Introduction
Renal cell carcinoma (RCC) is the most common
malignant neoplasms of the kidney (1). RCC represents 3–4% of adult solid tumors
globally, and metastasis is detected in ~30% of patients during
initial diagnosis due to the lack of symptoms typical of the
disease at the early stage (1).
Additionally, the 5-year survival of the patients with metastatic
RCC has been identified to be <10% (2). The major therapeutic tool for RCC is
surgery, though targeted therapy may be opted for in cases where
surgery does not prove effective. Therefore, attention has been
focused on the diagnosis and targeted treatment of RCC. As
previously described, RCC development is induced by metabolic
factors (3). Genes, including Von
Hippel-Lindau tumor suppressor, MET, folliculin, fumarate
hydratase, SDH, tuberous sclerosis (TSC) 1 and TSC2, are typically
mutated in RCC, all of which have fundamental roles in the
regulation of metabolic processes (3). This suggests that metabolic disorders
involved in energy and nutrient sensing are key factors in the
development of RCC, offering novel targets for tumor diagnosis and
treatment. Therefore, metabolic factors, which underlie RCC
development, may provide novel targets for screening and developing
more effective therapeutic strategies.
Nucleobindin 2 (NUCB2), as a metabolic factor, is a
neuropeptide that serves important roles in regulating food intake
and energy homeostasis (4). The
protein, NUCB2/nesfatin-1 was identified by Oh et al
(4) in 2006 as a satiety molecule
expressed in the hypothalamus. The NUCB2 gene is located on
chromosome 11, which consists of 14 exons spanning 54,785
nucleotides and codes a precursor 396-amino-acid protein NUCB2
(5,6).
NUCB2 was first reported to be expressed in the arcuate nucleus,
lateral hypothalamus, paraventricular nucleus and supraoptic
nucleus of the hypothalamus (4,5). Following
this, a number of studies have suggested that the primary
peripheral source of NUCB2 was originated from stomach mucosa, and
NUCB2 was also peripherally expressed in pancreas and fat tissues
(7,8).
In the hypothalamus, NUCB2 inhibits food intake and promotes body
metabolism (4). As previously
described, it has been report that the fasting plasma NUCB2 levels
were significantly decreased in patients with type 2 diabetes
compared with healthy or type 1 diabetes subjects (9). Additionally, plasma NUCB2 appears to
regulate glucose and fatty acid metabolism (10,11). NUCB2
also inhibits the apoptosis of pancreatic cells, which tends to
accelerate the development of metabolic disorders (12). In previous years, an increasing number
of studies have focused on the association between NUCB2 and
carcinoma development. Suzuki et al (13) suggested that NUCB2, as a tumor
promoter, served an important role in the process of development
and metastasis in breast carcinomas. Additionally, an association
between NUCB2 expression and biochemical recurrence and progression
of prostatic cancer has been reported (6,14,15). In a recent study, clear cell RCC
tissues were observed to express NUCB2 protein, which served a
critical function in tumorigenesis and tumor development (16).
However, there is a lack of studies investigating
the effects of NUCB2 protein on the invasion and apoptosis rates of
tumors, particularly in RCC. In the present study, NUCB2 expression
was first analyzed in various urological carcinoma cell lines, and
an in vitro study investigating the effect of NUCB2 on RCC
invasion and development was designed.
Materials and methods
Cell culture
All of the cell lines (293, 786-O, ACHN, PC-3 and
LNCaP) were purchased from the American Type Culture Collection
(Manassas, VA, USA) and maintained at −80°C with Cell Culture
Freezing medium (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
All the cell lines were cultured and maintained in RPMI-1640 medium
(HyClone; GE Healthcare Life Sciences, Logan, UT, USA), 10% fetal
bovine serum (Thermo Fisher Scientific, Inc.) and 50 µg/ml
streptomycin. All cells were cultured at 37°C in an atmosphere of
5% CO2 under saturated humidity. The 786-O cells were
divided into three groups: i) non-transfected control cells (NT
group); ii) β-actin (ACTB)-transfected cells (lenti-NC cells) and
iii) NUCB2-knocked down 786-O cells (lenti-NUCB2-KD cells).
Western blotting
The cells were lysed in ice-cold standard RIPA lysis
buffer (Beyotime Institute of Biotechnology, Haimen, China), and
the protein concentration was determined using a bicinchoninic
protein assay kit. In total, 50 µg of each protein was loaded per
lane and separated using SDS-PAGE with a 10% gel and then
transferred onto polyvinylidene fluoride membranes (EMD Millipore,
Billerica, MA, USA). Following blocking with 5% bovine serum
albumin (Solarbio, Beijing, China) for 1 h at 37°C, primary
antibodies against NUCB2 (1 µg/ml; cat. no. AF5949; R&D
Systems, Inc., Minneapolis, MN, USA) and GAPDH (1:3,000; cat. no.
8884; Cell Signaling Technology, Inc., Danvers, MA, USA) were used.
The secondary antibodies were peroxidase-conjugated immunoglobulin
G (rabbit anti-sheep, ZDR-5308; sheep anti-rabbit, ZDR-5306;
Beijing Zhongshan Jinqiao Biotechnology Co., Ltd., Beijing, China)
and were used at a dilution of 1:8,000. The intensity of bands was
analyzed using Quantity One software (version 4.6.2; Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Three replicates were
performed.
Quantitative reverse transcription
polymerase chain reaction (RT-qPCR)
The total RNA was extracted using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) according to the manufacturer's protocol. Following this, 20
µl RNase-free water was added into the RNA, and ultraviolet
analysis was used to measure the RNA content. RNA was then
subjected to RT to obtain cDNA for qPCR analysis. The sequences of
primers are summarized in Table I,
and the human NUCB2 and ACTB primers were designed according to the
sequences listed. Subsequent to the synthesis of the probes,
RT-qPCR was performed according to the manufacturer's protocol of
the PrimeScript™ RT-PCR kit (Takara Biotechnology Co., Ltd.,
Dalian, China). The qPCR was performed with the PCR mix containing
2X SYBR Green mix (Takara Biotechnology Co., Ltd.). The Cq value
was automatically recorded using the PCR machine. Thermal cycler
PCR conditions consisted of heating samples to 95°C for 2 min; 40
cycles of 95°C for 15 sec, 60°C for 20 sec, and 72°C for 20 sec;
and 72°C for 30 min. The relative expression levels of NUCB2 mRNA
were calculated using Quantity One software (version 4.6.2; Bio-Rad
Laboratories, Inc., Hercules, CA, USA) using the 2−ΔΔCq
method: ΔΔCq = (Cqtarget gene - Cqreference
gene)test group - (Cqtarget gene -
Cqreference gene)blank control group
(17). Three replicates were
performed and the HEK-293 cell line was used as the control
group.
 | Table I.List of primer sequences for
polymerase chain reaction. |
Table I.
List of primer sequences for
polymerase chain reaction.
| Target | Primer sequence
(5′-3′) | Size of product
(bp) |
|---|
| h ACTB-F2 |
GCACCCAGCACAATGAAGA | 254 |
| h ACTB-R2 |
AATAAAGCCATGCCAATCTCA |
|
| h NUCB2-F1 |
AAAAGGCAAGAAGTAGGAAGGTT | 124 |
| h NUCB2-R1 |
CAGGATTCAGGTGGTTTAGGTG |
|
Lentivirus infection
5′-GAGGACCACTGCTACAGTA-3′, 5′-TTTAGTAACACCATGTGAG-3′
and 5′-GCTCAGAATGGAATATCAT-3′, (lenti-NUCB2-KD1, lenti-NUCB2-KD2
and lenti-NUCB2-KD3, respectively) were selected to be knocked down
in the present study. A β-actin (ACTB) cDNA plasmid was used as a
negative control, and the lenti-NC cells were treated as the
negative control group. Human NUCB2 and ACTB cDNA plasmids were
transfected with ViraPower packaging mix using Lipofectamine 2000
reagent (Thermo Fisher Scientific, Inc.) into 786-O cells for 48 h
to generate the lentivirus according to the manufacturer's
protocol. The viral supernatant was then collected and used to
infect the 786-O renal cells. Additional analysis was performed 72
h post-transfection. Following this procedure, qPCR and western
blotting were used for the analysis of gene expression.
Apoptosis analysis
Subsequent to successful transfection, the cells
were collected by centrifugation (1,000 × g) for 5 min from the
supernatant at room temperature. Following trypsinization and
washing with PBS twice, the collected cells were mixed with 400 µl
Annexin V-fluorescein isothiocyanate binding buffer (Beyotime
Institute of Biotechnology). Subsequently, 5 µl adenomatous
polyposis coli and propidium iodide were added and mixed. The cell
mixture was incubated for 30 min at 20°C and detected by flow
cytometry with fluorescence-activated cell sorting analysis
following the use of FACS Calibur (BD Biosciences, San Jose, CA,
USA). Apoptotic rate was calculated as follows: Apoptotic rate =
apoptotic cells/(apoptotic cells + normal cells).
Lenti-NUCB2-KD3 was compared with the 786-O cell line
and the lenti-NC cell line.
Transwell invasion assay
Matrigel invasion chambers (Corning Incorporated,
Corning, NY, USA) were used for the invasion assay. The cells were
added to the upper chamber at a density of 5.0×105
cells/ml. A total of 0.1 ml cell suspension liquid was added and
0.6 ml complete medium (RPMI-1640) was added to the bottom of the
chambers. Subsequent to observation at 24–30 h following treatment,
the supernatant was removed from the chamber, and the upper layer
of the filters was wiped with cotton swabs. The cells that had
infiltrated were fixed with 4% paraformaldehyde solution and
stained with 1% crystal violet for 30 min at 37°C. Images of 5
different fields were captured using an Olympus fluorescence
microscope (magnification, ×100; Olympus CKX41; Olympus
Corporation, Tokyo, Japan) equipped with a camera and the number of
invading cells was counted. The control and negative control were
the same as aforementioned in the present study.
Statistical analysis
All data are presented as the mean ± standard error
of the mean. Statistically significant differences between groups
were assessed using one-way analysis of variance followed by the
LSD test. All statistical analysis was performed using a
commercially available statistical package (SPSS 17.0; SPSS Inc.,
Chicago, IL, USA). P≤0.05 was considered to indicate a
statistically significant difference.
Results
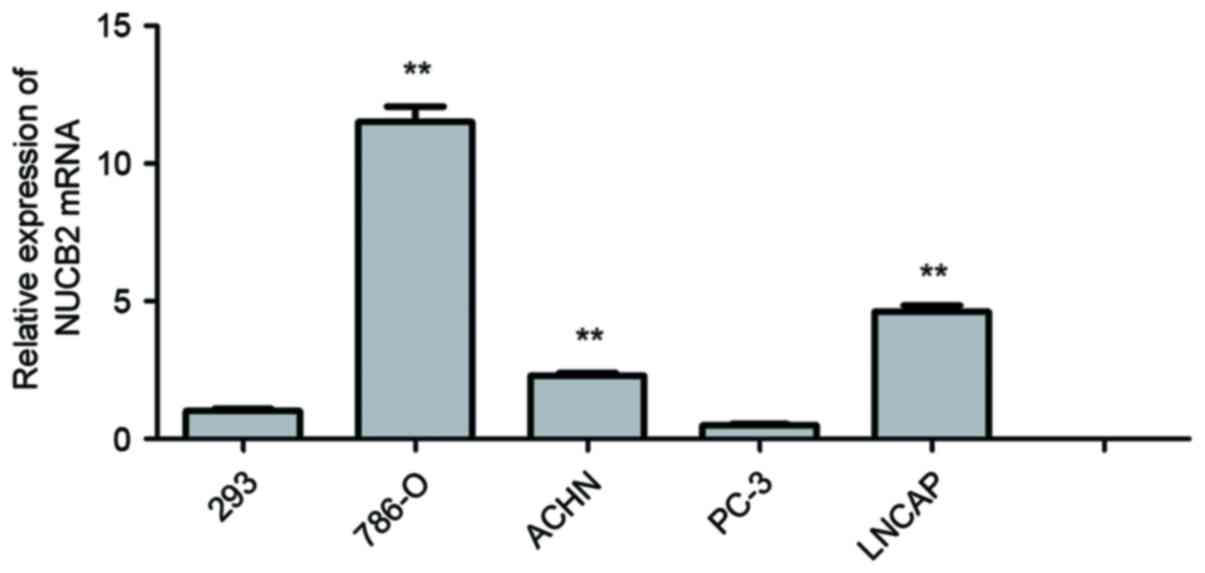
Analysis of NUCB2 gene expression
In the 5 cell lines, the expression of NUCB2 in
786-O (11.50±0.57; P<0.01), ACHN (2.30±0.10; P<0.01) and
LNCaP cell lines (4.62±1.08; P<0.01) was observed to be
significantly increased compared with HEK-293 cells (1.00±0.09).
Among the RCC cell lines, the expression of NUCB2 in 786-O and ACHN
cells was increased most significantly. NUCB2 expression was also
increased in LNCaP cells compared with 293 cells, while there was a
low expression of NUCB2 in PC3 cells compared with HEK-293 cells
(0.50±0.03 vs. 1.00±0.09; P=0.23). These data are presented in
Fig. 1.
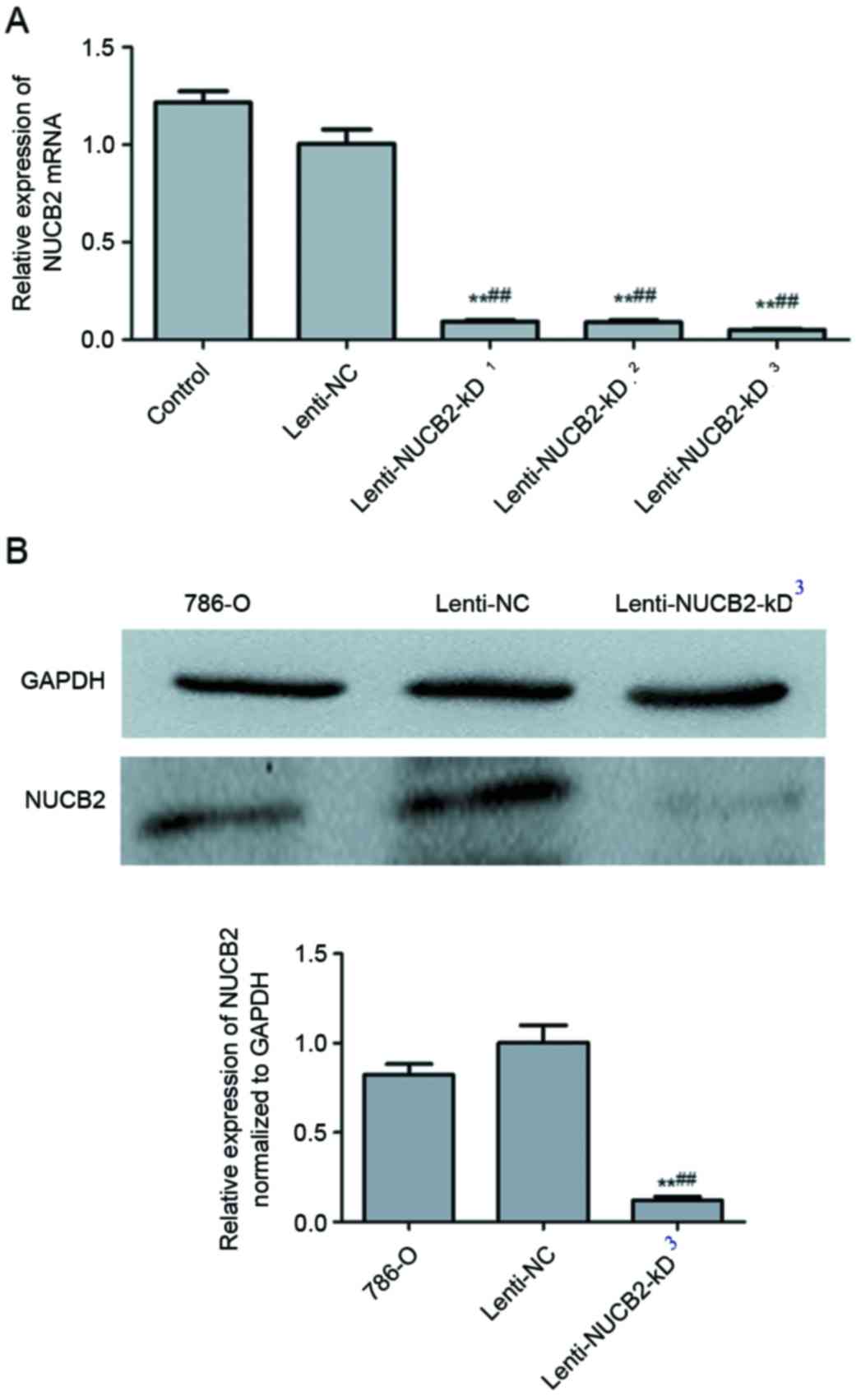
Construction of NUCB2 lentiviral
vector and expression analysis
Subsequent to the construction of the NUCB2
lentiviral vector according to the protocol aforementioned in the
present study, the levels of NUCB2 mRNA expression were detected by
qPCR in the normal 786-O cells, negative control and lenti-NUCB2-KD
group. Significant differences were observed in the lenti-NUCB2-KD
group compared with the 786-O cell (P<0.01) and negative control
group (P<0.01). The present study used 3 different gene segments
for knockdown: i) 5′-GAGGACCACTGCTACAGTA-3′, ii)
5′-TTTAGTAACACCATGTGAG-3′ and iii) 5′-GCTCAGAATGGAATATCAT-3′. NUCB2
mRNA expression was significantly decreased at all three knockdown
groups, while the decrease in the third group was the most marked
(Fig. 2A). Therefore, in subsequent
analyses, the third gene fragment, 5′-GCTCAGAATGGAATATCAT-3′, was
used. Compared with the negative control, NUCB2 protein expression
was significantly inhibited in the lenti-NUCB2-kD3 group
(P<0.01) as determined by western blotting (Fig. 2B).
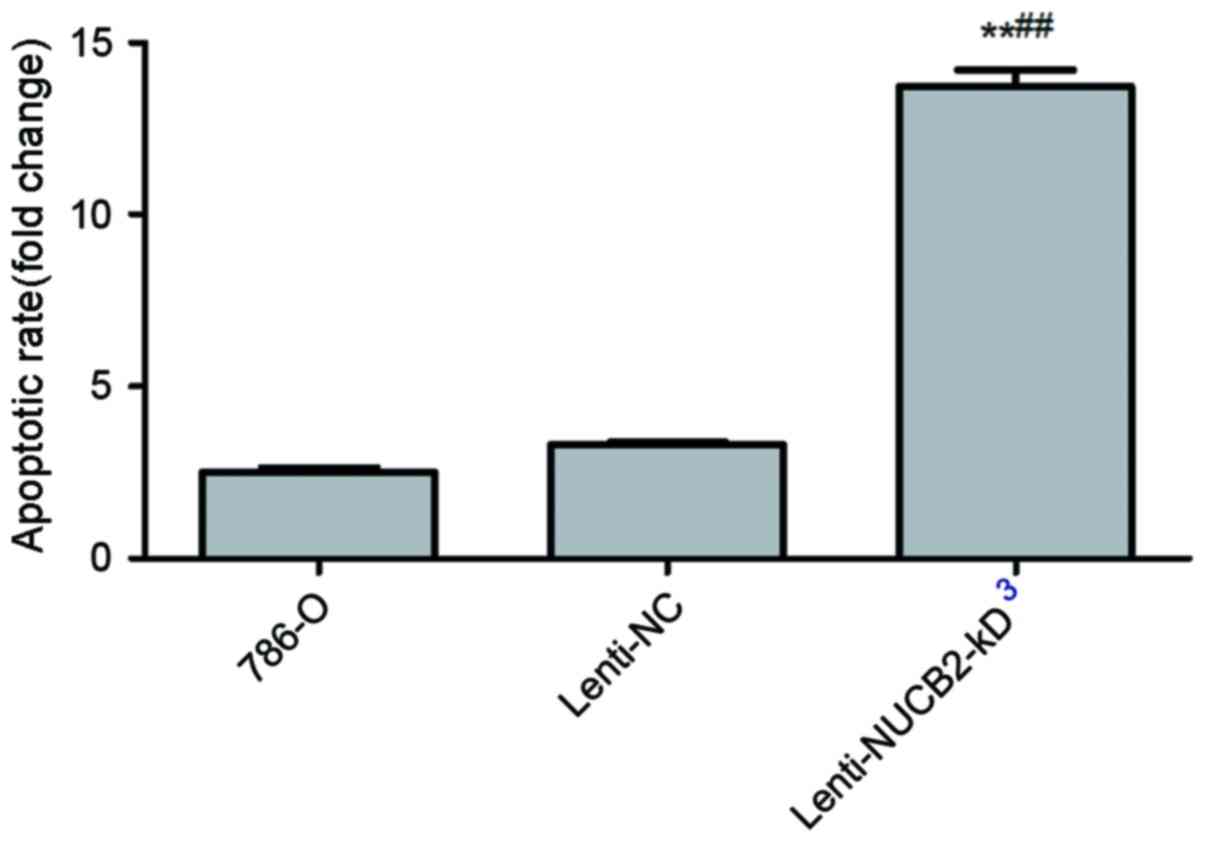
Downregulation of NUCB2 facilitated
apoptosis of renal carcinoma cells
As indicated in Fig.
3, it was observed that the NUCB2 appeared to inhibit apoptosis
of 786-O cells. The results indicated that downregulation of NUCB2
resulted in an increase in the rate of apoptosis compared with the
negative control from 3.32±0.10 to 13.72±0.84% (P<0.01), as
determined by flow cytometry.
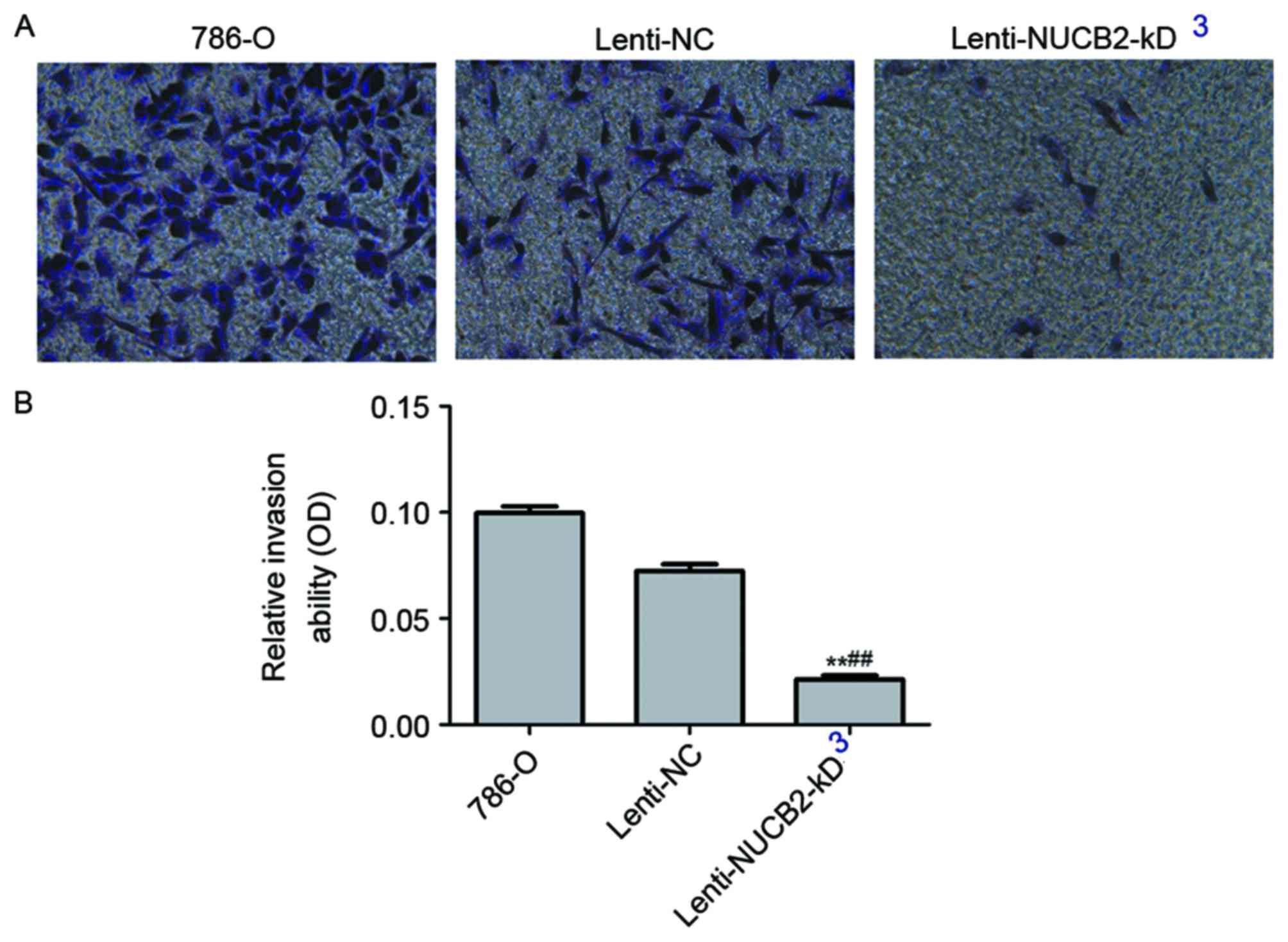
Invasive and migratory abilities of
transfected 786-O cells
Downregulation of NUCB2 resulted in a significant
decreased invasive ability in 786-O cells as determined by
Transwell migration assay (lenti-NUCB2-KD group vs. negative
control group; 0.02±0.002 vs. 0.07±0.003; P<0.01).
Representative images were captured following transfection, and the
cells were stained using crystal violet (Fig. 4).
Discussion
RCC is the most common malignant tumor in human
kidney cancer (2,18). However at present, the etiology of RCC
has not been completely expounded (2). RCC is a heterogeneous and multifactorial
disease with clinical outcomes that are difficult to predict. An
improved understanding of RCC etiology is essential for the
efficient management and early diagnosis, which is of great concern
for its prognosis (2,3). It has been suggested that RCC is a
heterogeneous genetic disease with variable biological and clinical
features (19). Therefore, a number
of studies have focused on the identification of the diagnostic
markers to predict the prognosis of this multifaceted disease
process and have identified a number of genes, which are involved
in the regulation of metabolic disorders. It has been suggested
that all of these genes are important for the development of RCC
(3,20,21).
To the best of our knowledge, the present study is
the first to investigate the novel function of NUCB2 in RCC
development and invasion. The main results consist of three points.
NUCB2 expression was detected in various cell lines of urinary
carcinoma, which may provide a basis for further study of the
mechanism of action of NUCB2 within urologic tumors. Additionally,
to the best our knowledge, the present study first observed that
the downregulation of NUCB2 leads to apoptosis of renal cancer
cells (786-O cell line) in vitro, which indicated the
proliferative effect of NUCB2 on renal carcinoma (16). Finally, the present study identified a
novel function of NUCB2, in that it contributes to renal cancer
invasion as demonstrated by results from the transwell assay.
As a metabolism-regulated peptide, NUCB2 has been
widely studied in the field of glucose and fatty acid metabolism,
particularly in the pancreas, fat tissue and hypothalamus (4). In recent years, increased attention has
focused on the metabolic pathways regulating tumor development.
Numerous neuropeptides have been discussed due to their
tumor-promoting effects (18). NUCB2
was suggested to promote the proliferation of various cells,
including adipocytes and pancreatic cells (12,22,23). In
addition, it has been suggested that NUCB2 is highly expressed in
the cancer of breast, stomach, ovarian epithelium, prostate and
kidneys, according to serological identification and expression
analysis (6,13–16,24). In
breast carcinoma, it has been suggested that NUCB2 may be
upregulated by estrogen and serve an important role in increased
proliferative, migratory and invasive properties in MCF-7 and
SK-BR-3 breast cancer cells (13). It
has also been suggested in gastric carcinoma that NUCB2 is a
candidate biomarker gene for cancer, which may be identified as a
novel potential target for immunotherapeutic treatment (25). Furthermore, a study investigating
ovarian epithelial carcinoma in 2013 by Xu et al (24) revealed a notable function of
NUCB2/nesfatin-1, where it prevented proliferation in the human
ovarian epithelial carcinoma HO-8910 cell line through apoptosis
signaling, via the mechanistic target of rapamycin-RhoA/RhoA kinase
signaling pathway, which serves an important role in cancer cell
proliferation and invasion. In a clinical study of prostate cancer,
it was demonstrated that high NUCB2 expression was associated with
a shorter biochemical recurrence-free survival time compared with
low NUCB2 expression, indicating a potential target for
ameliorating the diagnosis, prognosis and treatment methods of
prostate cancer (6,14,15).
Recently, overexpression of NUCB2 in clear cell RCC was observed to
contribute to malignant clinical pathology and a poor prognosis in
renal cancer, indicating that it may be a novel biomarker (16). Based on a previous publication by the
present authors (10), in the present
study NUCB2 expression in ACHN, 786-O, PC-3 and DU 145 cells was
verified, and a significant high expression was demonstrated in
renal carcinoma 786-O cells. Therefore, the NUCB2 gene was knocked
down in the 786-O cells in order to study its function. The results
in Fig. 2 indicated that the levels
of mRNA and protein were significantly decreased in the
lenti-NUCB2-KD group compared with the control group. Furthermore,
this effect was examined in a subsequent in vitro study,
which indicated that lenti-NUCB2 KD upregulated the apoptotic rate
of the renal carcinoma cells, indicating a proliferation-promoting
function of NUCB2 in renal carcinoma cells. Therefore, gene
production appears to be involved in tumorigenesis and development
of RCC. Notably, a novel function was observed in invasion
analysis. Invasion was significantly decreased in NUCB2-knocked
down cells and a 0.21-fold decrease was identified in the
lenti-NUCB2-KD renal cancer cells compared with the lenti-NC cells.
Therefore, this suggested that NUCB2 contributed to renal carcinoma
invasion. These results contributed to the hypothesis that
neuro-hormonal factors serve an important role in tumorigenesis and
invasion.
One limitation of the present study was that only
the proliferation and invasion of NUCB2-down regulated cells was
examined. The specific signaling pathway requires additional
clarification, according to the cytological results of the present
study. In conclusion, the results of the present study suggest a
novel and important function for NUCB2 in cancer progression,
particularly in the process of development and invasion in renal
cell carcinoma. Therefore, NUCB2 should be considered as a potent
prognostic factor and target in the diagnosis and treatment of
human RCC.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81172450),
the Key Disciplines Group Construction Project of Pudong Health
Bureau of Shanghai (grant no. PWZxq2014-11) and The Program for
Outstanding Medical Academic Leader.
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ljungberg B, Bensalah K, Canfield S,
Dabestani S, Hofmann F, Hora M, Kuczyk MA, Lam T, Marconi L,
Merseburger AS, et al: EAU guidelines on renal cell carcinoma: 2014
update. Eur Urol. 67:913–924. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Massari F, Ciccarese C, Santoni M,
Brunelli M, Piva F, Modena A, Bimbatti D, Fantinel E, Santini D,
Cheng L, et al: Metabolic alterations in renal cell carcinoma.
Cancer Treat Rev. 41:767–776. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oh-I S, Shimizu H, Satoh T, Okada S,
Adachi S, Inoue K, Eguchi H, Yamamoto M, Imaki T, Hashimoto K, et
al: Identification of nesfatin-1 as a satiety molecule in the
hypothalamus. Nature. 443:709–712. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cowley MA and Grove KL: To be or NUCB2, is
nesfatin the answer? Cell Metab. 4:421–422. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang H, Qi C, Wang A, Yao B, Li L, Wang Y
and Xu Y: Prognostication of prostate cancer based on NUCB2 protein
assessment: NUCB2 in prostate cancer. J Exp Clin Cancer Res.
32:772013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cao X, Liu XM and Zhou LH: Recent progress
in research on the distribution and function of NUCB2/nesfatin-1 in
peripheral tissues. Endocr J. 60:1021–1027. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
García-Galiano D, Navarro VM, Gaytan F and
Tena-Sempere M: Expanding roles of NUCB2/nesfatin-1 in
neuroendocrine regulation. J Mol Endocrinol. 45:281–290. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li QC, Wang HY, Chen X, Guan HZ and Jiang
ZY: Fasting plasma levels of nesfatin-1 in patients with type 1 and
type 2 diabetes mellitus and the nutrient-related fluctuation of
nesfatin-1 level in normal humans. Regul Pept. 159:72–77. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dong J, Xu H, Xu H, Wang PF, Cai GJ, Song
HF, Wang CC, Dong ZT, Ju YJ and Jiang ZY: Nesfatin-1 stimulates
fatty-acid oxidation by activating AMP-activated protein kinase in
STZ-induced type 2 diabetic mice. PLoS One. 8:e833972013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Su Y, Zhang J, Tang Y, Bi F and Liu JN:
The novel function of nesfatin-1: Anti-hyperglycemia. Biochem
Biophys Res Commun. 391:1039–1042. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gonzalez R, Reingold BK, Gao X, Gaidhu MP,
Tsushima RG and Unniappan S: Nesfatin-1 exerts a direct,
glucose-dependent insulinotropic action on mouse islet β- and MIN6
cells. J Endocrinol. 208:R9–R16. 2011.PubMed/NCBI
|
|
13
|
Suzuki S, Takagi K, Miki Y, Onodera Y,
Akahira J, Ebata A, Ishida T, Watanabe M, Sasano H and Suzuki T:
Nucleobindin 2 in human breast carcinoma as a potent prognostic
factor. Cancer Sci. 103:136–143. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang H, Qi C, Li L, Luo F and Xu Y:
Clinical significance of NUCB2 mRNA expression in prostate cancer.
J Exp Clin Cancer Res. 32:562013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang H, Qi C, Wang A, Li L and Xu Y: High
expression of nucleobindin 2 mRNA: An independent prognostic factor
for overall survival of patients with prostate cancer. Tumour Biol.
35:2025–2028. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qi C, Ma H, Zhang HT, Gao JD and Xu Y:
Nucleobindin 2 expression is an independent prognostic factor for
clear cell renal cell carcinoma. Histopathology. 66:650–657. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Siegel R, Naishadham D and Jemal A: Cancer
Statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Muriel L, ópez C, Esteban E, Berros JP,
Pardo P, Astudillo A, Izquierdo M, Crespo G, Sanmamed M, Fonseca PJ
and Martínez-Camblor P: Prognostic factors in patients with
advanced renal cell carcinoma. Clin Genitourin Cancer. 10:262–270.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen K, Zeng J, Xiao H, Huang C, Hu J, Yao
W, Yu G, Xiao W, Xu H and Ye Z: Regulation of glucose metabolism by
p62/SQSTM1 through HIF1α. J Cell Sci. 129:817–830. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Aboudehen K, Kim MS, Mitsche M, Garland K,
Anderson N, Noureddine L, Pontoglio M, Patel V, Xie Y, DeBose-Boyd
R and Igarashi P: Transcription factor hepatocyte nuclear factor-1β
regulates renal cholesterol metabolism. J Am Soc Nephrol.
27:2408–2421. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ramanjaneya M, Tan BK, Rucinski M, Kawan
M, Hu J, Kaur J, Patel VH, Malendowicz LK, Komarowska H, Lehnert H
and Randeva HS: Nesfatin-1 inhibits proliferation and enhances
apoptosis of human adrenocortical H295R cells. J Endocrinol.
226:1–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dai H, Li X, He T, Wang Y, Wang Z, Wang S,
Xing M, Sun W and Ding H: Decreased plasma nesfatin-1 levels in
patients with acute myocardial infarction. Peptides. 46:167–171.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu Y, Pang X, Dong M, Wen F and Zhang Y:
Nesfatin-1 inhibits ovarian epithelial carcinoma cell proliferation
in vitro. Biochem Biophys Res Commun. 440:467–472. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Line A, Stengrēvics A, Slucka Z, Li G,
Jankevics E and Rees RC: Serological identification and expression
analysis of gastric cancer-associated genes. Br J Cancer.
86:1824–1830. 2002. View Article : Google Scholar : PubMed/NCBI
|