Introduction
Osteosarcoma (OS), the most common primary malignant
bone tumor in children and adolescents, generally originates from
the metaphysis of the long bone and has a high propensity for local
invasion and distant metastases, most commonly the lung (1–3). In spite
of aggressive surgery, intensive chemotherapy and multidisciplinary
treatment, the survival rate of OS has remained constant over the
last few decades (4–6). The long-term survival rate of patients
with metastasis or recurrence remains at <20% (7). The metastatic biology of OS is now
beginning to be understood, which holds promise for the development
of novel strategies to improve survival rates in patients with
OS.
The p21-activated kinases (PAKs) are a family of
Rac-α serine/threonine protein kinase (Rac)/cell division cycle 42
(Cdc42)-associated serine/threonine protein kinases that are
characterized by a highly-conserved N-terminal
Cdc42/Rac-interactive binding domain and a C-terminal kinase domain
(8). PAKs serve important roles in
regulating a variety of cellular functions, including cytoskeletal
dynamics (9), morphology, cell
survival (10), migration (11) and mitosis (12). The six human PAK isoforms are
classified into two families on the basis of their amino acid
sequence and function: Group I PAKs (PAK1, PAK2 and PAK3) exhibit
between 80 and 90% sequence identity within their catalytic
domains, whereas group II PAKs (PAK4, PAK5 and PAK6) exhibit
between 40 and 50% sequence identity with the kinase domains of the
group I PAKs (13).
PAK5, the most recently discovered PAK family
member, exhibits unique characteristics that differentiate it from
the other PAKs, and its expression was primarily identified in
neuronal tissues (14). Unlike PAK1,
PAK5 has no ability to complement serine/threonine protein kinase
STE20 function, and its high basal activity is not regulated by
GTPases, although it is able to bind to Cdc42 in preference to Rac
(15). Furthermore, PAK5 localizes to
the mitochondria, protects cells from pro-apoptotic stimuli by
phosphorylating B-cell lymphoma 2-associated death promoter (Bad)
on Ser-112 and inhibits apoptosis by preventing Bad from entering
the mitochondria (15). PAK5 is
reportedly overexpressed in pancreatic cancer (16), colorectal cancer (17), gastric cancer (18), breast cancer (19) and glioma (20), and its potential ability to promote
tumor behavior in carcinogenesis has been investigated. However,
the underlying molecular mechanisms of PAK5 in OS progression
remain unclear.
In the present study, we identified that PAK5
expression was upregulated in OS tissue and OS cell lines. PAK5
expression was significantly increased in OS patients with
metastasis. Therefore, the effect and underlying molecular
mechanism of PAK5 in OS progression was investigated. The results
of the present study suggested that PAK5 knockdown is able to
inhibit cell migration and invasion in human OS cell lines. Matrix
metalloproteinase 2 (MMP2), a downstream signaling molecule, was
identified to participate in the regulation of the process.
Therefore, it is proposed that PAK5 may serve an essential role in
OS development and that PAK5 is a potential therapeutic target.
Materials and methods
Tissue samples of patients with
OS
The present study was a retrospective analysis.
Owing to loss to follow-up, 2 years was selected as the survival
duration. A total of 65 patients with OS who underwent surgery were
enrolled. Among them, 13 exhibited lung metastasis. Primary tumor
tissue and pulmonary metastasis tissue were collected to analyze
the expression of PAK5 and MMP2 at the mRNA and protein levels
using the reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) and immunohistochemistry (IHC). The samples were
collected from Shanghai Jiao Tong University Affiliated Sixth
People's Hospital (Shanghai, China) from January 2009 to January
2012. The patients' clinical characteristics included age at
diagnosis, sex, tumor size, clinical stage, pathological type,
necrosis rate and metastasis. The present study was conducted
following approval from the Ethics Committee of Shanghai Jiao Tong
University Affiliated Sixth People's Hospital and written informed
consent was provided by all patients.
Immunohistochemistry
Primary osteosarcoma tissue and pulmonary metastasis
tissue, which were paraffin-embedded, were sliced into 4-µm
sections. Paraffin-embedded tissue slides were heated for 45 min at
60°C, deparaffinized in xylene, rehydrated in alcohol gradient and
then non-specifically blocked using H2O2 for
10 min and non-specific serum (Maixin-Bio, Fuzhou, China) for 15
min at room temperature subsequent to antigens being exposed with
citric acid buffer (Maixin-Bio) (pH 6.0; 0.1 M) and heated by
microwave for 10 min. The paraffin sections were incubated with
primary antibody against PAK5 (catalog no., ab110069) and MMP2
(catalog no., ab37150) (dilution, 1:50; Abcam, Cambridge, UK).
Rabbit IgG (dilution, 1:100; catalog no., ab172730, Abcam) was used
as a negative control. Immunohistochemical scoring was performed
without prior knowledge of the clinical response. Tissue sections
were observed using an AX10-Imager A1 light microscope (Zeiss GmbH,
Jena, Germany), and all images were captured using AxioVision
microscopy software (version 4.7; Zeiss GmbH). The tissue sections
were then manually scored semi-quantitatively for cytoplasmic
staining. The positive particles appeared brown in the cytoplasm.
The dominant staining intensity in tumor cells was scored as: 0,
0%; 1, between 1 and 25%; 2, between 26 and 50%; 3, between 51 and
75%; and 4, between 76 and 100%.
Cell culture
The human OS cell lines Saos-2 and MG63 were
obtained from the American Type Culture Collection (Manassas, VA,
USA). Cells were cultured in Dulbecco's modified Eagle's medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and 100 units penicillin/streptomycin
(Invitrogen; Thermo Fisher Scientific, Inc.) and maintained at 37°C
in a humidified atmosphere containing 5% CO2.
Construction of recombinant lentivirus
and lentivirus infection
The cDNA sequence of PAK5 was obtained from
GenBank® (www.ncbi.nlm.nih.gov/genbank; accession number
NM_177990). The PAK5 short hairpin RNA (shRNA)
5′-CTAGCCGGGATTACCACCATGACAATTTCAAGAGAATTGTCATGGTGGTAATCCCGTTTTTTGGAATTAAT-3′)
that targets nucleotides 1,934–1,954 of the PAK5 mRNA was inserted
into the plasmid pFH-L (Shanghai Hollybio, Shanghai, China).
Non-silencing shRNA
(5′-CTAGCCCGGTTCTCCGAACGTGTCACGTATCTCGAGATACGTGACACGTTCGGAGAATTTTTTTAAT−3′)
was used as a control that does not target any genes in humans,
mice or rats as determined by NCBI Reference Sequence Database
(www.ncbi.nlm.nih.gov/refseq)
screening. The recombinant vector was confirmed by digestion
analysis using restriction endonucleases (NotI and
BamHI), and all inserted sequences were verified by DNA
sequencing by external company (Invitrogen; Thermo Fisher
Scientific, Inc.). The lentiviruses were generated by triple
transfection of 80% confluent human embryonic kidney HEK-293 cells
with PAK5 (or control) shRNA-expressing vector and the
virion-packaging elements (pVSVG-I and pCMVΔR8.92) using
Lipofectamine™ 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
The lentiviruses were harvested in serum-free medium after 3 days
and filtered through a 0.45-µm filter (EMD Millipore, Billerica,
MA, USA).
Saos-2 and MG63 cells were plated at
5×104 cells/well in 6-well plates with recombinant
lentivirus-encoding shRNA against PAK5 (Lv-shPAK5) at a
multiplicity of infection of 60 in serum-free DMEM at 37°C. After 2
h, serum-enriched DMEM was added to the cells, and the growth
medium was replaced after 24 h. At 3 days post-transfection,
reporter gene expression (green fluorescent protein) was examined
using fluorescence microscopy.
RT-qPCR
Total cellular RNA from Saos-2 and MG63 cells was
isolated using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) and amplified using an RT-qPCR kit
(Platinum® SYBR® Green qPCR SuperMix-UDG;
Applied Biosystems; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. A 1 µg sample of total RNA was used
for first-strand cDNA synthesis using Moloney murine leukemia virus
reverse transcriptase at the following temperature protocol: 5 min
at 25°C; 60 min at 42°C; and 5 min at 70°C. The primer sequences
used for PAK5 were 5′-GGCGTCCTCTTGTGTCTTC-3′ (forward) and
5′-GTACTGAGTCCTTCTGATTTGC-3′ (reverse). The primer sequences for
MMP2 were: 5′-CTTCTTCCCTCGCAAGCC-3′ (forward) and
5′-ATGGATTCGAGAAAACCG-3′ (reverse). β-actin was used as the
internal control and amplified with the primers
5′-GTGGACATCCGCAAAGAC-3′ (forward) and 5′-AAAGGGTGTAACGCAACTA-3′
(reverse). qPCR was conducted using the CFX Connect™ Real-Time PCR
Detection system (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
The qPCR conditions were as follows: 2 min at 50°C; 10 min at 95°C;
35 cycles of 10 sec at 95°C; and 1 min at 60°C. All reactions were
performed in triplicate. Relative expression levels of PAK5 in
non-transduced and transduced Saos-2 and MG63 cells were determined
using the 2−ΔΔCq method by Microsoft Excel 2007
(21).
Western blot analysis
The non-transduced and transduced Saos-2 and MG63
cells were lysed in 50 µl lysis buffer (100 mM Tris-HCl, pH 6.8, 10
mM EDTA, 4% SDS and 10% glycine) on ice for 15 min. The lysates
were clarified by centrifugation at 12,000 × g for 30 min at 4°C.
The supernatants were collected and protein concentrations were
determined using the bicinchoninic acid method. Equal amounts (30
µg protein) of lysate were subjected to SDS-PAGE (12% gels) prior
to being transferred onto a polyvinylidene fluoride membrane using
an electroblotting apparatus. The membrane was blocked for 1 h at
room temperature with 5% non-fat dried milk in TBS and Tween-20
(TBST) solution and incubated at 4°C overnight with anti-PAK5
(catalog no., ab110069; Abcam), anti-MMP2 (catalog no., ab37150;
Abcam), anti-epithelial (E-)cadherin (catalog no., ab40772; Abcam),
anti-vimentin (catalog no., ab92547; Abcam) and anti-fibronectin
(catalog no., ab2413; Abcam) antibody. Following three washes with
TBST solution, the membrane was incubated with anti-rabbit
horseradish peroxidase-conjugated secondary antibody (dilution,
1:5,000; catalog no., sc-2004; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) at room temperature for 2 h. The signals of the
detected proteins were visualized on an Electrochemiluminescence
Plus Western Blotting Detection system (GE Healthcare Life
Sciences, Chalfont, UK). GAPDH protein levels were used as a
control.
Scratch wound healing assay
Cells were grown in 6-well tissue culture dishes
until reaching confluence. Cultures were incubated for 10 min with
PBS buffer. A scrape was made through the confluent monolayer with
a 1-mm diameter plastic pipette tip. Subsequently, dishes were
washed twice and incubated at 37°C in fresh DMEM (Gibco; Thermo
Fisher Scientific, Inc.) containing 10% FBS. At the bottom of each
dish, two arbitrary places were marked where the width of the wound
was measured using an IX71 inverted microscope (×4 objective)
(Olympus Corporation, Tokyo, Japan). Wound healing was expressed as
the mean ± standard error of the mean (SEM) at 0 and 24 h.
Cell migration assay
Transwell migration assays were performed as
previously described (22). Assays
were performed using 8-µm Transwell inserts (EMD Millipore).
Migrated cells were fixed for 30 min with 4% paraformaldehyde and
stained with 0.1% crystal violet for 20 min. The assays were
monitored over a 24-h period. Images of five random fields were
captured using an IX71 inverted microscope. The cells were counted
in ten random fields using high power at ×200 magnification. All
experiments were performed in triplicate.
In vivo study
Male BALB/c nude mice (n=3 per group, between 5 and
6 weeks of age) weighing ~20 g each were purchased from Shanghai
Rubicam Laboratory Animal Ltd. (Shanghai, China) and housed in a
controlled environment at 22°C with a relative humidity of 65% and
a 12-h light/12-h dark cycle. All mice had free access to water and
food. Non-transduced and transduced Saos-2 cells at ~92% viability
by trypan blue stain assay were subcutaneously injected into the
right flank of each mouse (2×107 cells/mouse). Solid
tumor volumes (in mm3) were determined using digital
Vernier caliper measurements and the formula a × b2
×0.5, where a is the longest diameter of the tumor and b is the
shortest diameter of the tumor. Following tumor growth for 40 days,
the mice were sacrificed and the tumors were excised. All animal
procedures were approved by the Animal Ethics Committee of Shanghai
Jiao Tong University Affiliated Sixth People's Hospital.
Statistical analysis
For statistical analysis, SPSS software (version
13.0; SPSS Inc., Chicago, IL, USA) was used. All results are
presented as the mean ± standard deviation All results were
analyzed using one-way analysis of variance to compare the
measurement data with heterogeneity of variance or enumeration
data. The Kaplan-Meier estimator method was used to analyze
survival. P<0.05 was considered to indicate a statistically
significant difference.
Results
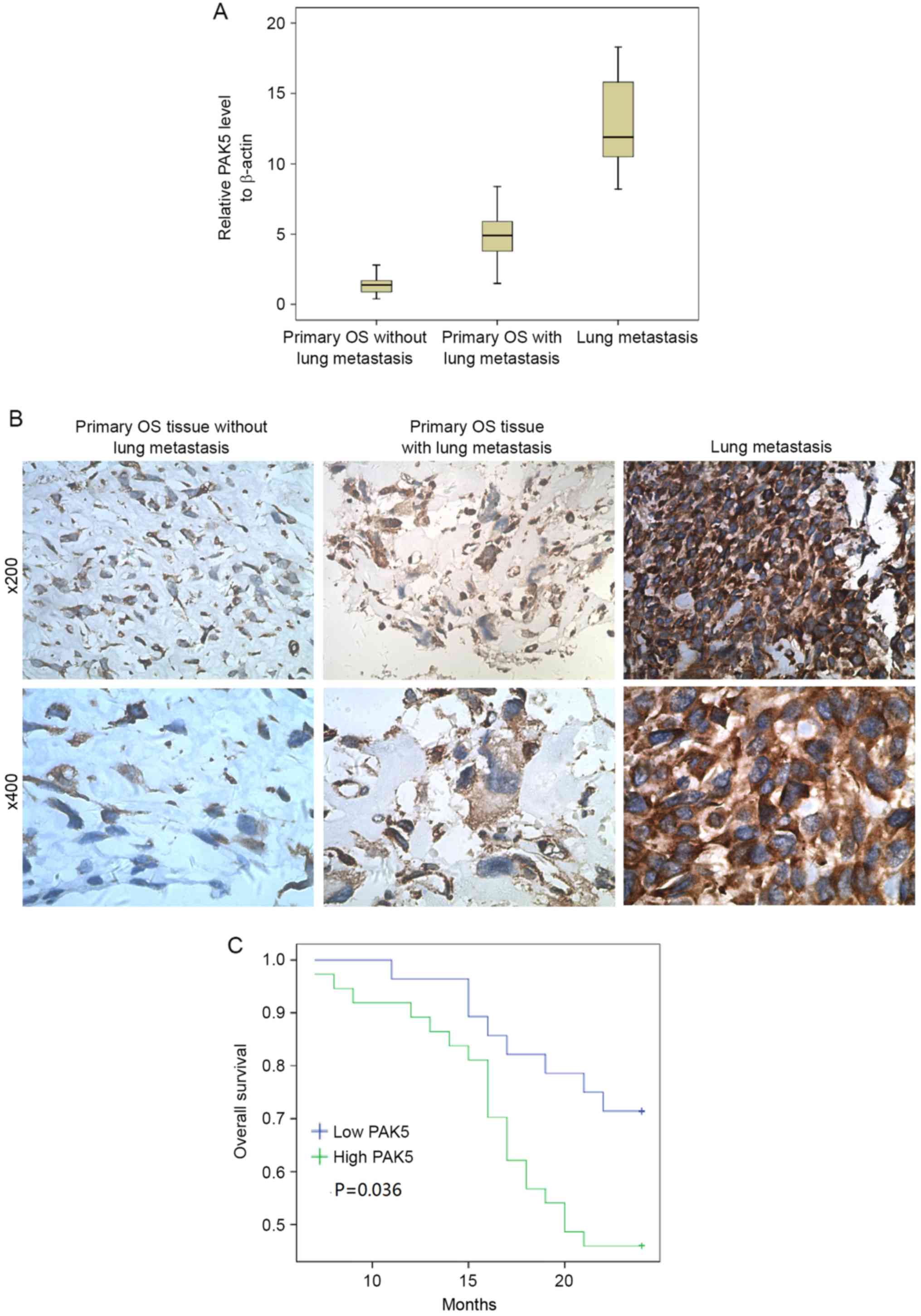
PAK5 overexpression during OS
progression
Aberrant PAK5 signaling is integral for breast
cancer (23), ovarian cancer
(24) and hepatocellular carcinoma
cells (25). In the present study,
primary samples from 65 patients with OS and 13 pulmonary
metastasis samples were analyzed for PAK5 expression using RT-qPCR
and IHC (Fig. 1A and B). The
clinicopathological characteristics of 65 patients are presented in
Table I. PAK5 expression was
identified to be significantly associated with clinical stage
(P=0.035), tumor cell necrosis rate (P=0.035) and metastasis
(P=0.010). No significant differences were identified for other
clinical features including sex, age, tumor location and
pathological type between individuals with high and low PAK5
expression. Fig. 1A and B indicate
that the greatest increase in PAK5 expression occurred in lung
metastasis tissue compared with the other tissues. PAK5 expression
in tissue from patients with OS with lung metastasis was increased
compared with those without lung metastasis. Furthermore, the
prognostic significance of PAK5 expression was assessed in 65
patients with OS with clinical follow-up records. Kaplan-Meier
estimator analysis of survival demonstrated that the high PAK5
group tended to have significantly shorter overall survival times
(Fig. 1C) compared with the low PAK5
group (P=0.036).
 | Table I.Analysis of PAK5 expression and the
clinical characteristics of patients with osteosarcoma. |
Table I.
Analysis of PAK5 expression and the
clinical characteristics of patients with osteosarcoma.
|
Characteristics | Low level of
PAK5 | High level of
PAK5 | P-value |
|---|
| Age, years |
|
| 0.235 |
|
<15 | 15 | 24 |
|
|
≥15 | 13 | 13 |
|
| Sex |
|
| 0.590 |
|
Male | 14 | 27 |
|
|
Female | 14 | 10 |
|
| Anatomical
location |
|
| 0.460 |
|
Tibia/femur | 17 | 19 |
|
|
Elsewhere | 11 | 18 |
|
| Tumor size, cm |
|
| 0.095 |
| ≤8 | 18 | 16 |
|
|
>8 | 10 | 21 |
|
| Clinical stage |
|
| 0.035 |
|
IIA | 18 | 14 |
|
|
IIB/III | 10 | 23 |
|
| Necrosis rate,
% |
|
| 0.035 |
|
≤25 | 9 | 19 |
|
|
>25 | 19 | 18 |
|
| Pathological
type |
|
| 0.863 |
|
Conventional | 18 | 23 |
|
|
Non-conventional | 10 | 14 |
|
| Metastasis |
|
| 0.010 |
|
Negative | 23 | 19 |
|
|
Positive | 5 | 18 |
|
| PAK5 score | 34 | 130 | <0.001 |
Knockdown of PAK5 inhibits OS cell
migration ability and decreases the expression of MMP2
The results of the present study indicated that the
PAK5 expression level was associated with metastasis. In order to
investigate this association further, PAK5 was knocked down to
determine its function. It was identified that PAK5 shRNA
effectively knocked down PAK5 expression at the RNA and protein
level. The inhibition ratio was >75%. Furthermore, certain
factors, which were associated with cell migration, including MMP2,
E-cadherin, vimentin and fibronectin, were identified. Only MMP2
expression was identified to be significantly decreased at the RNA
and protein levels (Fig. 2A and B).
Migration is the most important metastasis-associated process. It
was investigated whether PAK5 knockdown was able to decrease the
migration capability of OS cells using an in vitro scratch
wound healing assay and a Transwell migration assay. The results
identified that the PAK5 shRNA group markedly decreased the
migratory ability (Fig. 2C and D),
indicating that PAK5 is a potential migration regulator.
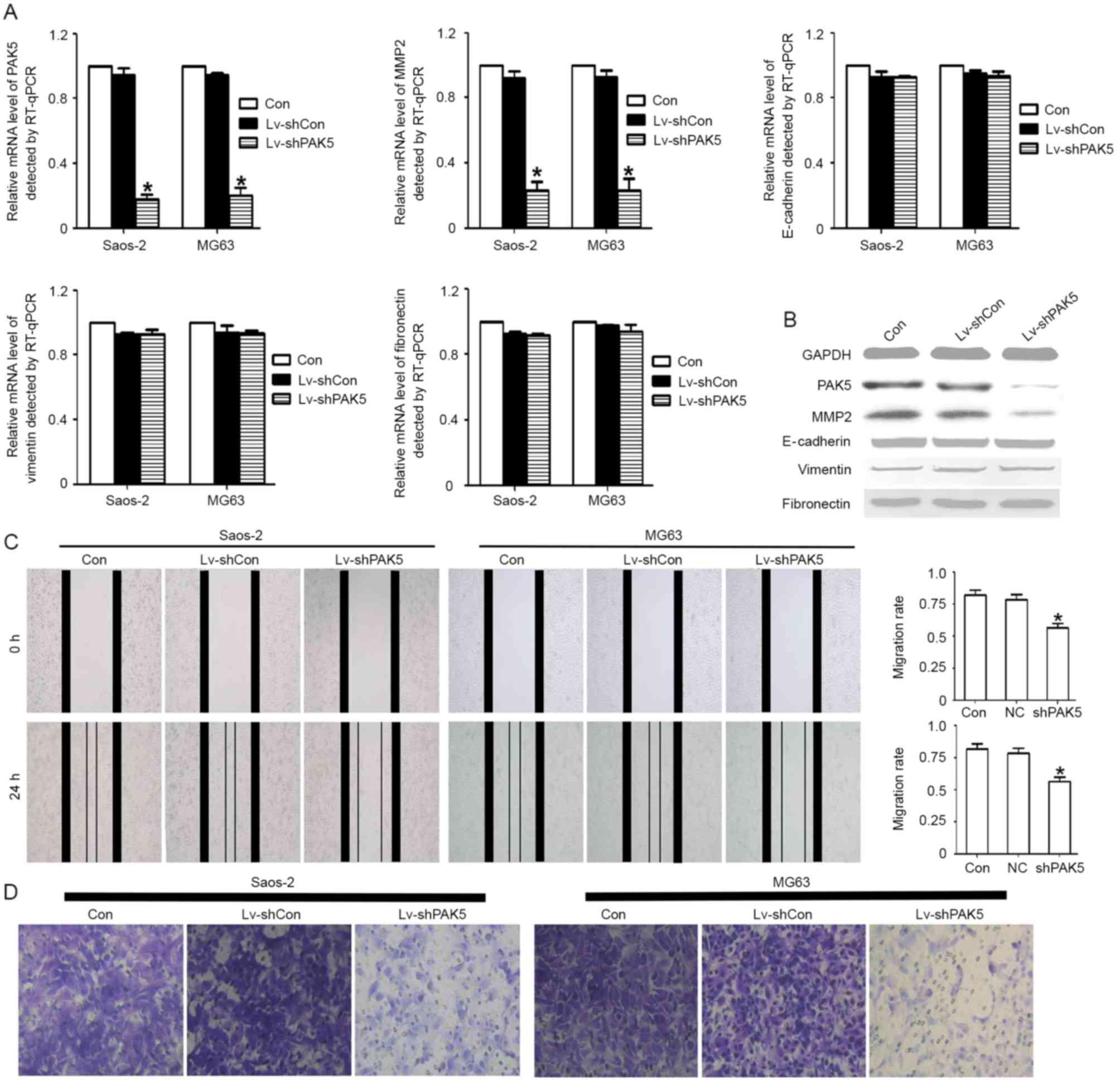 | Figure 2.Association between PAK5 and
osteosarcoma cell migratory ability. (A) The PAK5 gene was knocked
down using Lv-shPAK5 in Saos2 and MG63 cells. The mRNA level of
PAK5 and MMP2 was decreased significantly (>75%) in the two cell
lines (*P<0.05). No statistically significant difference was
identified for E-cadherin, vimentin or fibronectin levels. (B)
Western blotting results indicated a marked difference for PAK5 and
MMP2 protein expression among these groups. No significant
differences were identified for E-cadherin, vimentin and
fibronectin. (C) A scratch wound healing assay identified that
Lv-shPAK5-transfected Saos2 and MG63 cells exhibited a
significantly decreased wound healing rate compared with untreated
and Lv-shCon cells. Results are expressed as the mean ± standard
deviation. *P<0.05. (D) An in vitro Transwell migration
assay identified that PAK5 knockdown attenuated cell migration. The
untreated, Lv-shCon and Lv-shPAK5 cells were seeded in the upper
Transwell chambers and incubated for 24 h. Images were captured
using a light microscope at ×200 magnification of the cells that
had migrated into the lower chamber. PAK5, p21-associated kinase 5;
Lv-shPAK5, lentivirus-transduced short hairpin RNA targeting PAK5;
MMP2, matrix metalloproteinase 2; E-cadherin, epithelial cadherin;
Lv-shCon/NC, lentivirus-transduced negative control short hairpin
RNA; RT-qPCR, reverse transcription-quantitative polymerase chain
reaction; Con, control. |
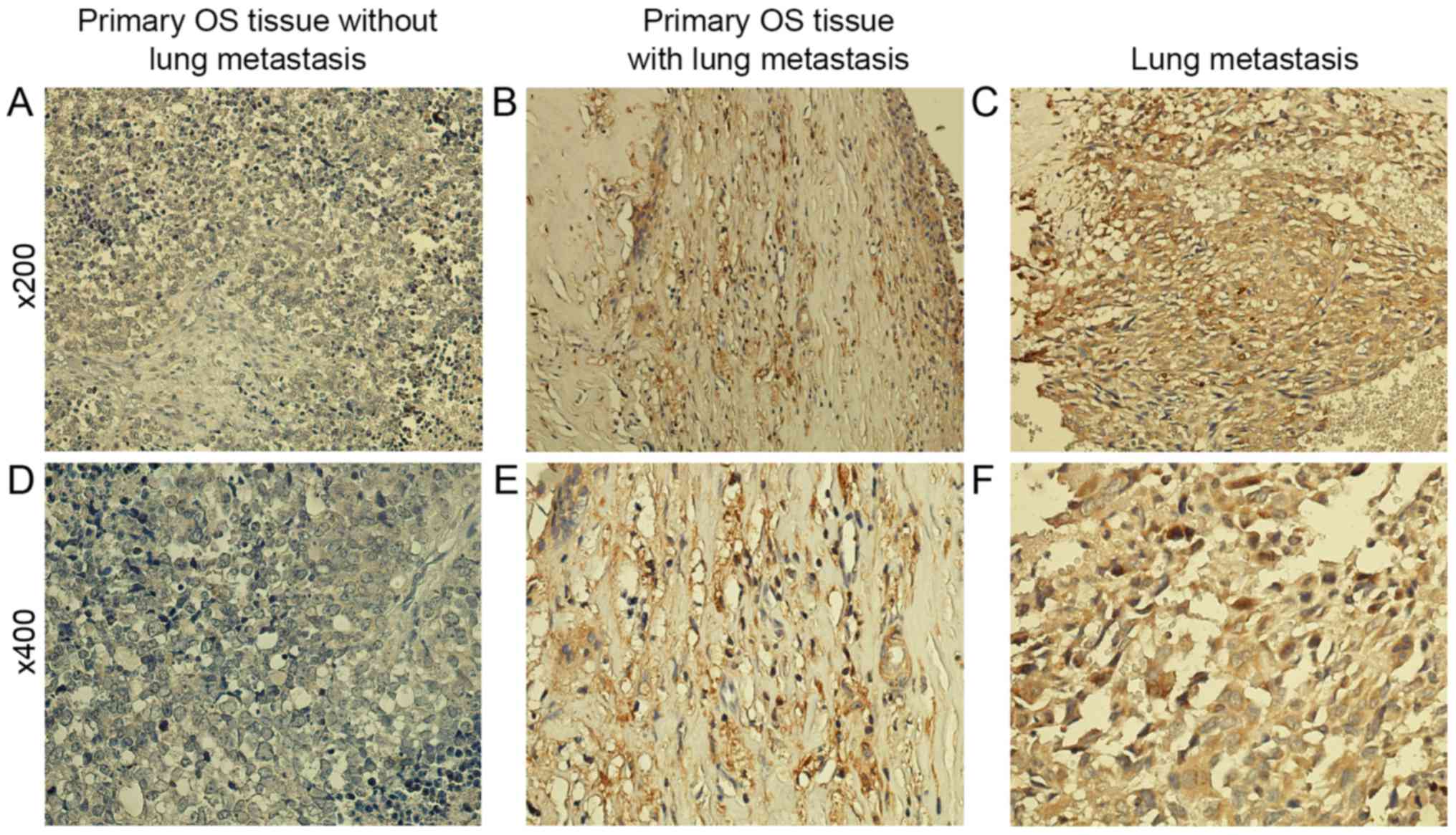
MMP2 overexpression in lung
metastasis
The PAK5-early growth response protein 1 (Egr1)-MMP2
signaling pathway contributes to the migration ability in breast
cancer and glioma. Therefore, it was analyzed whether MMP2 and PAK5
exhibited the same expression tendencies in OS tissue using IHC,
and it was identified that lung metastasis tissue exhibited
increased MMP2 expression compared with primary OS tissue (Fig. 3). A marked association was identified
between PAK5 and MMP2 expression with a correlation coefficient of
0.821 by Pearson's rank correlation analysis.
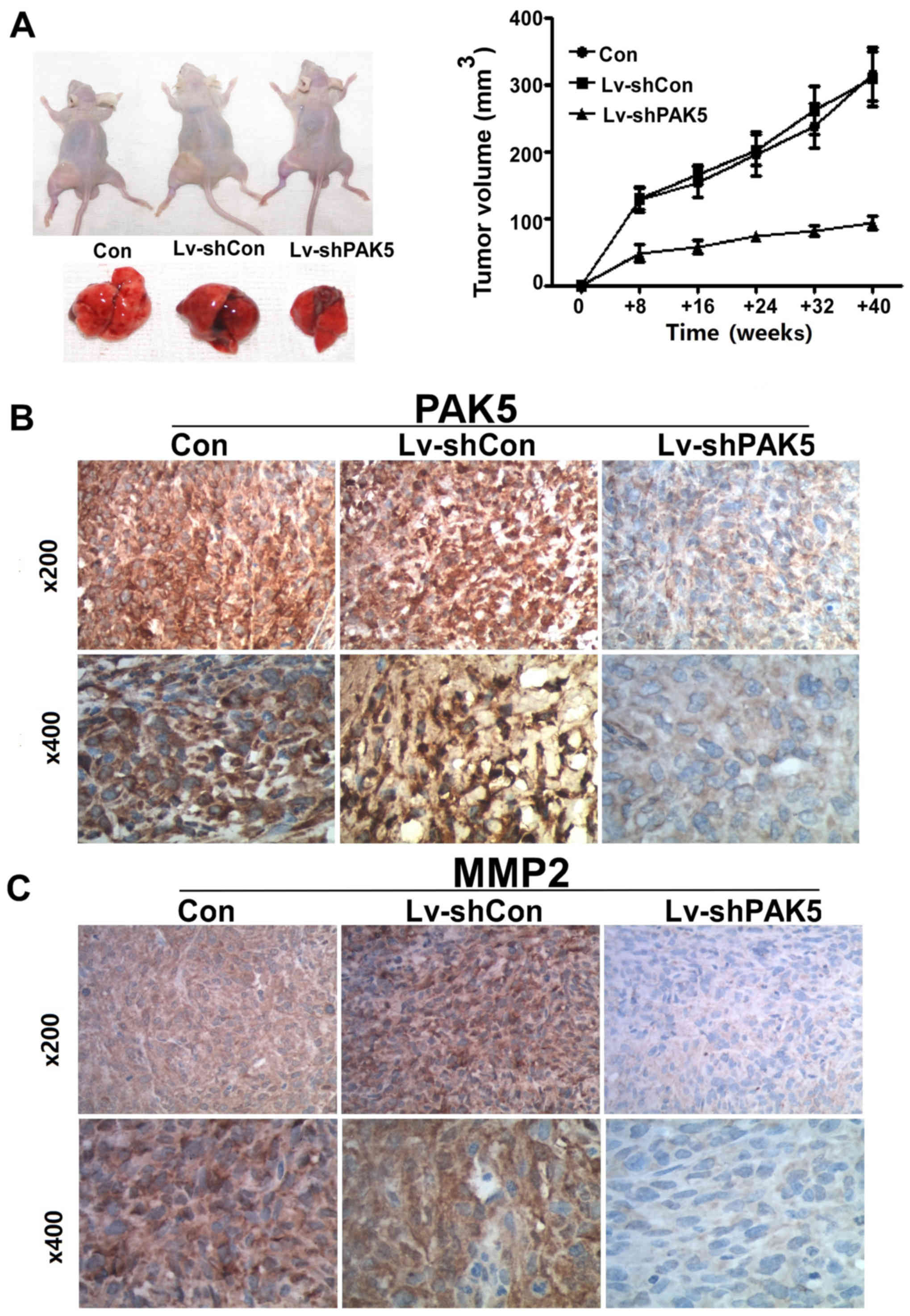
PAK5 silencing decreases xenograft
growth
The growth rate of subcutaneous xenografts in the
PAK5 shRNA group was significantly increased compared with that in
the non-transduced MG63 cells. At the fourth week, tumor volume and
weight were significantly increased in the two control groups
compared with those in the PAK shRNA group (volume, 315.67±69.15
and 308.67±71.43 vs. 93.33±16.1 mm3; P=0.015; Fig. 4A). PAK5 and MMP2 staining was more
intense in the two control group xenografts compared with that in
the treated group (Fig. 4B and
C).
Discussion
PAKs are key effectors of the Rho family GTPases
that are divided into group I PAKs (PAK1, PAK2 and PAK3) and group
II PAKs (PAK4, PAK5 and PAK6). PAK5 exhibits sequence identity with
other PAKs in the p21 GTPase-binding and kinase domains. However,
it is completely distinct in other regions compared with other PAKs
(26). It has been demonstrated that
PAK5 functions as a dynamic signaling node in cancer and is central
to a variety of cellular functions, including proliferation,
apoptosis and migration. PAK5 overexpression has been observed in a
number of tumors (9–12,16–20).
Metastasis due to tumor progression is the primary cause of the
majority of cancer-related mortality, particularly in patients with
OS (27). In total, <80% of
patients with OS exhibit lung metastasis. OS has increased local
aggressiveness, and occasionally migrates to the lung and other
bone sites with uncommon lymphoid metastasis (28–31). The
5-year overall survival rate in patients with OS and metastases is
between 10 and 30% (32,33). However, the underlying molecular
mechanisms of metastasis in OS remain to be further elucidated.
Previous research has focused on what facilitates OS cell
metastasis (34–36). The focus of the present study was on
PAK5, which decreases cell adhesion by promoting cytoskeletal
reorganization including filopodia formation and neurite outgrowth
(37). The association between PAK5
expression and the outcomes of 65 cases of primary OS tissues was
analyzed using RT-qPCR and IHC. A total of 13 lung metastasis
tissues were also examined. PAK5 levels were increased in lung
metastasis tissues compared with in primary OS tissue. Almost all
of the samples exhibited intense positive tan particles with large
amounts of cytoplasm. The primary OS tissues were divided into two
groups: Low PAK5 and high PAK5. Kaplan-Meier estimator analysis of
survival indicated that increased PAK5 expression was associated
with a shorter survival time of the patients. No significant
differences were identified in certain clinical features including
sex, age, tumor location and pathological type between individuals
with high and low PAK5 expression. PAK5 expression is associated
with clinical stage, necrosis rate and metastasis. In order to
investigate the function of PAK5, shRNA was used as an effective
way to silence gene expression. The shRNA was able to inhibit PAK5
expression by >75% at the mRNA and protein levels. The
downstream factors of PAK5 were also examined. A previous study
identified that PAK5 was able to promote metastasis by regulating
the Egr1-MMP2 signaling pathway (38). The results of the present study also
clarified that, following PAK5 silencing, MMP2, but not E-cadherin,
vimentin or fibronectin, was downregulated. MMP2 has been
identified as a key extracellular matrix factor that participates
in cell migration associated with tumor metastasis (39). PAK5 shRNA decreases the migration
ability of OS cells associated with decreased MMP2. The results of
the present study also identified that PAK5 serves an important
role in the migration capacity of OS cells via MMP2.
The effects of PAK5 were also investigated in
vivo. Following subcutaneous injection of MG63 cells with or
without PAK5 shRNA transduction in nude mice bearing OS xenografts,
the PAK5 shRNA exhibited superior antitumor efficacy in vivo
compared with the two control groups. It is noteworthy that the
in vivo PAK5 shRNA treatment led to a significant decrease
in tumor growth. Additionally, ex vivo histological analysis
revealed that the PAK5 shRNA group exhibited decreased expression
of PAK5 and MMP2 compared with the control groups.
The results of the present study indicate that PAK5
and MMP2 overexpression are directly associated with metastasis and
lead to poor prognosis. This retarded PAK5 shRNA-associated OS cell
growth in vitro and in vivo in a nude mouse model
indicated that PAK5 regulates MMP2 expression and inhibits tumor
metastasis. Therefore, the results of the present study indicate
that PAK5 is a potential prognostic indicator for human OS.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81172548 and
8137273).
References
|
1
|
Chi S, Conklin L, Qin J, Meyers P, Huvos
AG, Healey JH and Gorlick R: The patterns of relapse in
osteosarcoma: The Memorial Sloan-Kettering experience. Pediatr
Blood Cancer. 42:46–51. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Crompton BD, Goldsby RE, Weinberg VK,
Feren R, O'Donnell RJ and Ablin AR: Survival after recurrence of
osteosarcoma: A 20-year experience at a single institution. Pediatr
Blood Cancer. 47:255–259. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gelderblom H, Jinks RC, Sydes M, Bramwell
VH, van Glabbeke M, Grimer RJ, Hogendoorn PC, McTiernan A, Lewis
IJ, Nooij MA, et al: Survival after recurrent osteosarcoma: Data
from 3 European Osteosarcoma Intergroup (EOI) randomized controlled
trials. Eur J Cancer. 47:895–902. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hattinger CM, Pasello M, Ferrari S, Picci
P and Serra M: Emerging drugs for high-grade osteosarcoma. Expert
Opin Emerg Drugs. 15:615–634. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ray-Coquard I and Le Cesne A: A role for
maintenance therapy in managing sarcoma. Cancer Treat Rev.
38:368–378. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang J and Zhang W: New molecular insights
into osteosarcoma targeted therapy. Curr Opin Oncol. 25:398–406.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Luetke A, Meyers PA, Lewis I and Juergens
H: Osteosarcoma treatment-where do we stand? A state of the art
review. Cancer Treat Rev. 40:523–532. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sells MA and Chernoff J: Emerging from the
Pak: The p21-activated protein kinase family. Trends Cell Biol.
7:162–167. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Edwards DC, Sanders LC, Bokoch GM and Gill
GN: Activation of LIM-kinase by Pak1 couples Rac/Cdc42 GTPase
signalling to actin cytoskeletal dynamics. Nat Cell Biol.
1:253–259. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Marlin JW, Eaton A, Montano GT, Chang YW
and Jakobi R: Elevated p21-activated kinase 2 activity results in
anchorage independent growth and resistance to anticancer
drug-induced cell death. Neoplasia. 11:286–297. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Somanath PR, Vijai J, Kichina JV, Byzova T
and Kandel ES: The role of PAK-1 in activation of MAP kinase
cascade and oncogenic transformation by Akt. Oncogene.
28:2365–2369. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maroto B, Ye MB, von Lohneysen K,
Schnelzer A and Knaus UG: P21-activated kinase is required for
mitotic progression and regulates Plk1. Oncogene. 27:4900–4908.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jaffer ZM and Chernoff J: p21-activated
kinases: Three more join the Pak. Int J Biochem Cell Biol.
34:713–717. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dan C, Nath N, Liberto M and Minden A:
PAK5, a new brainspecific kinase, promotes neurite outgrowth in
N1E-115 cells. Mol Cell Biol. 22:567–577. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cotteret S, Jaffer ZM, Beeser A and
Chernoff J: p21-activated kinase 5 (Pak5) localizes to mitochondria
and inhibits apoptosis by phosphorylating BAD. Mol Cell Biol.
23:5526–5539. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Giroux V, Dagorn JC and Iovanna JL: A
review of kinases implicated in pancreatic cancer. Pancreatology.
9:738–754. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gong W, An Z, Wang Y, Pan X, Fang W, Jiang
B and Zhang H: P21-activated kinase 5 is overexpressed during
colorectal cancer progression and regulates colorectal carcinoma
cell adhesion and migration. Int J Cancer. 125:548–555. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gu J, Li K, Li M, Wu X, Zhang L, Ding Q,
Wu W, Yang J, Mu J, Wen H, et al: A role for p21-activated kinase 7
in the development of gastric cancer. FEBS J. 280:46–55. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang XX, Cheng Q, Zhang SN, Qian HY, Wu
JX, Tian H, Pei DS and Zheng JN: PAK5-Egr1-MMP2 signaling controls
the migration and invasion in breast cancer cell. Tumor Biol.
34:2721–2729. 2013. View Article : Google Scholar
|
|
20
|
Han ZX, Wang XX, Zhang SN, Wu JX, Qian HY,
Wen YY, Tian H, Pei DS and Zheng JN: Downregulation of PAK5
inhibits glioma cell migration and invasion potentially through the
PAK5-Egr1-MMP2 signaling pathway. Brain Tumor Pathol. 31:234–241.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lin F, Shen Z, Tang LN, Zheng SE, Sun YJ,
Min DL and Yao Y: KLF8 knockdown suppresses proliferation and
invasion in human osteosarcoma cells. Mol Med Rep. 9:1613–1617.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Y, Ke Q, Shao Y, Zhu G, Li Y, Geng N,
Jin F and Li F: GATA1 induces epithelial-mesenchymal transition in
breast cancer cells through PAK5 oncogenic signaling. Oncotarget.
6:4345–4356. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li D, Yao X and Zhang P: The
overexpression of P21-activated kinase 5 (PAK5) promotes
paclitaxel-chemoresistance of epithelial ovarian cancer. Mol Cell
Biochem. 383:191–199. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhai J, Qu S, Li X, Zhong J, Chen X, Qu Z
and Wu D: miR-129 suppresses tumor cell growth and invasion by
targeting PAK5 in hepatocellular carcinoma. Biochem Biophys Res
Commun. 464:161–167. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pandey A, Dan I, Kristiansen TZ, Watanabe
NM, Voldby J, Kajikawa E, Khosravi-Far R, Blagoev B and Mann M:
Cloning and characterization of PAK5, a novel member of mammalian
p21-activated kinase-II subfamily that is predominantly expressed
in brain. Oncogene. 21:3939–3948. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Farfalli GL, Albergo JI, Lobos PA, Smith
DE, Streitenberger PD, Rodríguez MG Pallotta and Aponte-Tinao LA:
Osteosarcoma lung metastases. Survival after chemotherapy and
surgery. Medicina (B Aires). 75:87–90. 2015.(In Spanish).
|
|
28
|
Poletajew S, Fus L and Wasiutynski A:
Current concepts onpathogenesis and biology of metastatic
osteosarcoma tumors. Ortop Traumatol Rehabil. 13:537–545. 2011.(In
English, Polish). View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Endo-Munoz L, Evdokiou A and Saunders NA:
The role of osteoclasts and tumour-associated macrophages in
osteosarcoma metastasis. Biochim Biophys Acta. 1826:434–442.
2012.PubMed/NCBI
|
|
30
|
Friedrich P, Ortiz R, Strait K, Fuentes S,
Gamboa Y, Arambú I, Ah-Chu-Sanchez M, London W, Rodríguez-Galindo C
and Antillón-Klussmann F: Pediatric sarcoma in Central America:
Outcomes, challenges, and plans for improvement. Cancer.
119:871–879. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rainusso N, Wang LL and Yustein JT: The
adolescent and oung adult with cancer: State of the art-bone
tumors. Curr Oncol Rep. 15:296–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hiller C, Wegler JL and Forest CP:
Osteosarcoma: Accurately diagnosing this bone-chilling disease.
JAAPA. 29:29–35. 2016.PubMed/NCBI
|
|
33
|
Meyers PA: Muramyl tripeptide
(mifamurtide) for the treatment of osteosarcoma. Expert Rev
Anticancer Ther. 9:1035–1049. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Seong BK, Lau J, Adderley T, Kee L,
Chaukos D, Pienkowska M, Malkin D, Thorner P and Irwin MS: SATB2
enhances migration and invasion in osteosarcoma by regulating genes
involved in cytoskeletal organization. Oncogene. 34:3582–3592.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lamora A, Talbot J, Bougras G, Amiaud J,
Leduc M, Chesneau J, Taurelle J, Stresing V, Le Deley MC, Heymann
MF, et al: Overexpression of smad7 blocks primary tumor growth and
lung metastasis development in osteosarcoma. Clin Cancer Res.
20:5097–5112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Luther GA, Lamplot J, Chen X, Rames R,
Wagner ER, Liu X, Parekh A, Huang E, Kim SH, Shen J, et al: IGFBP5
domains exert distinct inhibitory effects on the tumorigenicity and
metastasis of human osteosarcoma. Cancer Lett. 336:222–230. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wong LE, Reynolds AB, Dissanayaka NT and
Minden A: p120-catenin is a binding partner and substrate for Group
B Pak kinases. J Cell Biochem. 110:1244–1254. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Han ZX, Wang XX, Zhang SN, Wu JX, Qian HY,
Wen YY, Tian H, Pei DS and Zheng JN: Downregulation of PAK5
inhibits glioma cell migration and invasion potentially through the
PAK5-Egr1-MMP2 signaling pathway. Brain Tumor Pathol. 31:234–241.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu J, Ping W, Zu Y and Sun W:
Correlations of lysyl oxidase with MMP2/MMP9 expression and its
prognostic value in non-small cell lung cancer. Int J Clin Exp
Pathol. 7:6040–6047. 2014.PubMed/NCBI
|