% Triton X-100 for 30 min at room
temperature, and incubated for 30 min with anti-dioxigenin alkaline
phosphatase-conjugated antibodies (Roche Diagnostics) diluted at
1:4,000 in TBS containing 0.03% Triton X-100 and a 1% blocking
reagent. Subsequent to being washed three times, for 15 min in TBS
and 0.05% Tween 20, the slides were rinsed in a diammonimum
phosphate (DAP)-buffer [100 mmol/l Tris (pH 9.5) 100 mmol/l NaCl,
50 mmol/l MgCl2] and hybridization signals were subsequently
visualized using nitroblue tetrazolium and
5-bromo-4-chloro-3-indolyl phosphate as substrates [DAP-buffer (100
mmol/l Tris, pH 9.5, 100 mmol/l NaCl and 50 mmol/l MgCl2) in 10%
PVA (341584; Sigma Aldrich)].
Extraction of DNA from
paraffin-embedded tissues
DNA was extracted from paraffin-embedded breast
cancer and breast benign lesions (fibroadenoma, breast hyperplasia)
tissues. Areas of the tissue and cytology slides containing the
tumor cells of interest were marked by a cytopathologist using a
pen. A diamond-tipped pencil was then used to mark the underside of
the slide. Following removal of the cover slip, tumor cells were
scraped with a 26-gauge needle. Briefly, 50–100 µl of DNA
extraction buffer solution (50 mM Tris buffer, pH 8.3; 1 mM EDTA,
pH 8.0; 5% Tween-20 and 200 µg/ml proteinase K) with 10% resin was
added to the scraped cells. Following incubation at 56°C for ~1 h,
the tube was heated to 100°C for 20 min followed by centrifugation
at 900 × g for 10 min at 4°C to pellet the debris. The recovered
supernatant was used for the PCR.
DNA sequencing
Amplified specimens processed by PCR were sent to
Sangon Biotech Co., Ltd., (Shanghai, China) for sequencing and the
genotypes of specimens were analyzed using the obtained sequence
diagram. The obtained sequence was analyzed using the 2014 UCSC
Genome Browser (http://genome.ucsc.edu) (14).
Statistical analysis
Data was analyzed using SPSS 20.0 (IMB Corp.,
Armonk, NY, USA). Student's t-test was performed for between-group
comparisons, χ2 for comparisons between count data rate
and correlations were determined using Pearson's correlation
analysis. P<0.05 was considered to indicate a statistically
significant difference.
Results
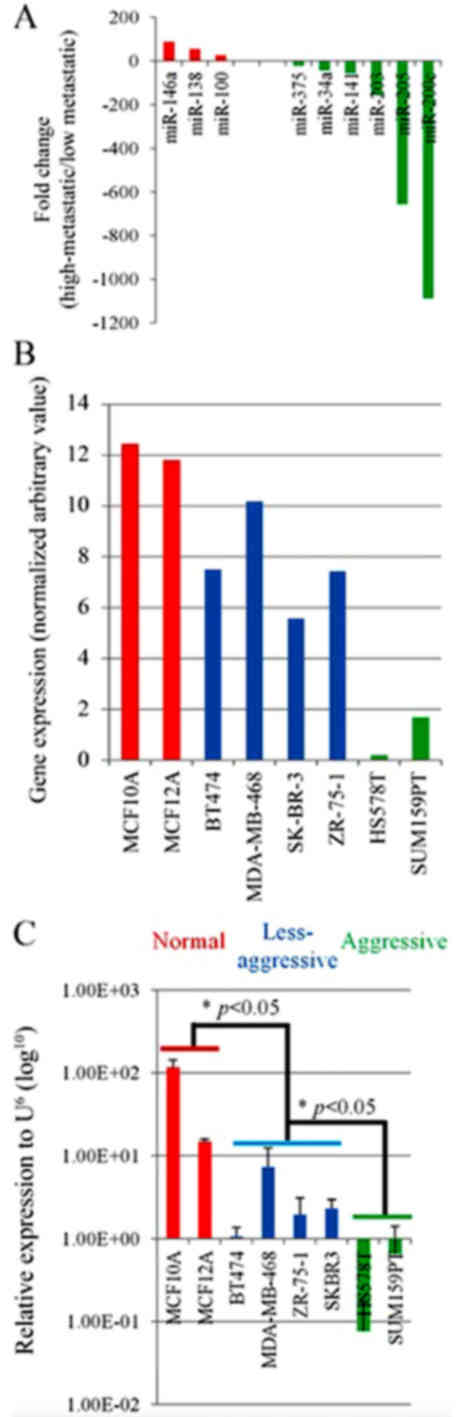
Analysis of miRNA expression profile
chip in breast cancer cells with more or less metastasis
A total of 4 highly metastatic breast cancer cell
lines and 6 less metastatic breast cancer cell lines, as well as 2
immortalized cell lines were selected to determine miRNA expression
profiles. Following data processing and statistical analysis of
chip results, it was identified that there were 9 differentially
expressed miRNAs among the 12 cell lines (P<0.01, numerical
value ≥20 or ≤-20). Compared with the less metastatic group, levels
of miR-100, miR-138 and miR-146a were increased in the highly
metastatic cell lines, whereas levels of miR-375, miR-34a, miR-141,
miR-203, miR-205 and miR-200c were decreased in the highly
metastatic group (Fig. 1A).
The expression of miR-205 in more and
less metastatic breast cancer cells
A total of 8 breast cancer cell lines with different
metastatic properties underwent RT-qPCR to measure the expression
of miR-205. The results were consistent with the aforementioned
miRNA chip data, whereby miR-205 was expressed in increased amounts
in the less metastatic cell lines compared with more metastatic
cell lines (P<0.05; Fig. 1B and
C).
miR-205 expression in breast cancer
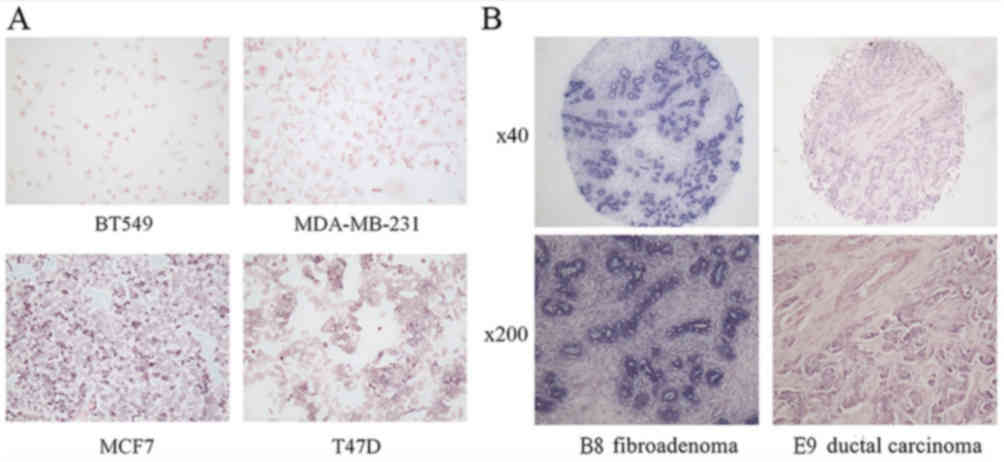
cells and tissue chips measured by in-situ hybridization
In-situ hybridization was performed to
analyze the expression of miR-205 following the fixation of 4
breast cancer cell lines with formalin; the concentration of
miR-205 of the digoxigenin-labelled cDNA probe was 100 nmol/l. The
results revealed that the expression of miR-205 in BT549 and
MDA-MB-231 was decreased compared with that in MCF7 and T47D
(Fig. 2A). The presence of blue or
blue purple granules located in breast glandular epithelium or
cancer cell cytoplasm and karyon indicated the positive expression
of miR-205, and blue or blue purple granules located in karyon of
breast ductal epithelium or mesenchymal cells indicated the
positive expression of U6. Following the in-situ
hybridization of breast normal tissues, no blue granules were
exhibited; the results of the U6 probe hybridization revealed
positive signal of blue purple granules located in karyon of breast
ductal epithelium or mesenchymal cells (Fig. 2B). The results of the miR-205 probe
hybridization revealed a positive signal with blue purple granules
located in the breast ductal epithelium or cancer cell cytoplasm
and karyon (Fig. 2B). In the 36 cases
of normal and benign breast lesions, 33 cases (91.67%), indicating
positive expression (+) of miR-205; in the 36 cases of breast
cancer, 23 cases (63.89%). MiR-205 expression in benign breast
lesions was deemed to be significantly higher than in breast cancer
tissue (Table I; P=0.011). The
expression of miR-205 was not associated with tumor-node-metastasis
stage or clinical stage of breast cancer.
 | Table I.Analysis of the expression of miR-205
in normal, benign breast and breast carcinoma. |
Table I.
Analysis of the expression of miR-205
in normal, benign breast and breast carcinoma.
| Case | Total, n | Positive miR-205
expression, n (%) | P-value | χ2 |
|---|
| Normal, benign
breast | 36 | 33 (91.67) | 0.011a | 6.509 |
| Breast carcinoma | 23 | 23 (63.89) |
|
|
Analysis of PCR amplification and
sequencing of miR-205
A total of 10 cell lines were cultured for this
experiment, including 4 breast cancer cell lines (BT-549,
MDA-MB-231, MCF7 and T47D), 2 lung cancer cell lines (95-D and
A549), 2 stomach cancer cell lines (AGS and MGC-803) and a liver
cancer cell line (SMMC-7721). Cells were collected and RNA and DNA
were extracted separately. The SinoBio PCR system was used to
amplify miR-205 genes and the electrophoresis results of amplified
products are presented in Fig. 3A.
Fig. 3B demonstrates the site map of
SNPs of the miR-205 gene as determined using the UCSC Genome
Browser. Amplified products were sent to Sangon Biotech for
sequencing. Sequencing results revealed that miR-205-rs3842642 was
present in a number of these 10 cell lines and was broadly divided
into four types (Fig. 3C-E); 9/9
homozygous genotype, 13/13 homozygous genotype, 9/13 heterozygous
genotype and 7/9 special genotype. Among them, AGS, BT-549 and MCF7
belong to the 9/9 homozygous genotype, 95-D, A549, MGC-803, HEPG2
and SMMC-7721 belong to the 9/13 heterozygous genotype, MDA-MB-231
is a 13/13 homozygous genotype and T47D is a special genotype.
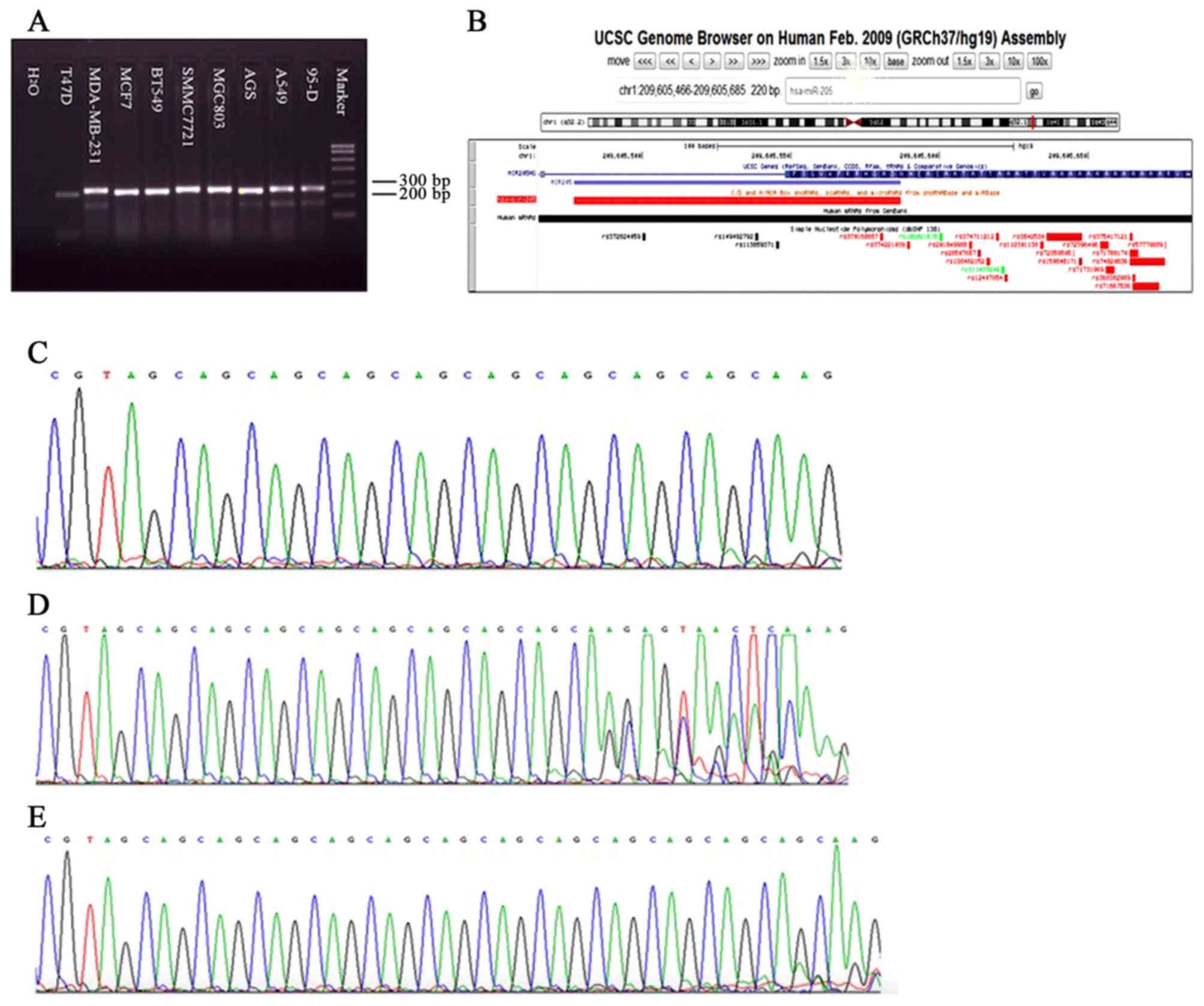 | Figure 3.Analysis of polymerase chain reaction
amplification and sequencing of miR-205. (A) Amplification of
miR-205 gene in 10 cell lines. (B) Gene SNP sites of miR-205;
miR-205-rs3842642 commonly existed in these 10 cell lines and was
broadly divided into four types: 9/9 homozygous genotype, 13/13
homozygous genotype, 9/13 heterozygous genotype and 7/9 special
genotype. Among them, AGS, BT-549 and MCF7 belong to the 9/9
homozygous genotype, 95-D, A549, MGC-803, HEPG2 and SMMC-7721
belong to the 9/13 heterozygous genotype, MDA-MB-231 is the 13/13
homozygous genotype and T47D is the special genotype. (C) rs3842530
9/9 homozygous genotype,
CGT(AGC)9AAG/CGT(AGC)9AAG. (D) rs3842530 9/13
heterozygous genotype,
CGT(AGC)9AAG/CGT(AGC)13AAG. (E) rs3842530
13/13 homozygous genotype,
CGT(AGC)13AAG/CGT(AGC)13AAG. SNP, single
nucleotide polymorphism; miR, microRNA. |
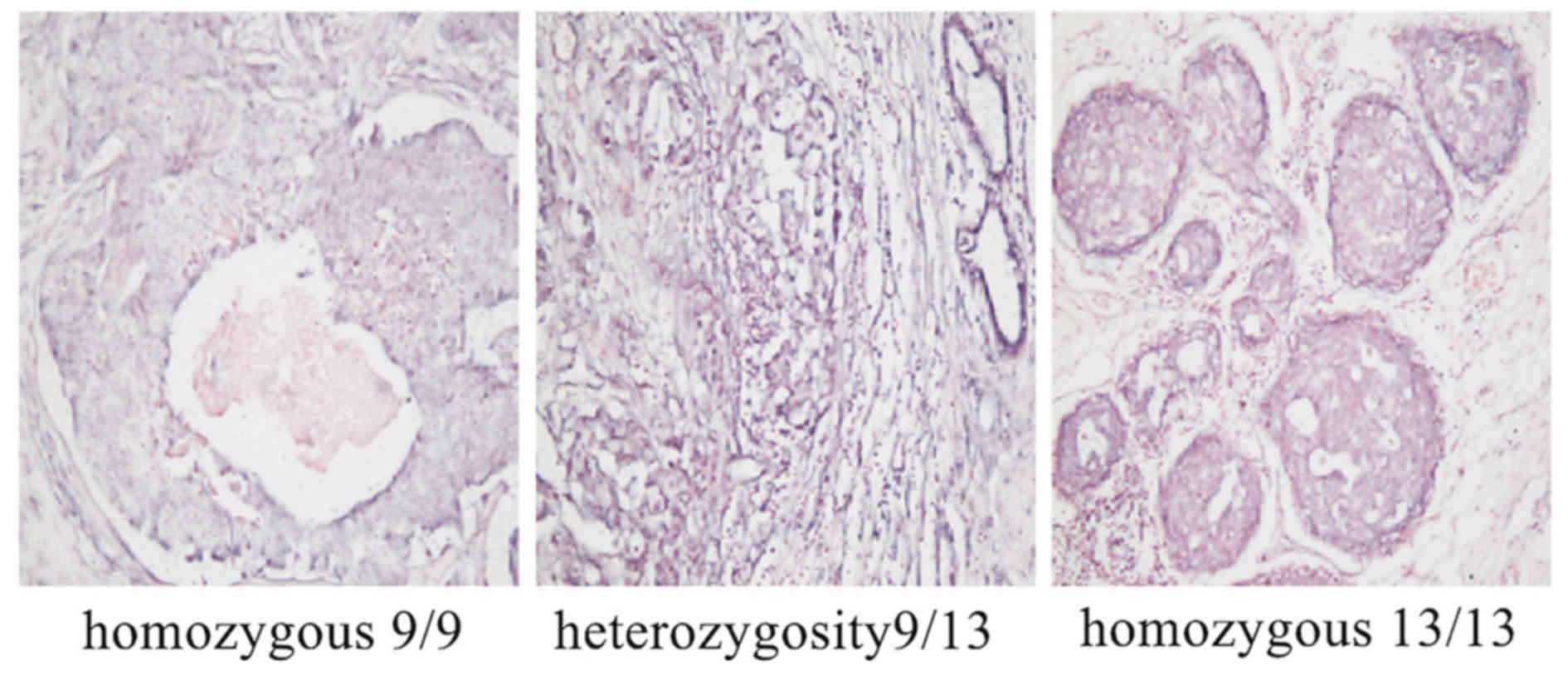
Expression of miR-205 in specimens of
different rs3842642 polymorphisms
DNA was extracted from paraffin-embedded breast
cancer specimens using xylene solvent. These fragments were then
amplified using PCR and sent to Sangon Biotech for sequencing. By
analyzing the genotype of various specimens according to the
returned sequencing results, it was identified that there were 106
cases of deletion of homozygosis, 69 cases of deletion of
heterozygosity, 14 cases of wild genotype and 4 cases of special
genotype, in which 3 cases of homozygosis (9/9), homozygosis
(13/13) and heterozygote (9/13) were respectively selected for
in-situ hybridization investigation of miR-205 expression,
the results demonstrated that all were negative (Fig. 4).
Analysis of rs3842642 polymorphism in
breast cancer and benign breast lesions
DNA was extracted from the paraffin-embedded tissue
specimens of breast benign lesions and sent for sequencing. The
results of sequencing revealed 119 cases of deletion of
homozygosis, 63 cases of heterozygous genotype, 13 cases of wild
genotype and 2 cases of special genotype. An association analysis
was conducted between obtained results for benign lesions and the
results from the breast cancer tissue (Table II). Differences in SNP genotyping
between breast cancer and benign lesion tissues were not
significant as determined by the χ2 analysis; however,
compared with breast cancer tissue, benign lesion tissues exhibited
an increased ratio of 9/9 homozygous genotype. The case number of
SNP genotyping corresponding to statistical pathological indicators
[estrogen receptor (ER), progesterone receptor (PR), erb-B2
receptor tyrosine kinase 2 (ERBB2)] and association analysis
between SNP genotyping and specific pathological indicators were
conducted (Table II).
| Table II.Single nucleotide polymorphism
genotyping and statistical pathological indicators. |
Table II.
Single nucleotide polymorphism
genotyping and statistical pathological indicators.
|
|
| rs3842530
polymorphism |
|---|
|
|
|
|
|---|
| Characteristic | n | Homozygous 9/9, n
(%) | Heterozygosity 9/13,
n (%) | P-value |
|---|
| Breast lesions |
|
|
| 0.362 |
| Benign
lesion | 195 | 119 (61.03) | 76 (38.97) |
|
| Breast
cancer | 189 | 106 (56.08) | 83 (43.92) |
|
| Breast ER
expression |
|
|
| 0.014a |
| High | 45 | 19 (42.22) | 26 (57.78) |
|
|
Low | 62 | 41 (66.13) | 21 (33.87) |
|
| Breast PR
expression |
|
|
| 0.112 |
|
High | 26 | 11 (42.31) | 15 (57.69) |
|
|
Low | 66 | 40 (60.61) | 26 (39.39) |
|
| Breast ERBB2
expression |
|
|
| 0.391 |
|
High | 18 | 9 (50.00) | 9 (50.00) |
|
|
Low | 77 | 47 (61.04) | 30 (38.96) |
|
Discussion
Breast cancer is one of the most common tumors
diagnosed in women and an important cause of mortality in breast
cancer is the metastasis of cancer cells (15). The epithelial-mesenchymal transition
(EMT), as a primary stage of transfer, has been a key area of
research into tumor metastasis, and is a process associated with a
decrease in E-cadherin expression (16). The expression of E-cadherin is
regulated by multiple genes, including nuclear factor-κB and the
matrix metalloproteinase family (17). These genes and their associated cell
signaling pathways are influenced by miRNAs, which serve important
functions in promoting breast cancer progression by stimulating the
EMT process (18,19). MiR-205 is a conserved RNA molecule
widely distributed in squamous epithelial tissues of higher living
organisms, as its regulation and control of numerous target genes
is associated with the onset and development of tumors. Studies
investigating miR-205 are primarily focused in the field of tumor
biology and have revealed that miR-205 expression may affect the
incidence and development of a number of tumors; however, miR-205
serves distinct functions in different types of tumors (20). The expression of miR-205 in breast
cancer is generally downregulated; this may be due to the
regulation of upstream genes, such as ERBB2 (21), by miR-205 or may be due to SNP
variations in the miR-205 sequence. The different functions of
miR-205 in breast cancer may be associated with the cellular
environment, cancer subtype, origin of the tumor, and the
development phase of the tumor (22).
The present study revealed that, compared with less metastatic
breast cancer cell lines, the expression of miR-205 was decreased
in highly metastatic breast cancer cells, suggesting that miR-205
may suppress the metastasis of breast cancer. The results of the
present study and the aforementioned studies are in agreement with
the results of investigations into the breast cancer cell line and
clinical specimens as identified by Wang et al (23) and Liu et al (24).
Numerous different genes regulate the onset and
development of breast cancer. MiRNA and various other genes
constitute this vast regulatory network and SNPs are present in the
various genes involved. SNP variation in these genes may lead to a
variation in gene expression, thus affecting breast cancer risk
(25). Previous studies have
demonstrated that, Breast Cancer Association Consortium rs1045485
and rs1982073 SNPs are associated with an increased risk of breast
cancer (26). Wang et al
conducted an investigation into 491 Chinese patients with breast
cancer and 502 healthy individuals (27), and analyzed the distributional
difference of genotypes between them. It was identified that
rs10889677 and rs1884444 in the interleukin 23 receptor 3′-UTR
region were associated with an increased risk of breast cancer
among Chinese women, which also affected the early development of
tumors. It was also identified that the cytoskeleton gene IQ motif
containing GTPase activating protein 1 (IQGAP) was regulated by
miR-124. The results of a study in 1,541 patients with breast
cancer and 1,598 healthy controls indicated that the variation in
rs1042538 sites in the IQGAP 3′-UTR region affected the binding and
regulation of miR-124 to this gene; thus, this may be developed as
a potential molecular marker to evaluate the risk and prognosis of
patients with breast cancer (28).
The results of the aforementioned studies indicate that SNP
variation in genes associated with breast cancer is associated with
occurrence and development of breast cancer. The important
regulatory effect of miRNA in breast cancer has enabled research on
miRNA SNP to attract increased attention, with more studies being
performed investigating the expression of miR-205 in breast cancer;
however, to the best of our knowledge, there have been no studies
investigating the association between SNP variation of miR-205 and
breast cancer. Following verification that miR-205 is
differentially expressed in breast cancer, the present study
explored whether SNP variation is associated with the differential
expression of miR-205.
In order to improve detection of SNP distribution in
miR-205, two lines of lung cancer, gastric cancer and liver cancer
cells were investigated along with the breast cancer cell lines.
SNP distribution in each cell was detected using DNA sequencing and
through this analysis, the presence of rs3842642 was identified and
four genotypes within the 10 cell lines that were investigated were
also identified: A 9/9 homozygous genotype, a 9/13 heterozygous
genotype, a 13/13 homozygously inserted genotype and a special
genotype, the high frequency of such sites in cancer cells suggest
that they may be associated with tumorigenesis. By detecting the
expression of miR-205 in the aforementioned cell lines using
RT-qPCR and comparing SNP genotypes in each cell line, it was
identified that the expression of miR-205 increased in breast
cancer cell lines with an increase of the number of AGC deletions
at this site. This association was observed in 4 breast cancer cell
lines and it was unclear whether variation at this site
significantly affected the expression of miR-205; an increased
number of samples are required verify this phenomenon. Therefore,
193 cases of breast cancer specimens were used for sequencing
analysis in order to determine the genotype of each specimen. The
analysis revealed that rs384242 widely existed in these specimens
including these four genotypes of the site. The expression of
miR-205 in in-situ hybridization was analyzed by separately
selecting 3 cases of homozygous deletion, heterozygous deletion and
homozygous insertion in breast cancer specimens However the
specimen sample size selected was relatively small, thus no
significant association was identified in these specimens. A larger
sample size may assist in verifying the aforementioned
association.
Under normal conditions, miR-205 serves a tumor
suppressive function in breast cancer and the expression of miR-205
in breast cancer is inhibited (29).
If there is a distributional difference of these three genotypes of
rs3842642 in breast cancer specimens and breast benign lesions, the
variation in this site may be associated with miR-205 expression
and t breast cancer progression. Statistical analysis indicated a
P-value >0.05 when comparing genotypes between breast cancer and
benign specimens, suggesting that there was no significant
difference. However, the proportion of 9/9 homozygous deletion in
benign breast lesion tissue was slightly increased compared with
breast cancer tissue specimens. The differential distribution of
distinct genotypes of rs3842642 in breast cancer and benign breast
lesions may be identified in a larger sample size.
ER, PR and ERBB2 are the primary indicators of
breast cancer used in pathological examinations (30). Notably, the results of the present
study demonstrated that there was a significant association between
rs3842642 polymorphism and ER expression in breast cancer tissues
(P=0.014); however, there was no association between PR or ERBB2
and breast cancer. The proportion of 9/9 homozygous deletion in the
high expression group was increased compared with that of the low
expression group. The rs3842642 site of miR-205 should be
investigated further in a larger sample of patients; with more
in-depth study, this site may become a new molecular marker for the
diagnosis of breast cancer.
The expression of miR-205 was detected at the
cellular level using RT-qPCR, however, this method was not suitable
to detect the expression of miR-205 in paraffin-embedded tissues.
In-situ hybridization was adopted to quantify the expression
of miR-205 in breast cancer tissues. In the present study, more and
less metastatic breast cancer cells were selected for cells grown
on coverslips. Cells were collected and fixed and the expression of
miR-205 was detected using in-situ hybridization.
Subsequently, the expression of miR-205 in various cancer cells was
determined by RT-qPCR. The results of RT-qPCR were consistent with
those of in-situ hybridization, further confirming the
feasibility and creditability of in-situ hybridization to
determine the expression of miR-205.
In conclusion, the present study identified the
differential expression of miR-205 in breast cancer cells at the
cellular level. Decreased expression of miR-205 was associated with
metastatic breast cancer and rs3842642 was identified in various
tumor cells. The increase in the missing number of AGC repeats at
these sites in breast cancer cells was accompanied by an increased
expression of miR-205, however this has not been verified in breast
cancer specimens. A significant association was identified between
genotypes of rs3842642 and ER+ groups in breast cancer tissue
specimens, although no significant association was identified
between PR and ERBB2 positive groups and breast cancer/breast
benign lesions groups. The present study identified an association
between SNP variation of miR-205 and breast cancer, however the
association between miR-205 rs3842642 and SNP variation at other
sites in breast cancer should be investigated further.
Acknowledgements
The present study was supported by the Natural
Science Foundation of Zhejiang (grant no. LY15H160049) and Zhejiang
Province Public Technology Application Research Program (grant no.
2016F81SA700015).
References
|
1
|
Venter JC, Adams MD, Myers EW, Li PW,
Mural RJ, Sutton GG, Smith HO, Yandell M, Evans CA, Holt RA, et al:
The sequence of the human genome. Science. 291:1304–1351. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jupe ER, Badgett AA, Neas BR, Craft MA,
Mitchell DS, Resta R, Mulvihill JJ, Aston CE and Thompson LF:
Single nucleotide polymorphism in prohibitin 3′ untranslated region
and breast-cancer susceptibility. Lancet. 357:1588–1589. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gudmundsdottir ET, Barkardottir RB, Arason
A, Gunnarsson H, Amundadottir LT, Agnarsson BA, Johannsson OT and
Reynisdottir I: The risk allele of SNP rs3803662 and the mRNA level
of its closest genes TOX3 and LOC643714 predict adverse outcome for
breast cancer patients. BMC Cancer. 12:6212012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sun K and Lai EC: Adult-specific functions
of animal microRNAs. Nat Rev Genet. 14:535–548. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mishra PJ, Mishra PJ, Banerjee D and
Bertino JR: MiRSNPs or MiR-polymorphisms, new players in microRNA
mediated regulation of the cell: Introducing microRNA
pharmacogenomics. Cell Cycle. 7:853–858. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu C, Gong Y, Sun A, Zhang Y, Zhang C,
Zhang W, Zhao G, Zou Y and Ge J: The human MTHFR rs4846049
polymorphism increases coronary heart disease risk through
modifying miRNA binding. Nutr Metab Cardiovasc Dis. 23:693–698.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xu Y, Ma H, Yu H, Liu Z, Wang LE, Tan D,
Muddasani R, Lu V, Ajani JA, Wang Y and Wei Q: The miR-184
binding-site rs8126 T>C polymorphism in TNFAIP2 is associated
with risk of gastric cancer. PLoS One. 8:e649732013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guo Z, Wang H, Li Y, Li B, Li C and Ding
C: A microRNA-related single nucleotide polymorphism of the XPO5
gene is associated with survival of small cell lung cancer
patients. Biomed Rep. 1:545–548. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang N, Huo Q, Wang X, Chen X, Long L,
Jiang L, Ma T and Yang Q: A genetic variant in pre-miR-27a is
associated with a reduced breast cancer risk in younger Chinese
population. Gene. 529:125–130. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gilam A, Edry L, Mamluk-Morag E, Bar-Ilan
D, Avivi C, Golan D, Laitman Y, Barshack I, Friedman E and Shomron
N: Involvement of IGF-1R regulation by miR-515-5p modifies breast
cancer risk among BRCA1 carriers. Breast Cancer Res Treat.
138:753–760. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Linhares JJ, Azevedo M Jr, Siufi AA, de
Carvalho CV, Mdel C Wolgien, Noronha EC, Bonetti TC and da Silva
ID: Evaluation of single nucleotide polymorphisms in microRNAs
(hsa-miR-196a2 rs11614913 C/T) from Brazilian women with breast
cancer. BMC Med Genet. 13:1192012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Smith RA, Jedlinski DJ, Gabrovska PN,
Weinstein SR, Haupt L and Griffiths LR: A genetic variant located
in miR-423 is associated with reduced breast cancer risk. Cancer
Genomics Proteomics. 9:115–118. 2012.PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Karolchik D, Barber GP, Casper J, Clawson
H, Cline MS, Diekhans M, Dreszer TR, Fujita PA, Guruvadoo L,
Haeussler M, et al: The UCSC genome browser database: 2014 update.
Nucleic Acids Res. 42(Database Issue): D764–D670. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mundy GR: Metastasis to bone: Causes,
consequences and therapeutic opportunities. Nat Rev Cancer.
2:584–593. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hay ED: The mesenchymal cell, its role in
the embryo, and the remarkable signaling mechanisms that create it.
Dev Dyn. 233:706–720. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Brinckerhoff CE and Matrisian LM: Matrix
metalloproteinases: A tail of a frog that became a prince. Nat Rev
Mol Cell Biol. 3:207–214. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao X, Lu Y, Nie Y and Fan D: MicroRNAs
as critical regulators involved in regulating
epithelial-mesenchymal transition. Curr Cancer Drug Targets.
13:935–944. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ding XM: MicroRNAs: Regulators of cancer
metastasis and epithelial-mesenchymal transition (EMT). Chin J
Cancer. 33:140–147. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chao CH, Chang CC, Wu MJ, Ko HW, Wang D,
Hung MC, Yang JY and Chang CJ: MicroRNA-205 signaling regulates
mammary stem cell fate and tumorigenesis. J Clin Invest.
124:3093–3106. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Adachi R, Horiuchi S, Sakurazawa Y,
Hasegawa T, Sato K and Sakamaki T: ErbB2 down-regulates
microRNA-205 in breast cancer. Biochem Biophys Res Commun.
411:804–808. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Orang AV, Safaralizadeh R and Feizi MA
Hosseinpour: Insights into the diverse roles of miR-205 in human
cancers. Asian Pac J Cancer Prev. 15:577–583. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Z, Liao H, Deng Z, Yang P, Du N,
Zhanng Y and Ren H: miRNA-205 affects infiltration and metastasis
of breast cancer. Biochem Biophys Res Commun. 441:139–143. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu J, Mao Q, Liu Y, Hao X, Zhang S and
Zhang J: Analysis of miR-205 and miR-155 expression in the blood of
breast cancer patients. Chin J Cancer Res. 25:46–54.
2013.PubMed/NCBI
|
|
25
|
Khan S, Greco D, Michailidou K, Milne RL,
Muranen TA, Heikkinen T, Aaltonen K, Dennis J, Bolla MK, Liu J, et
al: MicroRNA Related Polymorphisms and Breast Cancer Risk. Plos
One. 9:e109973. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Michailidou K, Hall P, Gonzalez-Neira A,
Ghoussaini M, Dennis J, Milne RL, Schmidt MK, Chang-Claude J,
Bojesen SE, Bolla MK, et al: Large-scale genotyping identifies 41
new loci associated with breast cancer risk. Nat Genet. 45:353–361.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang L, Liu W, Jiang W, Lin J, Jiang Y, Li
B and Pang D: A miRNA binding site single-nucleotide polymorphism
in the 3′-UTR region of the IL23R gene is associated with breast
cancer. PLoS One. 7:e498232012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zheng H, Song F, Zhang L, Yang D, Ji P,
Wang Y, Almeida M, Calin GA, Hao X, Wei Q, et al: Genetic variants
at the miR-124 binding site on the cytoskeleton-organizing IQGAP1
gene confer differential predisposition to breast cancer. Int J
Oncol. 38:1153–1161. 2011.PubMed/NCBI
|
|
29
|
Wu H, Zhu S and Mo YY: Suppression of cell
growth and invasion by miR-205 in breast cancer. Cell Res.
19:439–448. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bartsch R, Ziebermayr R, Zielinski CC and
Steger GG: Triple-negative breast cancer. Wien Med Wochenschr.
160:174–181. 2010. View Article : Google Scholar : PubMed/NCBI
|