Introduction
Lung cancer remains a leading cause of
cancer-associated mortalities worldwide, despite considerable
progress in the development of therapies. The prevalence and
mortality rates of lung cancer have increased worldwide (1–3). The
detection and pathological research of lung cancer are important in
the diagnosis and therapy of cancer diseases (4,5). In view
of this, it is necessary to elucidate the molecular mechanisms
underlying cell proliferation and apoptosis in lung cancer
cells.
The imbalance between cell proliferation and
apoptosis has been implicated to serve key roles in cancer
pathogenesis (6,7). Accumulating evidence has indicated that
microRNAs (miRNAs or miRs) serve a role in a number of cancer
processes, including lung cancer (8–10). The
results of previous studies have demonstrated that targeting or
regulating miRNAs may identify novel ways to effectively overcome
lung cancer resistance and personalize lung cancer therapy
(11,12). Therefore, as a type of small
non-coding RNA, miRNA potentially serves roles in tumorigenesis and
has attracted increasing attention.
Van Keuren-Jensen et al (8) recently revealed that miR-122-3p was
significantly downregulated between early and advanced fibrosis of
the liver. Another recent study by Wang et al (13) demonstrated that miR-122-3p exhibited
significant association with either plasma cytokine and chemokine
levels, or clinical features in patients with rheumatoid arthritis.
Increasing evidences have indicated that miR-122-3p was
downregulated in a number of types of cancer in humans, which
suggested that miR-122-3p may serve a key role in tumorigenesis
(14).
p27 and p21 are cell cycle regulators, which are
reported to be associated with numerous types of tumor cell cycles,
including liver, cervical and gastric cancer (15–17).
Although these studies have demonstrated that p27 and p21 serve
important roles in cell proliferation regulation, the mechanism
underlying lung cancer remains to be elucidated.
Furthermore, Forkhead box O (FOXO) proteins belong
to a family of proteins including FOXO1, FOXO3a, FOXO4 and FOXO6 in
humans. Accumulating evidences have suggested that FOXO proteins
function as tumor suppressors (18,19).
The aim of the present study was to clarify the role
of miR-122-3p in tumor growth of lung cancer cells. In the present
study, A549 cells were transfected with control, miR-122-3p mimic
or miR-122-3p inhibitor to affect miR-122-3p expression.
Subsequently, an MTT assay was performed to detect cell
proliferation. The results revealed that overexpression of
miR-122-3p suppressed lung cancer cell proliferation. Subsequently,
5-bromo-2-deoxyuridine (BrdU) staining assay was performed to
further investigate the mechanisms underlying miR-122-3p-induced
cell growth inhibition. In addition, p27 was revealed to be
upregulated by miR-122-3p. Finally, the present study analyzed
FOXO, bim, and pro- and activated caspase-3 expression levels to
clarify the mechanism underlying the effect of miR-122-3p on cell
apoptosis regulation. The results demonstrated that miR-122-3p
induced apoptosis by targeting FOXO in A549 cells. The present
study provides theoretical basis and a new insight into the
treatment of lung cancer.
Materials and methods
Cell culture
A549 cells were obtained from American Type Culture
Collection (Manassas, VA, USA) and cultured in Dulbecco's modified
Eagle's medium (DMEM; Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C in 5%
CO2. The cells were transfected with mature miR-122-3p
mimic (5′-AACAGCACAAACUACUACCUCA-3′) or miR-122-3p inhibitor
(5′-UAUUUAGUGUGAUAAUGGCGUU-3′; both from Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. U6 (5′-AACGCTTCACGAATTTGCGT-3′;
Sigma-Aldrich; Merck KGaA) was selected as a miRNA control. In
brief, miR-122-3p mimic or inhibitor and Lipofectamine®
2000 reagent, diluted with 50 µl antibiotic-free DMEM, were mixed
and incubated for 20 min at room temperature. The cells
(5×103 cells/well) were seeded onto a 96-well plate and
cultured with 400 µl antibiotic-free DMEM. Then, the mixture was
added to the cell culture plates and incubated at 37°C in 5%
CO2. Following 48 h transfection, the stably transfected
cells were maintained for ~4 weeks by the culture medium containing
0.5 mg/ml G418 (Sigma-Aldrich; Merck KGaA). G418-resistant cells
were directly collected for use in subsequent experiments.
Cell proliferation and MTT assay
A549 cells were digested with 0.25% trypsin for 3
min at room temperature, re-suspended in 3 ml culture medium and
counted using a hemocytometer following exposure to Danshen extract
(Xi'an Feida Bio-tech Co., Xi'an, China) every 2 h for 12 h (a
total of 6 times). For the MTT assay, 5×103 cells/well
were seeded onto 96-well culture plates, exposed to 5 µg/ml Danshen
and incubated for 2 h at 37°C. Cell viability was then analyzed by
adding 20 µl 10 mg/ml MTT (Sigma-Aldrich) to 0.2 ml culture medium
and incubated for 3 h at 37°C. Following removal of the medium,
formazan (in dimethylsulfoxide) was added, and the 590 nm optical
density was recorded using a Multiskan EX (Thermo Fisher
Scientific, Inc.) (20).
BrdU assay
Cells were seeded in 6-well plates at a density of
2×104 cells/well on sterilized coverslips for 72 h at
37°C. BrdU (Sigma-Aldrich; 10 µM) was added to the medium and
incubated at 37°C for 5 h. Cells were fixed in 70% ethanol for 5
min at 4°C followed by addition of 1.5 M HCl to the cells for 30
min at room temperature to denature DNA. Immunofluorescence to
visualize incorporated BrdU was performed by using a mouse
anti-BrdU antibody (cat no. 555627; 1:1,000; BD Biosciences, San
Jose, CA, USA) to incubate cells for 1 h at 37°C. Then, incubation
with secondary green-fluorescence dye-conjugated antibody
(Alexafluor® 594; 1:1,000; Invitrogen; Thermo Fisher
Scientific, Inc.) was subsequently performed for 45 min at 37°C.
VECTASHIELD mounting medium with DAPI (Vector Laboratories, Inc.,
Burlingame, CA, USA) was used to stain the nuclei for 5 min at room
temperature. Images were visualized using a Leica inverted fully
automated microscope (DMI6000B) at magnification, ×40 with a
digital camera DFC 420 RGB (Leica Microsystems GmbH, Wetzlar,
Germany) (21). The Brdu labeling
index (LI) was the number of Brdu-positive cells/the total number
of cells in 10 fields and analyzed using SPSS version 19.0 (IBM
Corp., Armonk, NY, USA).
Apoptosis assay
At 24 h following transfection, cells were harvested
and suspended in Annexin-binding buffer (Thermo Fisher Scientific,
Inc.) and then incubated with Annexin V-FITC Apoptosis Detection
kit (Sigma-Aldrich; Merck KGaA) for 30 min in the dark at 25°C.
Apoptotic cells were subsequently analyzed using a FACS can
(Beckman Coulter, Inc., Brea, CA, USA) and FlowJo version 10.4
(FlowJo LLC, Ashland, OR, USA) (6).
Western blot analysis
Cells were washed once with PBS and lysed in a
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) supplemented with
Phenylmethanesulfonyl fluoride (Beyotime Institute of
Biotechnology) for 30 min at 0°C. Protein samples were measured the
concentrations using Bradford Protein Assay kit (Beyotime Institute
of Biotechnology) for 10 min at room temperature, boiled for 10 min
in SDS sample buffer, then separated by SDS-PAGE and transferred to
a nitrocellulose membrane. The membranes were incubated with
primary antibodies (all dilution 1:1,000) overnight at 4°C
following blocking with 5% skim milk in TBS Tween-20 (TBST) for 1 h
at room temperature. Membranes were washed once with TBST and
incubated with horseradish peroxidase-conjugated secondary
anti-rabbit antibody (cat. no., ab191866; 1 mg/ml; Abcam,
Cambridge, UK) for 2 h at room temperature. WEST-ZOL-plus Western
Blot Detection system (iNtRON Biotechnology, Sungnam, Korea) was
used to visualize the protein bands. Antibodies specific to actin
(cat no. 8457) were purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA) and served as the internal control. Activated
caspase-3 (cat no. ab2302), p27 (cat no. ab54563), p21 (cat no.
ab7960) and pro-caspase-3 (cat no. ab32150) antibodies were
obtained from Abcam. Bim (cat no. 2933), FOXO1 (cat no. 2880),
FOXO3 (cat no. 2497) and phosphorylated FOXO1/3 (cat no. 9464)
antibodies were purchased from Cell Signaling Technology, Inc.
(22).
Statistical analysis
All experiments were repeated three times. Data are
presented as the mean ± standard deviation (SD). SPSS version 19.0
(IBM Corp.) was used for statistical analysis of data. Statistical
significance was evaluated by one-way analysis of variance for
multi-group comparisons with Bonferroni analyses or Student's
t-test for two group comparisons. P<0.05 was considered to
indicate a statistically significant difference.
Results
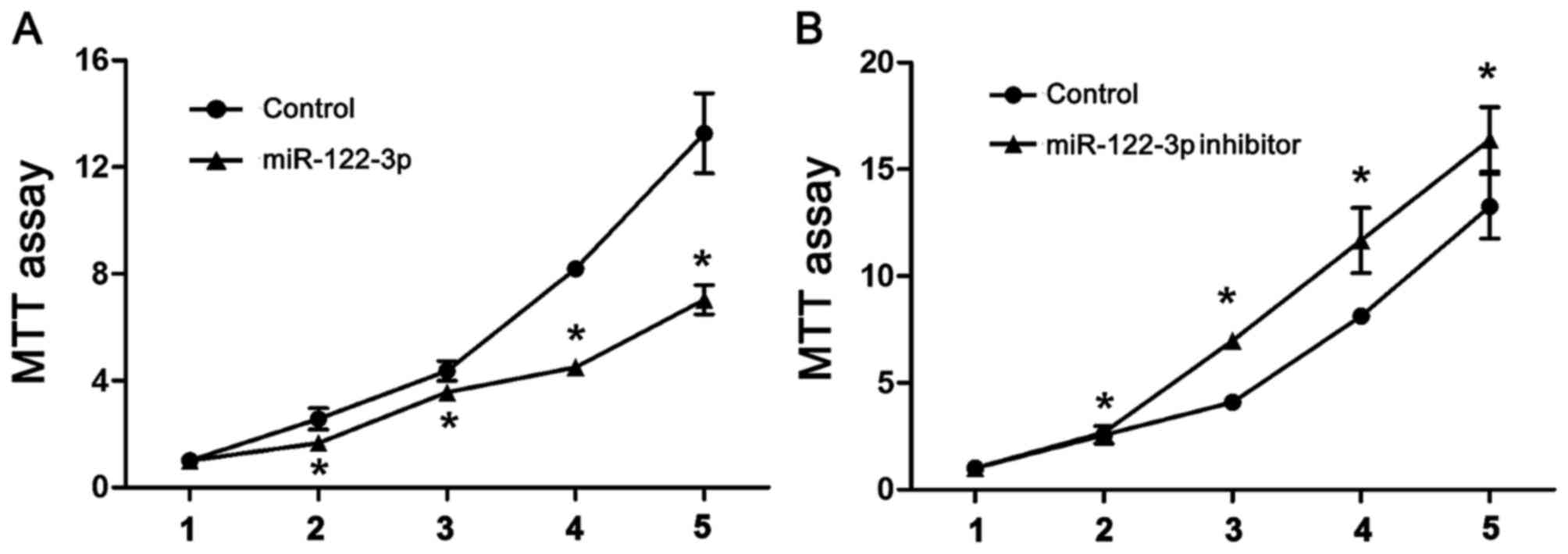
Overexpression of miR-122-3p reduces
cell proliferation in A549 cells
To analyze the effect of miR-122-3p on cell
proliferation, A549 cells were transfected with the control (U6),
miR-122-3p mimic or miR-122-3p inhibitor, and an MTT assay was
performed. As demonstrated in Fig. 1,
miR-122-3p overexpression significantly inhibited the proliferation
of A549 cells compared with controls. Correspondingly, cells
transfected with miR-122-3p inhibitor significantly increased A549
cell proliferation compared with controls (P<0.05).
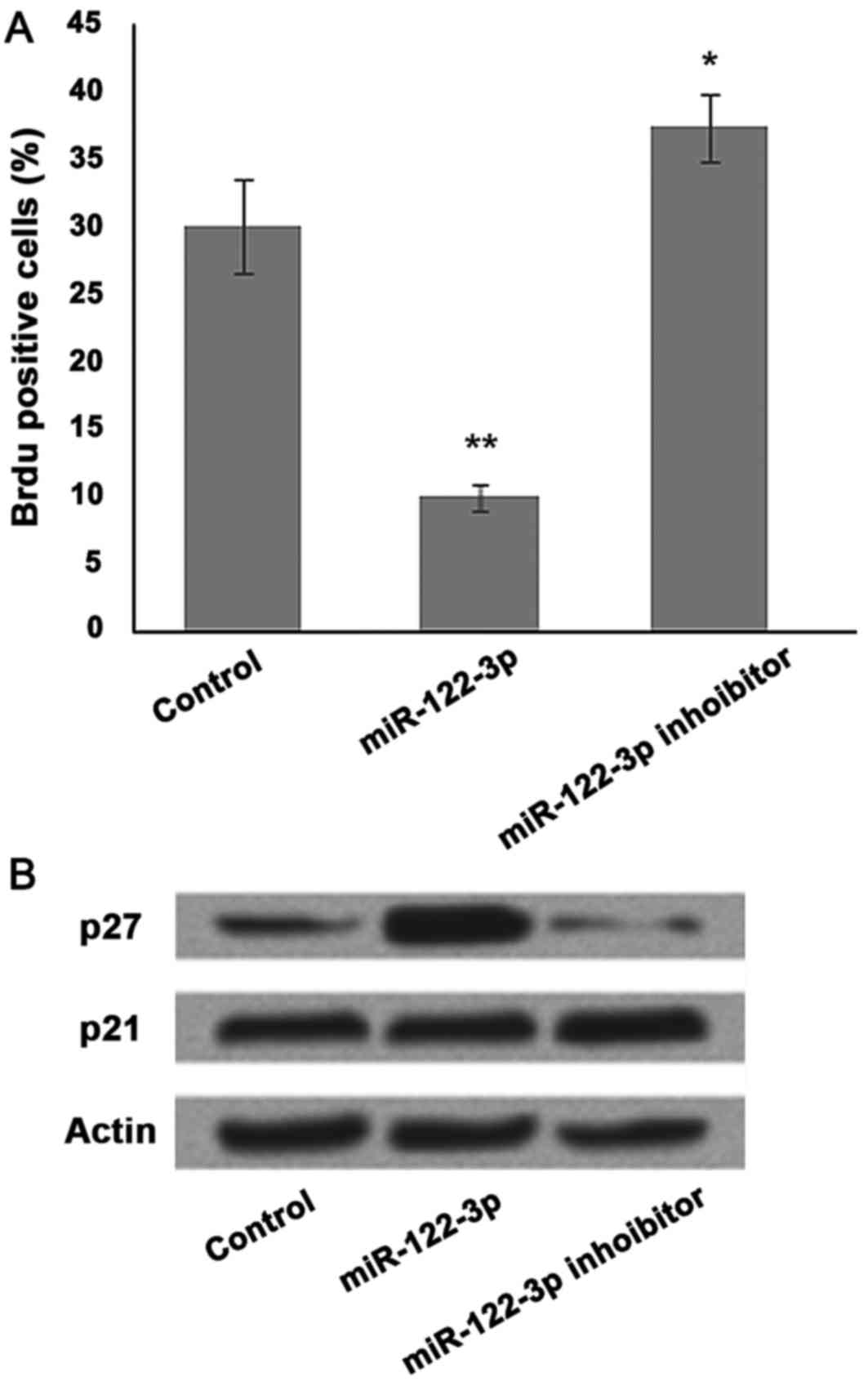
miR-122-3p overexpression induces cell
growth arrest via upregulation of p27 in A549 cells
To analyze the effect of miR-122-3p on cell
proliferation, A549 cells were transfected with the control,
miR-122-3p mimic or inhibitor, and BrdU staining was performed. To
further investigate the mechanisms underlying miR-122-3p-induced
cell growth inhibition, the present study analyzed the effect of
miR-122-3p on various cell cycle regulators in A549 cells. BrdU
positive cells were significantly decreased in the miR-122-3p mimic
group and increased following miR-122-3p suppression compared with
the control group (P<0.05 and P<0.01, respectively; Fig. 2A). Marked accumulation of p27 was
detected in response to miR-122-3p expression. Conversely, p21
expression was not markedly affected by miR-122-3p expression
(Fig. 2B).
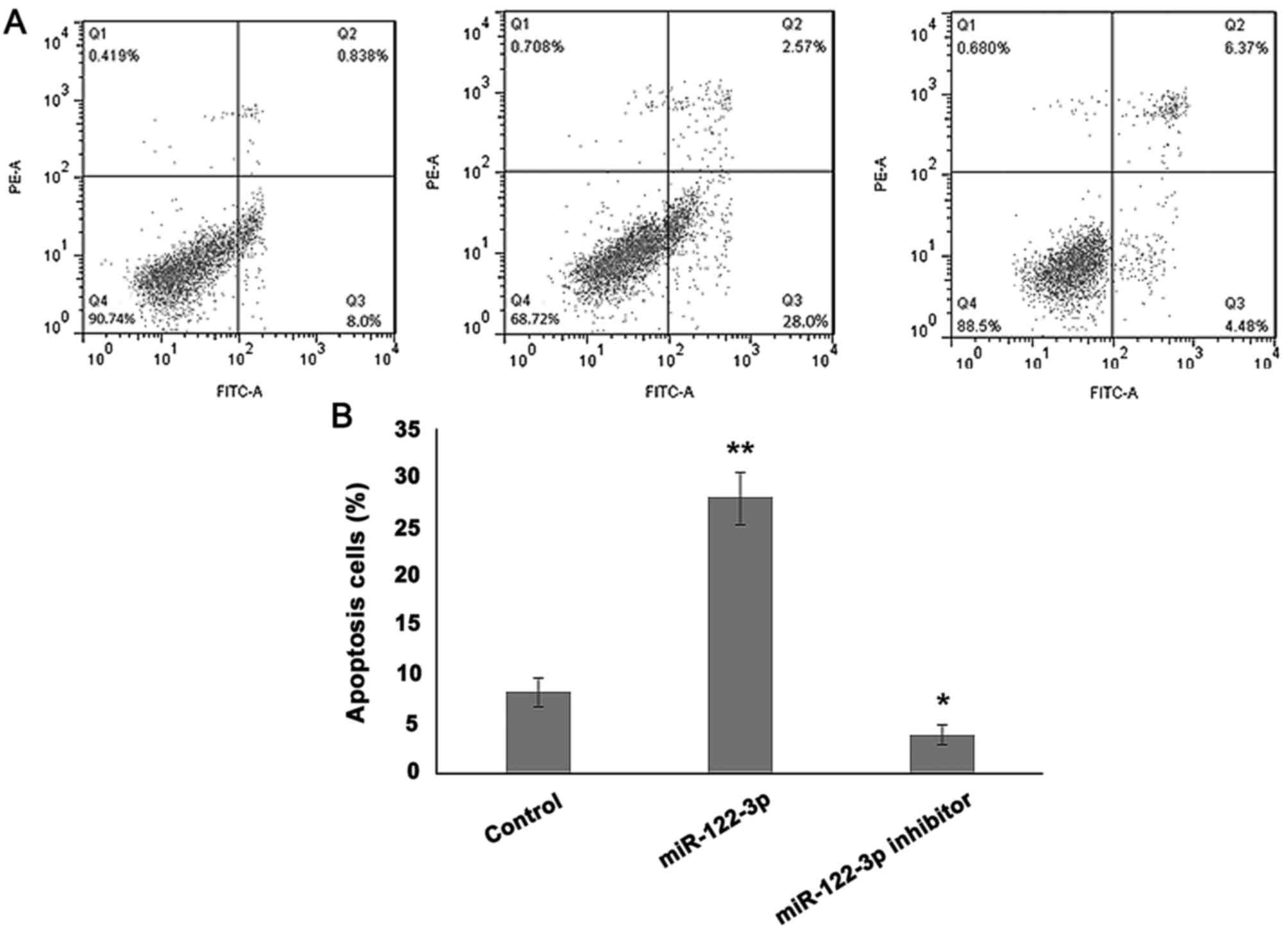
Overexpression of miR-122-3p induces
apoptosis in A549 cells
To analyze the effect of miR-122-3p on cell death,
A549 cells were transfected with the control, miR-122-3p mimic or
inhibitor, and an apoptosis assay was performed. As presented in
Fig. 3, overexpression of miR-122-3p
significantly increased the rate of apoptosis compared with the
control group (P<0.01), while miR-122-3p suppression decreased
apoptosis in A549 cells (P<0.05).
miR-122-3p induces apoptosis by
targeting FOXO in A549 cells
To further investigate the mechanisms underlying the
effect of miR-122-3p on apoptosis, A549 cells were transfected with
the control, miR-122-3p mimic or inhibitor, and the expression
levels of FOXO, bim, and pro- and activated caspase-3 were
evaluated (Fig. 4). The results
confirmed that A549 cell apoptosis was associated with the
expression of FOXO. The western blot results of FOXO, bim, pro and
activated caspase-3 expressions demonstrated that miR-122-3p
promoted cell apoptosis by targeting FOXO.
Discussion
As one of the most common types of malignant tumors
worldwide, lung cancer research has attracted increasing attention
in research; however, there is currently no effective treatment
strategy (23,24). miRNA is a type of non-coding RNA
molecule that inhibits the expression of target genes at multiple
stages (25). As one of the most
important regulators in the pathogenesis of human diseases, there
are between 800 and 1,000 known miRNAs in the human genome that
serve important roles in cancer cell progress regulation via the
regulation of gene expression (25,26).
Certain miRNAs have been reported to serve key roles in the
progression of human lung cancer, including miR-145 and miR-21
(27,28).
The present study investigated the association
between miR-122-3p and lung cancer, and revealed that miR-122-3p
was associated with lung cancer cell proliferation. Furthermore,
cell apoptosis was also an important mechanism associated with the
progression of cancer. Therefore, a cell apoptosis assay was
performed, which confirmed that miR-122-3p induced lung cancer
apoptosis.
p27 and p21 are cell cycle regulators that are
associated with numerous types of disease, demonstrating prognostic
importance (29). Skirnisdottir and
Seidal (30) previously evaluated the
prognostic value of p21 alone and in combination with p27, and
revealed that patients with non-serous tumors and concomitant p21
and p27 positivity patients exhibited excellent survival rates. In
another study, p21 and p27 were reported to be associated with the
cellular proliferation of lung cancer (31).
To fully elucidate the underlying mechanism of
miR-122-3p on cancer cell proliferation of lung cancer, the present
study performed BrdU staining and western blot analysis to evaluate
the expression levels of p21 and p27. The results demonstrated that
p27 was markedly accumulated in response to miR-122-3p expression,
whereas p21 was not affected by miR-122-3p expression. Therefore,
the present study concluded that miR-122-3p inhibits cell
proliferation of lung cancer by upregulating p27 expression.
FOXO proteins, as tumor suppressors, have been
confirmed to regulate a number of cellular functions, including
cell cycle, apoptosis, DNA damage repair and energy metabolism
(32). In addition, FOXO
overexpression and nuclear accumulation have been reported to be
associated with reduced tumor growth, controlling cell homeostasis
and cell fate (33).
In the present study, A549 cells were transfected
with the control, miR-122-3p mimic or inhibitor, and the expression
levels of FOXO, bim, and pro- and activated caspase-3 were
determined. Apoptosis assay and western blot analysis were also
performed. The results confirmed that miR-122-3p promotes cell
apoptosis by targeting FOXO.
In summary, miR-122-3p inhibits cell proliferation
by upregulating p27 expression and induces cell apoptosis by
targeting FOXO in A549 lung cancer cells. These findings suggest
that miR-122-3p may be associated with the progression of lung
cancer and be a new therapeutic target for this disease.
References
|
1
|
Ferlay J, Steliarova-Foucher E,
Lortet-Tieulent J, Rosso S, Coebergh JW, Comber H, Forman D and
Bray F: Cancer incidence and mortality patterns in Europe:
Estimates for 40 countries in 2012. Eur J Cancer. 49:1374–1403.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Utzschneider S, Wicherek E, Weber P,
Schmidt G, Jansson V and Dürr HR: Surgical treatment of bone
metastases in patients with lung cancer. Int Orthop. 35:731–736.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ávila-Moreno F, Armas-López L,
Álvarez-Moran AM, López-Bujanda Z, Ortiz-Quintero B,
Hidalgo-Miranda A, Urrea-Ramírez F, Rivera-Rosales RM,
Vázquez-Manríquez E, Peña-Mirabal E, et al: Overexpression of MEOX2
and TWIST1 is associated with H3K27me3 levels and determines lung
cancer chemoresistance and prognosis. PLoS One. 9:e1141042014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tan C, Chen H and Wu T: Classification
models for detection of lung cancer based on nine element
distribution of urine samples. Biol Trace Elem Res. 142:18–28.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ironmonger L, Ohuma E, Ormiston-Smith N,
Gildea C, Thomson CS and Peake MD: An evaluation of the impact of
large-scale interventions to raise public awareness of a lung
cancer symptom. Br J Cancer. 112:207–216. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang N, Su Y and Xu L: Targeting PKCε by
miR-143 regulates cell apoptosis in lung cancer. FEBS Lett.
587:3661–3667. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han SW and Roman J: Targeting apoptotic
signaling pathways in human lung cancer. Curr Cancer Drug Targets.
10:566–574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Van K, euren-Jensen KR, Malenica I,
Courtright AL, Ghaffari LT, Starr AP, Metpally RP, Beecroft TA,
Carlson EW, Kiefer JA, Pockros PJ and Rakela J: microRNA changes in
liver tissue associated with fibrosis progression in patients with
hepatitis C. Liver Int. 36:334–343. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maftouh M, Avan A, Galvani E, Peters GJ
and Giovannetti E: Molecular mechanisms underlying the role of
microRNAs in resistance to epidermal growth factor
receptor-targeted agents and novel therapeutic strategies for
treatment of non-small-cell lung cancer. Crit Rev Oncog.
18:317–326. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rolfo C, Fanale D, Hong DS, Tsimberidou
AM, Piha-Paul SA, Pauwels P, Van Meerbeeck JP, Caruso S, Bazan V,
Cicero G, et al: Impact of microRNAs in resistance to chemotherapy
and novel targeted agents in non-small cell lung cancer. Curr Pharm
Biotechnol. 15:475–485. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gong Z, Yang J, Li J, Yang L, Le Y, Wang S
and Lin HK: Novel insights into the role of microRNA in lung cancer
resistance to treatment and targeted therapy. Curr Cancer Drug
Targets. 14:241–258. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
MacDonagh L, Gray SG, Finn SP, Cuffe S,
O'Byrne KJ and Barr MP: The emerging role of microRNAs in
resistance to lung cancer treatments. Cancer Treat Rev. 41:160–169.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang W, Zhang Y, Zhu B, Duan T, Xu Q, Wang
R, Lu L and Jiao Z: Plasma microRNA expression profiles in Chinese
patients with rheumatoid arthritis. Oncotarget. 6:42557–42568.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi Y and Huang A: Effects of sorafenib on
lung metastasis in rats with hepatocellular carcinoma: The role of
microRNAs. Tumour Biol. 36:8455–8463. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim JH, Choi YK, Byun JK, Kim MK, Kang YN,
Kim SH, Lee S, Jang BK and Park KG: Estrogen-related receptor γ is
upregulated in liver cancer and its inhibition suppresses liver
cancer cell proliferation via induction of p21 and p27. Exp Mol
Med. 48:e2132016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Thacker PC and Karunagaran D: Curcumin and
emodin down-regulate TGF-β signaling pathway in human cervical
cancer cells. PLoS One. 10:e01200452015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang E, Yin D, Han L, He X, Si X, Chen W,
Xia R, Xu T, Gu D, De W, et al: E2F1-induced upregulation of long
noncoding RNA LINC00668 predicts a poor prognosis of gastric cancer
and promotes cell proliferation through epigenetically silencing of
CKIs. Oncotarget. 7:23212–23226. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang H and Tindall DJ: Regulation of FOXO
protein stability via ubiquitination and proteasome degradation.
Biochim Biophys Acta. 1813:1961–1964. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang F, Chan CH, Chen K, Guan X, Lin HK
and Tong Q: Deacetylation of FOXO3 by SIRT1 or SIRT2 leads to
Skp2-mediated FOXO3 ubiquitination and degradation. Oncogene.
31:1546–1557. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu L, Li C, Li D, Wang Y, Zhou C, Shao W,
Peng J, You Y, Zhang X and Shen X: Cryptotanshinone inhibits human
glioma cell proliferation by suppressing STAT3 signaling. Mol Cell
Biochem. 381:273–282. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fernandez S, Risolino M, Mandia N, Talotta
F, Soini Y, Incoronato M, Condorelli G, Banfi S and Verde P:
miR-340 inhibits tumor cell proliferation and induces apoptosis by
targeting multiple negative regulators of p27 in non-small cell
lung cancer. Oncogene. 34:3240–3250. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nam KS, Oh S, Lee KM, Yoo SA and Shin I:
CD44 regulates cell proliferation, migration, and invasion via
modulation of c-Src transcription in human breast cancer cells.
Cell Signal. 27:1882–1894. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Zhao J, Qiu L, Zhang P, Li J,
Yang D, Wei X, Han Y, Nie S and Sun Y: Co-expression of ILT4/HLA-G
in human non-small cell lung cancer correlates with poor prognosis
and ILT4-HLA-G interaction activates ERK signaling. Tumor Biol.
37:11187–11198. 2016. View Article : Google Scholar
|
|
24
|
Lysov Z, Dwivedi DJ, Gould TJ and Liaw PC:
Procoagulant effects of lung cancer chemotherapy: Impact on
microparticles and cell-free DNA. Blood Coagul Fibrinolysis.
28:72–82. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yongchun Z, Linwei T, Xicai W, Lianhua Y,
Guangqiang Z, Ming Y, Guanjian L, Yujie L and Yunchao H:
MicroRNA-195 inhibits non-small cell lung cancer cell
proliferation, migration and invasion by targeting MYB. Cancer
Lett. 347:65–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Koturbash I, Zemp FJ, Pogribny I and
Kovalchuk O: Small molecules with big effects: The role of the
microRNAome in cancer and carcinogenesis. Mutat Res. 722:94–105.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hu J, Qiu M, Jiang F, Zhang S, Yang X,
Wang J, Xu L and Yin R: MiR-145 regulates cancer stem-like
properties and epithelial-to-mesenchymal transition in lung
adenocarcinoma-initiating cells. Tumour Biol. 35:8953–8961. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu L, Huang Y, Chen D, He J, Zhu W, Zhang
Y and Liu X: Downregulation of miR-21 increases cisplatin
sensitivity of non-small-cell lung cancer. Cancer Genet.
207:214–220. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang M, Li J, Wang L, Tian Z, Zhang P, Xu
Q, Zhang C, Wei F and Chen W: Prognostic significance of p21, p27
and survivin protein expression in patients with oral squamous cell
carcinoma. Oncol Lett. 6:381–386. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Skirnisdottir I and Seidal T: Association
of p21, p21 p27 and p21 p53 status to histological subtypes and
prognosis in low-stage epithelial ovarian cancer. Cancer Genomics
Proteomics. 10:27–34. 2013.PubMed/NCBI
|
|
31
|
Li ZC, Zhang LM, Wang HB, Ma JX and Sun
JZ: Curcumin inhibits lung cancer progression and metastasis
through induction of FOXO1. Tumour Biol. 35:111–116. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shukla S, Rizvi F, Raisuddin S and Kakkar
P: FoxO proteins' nuclear retention and BH3-only protein Bim
induction evoke mitochondrial dysfunction-mediated apoptosis in
berberine-treated HepG2 cells. Free Radic Biol Med. 76:185–199.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bucur O, Stancu AL, Muraru MS, Melet A,
Petrescu SM and Khosravifar R: PLK1 is a binding partner and a
negative regulator of FOXO3 tumor suppressor. Discoveries
(Craiova). 2:pii: e162014. View Article : Google Scholar
|