Introduction
The occurrence and development of tumors is
modulated by the dual regulation of genetic instability and the
tumor microenvironment. Several studies have shown that the tumor
microenvironment is important in the development and progression of
malignant tumors; however, its mechanisms remain to be fully
elucidated. Metabolism is one of the basic biological life
activities in the body; changes in the biological properties of
normal cells when converted to malignant cells include metabolic
reprogramming. In previous years, studies have revealed that the
activation of certain oncogenes or tumor suppressor inactivation
can regulate metabolic homeostasis, and are involved in promoting
or inhibiting cancer progression (1,2). Certain
metabolic enzymes can themselves be used as oncogenes or tumor
suppressors involved in the process of cancer development.
Cardiolipin (CL) is a major mitochondrial membrane
polyglycerophospholipid (3,4). It has been shown to be required for the
activation of several pivotal mitochondrial membrane enzymes
involved in ATP synthesis and mitochondrial respiratory complexes
(5). CL may be involved in the
pathway leading to cellular apoptosis (6,7). The final
step for the de novo mitochondrial synthesis of CL is based
on the substrates phosphatidylglycerol and cytidine diphosphate
choline-diacylglycerol. This reaction is catalyzed by the enzyme
cardiolipin synthase 1 (CRLS1) (8),
which is ubiquitous in eukaryotes, archaebacteria and eubacteria.
The human CRLS1 gene has previously been cloned and
characterized (9,10).
Although CRLS1 has been demonstrated to be
functional in lipid anyagcsere, the exact role in tumorigenesis
remains to be fully elucidated, with the exception of
investigations in patients with solid and non-solid malignancies,
which showed that anti-cardiolipin antibodies were associated with
an increased rate of thrombosis (11). The expression, prognostic effects and
potential mechanism of CRLS1 in tumors remain to be elucidated. It
is known that lung cancer is one of the malignant tumors with high
morbidity and mortality rates in the world. Non-small cell lung
cancer (NSCLC) accounts for almost 85% of all lung cancer cases and
generally presents at an advanced stage at diagnosis with a 5-year
survival rate of <5% (12,13). Although standard chemotherapy and
molecular targeted therapy have improved overall survival (OS) and
quality of life, the survival benefit is limited to those who lack
a driver mutation or show resistance to drugs (14). Therefore, it is important to
investigate the underlying and intertwined mechanisms of lung
tumorigenesis and tumor progression to provide novel ideas and a
theoretical basis for the treatment of NSCLC.
The present study focused on the association between
metabolism-related genes and CRLS1 in addition to NSCLC, and
comprehensively analyzed the mRNA expression of CRLS1 in tumors and
patient prognosis. Furthermore, the mechanism in tumor development
was examined using web-based and soft-based microarray databases.
Together, the results suggested that CRLS1 is a promising
candidate gene for prognostic and diagnostic approaches in NSCLC,
and warrants further investigation.
Materials and methods
Oncomine analysis
Oncomine, a cancer microarray database and
integrated data-mining platform, was used to facilitate the
identification from genome-wide expression analyses, and the
transcriptome data in major types of cancer were compared with
respective normal tissues (15,16). The
metabolism-related gene expression of CRLS1 (Affymetrix ID,
241741_at) was analyzed using Oncomine™ research premium edition
(http://oncomine.org). The mRNA levels in NSCLC
vs. normal patient datasets were compared. P<0.0001,
fold-change=2, and top 10% gene rank were adjusted as the
thresholds. Subsequently, the log2 median centered intensity, and
the 10 and 90th percentile data from Oncomine concerning
CRLS1 were plotted using GraphPad Prism software (version
5.0; GraphPad Software, Inc., La Jolla, CA, USA).
Analysis of survival prognosis using
the Kaplan-Meier plotter
The correlation between survival prognosis and the
mRNA expression of CRLS1 in NSCLC was evaluated using the
Kaplan-Meier plotter (www.kmplot.com) (17),
an online database containing gene expression data and clinical
data for lung cancer, breast cancer, stomach cancer and ovarian
cancer. The patient samples were divided into two cohorts according
to the median expression of the gene (high vs. low expression);
this limited the cohorts in accordance with the clinical
characteristics of patients, including sex, pathological type,
clinical stage and treatment. OS was analyzed in patients with
NSCLC using a Kaplan-Meier survival plot and CRLS1 was
uploaded into the database to obtain the Kaplan-Meier survival
plots. Subsequently, the plot data were exported as text and the
Kaplan-Meier survival curve was plotted using GraphPad Prism
software.
Potential mechanism in NSCLC
meta-analysis of CRLS1
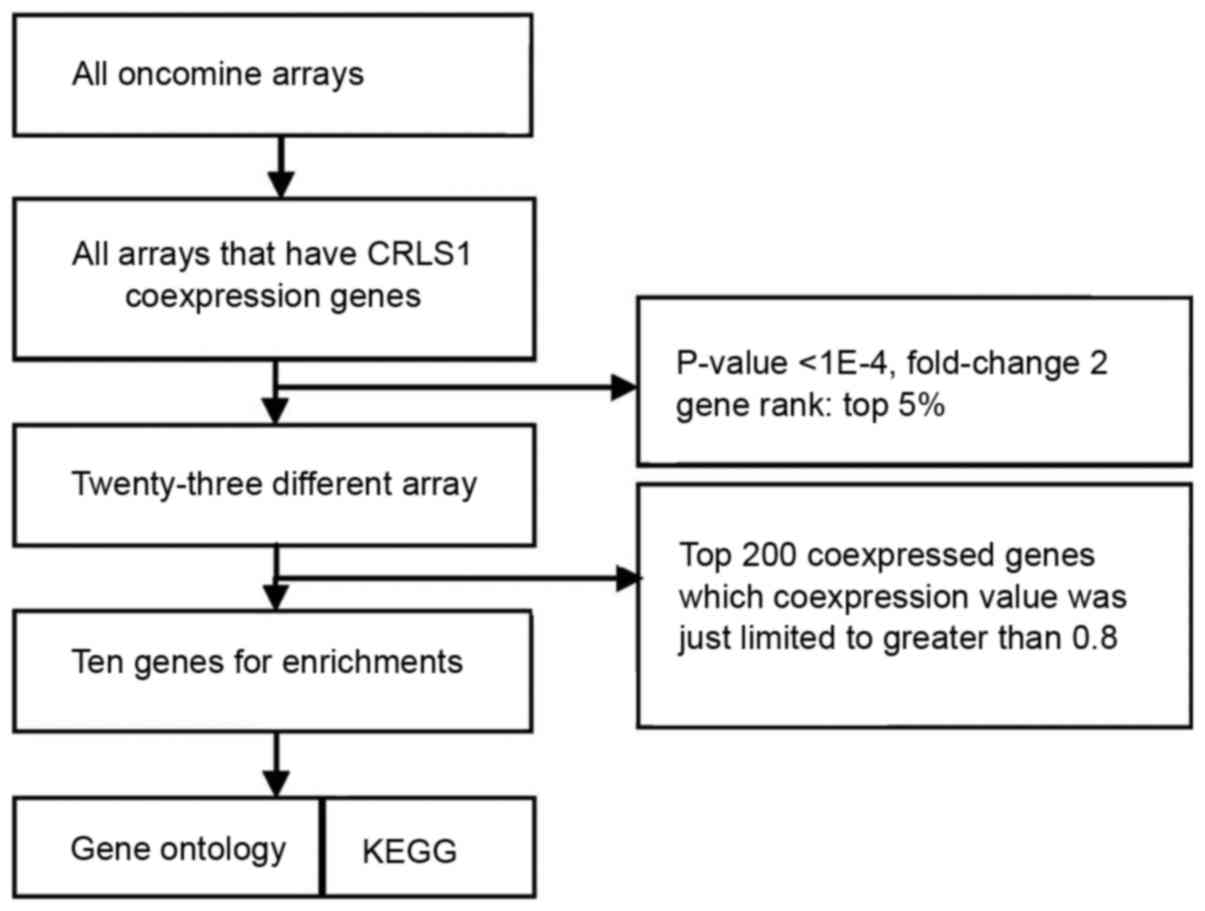
To understand the function of CRLS1, co-expression
analysis was performed using the Oncomine database (http://oncomine.org). The threshold was adjusted to
P<0.0001, fold-change=2, and gene rank of top 5%. A total of 23
different arrays fulfilled these criteria and the top 200
co-expressed genes, where the co-expression value was limited to
>0.8, were extracted and filtered to obtain one representative
gene per study following the removal of duplicates. These filtered
gene lists were then compared with searches for repeatedly
co-expressed genes over multiple studies, with the two studies
(>30% of five studies) frequency cut-off. Through this, a
meta-analysis list for CRLS1 was generated, and enriched Gene
Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG)
pathway analyses were performed for the terms of the gene lists
produced via co-expression data analysis on account of the
web-based Database for Annotation, Visualization and Integrated
Discovery (DAVID; http://david.abcc.ncifcrf.gov) (18,19). The
Benjamini and Hochberg false discovery rate correction was used to
correct the results for multiple testing.
Results
Gene expression of CRLS1 in lung
cancer
Through Oncomine analysis, the present study
investigated the mRNA levels of CRLS1 in human cancer cases. There
were more datasets showing a downregulation of CRLS1 than an
upregulation of CRLS1 in the various types of tumors, compared with
the normal tissues (Table I). This
was similar in lung cancer, vs. normal tissues, for which three
datasets showed a downregulation of CRLS1 and none showed an
upregulation of CRLS1 under the threshold of P=0.05,
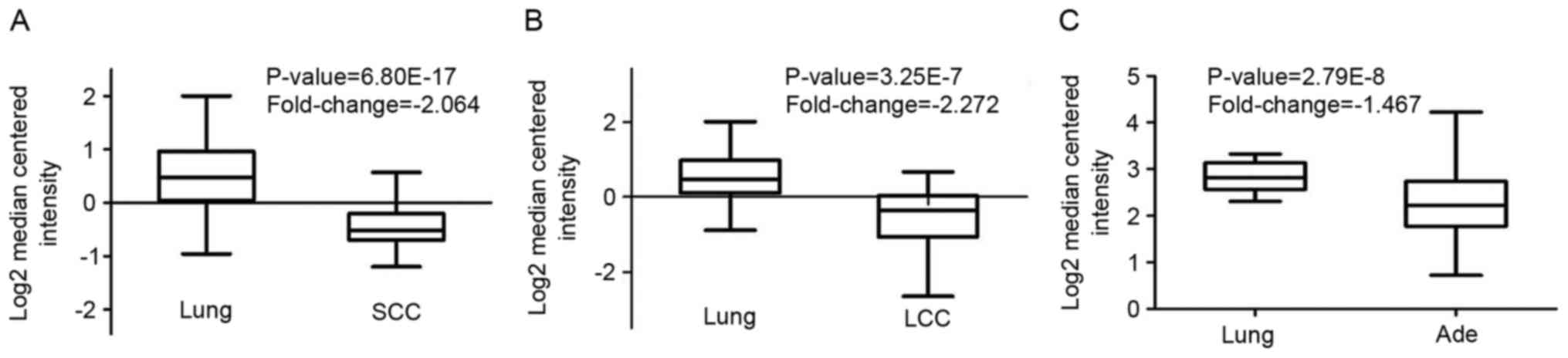
fold-change=1.5, and gene rank of 10%. The mRNA levels of CRLS1 in
squamous cell carcinoma (SCC) and large cell lung cancer (LCC) were
lower, compared with those in normal tissues (fold-change, −2.046
and −2.272, respectively; Fig. 1A and
B). However, compared with the normal tissues, no changes in
the mRNA levels of CRLS1 were found in adenocarcinoma (Ade), with a
fold change of −1.467 (Fig. 1C).
Taken together, these data confirmed that CTLS1 was downregulated
in lung tumors, compared to normal tissues (Table II).
 | Table I.Number of datasets correlated with
upregulation and downregulation of the CRLS1 gene in cancer vs.
normal tissues, determined via oncomine analysis at different
P-values. |
Table I.
Number of datasets correlated with
upregulation and downregulation of the CRLS1 gene in cancer vs.
normal tissues, determined via oncomine analysis at different
P-values.
|
| Datasets for
CRLS1 |
|---|
|
|
|
|---|
| P-value | Upregulated
(n) | Downregulated
(n) |
|---|
| 0.05 | 8 | 10 |
| 0.01 | 8 | 10 |
| 1E-03 | 8 | 8 |
| 1E-04 | 5 | 8 |
| 1E-05 | 4 | 8 |
| 1E-06 | 3 | 7 |
| 1E-07 | 2 | 6 |
| 1E-08 | 1 | 6 |
| Table II.Gene expression of CRLS1 is elevated
in lung cancer, determined using the Oncomine database. |
Table II.
Gene expression of CRLS1 is elevated
in lung cancer, determined using the Oncomine database.
|
|
|
| Sample size
(n) |
|---|
|
|
|
|
|
|---|
| Comparison | P-value | Fold-change | Dataset | Normal | Cancer | Total |
|---|
| Lung Ade vs.
normal | 2.79E-8 | −1.567 | Okayama | 20 | 226 | 246 |
| Squamous cell
carcinoma vs. normal | 6.80E-17 | −10.564 | Hou | 65 | 27 | 92 |
| Large cell
carcinoma vs. normal | 3.25E-7 | −6.603 | Hou | 65 | 19 | 84 |
Survival prognosis analysis
The association between the mRNA expression of CRLS1
and clinical outcomes was examined using the Kaplan-Meier plotter
(www.kmplot.com). Survival curves were plotted for
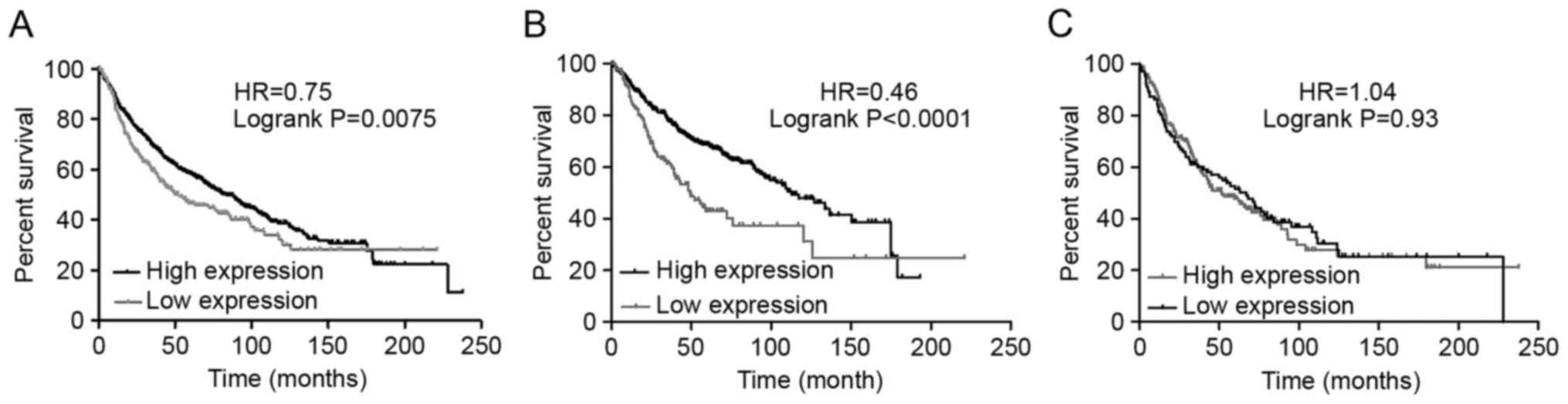
all patients with NSCLC (Fig. 2A),
Ade (Fig. 2B), and SCC (Fig. 2C). Consequently, a high mRNA
expression of CRLS1 was found to be associated with significantly
improved OS for all patients with NSCLC [HR=0.75 (0.61–0.93);
P=0.0075]. In addition, a high mRNA expression of CRLS1 was found
to be associated with significantly improved OS in Ade [HR=0.46
(0.33–0.64); P<0.0001]. However, no statistically significant
difference was found in the expression levels in patients with SCC
[HR=1.04 (0.76–1.42); P=0.83].
The association between CRLS1 and the clinical
characteristics for patients with NSCLC, including sex, smoking
status, tumor grade, clinical stage, lymph node status,
chemotherapy and radiotherapy, was also investigated. CRLS1 was
associated with significantly improved OS in stage I tumors and N0
lymph node status in patients with NSCLC. No significant difference
was found in stages II–IV tumors or N1 and N2 lymph node status.
CRLS1 was not significantly associated with sex or smoking status
in patients with NSCLC. No association between CRLS1 and
chemotherapy was observed, as the sample size was too small. The
results of the associations are shown in Table III.
| Table III.Correlation between CRLS1 and tumor
clinical characteristics of patients with non-small cell lung
cancer. |
Table III.
Correlation between CRLS1 and tumor
clinical characteristics of patients with non-small cell lung
cancer.
| Characteristic | Cases low (n) | Cases high (n) | HR (95% CI) | P-value |
|---|
| Stage |
|
|
|
|
| I | 224 | 225 | 0.36
(0.26–0.51) | 2.7E-09 |
| II | 80 | 81 | 0.69
(0.43–1.09) | 0.11 |
|
III | 22 | 22 | 0.54
(0.27–1.09) | 0.08 |
| Topography |
|
|
|
|
| I | 112 | 112 | 0.63
(0.42–0.94) | 0.023 |
| II | 95 | 95 | 0.81
(0.55–1.17) | 0.26 |
|
III | 14 | 15 | 1.06
(0.47–2.38) | 0.89 |
| Lymph node
status |
|
|
|
|
| N0 | 162 | 162 | 0.72
(0.52–0.98) | 0.037 |
| N1 | 51 | 51 | 0.81
(0.49–1.34) | 0.42 |
| N2 | 16 | 16 | 0.56
(0.27–1.2) | 0.13 |
| Metastasis |
|
|
|
|
| M0 | 231 | 231 | 0.69
(0.54–0.89) | 0.0037 |
| M1 | – | – | – | – |
| Smoking status |
|
|
|
|
| Never
Smoked | 151 | 149 | 0.51
(0.33–0.78) | 0.0018 |
|
Smoked | 70 | 71 | 0.33
(0.13–0.83) | 0.014 |
| Sex |
|
|
|
|
|
Male | 330 | 329 | 0.54
(0.44–0.67) | 8.7E-09 |
|
Female | 189 | 186 | 0.45
(0.31–0.65) | 8.6E-06 |
| Chemotherapy |
|
|
|
|
|
Yes | 17 | 17 | 0.41
(0.12–1.34) | 0.13 |
| No | 10 | 10 | 3.94
(0.46–33.95) | 0.18 |
CRLS1 is co-expressed with genes
involved in tumor metabolism and the mitogen-activated protein
kinase (MAPK) signaling pathway
Using the Oncomine cancer microarray database, a
search was performed of CRLS1 with co-expressed genes. The
retrieval flow of the meta-analysis and the selected multi-array
studies for CRLS1 are shown in Fig.
3. Following meta-analysis, 10 genes were found as being
co-expressed in two or more studies, comprising G protein subunit
γ10, membrane associated ring-CH-type finger 7 (MARCH7), MARCH5,
DAZ interacting zinc finger protein 1 (DZIP1), selenophosphate
synthetase 2 (SEPHS2), Obg-like ATPase 1 (OLA1), SEPHS1,
dual-specificity phosphatase 7 (DUSP7), protein phosphatase,
Mg2+/Mn2+-dependent 1A (PPM1A) and protein
tyrosine phosphatase receptor type R (PPTRR) in five datasets that
met the criteria (Jia Liver, Hoek Melanoma 2, Tomida Lung, Kreike
Breast, and Fandy Leukemia). Web-based DAVID was used to perform GO
term enrichment analysis to obtain the specific functions of the
above-mentioned co-expressed genes, which provided a list of gene
functions in a gene set. Analysis of the 10 CRLS1-co-expressed
genes with the DAVID functional annotation tool (GOTERM_BP/MF_FAT)
resulted in eight annotation clusters matching the statistical
criteria (P<0.05; count ≥2; fold enrichment >2; Table IV). Subsequently, the above-mentioned
DAVID functional annotation tool was used for the identification of
putative KEGG pathways associated with the CRLS1-co-expressed
genes. Consequently, one pathway associated with seleno-amino acid
metabolism and another associated with the MAPK signaling pathway
in cancer. These two signaling pathways were significantly enriched
with CRLS1-co-expressed genes (P<0.05 and fold enrichment >2;
Table V).
| Table IV.Functional enrichment of
CRLS1-co-expressed genes. |
Table IV.
Functional enrichment of
CRLS1-co-expressed genes.
| Term | Count | % | P-value | Fold-change |
|---|
|
GO:0004756-selenide, water dikinase
activity | 2 | 20 | 1.4E-3 | 1,298.3 |
|
GO:0016781-phosphotransferase
activity | 2 | 20 | 1.4E-3 | 1,298.3 |
| GO:0006470-protein
amino acid dephosphorylation | 3 | 30 | 2.6E-3 | 33.9 |
|
GO:0016311-dephosphorylation | 3 | 30 | 3.4E-3 | 29.3 |
|
GO:0004721-phosphoprotein phosphatase
activity | 3 | 30 | 5.5E-3 | 23.6 |
|
GO:0016791-phosphatase activity | 3 | 30 | 1.2E-2 | 15.6 |
| GO:0008430-selenium
binding | 2 | 20 | 2.2E-2 | 81.1 |
| GO:0004722-protein
serine/threonine phosphatase activity | 2 | 20 | 3.1E-2 | 57.7 |
| Table V.Pathway-based enrichment of
CRLS1-co-expressed genes. |
Table V.
Pathway-based enrichment of
CRLS1-co-expressed genes.
| Term | Count | % | P-value | Fold-change |
|---|
| Hsa04010:MAPK
signaling pathway | 3 | 30 | 2.5E-2 | 9.5 |
|
Hsa00450:Selenoamino acid metabolism | 2 | 20 | 3.8E-2 | 65.2 |
CRLS1 is differentially expressed in
various types of cancer
The expression of CRLS1 in various types of cancer
was investigated using publicly available gene expression data from
Oncomine, the results of which are shown in Table VI.
| Table VI.Differential expression of CRLS1 in
cancer tissue, compared with their normal tissue via the Oncomine
cancer microarray database. |
Table VI.
Differential expression of CRLS1 in
cancer tissue, compared with their normal tissue via the Oncomine
cancer microarray database.
| Type of cancer | Over expressed | Under
expressed | Dataset count |
|---|
| Brain tumor | + | + | 4/1 |
| Leucocythemia |
| + | 5 |
| Lung cancer |
| + | 2 |
| Pancreatic
carcinoma |
| + | 1 |
| Lymphoma | + |
| 1 |
| Tongue cancer | + |
| 1 |
| Mouth floor
carcinoma | + |
| 1 |
Discussion
Warburg first suggested that the prime cause of
cancer was impaired energy metabolism, which involved irreversible
injury to cellular respiration (20,21). The
majority of normal mammalian cells achieve a constant delta G' of
ATP hydrolysis of ~257 kJ/mol as energy required for normal life
activities through respiration (22,23). By
contrast, tumor cells maintain this level through a combination of
respiration and glycolysis (24,25).
Elevated glycolysis is a metabolic characteristic of almost all
types of tumor, which can be observed through the use of labeled
glucose analogs (26,27). Structural and biochemical
abnormalities that have occurred in tumor cell mitochondria can
compromise oxidative respiratory function. Several studies have
assessed the lipid composition of tumor mitochondria (28,29),
however, few have evaluated the content and the composition of CL
in tumor mitochondria, which is a complex mitochondrial-specific
phospholipid regulating numerous enzyme activities, particularly
those associated with oxidative phosphorylation and coupled
respiration (30,31). Kiebish et al investigated the
content and composition of CL for the first time in purified
mitochondria from mouse brain tumors. The results showed that the
composition and content of CL in mouse brain tumor mitochondria
differed distinctly from that derived from syngeneic host normal
brain tissue. The mouse brain tumors contained CL abnormalities,
which were unique to each tumor type. In addition, these CL
abnormalities were associated with significant hypofunction in the
electron transport chain (ETC). Taken together, the results
suggested that abnormal CL underlies irreversible respiratory
injury in tumors, which further supports the importance of CL in
maintaining the structural integrity of the inner mitochondrial
membrane and lined mitochondrial lipid defects to the Warburg
theory of tumorigenesis (30).
CRLS1, also known as C20 or f155, is the human
candidate gene for CL synthase, is localized in the mitochondria
and is expressed at high levels in tissues, including the heart,
skeletal muscle, lung and liver, belonging to the CDP-alcohol
phosphatidyltransferase class-I family. It predominantly catalyzes
the reversible phosphatidy l group transfer from one
phosphatidylglycerol molecule to another to form CL and glycerol.
As a result of these findings, the present study hypothesized that
the reduction of CRLS1 caused by various factors contributes either
directly or indirectly to abnormalities in CL synthesis or
remodeling, and leads to damage to mitochondrial intima integrity
and hypofunction in ETC activities. This suggests a reliance on
glycolysis for cell survival, eventually resulting in the
occurrence and development of tumors. In addition to the
enzyme-mediated inhibition of CL synthesis, inherited mutations, a
variety of epigenetic causes and environmental insults, including
necrosis, hypoxia, ischemia, dietary imbalances and reactive oxygen
species, can also produce mitochondrial defects to cause CL
abnormalities (31,32), contributing to tumor initiation and
progression. However, enzyme-mediated CL synthesis may be the most
direct element and was investigated as one of the primary aims of
the present study. In addition, based-wed DAVID GO term enrichment
analysis showed that CRLS1-co-expressed genes were involved in
phosphoprotein phosphatase activity, protein amino acid
dephosphorylation, protein serine/threonine phosphatase activity
and phosphate metabolic process. According to the co-expression
analysis theory, which is based on hierarchical clustering in which
groups of genes may have similar functions if they have similar
expression patterns under a given pathological condition, it is
possible to examine whether CRLS1 possesses putative and analogical
functional activities with its co-expressed genes. These functional
abnormalities caused by abnormalities in lipid metabolism may also
be complementary contributors in tumorigenesis.
In previous years, studies have found that oncogene
activation or tumor suppressor gene inactivation can regulate
metabolic homeostasis in order to promote or inhibit cancer.
Furthermore, certain metabolic enzymes can be used as oncogenes or
tumor suppressors involved in the process of cancer progression.
Members of the MAPK family, including the extracellular
signal-regulated protein kinase (ERK) pathway, c-Jun N-terminal
kinase (JNK) pathway, p38 pathway and ERK5 pathway, have been
reported to be important in cell proliferation and apoptosis
(33). From the extracellular
stimulus to the cells to the corresponding biological effects, the
MAPK signal transduction pathway involves a signaling cascade of
multistage protein kinases, including three key kinases, MAPK,
MAPKK and MAPKKK (34). The analysis
of KEGG pathways via DAVID showed that CRLS1 may be involved in the
MAPK signaling pathway as it is co-expressed with PPM1A, protein
tyrosine phosphatase receptor type R protein (PTPRR) and DUSP7.
PPM1A is a phosphatase belonging to the PP2C family, which
dephosphorylates and inactivates a broad range of substrates,
including MAPKs, p38 and JNK, and is involved in tumor suppression
(35,36), Su et al (37) characterized the tumor suppressor
function of PTPRR, particularly its role in metastasis via the MAPK
signaling pathway, in a cervical cancer model. Taken together, it
was suggested that PTPRR affects malignant phenotypes by
dephosphorylating substrates, including ERK5 and the p38 pathway,
which has also been observed in other species (38). However, the extent of kinases targeted
by PTPRR in NSCLC has not been investigated. DUSPs show marked
substrate preferences for specific MAPKs, which have been reported
to limit the duration of MAPK phosphorylation and reduce microglial
activation (39). Sharing a substrate
preference for ERK1/2 may be categorized as inducible or
constitutive according to their expression. Inducible DUSPs
dephosphorylate JNK, p38 and ERK, whereas constitutive DUSPs are
specific for ERK (40,41). Several studies of DUSPs in tumors have
focused on constitutive and inducible DUSP7, which downregulates
p38 and ERK in major inflammatory pathways (42,43). To
date, few studies have reported on the roles of DUSP6 and other
DUSPs in microglia.
The MAPK pathway consists of a three-tiered kinase
cascade and the sequential phosphorylation of each kinase causes
the activation of effector kinases JNK, p38, and ERK1/2. Three
classes of protein phosphatases modulate the activities of upstream
kinases: Type-2 Ser/Thr phosphatases (PP2A) including PPM1A, PTPs
including PTPRR, and dual-specificity (Thr/Tyr) phosphatases
(DUSPs), including DUSP7 (42). Using
the DAVID functional annotation chart, several subclasses of MAPK
signaling pathways may reveal the role of CRLS1 in tumorigenesis
based on the co-expression analysis theory. The JNKs are members of
the MAPK family and are activated by MAP2 kinases MKK-4 and MKK-7.
This subsequently translocates to the nucleus where it can regulate
the activity of multiple transcription factors. At the
post-translational level, JNK enhances its transcriptional activity
by double-phosphorylating Ser-63 and Ser-73 sites in the c-jun
activation region, and eventually affects cell cycle, apoptosis,
cell stress, and other physiological and pathological processes
(44). According to the KEGG pathways
and co-expression patterns identified in the present study, the
putative CRLS1 JNK/c-Jun MAPK signaling pathway was demonstrated
(Fig. 4A). CRLS1 prevents the
phosphorylation of MKK-4 and MKK-7 by dephosphorylation and
reducing its transcription by c-jun by suppressing the activation
of JNK. The p38 signaling pathway (45) is activated through phosphorylation by
the upstream MKK-3, which then translocates into the nucleus to
interact with downstream protein kinases, chromosome remodeling and
transcription factors, including mitogen-and stress-activated
protein kinase 1/2 (MSK1/2), MYC associated factor X (MAX) and ETS
transcription factor (ELK-1). Similarly, the putative CRLS1 p38
MAPK signaling pathway indicates the role of CRLS1 in tumors
(Fig. 4B). CRLS1 inhibits the
phosphorylation of MKK-3 and p38 by dephosphorylation, and reduces
the transcription of MSK1/2, MAX/GADD153 and ELK-1 by suppressing
the activation of p38. ERKs were the first subclass to be
investigated and are also the most important members of the MAPK
family (46). The ERK signaling
pathway is activated by growth factors and mitotic stimuli, which
are closely associated with cell proliferation, differentiation and
transcription. ERK is activated continuously, and ultimately
promotes cell proliferation and malignant transformation. The ERK
cascade contains a typical three-level MAPK sequence activation
process. Raf belongs to the MAPKKK family, which phosphorylates and
activates MEK1/2 (MAPKK), and subsequently activates ERK1/2 by
double phosphorylation. The same reaction model can also be applied
to the CRLS1 ERK MAPK signaling pathway (Fig. 4C). CRLS1 prevents the phosphorylation
of ERK by dephosphorylation, and reduces the transcription of MAP
kinase-interacting kinase 1/2 and Elk-1 by suppressing the
activation of ERK.
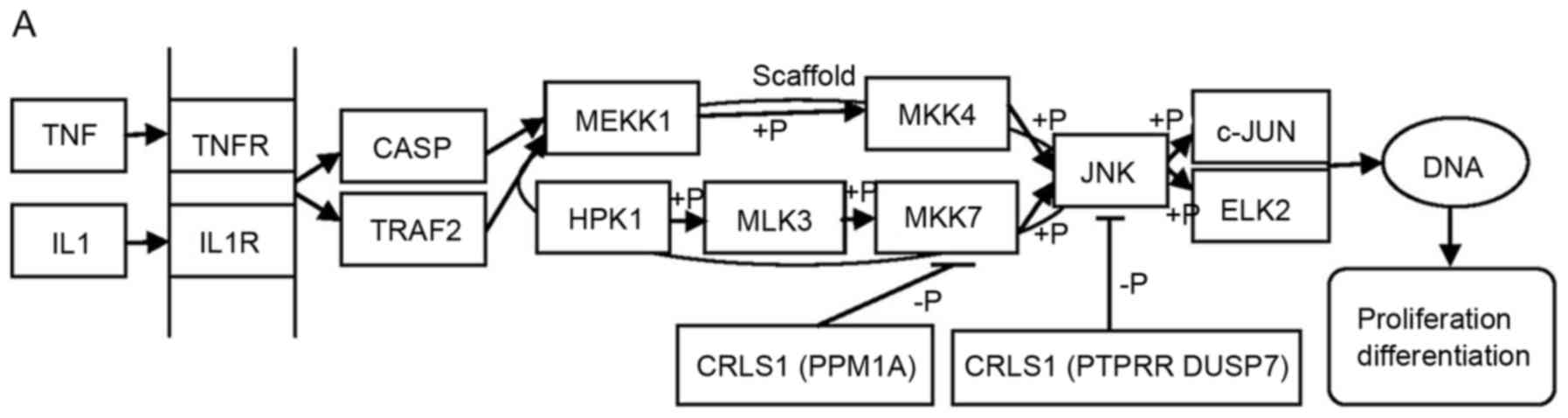 | Figure 4.MAPK signal pathway involved in CRLS1
co-expression genes by KEGG pathway analysis. (A) JNK/c-JUN MAPK
signaling pathway. (B) p38 MAPK signaling pathway. (C) ERK MAPK
signaling pathway. Arrows indicate promotion. T-shaped arrows
represent an inhibitory effect. Genes in brackets represent CRLS1
co-expression genes. +p, phosphorylation; -p, dephosphorylation;
CRLS1, cardiolipin synthase 1; KEGG, Kyoto Encyclopedia of Genes
and Genomes; MAPK, mitogen-activated protein kinase; JNK, c-Jun
N-terminal kinase; ERK, extracellular signal-regulated kinase; TNF,
tumor necrosis factor; IL1, interleukin 1; TNFR, TNF receptor;
CASP, caspase; TRAF2, TNFR-associated factor 2; MEKK1, MAPK kinase
kinase kinase 1; HPK1, hematopoietic progenitor kinase 1; MLK3,
mixed lineage kinase 3; MKK, MPAK kinase. |
In terms of seleno-amino acid metabolism, KEGG
pathways analysis via DAVID revealed that SEPHS1/2 is a
CRLS1-co-expression gene involved in tumorigenesis. In the 1970s
and 1980s, clinical data was collected examining the correlation
between selenomethionine (Se-Met) and cancer incidence in various
tumor types, including lung, skin and colon cancer (47). Several studies have identified
selenium and Se-Met as potential chemopreventive agents against
prostate cancer (48,49). SEPHS1/2 belongs to the selenophosphate
synthetase 1 family, class II subfamily, which predominantly
synthesizes selenophosphate from selenide and ATP (catalytic
activity: ATP + selenide + H2O=AMP +
selenophosphate+phosphate). Studies have also shown that SEPHS
affects cell viability upon ionization radiation through the
modulation of p53, which further suggests that SEPHS and its
reaction product selenophosphate may be involved in inhibiting
cancer progression in a p53-dependent manner and offers potential
as a novel anticancer measure (50,51).
However, the antitumor mechanisms of selenophosphate and SEPHS
described above have not been validated in the field of lung
cancer. By contrast, the co-expression analysis theory indicates a
similar role for CRLS1, which may be confirmed by further
investigations of NSCLC-related seleno-amino acid metabolism and
the p53 pathway.
In conclusion, using in silico approaches,
the differential expression of CRLS1 in NSCLC and normal tissue was
assessed in the present study, which revealed that the expression
level of CRLS1 in lung cancer was lower, compared with that in
normal lung tissues. In addition, a high expression of CRLS1 was
found to be associated with improved OS in all patients with NSCLC
and lung Ade. The meta-analysis for CRLS1-related co-expression
analysis revealed that this molecule may be important for
tumor-associated lipid metabolic processes, and its anti-oncogene
position was also assessed, involving the MAPK signaling pathway.
The above conclusions were obtained using various tools, software
and large-scale databases, notably in silico rather than wet
laboratory data. The results of the in silico experiments
generally require confirmation in laboratory experiments. However,
this remains an effective experimental approach for novel
developments in understanding carcinogenesis and makes it
convenient for further wet laboratory investigations.
References
|
1
|
van der Knaap JA and Verrijzer CP:
Undercover: Gene control by metabolites and metabolic enzymes.
Genes Dev. 30:2345–2369. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Saadi H, Seillier M and Carrier A: The
stress protein TP53INP1 plays a tumor suppressive role by
regulating metabolic homeostasis. Biochimie. 118:44–50. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hostetler K: Chapter 6
polyglycerophospholipids: Phosphatidylglycerol,
diphosphatidylglycerol and bis, (monoacylglycero) phosphate. New
Compr Biochem. 4:215–261. 1982. View Article : Google Scholar
|
|
4
|
Hatch GM: Cell biology of cardiac
mitochondrial phospholipids. Biochem Cell Biol. 82:99–112. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang M, Mileykovskaya E and Dowhan W:
Gluing the respiratory chain together. Cardiolipin is required for
supercomplex formation in the inner mitochondrial membrane. J Biol
Chem. 277:43553–43556. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Garrido C, Galluzzi L, Brunet M, Puig PE,
Didelot C and Kroemer G: Mechanisms of cytochrome C release from
mitochondria. Cell Death Differ. 13:1423–1433. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ott M, Zhivotovsky B and Orrenius S: Role
of cardiolipin in cytochrome c release from mitochondria. Cell
Death Differ. 14:1243–1247. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hostetler KY, Van den Bosch H and Van
Deenen LL: Biosynthesis of cardiolipin in liver mitochondria.
Biochim Biophys Acta. 239:113–119. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu B, Xu FY, Jiang YJ, Choy PC, Hatch GM,
Grunfeld C and Feingold KR: Cloning and characterization of a cDNA
encoding human cardiolipin synthase (hCLS1). J Lipid Res.
47:1140–1145. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Houtkooper RH, Akbari H, van Lenthe H,
Kulik W, Wanders RJ, Frentzen M and Vaz FM: Identification and
characterization of human cardiolipin synthase. FEBS Lett.
580:3059–3064. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Abdollahi A and Omranipour R: Is increase
of homocysteine, anti-cardiolipin, anti-phospholipid antibodies
associated with breast tumors? Acta Med Iran. 53:681–685.
2015.PubMed/NCBI
|
|
12
|
Schvartsman G, Ferrarotto R and Massarelli
E: Checkpoint inhibitors in lung cancer: Latest developments and
clinical potential. Ther Adv Med Oncol. 8:460–473. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Santarpia M, Giovannetti E, Rolfo C,
Karachaliou N, González-Cao M, Altavilla G and Rosell R: Recent
developments in the use of immunotherapy in non-small cell lung
cancer. Expert Rev Respir Med. 10:781–798. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cufer T, Ovcaricek T and O'Brien ME:
Systemic therapy of advanced non-small cell lung cancer:
Major-developments of the last 5-years. Eur J Cancer. 49:1216–1225.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: A cancer microarray database and integrated Data-mining
platform. Neoplasia. 6:1–6. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rhodes DR, Kalyana-Sundaram S, Mahavisno
V, Varambally R, Yu J, Briggs BB, Barrette TR, Anstet MJ,
Kincead-Beal C, Kulkarni P, et al: Oncomine 3.0: Genes, pathways
and networks in a collection of 18,000 cancer gene expression
profiles. Neoplasia. 9:166–180. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gyorffy B, Surowiak P, Budczies J and
Lánczky A: Online survival analysis software to assess the
prognostic value of biomarkers using transcriptomic data in
non-small-cell lung cancer. PLoS One. 8:e822412013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang da W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: Paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shirlaw JT: The metabolism of tumors. Br
Med J. 1:741931. View Article : Google Scholar
|
|
21
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Veech RL: The therapeutic implications of
ketone bodies: The effects of ketone bodies in pathological
conditions: Ketosis, ketogenic diet, redox states, insulin
resistance and mitochondrial metabolism. Prostaglandins Leukot
Essent Fatty Acids. 70:309–319. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Seyfried TN and Mukherjee P: Targeting
energy metabolism in brain cancer: Review and hypothesis. Nutr
Metab. 2:302005. View Article : Google Scholar
|
|
24
|
Wu M, Neilson A, Swift AL, Moran R,
Tamagnine J, Parslow D, Armistead S, Lemire K, Orrell J, Teich J,
et al: Multiparameter metabolic analysis reveals a close link
between attenuated mitochondrial bioenergetic function and enhanced
glycolysis dependency in human tumor cells. Am J Physiol Cell
Physiol. 292:C125–C136. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Galarraga J, Loreck DJ, Graham JF, DeLaPaz
RL, Smith BH, Hallgren D and Cummins CJ: Glucose metabolism in
human gliomas: Correspondence of in situ and in vitro metabolic
rates and altered energy metabolism. Metab Brain Dis. 1:279–291.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Baggetto LG, Clottes E and Vial C: Low
mitochondrial proton leak due to high membrane cholesterol content
and cytosolic creatine kinase as two features of the deviant
bioenergetics of Ehrlich and AS30-D tumor cells. Cancer Res.
52:4935–4941. 1992.PubMed/NCBI
|
|
27
|
Bergelson LD, Dyatlovitskaya EV, Sorokina
IB and Gorkova NP: Phospholipid compositon of mitochondria and
microsomes from regenerating rat liver and hepatomas of different
growth rate. Biochim Biophys Acta. 360:361–365. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chicco AJ and Sparagna GC: Role of
cardiolipin alterations in mitochondrial dysfunction and disease.
Am J Physiol Cell Physiol. 292:C33–C44. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hoch FL: Cardiolipins and biomembrane
function. Biochim Biophys Acta. 1113:71–133. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kiebish MA, Han X, Cheng H, Chuang JH and
Seyfried TN: Cardiolipin and electron transport chain abnormalities
in mouse brain tumor mitochondria: Lipidomic evidence supporting
the Warburg theory of cancer. J Lipid Res. 49:2545–2556. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hardy S, El-Assaad W, Przybytkowski E,
Joly E, Prentki M and Langelier Y: Saturated fatty acid-induced
apoptosis in MDA-MB-231 breast cancer cells. A role for
cardiolipin. J Biol Chem. 278:31861–31870. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mcmillin JB and Dowhan W: Cardiolipin and
apoptosis. Biochim Biophys Acta. 1585:97–107. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim EK and Choi EJ: Compromised MAPK
signaling in human diseases: An update. Arch Toxicol. 89:867–882.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim EK and Choi EJ: Pathological roles of
MAPK signaling pathways in human diseases. Biochim Biophys Acta.
1802:396–405. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang B, Zhou Z, Lin H, Lv X, Fu J, Lin P,
Zhu C and Wang H: Protein phosphatase 1A (PPM1A) is involved in
human cytotrophoblast cell invasion and migration. Histochem Cell
Biol. 132:169–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li R, Gong Z, Pan C, Xie DD, Tang JY, Cui
M, Xu YF, Yao W, Pang Q, Xu ZG, et al: Metal-dependent protein
phosphatase 1A functions as an extracellular signal-regulated
kinase phosphatase. FEBS J. 280:2700–2711. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Su PH, Lin YW, Huang RL, Liao YP, Lee HY,
Wang HC, Chao TK, Chen CK, Chan MW, Chu TY, et al: Epigenetic
silencing of PTPRR activates MAPK signaling, promotes metastasis
and serves as a biomarker of invasive cervical cancer. Oncogene.
32:15–26. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Blanco-Aparicio C, Torres J and Pulido R:
A novel regulatory mechanism of MAP kinases activation and nuclear
translocation mediated by PKA and the PTP-SL tyrosine phosphatase.
J Cell Biol. 147:1129–1136. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bhalla US, Ram PT and Iyengar R: MAP
kinase phosphatase as a locus of flexibility in a mitogen-activated
protein kinase signaling network. Science. 297:1018–1023. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bermudez O, Pagès G and Gimond C: The
dual-specificity MAP kinase phosphatases: Critical roles in
development and cancer. Am J Physiol Cell Physiol. 299:C189–C202.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Caunt CJ and Keyse SM: Dual-specificity
MAP kinase phosphatases (MKPs): Shaping the outcome of MAP kinase
signalling. FEBS J. 280:489–504. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ham JE, Oh EK, Kim DH and Choi SH:
Differential expression profiles and roles of inducible DUSPs and
ERK1/2-specific constitutive DUSP6 and DUSP7 in microglia. Biochem
Biophys Res Commun. 467:254–260. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Huo Y, Rangarajan P, Ling EA and Dheen ST:
Dexamethasone inhibits the Nox-dependent ROS production via
suppression of MKP-1-dependent MAPK pathways in activated
microglia. BMC Neurosci. 12:492011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zeke A, Misheva M, Reményi A and
Bogoyevitch MA: JNK signaling: Regulation and functions based on
complex protein-protein partnerships. Microbiol Mol Biol Rev.
80:793–835. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Segalés J, Perdiguero E and Muñoz-Cánoves
P: Regulation of muscle stem cell functions: A focus on the p38
MAPK signaling pathway. Front Cell Dev Biol. 4:912016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chang L and Karin M: Mammalian MAP kinase
signalling cascades. Nature. 410:37–40. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Schrauzer GN, White DA and Schnieder CJ:
Cancer mortality correlation studies-III: Statistical associations
with dietary selenium intakes. Bioinorg Chem. 7:23–31. 1997.
View Article : Google Scholar
|
|
48
|
Willis MS and Wians FH: The role of
nutrition in preventing prostate cancer: A review of the proposed
mechanism of action of various dietary substances. Clin Chim Acta.
330:57–83. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nyman DW, Stratton M Suzanne, Kopplin MJ,
Dalkin BL, Nagle RB and Jay Gandolfi A: Selenium and
selenomethionine levels in prostate cancer patients. Cancer Detect
Prev. 28:8–16. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Morey M, Corominas M and Serras F: DIAP1
suppresses ROS-induced apoptosis caused by impairment of the
selD/sps1 homolog in Drosophila. J Cell Sci. 116:4597–4604. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chung HJ, Yoon SI, Shin SH, Koh YA, Lee
SJ, Lee YS and Bae S: p53-Mediated enhancement of radiosensitivity
by selenophosphate synthetase 1 overexpression. J Cell Physiol.
209:131–141. 2006. View Article : Google Scholar : PubMed/NCBI
|