Introduction
Ovarian cancer is one of the most common
gynecological malignancies and the incidence rate has been
increasing annually (1). The most
prevalent histological type is the epithelial ovarian carcinoma
(EOC) which represents 85% of ovarian carcinoma (2). Despite advances in surgery and
chemotherapy in the last decades, the overall survival of patients
suffering from EOC is not satisfactory, with a 5-year survival rate
of only 30% (3). Such a poor
prognosis of patients with EOC has been largely correlated with
tumor metastasis. Thus, it is urgently needed to understand the
molecular mechanisms underlying ovarian cancer development and
progression.
Epithelial-mesenchymal transition (EMT) is an
important mechanism leading to invasion and metastasis of various
cancers (4–6). Cells undergoing EMT lose the epithelial
features and acquire some mesenchymal features, which is associated
with the upregulation of N-cadherin and downregulation of
E-cadherin (6). EMT causes the
failure of the intercellular connection that may facilitate the
cancer cell to pass through basement membrane (7). E-cadherin is also known as cadherin 1
(CDH1) and crucial for sustaining the polarity and structure of
normal epithelial cell (8). Reduction
of E-cadherin on the cell surface is clearly associated with poor
overall survival of patients with EOC (9).
Colon cancer-associated transcript 2 (CCAT2), a
novel lncRNA transcript reported by Ling et al, is located
on 8q24 and highly overexpressed in microsatellite-stable
colorectal cancer (10). Recently,
Qiu et al reported that CCAT2 was highly expressed in
non-small cell lung cancer (NSCLC). And the proliferation and
invasion of NSCLC cell lines were inhibited in vitro after
CCAT2 was silenced (11). Redis et
al found that CCAT2 appeared to have higher expression in
breast cancer tissue than in non-tumor tissue. Additionally, they
observed that CCAT2 promoted cell migration and downregulated
chemosensitivity to 5′FU in breast cancer (12). These studies indicated that CCAT2
acted as an oncogene in various human cancers. However, the
detailed function and potential molecular mechanisms of CCAT2 in
EOC are not fully understood and need to be investigated.
In the present study, we found that knockdown of
CCAT2 inhibited EMT by upregulating E-cadherin and downregulating
N-cadherin, Slug and Twist1 in SKOV3 and A2780. Furthermore, our
results indicate that CCAT2 may exert its biological functions
partly through Wnt/β-catenin signaling pathway in the ovarian
cancer cells. Therefore, the study revealed the important roles as
well as the potential mechanisms of CCAT2 in ovarian cancer
progression.
Materials and methods
Cell lines and cell culture
Two human EOC cell lines (SKOV3, A2780) were
purchased from the Institute of Biochemistry and Cell Biology of
Chinese Academy of Science (Shanghai, China). HO8910 was kindly
provided by the Key Laboratory, Harbin Medical University, Ministry
of Education (Heilongjiang, China). The EOC cell lines were
cultured in RPMI-1640 medium (Corning Inc., Corning, NY, USA)
containing 10% fetal bovine serum (FBS; Gibco Life Technologies,
Carlsbad, CA, USA), penicillin (100 U/ml) and streptomycin (100
mg/ml) (both from Beyotime Institute of Biotechnology, Jiangsu,
China). Human ovarian surface epithelial (HOSE) cell HUM-CELL-0088
was obtained from PriCells Biomedical Technology Co., Ltd. (Wuhan,
China). HUM-CELL-0088 was cultured in DMEM (Corning, Inc.) with 10%
FBS, penicillin (100 U/ml) and streptomycin (100 mg/ml). All of the
above cells were maintained in a humidified 5% CO2
incubator at 37°C.
Total RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was isolated from cell lines using TRIzol
reagent (Invitrogen, Auckland, New Zealand). RNA was reversely
transcribed into cDNAs using a Transcriptor First Strand cDNA
Synthesis kit (Roche Diagnostics, Indianapolis, IN, USA) according
to the manufacturer's instructions. qPCR reactions performed using
a qPCR System (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and
SYBR-Green PCR Master Mix (Roche Diagnostics). The primers used
were: CCAT2, 5′-CCAGGCAATAACTGTGCAACTC-3′ (sense) and
5′-ACTTACGTAGGGCATGCCAAA-3′ (antisense); E-cadherin (CDH1),
5′-TGCCCAGAAAATGAAAAAGG-3′ (sense) and 5′-GTGTATGTGGCAATGCGTTC-3′
(antisense); N-cadherin (CDH2), 5′-ACAGTGGCCACCTACAAAGG-3′
(sense) and 5′-CCGAGATGGGGTTGATAATG-3′ (antisense); Slug
(SNAI2), 5′-TTCGGACCCACACATTACCT-3′ (sense) and
5′-GCAGTGAGGGCAAGAAAAAG-3′ (antisense); Twist1 (TWIST1),
5′-GGAGTCCGCAGTCTTACGAG-3′ (sense) and 5′-TCTGGAGGACCTGGTAGAGG-3′
(antisense); glyceraldehyde 3-phosphate dehydrogenase
(GAPDH), 5′-GAGTCAACGGATTTGGTCGT-3′ (sense) and
5′-GACAAGCTTCCCGTTCTCAG-3′ (antisense). Relative expression was
calculated using the 2−ΔΔCt method. Each qPCR
amplification was performed in triplicate to verify the
results.
Small interfering RNAs (siRNAs) and
transfection
For the study in vitro, SKOV3 and A2780 cells
cultured on 6-well plate were transfected with either siRNAs
targeting CCAT2 or negative controls (GeneChem, Shanghai, China)
using lentivirus as vector (hU6-MCS-Ubiquitin-EGFP-IRES-puromycin)
according to the instructions provided by the manufacturer. The
siRNA sequences were as follows: 5′-AGGUGUAGCCAGAGUUAAUTT-3′
(sense) and 5′-AUUAACUCUGGCUACACCUTT-3′ (antisense). Cells were
harvested for RT-qPCR and other experiments at 48 h after
transfection.
Monolayer wound healing assay
Migration ability was measured using the wound
healing assay. SKOV3 and A2780 were grown in 6-well plates. The
cells were transfected with either siRNAs targeting CCAT2 or a
negative control (NC). When cell reached 80% density, wounds were
created in confluent cells using a 100 µl pipette tip and then
cells incubated in fresh medium for 48 h. Three different locations
were observed and photographed with an inverted phase contrast
microscope (10X objective; Nikon, Japan, Tokyo). Each experiment
was repeated at last three times.
Transwell invasion assay
Cell invasion assay was performed using Transwell
chambers (8 µm pore size; Corning Life Sciences, Tewksbury, MA,
USA). For this assay, 100 µl Matrigel (BD Biosciences, San Jose,
CA, USA) was coated onto the upper chamber. The infected cells
(5×104) were seeded in the upper chamber of the wells in
100 µl FBS-free medium, while the lower chambers were filled with
600 µl 20% FBS medium. Following incubation for 24 h, the top layer
of the insert was scrubbed with a sterile cotton swab to remove any
remaining cells. The invading cells on the bottom surface were
fixed with methanol for 30 min, stained with 0.1% crystal violet
for 10 min and imaged using digital microscopy (10X objective;
Nikon). Cell numbers were calculated in five random fields for each
chamber at least and then the average value was calculated.
Western blot analysis
The harvested cells were disrupted in RIPA lysis
buffer (Beyotime Institute of Biotechnology) containing protease
inhibitor [1 mM phenylmethylsulfonyl fluoride (PMSF)]. After
centrifugation, the supernatant fraction was harvested as the total
cellular protein extract. The nuclear and cytoplasmic protein was
extracted using Nuclear and cytoplasmic protein extraction kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's instructions. The protein concentration was
calculated using the Pierce BCA protein assay kit (Beyotime
Institute of Biotechnology). The cellular protein was separated by
sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) and
transferred to polyvinylidene fluoride (PVDF) membranes (Invitrogen
Life Technologies, Carlsbad, CA, USA). After blocking with 5%
fat-free milk for 1 h, the membrane was incubated with primary
antibodies against E-cadherin (1:200 dilution, ab219332),
N-cadherin (1:200 dilution, ab12221) (both from Abcam, Cambridge,
MA, USA), Slug (1:500 dilution, C19G7), Twist1 (1:500 dilution,
46702), β-catenin (1:500 dilution, 9562) (all from Cell Signaling
Technology, Inc., Danvers, MA, USA), α-tubulin (1:2,000 dilution,
ab7291), Lamin B1 (1:2,000 dilution, ab16048) and β-actin (1:2,000
dilution, ab8226) (all from Abcam) overnight at 4°C and then blots
were washed 3 times in 1X Tris-Buffered Saline with 0.5% Tween
(TBS-T), followed by incubation with an appropriate secondary
antibody (1:5,000 dilution, ab6721 or ab6728; Abcam) for 1 h. The
blots were visualized using an enhanced chemiluminescence kit
(Beyotime Institute of Biotechnology). α-Tubulin, β-actin and lamin
B1 were used as the loading control. Grayscale scanning for western
blot analyses of three independent experiments was performed for
quantitative analysis. The protein bands were analyzed using
Gel-Pro Analyzer 6.3.0 software (Media Cybernetics, Rockville, MD,
USA).
TOP-FLASH luciferase assay
Cells were transfected with 250 ng of the TOP-FLASH
reporter constructs together with 25 ng of the Renilla
luciferase vector. Luciferase activity was measured by the Dual
Luciferase Reporter Assay System (Promega, Madison, WI, USA) 48 h
after transfection. Renilla luciferase was used as the
internal control, and TOP values were normalized to Renilla
values. The TOP values were calculated and used as indicators of
the endogenous level of Wnt signaling.
Statistical analysis
All of the statistical analyses were performed with
SPSS 18.0 software (SPSS Inc., Chicago, IL, USA).
Differences/correlations between two groups were assessed by the
Student's t-test. P-values <0.05 were considered to indicate a
statistically significant difference.
Results
CCAT2 is upregulated in ovarian cancer
cell lines
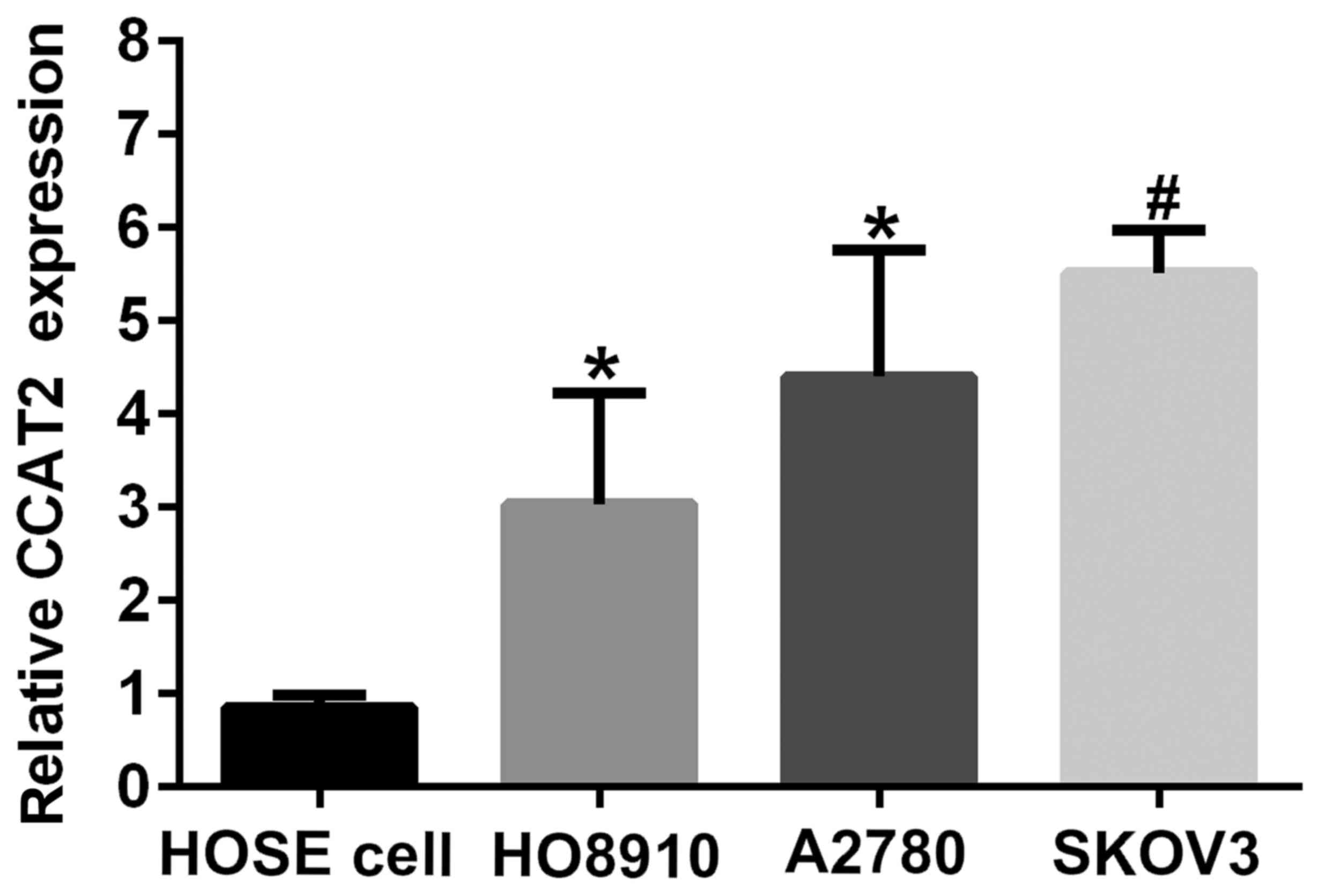
To know about the profile of CCAT2 in EOC cell lines
and HOSE cell, we characterized its expression by RT-qPCR in SKOV3,
A2780, HO8910 and HUM-CELL-0088 (Fig.
1). CCAT2 expression was upregulated in ovarian cancer cell
lines (SKOV3, A2780 and HO8910) compared with that in HOSE cell
(HUM-CELL-0088). The results also showed that the expression level
of CCAT2 in SKOV3 was the highest among three ovarian cancer cells,
followed by that in A2780. SKOV3 and A2780 cell lines were chosen
for subsequent experiments.
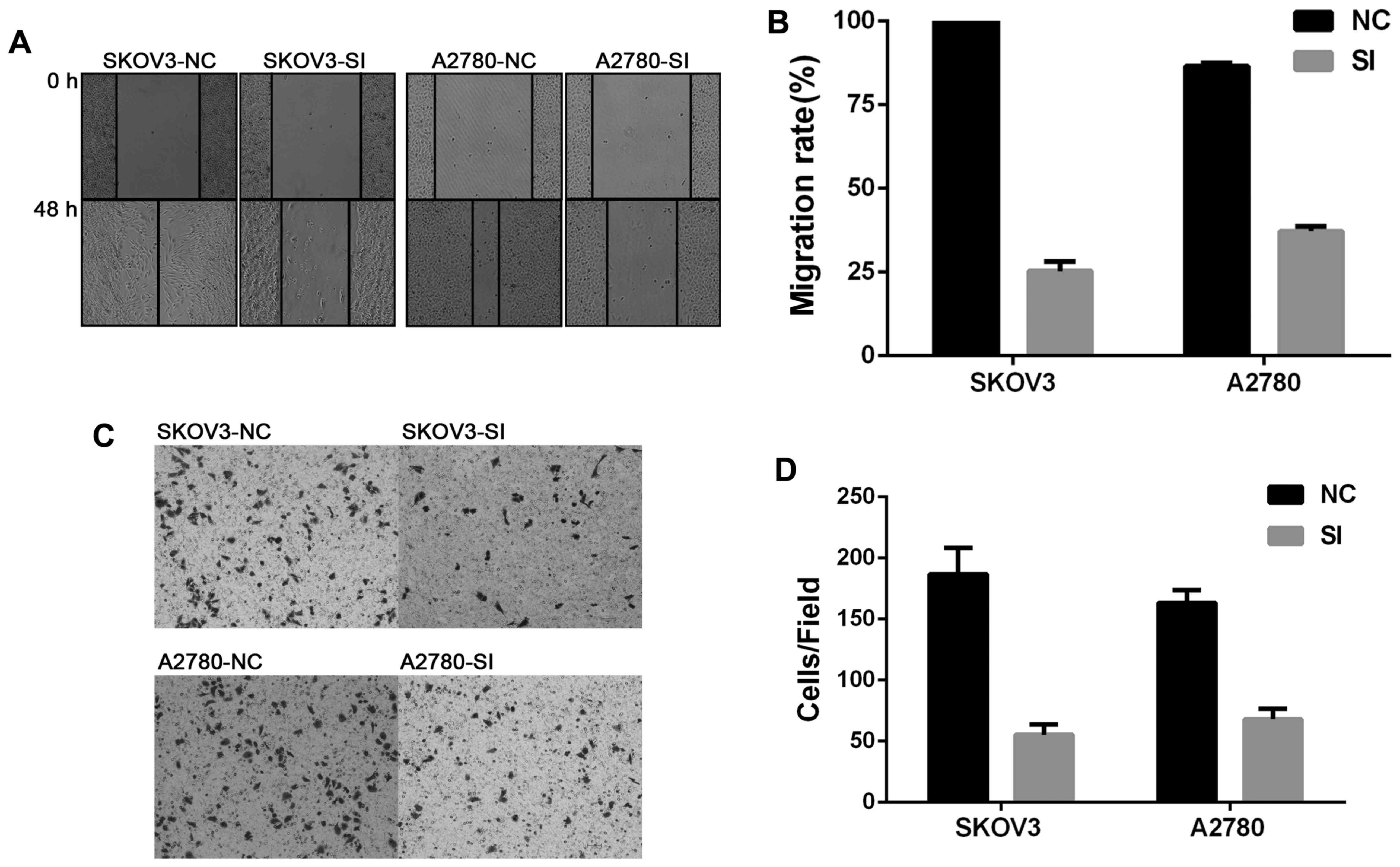
Silencing CCAT2 inhibited migration
and invasion of EOC cell lines in vitro
To investigate whether CCAT2 plays a key role in
facilitating cell migration and invasion, wound healing and
invasion assays were performed. As was shown in Fig. 2, knockdown of CCAT2 significantly
inhibited migration and invasion of EOC cells.
Knockdown of CCAT2 inhibits EMT in EOC
cells
To test the hypothesis that CCAT2 may influence EMT
in ovarian cancer cells, we examined a set of EMT marker genes,
including CDH1 (E-cadherin), CDH2 (N-cadherin) SNAI1 (Snail), SNAI2
(Slug) and Twist1. The data showed that knockdown of CCAT2
increased the expression of epithelial marker CDH1 but decreased
that of mesenchymal marker CDH2 at the mRNA and protein levels
(Figs. 3 and 4). Moreover, Slug and Twist1 were also
downregulated when CCAT2 was silenced in EOC cells (Figs. 3 and 4).
These results indicate that CCAT2 may be through the upregulation
of Slug and Twist1 to promote EMT in ovarian cancer cells.
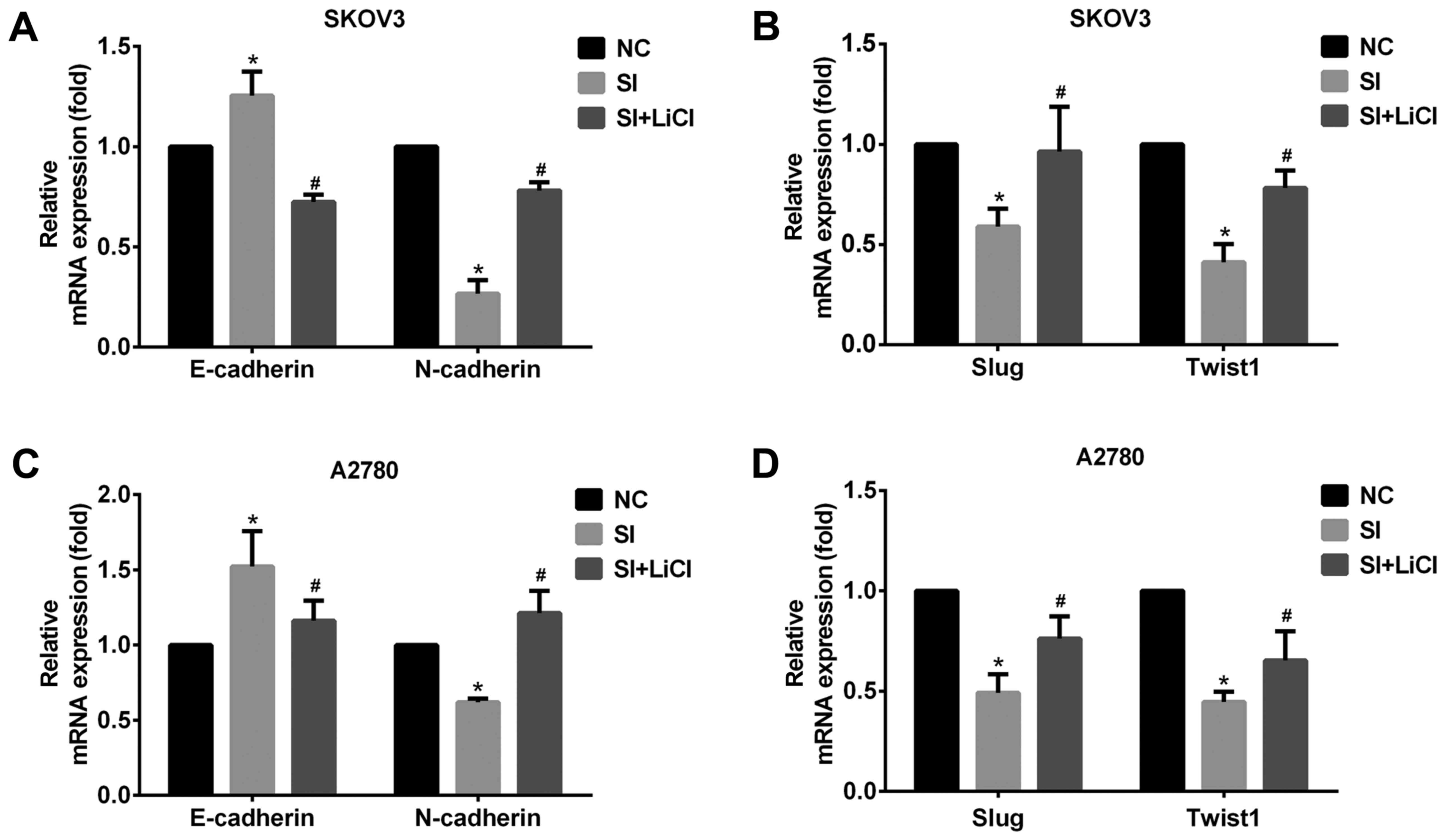 | Figure 3.CCAT2 promotes EMT involving
Wnt/β-catenin signaling pathway in EOC cells. (A) Knockdown of
CCAT2 increased the expression of E-cadherin and decreased that of
N-cadherin, (B) Slug and Twist1 at the mRNA level in SKOV3 cells
and (C) increased the expression of E-cadherin and decreased that
of N-cadherin, (D) Slug and Twist1 at the mRNA level in A2780. But,
that was reversed by the treatment with Wnt signaling activator
LiCl (20 mM for 24 h). The data represent the mean ± standard
deviation of three independent experiments. The values of the
controls were set as 1. *P<0.05 compared to NC group;
#P<0.05 compared to SI group (Student's t-test).
CCAT2, colon cancer-associated transcript 2; EMT,
epithelial-mesenchymal transition; EOC, epithelial ovarian
carcinoma; NC, negative control; siRNA, small interfering RNA;
LiCl, lithium chloride; E-cadherin, epithelial cadherin;
N-cadherin, neural cadherin; Slug, zinc finger protein SNAI;
Twist1, Twist-related protein 1. |
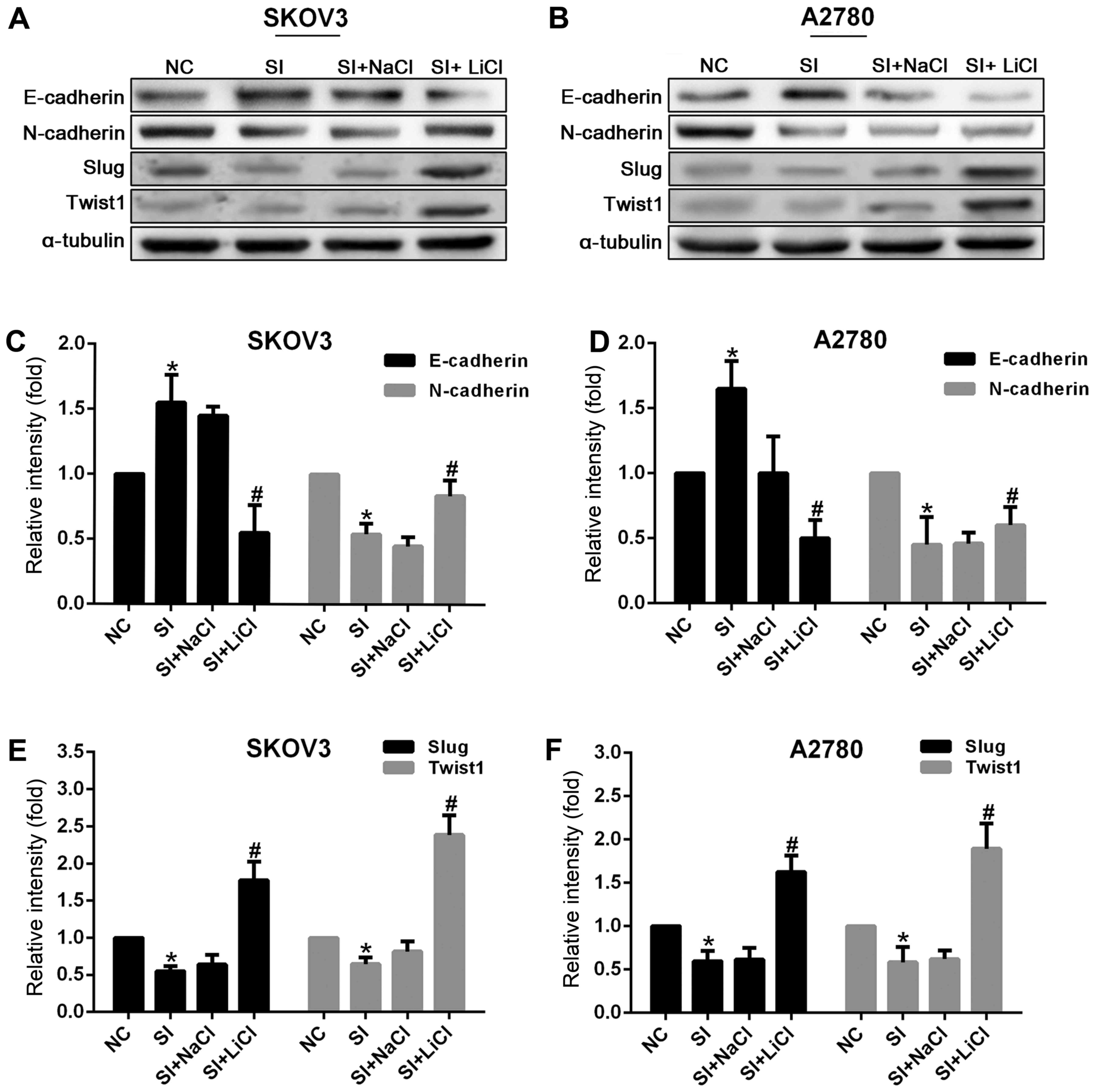 | Figure 4.Knockdown of CCAT2 increased the
expression of E-cadherin and decreased that of N-cadherin, Slug and
Twist1 at the protein level in SKOV3 and A2780 cells. But, that was
reversed by the treatment with Wnt signaling activator LiCl (20 mM)
for 24 h (20 mM NaCl as control). Representative images of western
blot analysis for (A) SKOV3 and (B) A2780 are presented. The data
represent the mean ± standard deviation of three independent
experiments. The values of the controls were set as 1. *P<0.05
compared to NC group; #P<0.05 compared to the SI+NaCl
group (Student's t-test). CCAT2, colon cancer-associated transcript
2; LiCl, lithium chloride; NC, negative control; siRNA, small
interfering RNA; E-cadherin, epithelial cadherin; N-cadherin,
neural cadherin; Slug, zinc finger protein SNAI; Twist1,
Twist-related protein 1. |
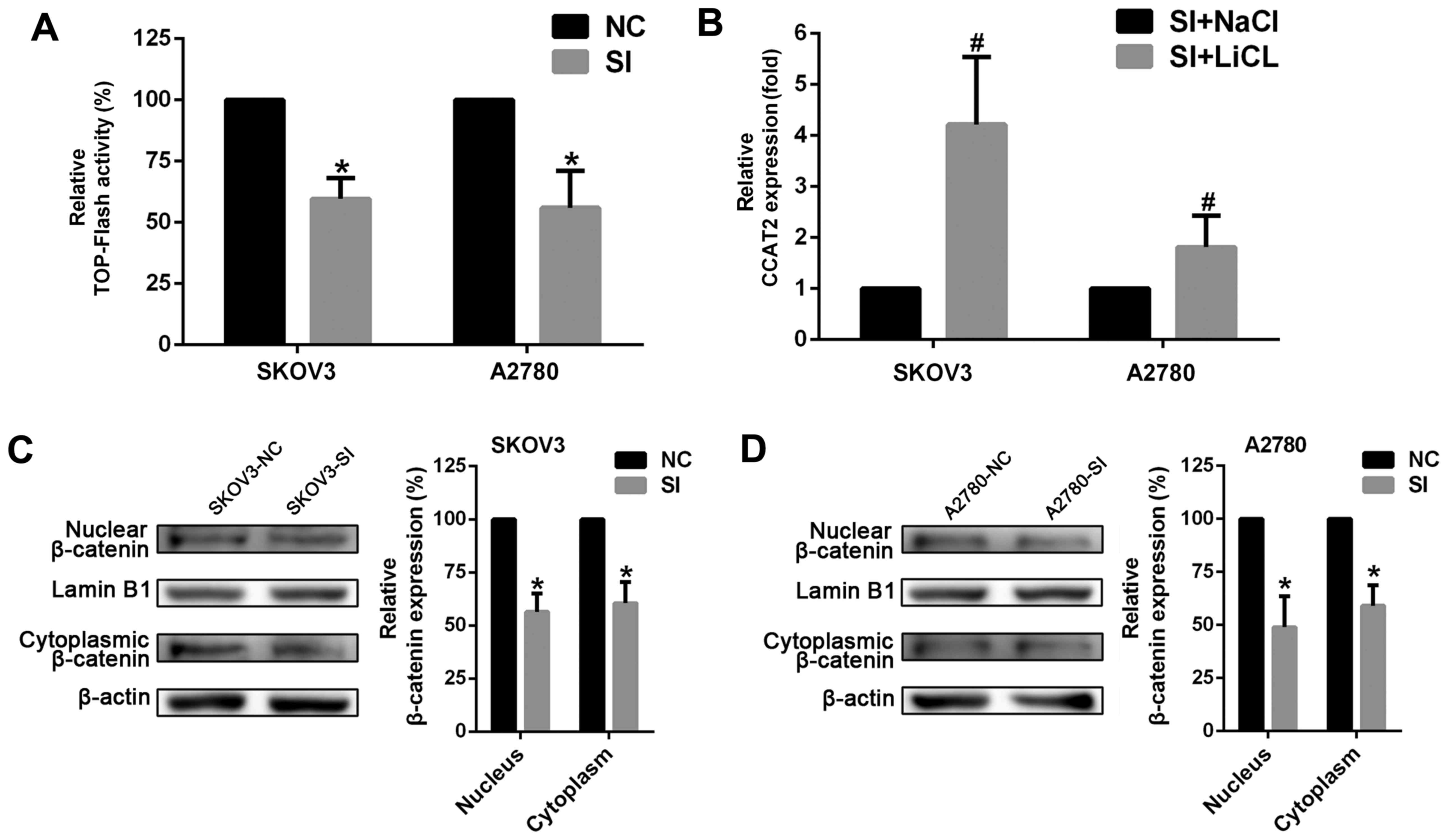
CCAT2 is involved in Wnt/β-catenin
signaling pathway
To better understand the detailed mechanisms of
CCAT2 in ovarian cancer, we tested whether suppressing CCAT2
affected Wnt signaling pathway whose aberration played a key role
in EOC development. The luciferase reporter system was employed in
SKOV3 and A2780 cells. The results showed that silencing CCAT2
decreased TCF/LEF activity in the cells (Fig. 5A). We next measured the expression
levels of β-catenin that is a key component of Wnt/β-catenin
signaling pathway. The data revealed that knockdown of CCAT2
decreased the expression of β-catenin both in the nucleus and
cytoplasm at the protein level (Fig. 5C
and D). In addition, we observed an obvious increase in CCAT2
expression after ovarian cancer cells were treated with lithium
chloride (LiCl; 20 mM for 24 h) in order to activate canonical
Wnt/β-catenin signaling through the inhibition of glycogen
synthase kinase 3β (GSK3B) (Fig. 5B).
These results strongly suggest that CCAT2 is involved in
Wnt/β-catenin signaling pathway and may elevate the signaling
activity in EOC cells.
CCAT2 promotes EMT involving
Wnt/β-catenin signaling pathway
The above results showed that knockdown of CCAT2
inhibited the process of EMT (Figs. 3
and 4) and the activity of Wnt
signaling (Fig. 5A and C). To further
study the underlying mechanism by which CCAT2 induced EMT, ovarian
cancer cells with knockdown of CCAT2 were treated with LiCl (20 mM)
for 24 h to activate canonical Wnt/β-catenin signaling. The results
showed that it downregulated the expression of CDH1 and upregulated
CDH2, Slug, Twist1 and CCAT2 (Figs.
3, 4 and 5B). Taken together, these data indicate that
CCAT2 may promote EMT involving Wnt/β-catenin signaling pathway in
EOC cells.
Discussion
Recently, lncRNAs were discovered to be dysregulated
in a variety of diseases, especially carcinomas (13–15). As a
novel lncRNA, CCAT2 was initially reported for its overexpression
in primary colorectal cancer and promotion of tumor growth,
metastasis and chromosomal instability (10). Redis et al revealed that CCAT2
modulated colorectal cancer metabolism by interaction with the
cleavage factor I (CFIm) complex in an allele-specific manner
(16). Their study indicated that the
mechanisms, underlying the regulation of metabolism, involved
lncRNA, protein complexes, oncogenes and transcription factors.
These findings provide some new and valuable clues to further
explore the biological functions of CCAT2 in EOC, since the
aberration of metabolism plays a really important role in ovarian
cancer progression (17,18). Wang et al reported that CCAT2
functioned as an oncogene in gastric cancer (GC) and was involved
in GC progression (19). Huang et
al found that CCAT2 expression was elevated in ovarian cancer
tissues and cell lines. They also demonstrated that high CCAT2
level was significantly positively correlated with FIGO stage,
tumor grade and distant metastasis of ovarian cancer (20). However, the molecular mechanisms by
which CCAT2 promotes EOC metastasis and progression are not fully
understood. In the current study, we investigated the effect of
CCAT2 on EMT and related molecular mechanisms in epithelial ovarian
cancer cells. Our results demonstrated that CCAT2 played a
functional role in promoting EMT of EOC cells.
As a significant biological process, EMT involves a
change from an epithelial to a mesenchymal phenotype in epithelial
cell. Downregulation of E-cadherin and upregulation of N-cadherin
are the hallmarks of EMT (21).
N-cadherin is a mesenchymal feature, while E-cadherin is an
epithelial adhesion glycoprotein whose decrease could make the
junctions between epithelial cells dissolved and facilitate
migratory and aggressive behavior (22). Previous studies demonstrated that the
expression of E-cadherin was reduced in various kinds of invasive
tumors including epithelial ovarian carcinoma (23–25). In
the present study, we observed that silencing CCAT2 inhibited cell
migration and invasion, as well as EMT of EOC cells. Furthermore,
we found that Slug and Twist1 were both downregulated at the mRNA
and protein levels after CCAT2 was knocked down. As is known, Slug
and Twist1 could facilitate the process of EMT by modulating
E-cadherin and N-cadherin. Therefore, these findings strongly
suggest that Slug and Twist1 may be the major targets by which
CCAT2 modulates EMT of ovarian cancer cells.
Canonical Wnt/β-catenin signaling pathway is
important for the induction of EMT (26,27).
Studies demonstrated that the aberrant signaling of Wnt pathway
played a key role in EMT during colorectal cancer progression
(28). Previous research has shown
that the suppression of Wnt signaling inhibited cell migration and
promoted cell adhesion in ovarian cancer cells, which was
attributed to the inhibition of EMT (29). By virtue of immunoprecipitation assay
targeting TCF7L2 (protein) and CCAT2 (RNA), Ling et al found
that CCAT2 might markedly modulate the downstream genes of Wnt
signaling pathway by combining with TCF7L2 and increasing its
transcriptional activity in colon cancer cells (10). In this study, we detected that
knockdown of CCAT2 inhibited the activity of TCF/LEF in SKOV3 and
A2780. Moreover, our investigation revealed that silencing CCAT2
decreased the expression of β-catenin both in the nucleus and
cytoplasm at the protein level. These results demonstrated that
knockdown of CCAT2 inhibited the activity of Wnt signaling in EOC
cells, which was in line with the finding in glioma and breast
cancer (30,31).
The above results revealed that knockdown of CCAT2
inhibited the process of EMT and the activity of Wnt signaling. To
further study the underlying mechanisms by which CCAT2 induced EMT,
cells with knockdown of CCAT2 were treated with LiCl to activate
canonical Wnt/β-catenin signaling. We found that LiCl treatment not
only reversed the effect of silencing CCAT2 on EMT, but also
greatly enhanced CCAT2 expression. These results suggest that CCAT2
may promote the process of EMT, at least partly, through
Wnt/β-catenin signaling pathway in epithelial ovarian carcinoma
cells. The data also indicate there might be a complex feedback
loop between CCAT2 and Wnt signaling that needs to be further
explored in the future.
In conclusion, the present study advances the
understanding of the molecular mechanisms by which CCAT2
facilitates EMT. Our findings suggest that CCAT2 may play an
important role in EOC progression. Thus, CCAT2 may serve as a
potential target for the treatment of ovarian cancer.
Acknowledgements
The authors thank Yufei Jiao (Department of
Pathology, the Second Affiliated Hospital of Harbin Medical
University), Xianli Zhou (Department of Ultrasound Medicine, the
Second Affiliated Hospital of Harbin Medical University) for
providing general support for this study. This study was supported
by Harbin Medical University.
References
|
1
|
Menon U, Gentry-Maharaj A and Jacobs I:
Ovarian cancer screening and mortality. JAMA. 306:1544–1545. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jelovac D and Armstrong DK: Recent
progress in the diagnosis and treatment of ovarian cancer. CA
Cancer J Clin. 61:183–203. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bast RC Jr, Hennessy B and Mills GB: The
biology of ovarian cancer: New opportunities for translation. Nat
Rev Cancer. 9:415–428. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fan WH, Du FJ, Liu XJ and Chen N:
Knockdown of FRAT1 inhibits hypoxia-induced
epithelial-to-mesenchymal transition via suppression of the
Wnt/β-catenin pathway in hepatocellular carcinoma cells. Oncol Rep.
36:2999–3004. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mo D, Yang D, Xiao X, Sun R, Huang L and
Xu J: miRNA-145 suppresses lung adenocarcinoma cell invasion and
migration by targeting N-cadherin. Biotechnol Lett. 1–710.
2017.PubMed/NCBI
|
|
6
|
Lim J and Thiery JP:
Epithelial-mesenchymal transitions: Insights from development.
Development. 139:3471–3486. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ramis-Conde I, Chaplain MA, Anderson AR
and Drasdo D: Multi-scale modelling of cancer cell intravasation:
The role of cadherins in metastasis. Phys Biol. 6:0160082009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
van Roy F: Beyond E-cadherin: Roles of
other cadherin superfamily members in cancer. Nat Rev Cancer.
14:121–134. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Faleiro-Rodrigues C, Macedo-Pinto I,
Pereira D and Lopes CS: Prognostic value of E-cadherin
immunoexpression in patients with primary ovarian carcinomas. Ann
Oncol. 15:1535–1542. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ling H, Spizzo R, Atlasi Y, Nicoloso M,
Shimizu M, Redis RS, Nishida N, Gafà R, Song J, Guo Z, et al:
CCAT2, a novel noncoding RNA mapping to 8q24, underlies metastatic
progression and chromosomal instability in colon cancer. Genome
Res. 23:1446–1461. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qiu M, Xu Y, Yang X, Wang J, Hu J, Xu L
and Yin R: CCAT2 is a lung adenocarcinoma -specific long non-coding
RNA and promotes invasion of non-small cell lung cancer. Tumour
Biol. 35:5375–5380. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Redis RS, Sieuwerts AM, Look MP, Tudoran
O, Ivan C, Spizzo R, Zhang X, de Weerd V, Shimizu M, Ling H, et al:
CCAT2, a novel long non-coding RNA in breast cancer: Expression
study and clinical correlations. Oncotarget. 4:1748–1762. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Crea F, Clermont PL, Parolia A, Wang Y and
Helgason CD: The non-coding transcriptome as a dynamic regulator of
cancer metastasis. Cancer Metastasis Rev. 33:1–16. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Richards EJ, Permuth-Wey J, Li Y, Chen YA,
Coppola D, Reid BM, Lin HY, Teer JK, Berchuck A, Birrer MJ, et al:
A functional variant in HOXA11-AS, a novel long non-coding RNA,
inhibits the oncogenic phenotype of epithelial ovarian cancer.
Oncotarget. 6:34745–34757. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Qiu JJ, Lin YY, Ye LC, Ding JX, Feng WW,
Jin HY, Zhang Y, Li Q and Hua KQ: Overexpression of long non-coding
RNA HOTAIR predicts poor patient prognosis and promotes tumor
metastasis in epithelial ovarian cancer. Gynecol Oncol.
134:121–128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Redis RS, Vela LE, Lu W, Ferreira de
Oliveira J, Ivan C, Rodriguez-Aguayo C, Adamoski D, Pasculli B,
Taguchi A, Chen Y, et al: Allele-specific reprogramming of cancer
metabolism by the long non-coding RNA CCAT2. Mol Cell. 61:520–534.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nieman KM, Kenny HA, Penicka CV, Ladanyi
A, Buell-Gutbrod R, Zillhardt MR, Romero IL, Carey MS, Mills GB,
Hotamisligil GS, et al: Adipocytes promote ovarian cancer
metastasis and provide energy for rapid tumor growth. Nat Med.
17:1498–1503. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Anderson AS, Roberts PC, Frisard MI,
Hulver MW and Schmelz EM: Ovarian tumor-initiating cells display a
flexible metabolism. Exp Cell Res. 328:44–57. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang YJ, Liu JZ, Lv P, Dang Y, Gao JY and
Wang Y: Long non-coding RNA CCAT2 promotes gastric cancer
proliferation and invasion by regulating the E-cadherin and LATS2.
Am J Cancer Res. 6:2651–2660. 2016.PubMed/NCBI
|
|
20
|
Huang S, Qing C, Huang Z and Zhu Y: The
long non-coding RNA CCAT2 is up-regulated in ovarian cancer and
associated with poor prognosis. Diagn Pathol. 11:492016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Onder TT, Gupta PB, Mani SA, Yang J,
Lander ES and Weinberg RA: Loss of E-cadherin promotes metastasis
via multiple downstream transcriptional pathways. Cancer Res.
68:3645–3654. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sawada K, Mitra AK, Radjabi AR, Bhaskar V,
Kistner EO, Tretiakova M, Jagadeeswaran S, Montag A, Becker A,
Kenny HA, et al: Loss of E-cadherin promotes ovarian cancer
metastasis via alpha 5-integrin, which is a therapeutic target.
Cancer Res. 68:2329–2339. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Poser I, Domı́nguez D, Herreros AG, Varnai
A, Buettner R and Bosserhoff AK: Loss of E-cadherin expression in
melanoma cells involves up-regulation of the transcriptional
repressor Snail. J Biol Chem. 276:24661–24666. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Arend RC, Londono-Joshi AI, Straughn Jr JM
and Buchsbaum DJ: The Wnt/β-catenin pathway in ovarian cancer: A
review. Gynecol Oncol. 131:772–779. 2001. View Article : Google Scholar
|
|
27
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gonzalez DM and Medici D: Signaling
mechanisms of the epithelial-mesenchymal transition. Sci Signal.
7:re82014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ford CE, Jary E, Ma SS, Nixdorf S,
Heinzelmann-Schwarz VA and Ward RL: The Wnt gatekeeper SFRP4
modulates EMT, cell migration and downstream Wnt signaling in
serous ovarian cancer cells. PLoS One. 8:e543622013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hua G, Hu G, Yang Q, Zhang P, Kuang W, Zhu
X and Wu L: Knockdown of long non-coding RNA CCAT2 suppressed
proliferation and migration of glioma cells. Oncotarget.
7:81806–81814. 2016.PubMed/NCBI
|
|
31
|
Cai Y, He J and Zhang D: Long noncoding
RNA CCAT2 promotes breast tumor growth by regulating the Wnt
signaling pathway. Onco Targets Ther. 8:2657–2664. 2015.PubMed/NCBI
|