Introduction
Osteosarcoma (OS), rooted in primitive bone-forming
mesenchymal cells and occurring primarily in pediatric patients, is
the most common malignant skeletal tumor (1,2).
Generally, OS patients experience severe pain and soft-tissue
swelling due to tumor cell invasion (3). In addition, the destructive effects on
cancer-associated tissues and cells in the majority of cases
(80–90%) are localized to the long bones, with fewer cases
occurring in the axial and irregular bones (2). Although OS comprises ≤0.5% of all types
of cancer (4), the risk of
pathological fracture in pediatric patients is up to 15% (5). Furthermore, the 5-year survival rate of
OS patients was revealed to be <60% even under the circumstances
of great improvement in surgical techniques and chemotherapy and/or
radiotherapy regimens (6). Therefore,
on the basis of these findings, a great deal of basic studies have
contributed toward understanding the mechanisms underpinning OS
development. However, these mechanisms remain unclear and require
further investigation in basic or clinical studies in order to be
elucidated.
MicroRNAs (miRNAs/miRs) consist of 18–25 nucleotides
and are a class of short non-coding RNA molecules. miRNAs, as
regulators of gene expression, are able to induce the
destabilization of mRNA and/or the inhibition of protein
translation by directly binding to the 3′-untranslated region (UTR)
of their target genes (7,8). It has been reported that miRNAs are
involved in the modulation of a variety of biological processes,
including cell proliferation, differentiation, apoptosis and
migration, and tumorigenesis (8,9). It has
also been reported that miR-499a-5p may contribute to the
progression of oral squamous cell carcinoma (10). However, as of yet, there are no
studies combining clinical and basic experiments regarding the
function of miR-499a-5p in regulating OS tissue and cell growth,
proliferation and metastasis.
In the present study, OS tissues and the adjacent
normal tissues were collected from 62 patients, and the expression
of protein phosphatase 1D (PPM1D), also known as wild-type
p53-inducible phosphatase-1 (Wip1), which has been reported
to be involved in the regulation of cellular proliferation and
metastasis (11–14). The objective of the present study was
to clarify the association between PPM1D and
osteosarcoma.
Materials and methods
Patient tissue samples
A total of 62 primary OS and adjacent non-tumorous
tissue samples were collected during surgical resection in the
Department of Pathology of Xi'an Jiaotong University Affiliated
Hong Hui Hospital (Xi'an, Shaanxi, China) between May 2007 and May
2016. All tissues were rapidly frozen in liquid nitrogen and
maintained at −70°C. None of the patients had undergone blood
transfusion, chemotherapy or radiotherapy prior to surgery. All the
samples were used for experiments with the patients' written
informed consent and the approval of the Ethics Committee of Hong
Hui Hospital Affiliated to Xi'an Jiaotong University. In order to
clarify the association between OS and miR-499a-5p expression, the
62 OS patient tissues were divided into a metastasis group (n=28)
and a non-metastasis (n=34) group according to the
Tumor-Node-Metastasis (TNM) staging system (15), among which the ratio of male and
female was 37: 25 and the average age was 19.3±4.5 years.
miRNA microarray analysis
The total RNA of the OS tissues and the adjacent
non-tumorous tissues was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
according to the manufacturer's protocol. miRNA microarray analysis
(v.16.0; Exiqon A/S, Vedbaek, Denmark) was applied to detect the
differential expression of miRNAs in the OS tissues and the
adjacent non-tumorous tissues according to the manufacturer's
protocol.
MG-63 cell culture and
transfection
Human OS MG-63 cells, purchased from the Type
Culture Collection of the Chinese Academy of Sciences (Shanghai,
China), were cultured in minimum essential medium/Earle's balanced
salt solution supplemented with 10% fetal bovine serum, 50 U/ml
penicillin and 50 mg/ml streptomycin (all Hyclone; GE Healthcare
Life Sciences, Logan, UT, USA) at 37°C in a humidified 5%
CO2 atmosphere. Cells were plated onto 6-well plates
(5×105 cells/well) and the medium was replaced every 3
days. Once the cells reached 70% confluence, 100 nM miR-499a-5p
mimic (forward 5′-GACACGGCTCCGTCAC-3′; reverse
5′-ATCCCTGATCAACTGTCCGCC-3′) or its negative control (miR-499a-5p
mimic NC) (forward 5′-AACCAGGCCTCCACCG-3′; reverse
5′-GCCACGCATGTCTTATACTGC-3′) (Sangon Biotech Co., Ltd., Shanghai,
China) was transfected into the MG-63 cells using Lipofectamine
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. After 4–6 h co-transfection, the
transfection reagent was exchanged with fresh normal medium, and 24
h later, total RNA or total protein was extracted.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from the OS tissues and
normal cells transfected with either miR-499a-5p or its NC using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.), and
then l mg total RNA was reverse transcribed using a PrimeScript RT
Reagent Kit and a gDNA Eraser kit (Takara Biotechnology Co., Ltd.,
Dalian, China) according to the manufacturer's protocol. Sequences
of all primers were as follows: miR-499a-5p forward,
5′-ATGTAGCGTGCGACCG-3′ and reverse, 5′-CAGGCTGACGCACTCTGTGCT-3′; U6
forward, 5′-CCATCGGAAGCTCGTATACGAAATT-3′ and reverse,
5′-GGCCTCTCGAACTTGCGTGTCAG-3′; PPM1D forward,
5′-TTCAGAGCTCCATAACAAGC-3′ and reverse, 5′-GGACCTATCGCATCCGACCG-3′;
and β-actin forward, 5′-GCTGTCCCTGTATGCCTCT-3′ and reverse,
5′-TGTCACGCACGATTTCC-3′. Subsequently, 1 µl miR-499a-5p or
PPM1D primers (Sangon Biotech Co., Ltd.), 10 µl SYBRGreen
(Takara Biotechnology Co., Ltd.), 6 µl diethylpyrocarbonate
(DEPC)-treated water (Sangon Biotech Co., Ltd.) and 2 µl cDNA, with
a total volume of 20 µl, was then processed in a Real-Time PCR
system (Roche Diagnostics, Basel, Switzerland). The following
thermocycling conditions were used: 5 sec at 95°C, followed by 40
cycles of 1 sec at 95°C and 20 sec at 65°C. The comparative
2−ΔΔCq method was used for relative quantification of
gene expression (16). Each sample
was analyzed ≥3 times.
Luciferase activity assay
A fragment that included the 3′-UTR of human
PPM1D (NCBI Reference Sequence: NG_023265.1) was amplified
from human genomic DNA using the following primer pair:
5′-TCTTGAACTCTGAGCTCAAGCGATCCACCCACCTCGGCCTCC-3′ and
5′-GTTAAGAAAGTCCCTTAATATTTCAACTGAGATCTGAGTAGC-3′. pGL3-PPM1D-wild
reporter was constructed by interception of PCR products of the
3′-UTR of PPM1D, which was cut with XbaI and cloned
into the corresponding sites of the pGL3 vector (Promega
Corporation, Madison, WI, USA). The QuikChange II Site-Directed
Mutagenesis kit (Takara Biotechnology Co., Ltd.) was used to take
the mutation into the putative sites of miR-499a-5p recognition,
according to the manufacturer's protocol. A total of 400 ng
Luciferase reporter, 100 nM miRNA-499a-5p mimic or its NC and 30 ng
pRL-TK Renilla Luciferase Reporter Vector (Promega Corporation)
were added to each 24-well plate (4–5×104 cells/well).
Following transfection with Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) for 48 h, cells were collected and
luciferase activity was measured using a
Dual-Luciferase® Reporter assay system to compare with
renilla luciferase activity (Promega Corporation).
Bioinformatic resources
The online Targetscan software (TargetScanHuman 7.1;
http://www.targetscan.org/) was used to
predict the target genes of miR-499a-5p with reference to the human
gene sequence. The predicted genes identified by the software
program were intersected for further analysis.
Western blot analysis
Total protein was extracted from collected tissues
and cells using RIPA Lysis Buffer (Beyotime Institute of
Biotechnology, Haimen, China) supplemented with 100X proteinase
inhibitor and phosphatase inhibitor. Protein quantity was
determined using a bicinchoninic acid assay kit (Beyotime Institute
of Biotechnology) according to the manufacture's protocol. Proteins
(25 µg) were separated with 10% SDS-PAGE. Subsequently, the gel was
transferred onto a polyvinylidene difluoride membrane (Merck KGaA,
Darmstadt, Germany) and blocked with 5% skimmed milk for 1 h at
room temperature. The following rabbit monoclonal primary
antibodies were used: Anti-PPM1D (cat. no. 11901; 1:1,000
dilution), anti-p-AKT (ser473; cat. no. 12694; 1:1,000 dilution),
anti-p-GSK-3β (ser9; cat. no. 14026; 1:500 dilution), anti-Akt
(cat. no. 2938; 1:1,000 dilution), anti-GSK-3β (cat. no. 9315;
1:500 dilution) and anti-β-actin (cat. no. 4970; 1:1,000 dilution)
(all Cell Signaling Technology, Inc., Danvers, MA, USA). A
secondary mouse anti-rabbit antibody was then used (1:5,000
dilution; A0208; Beyotime, Shanghai, China). The signal was
visualized using the enhanced chemiluminescence kit (Invitrogen;
Thermo Fisher Scientific, Inc.). The gray bands were analyzed with
Image J software (version 1.46; National Institutes of Health,
Bethesda, MD, USA) to compare the expression between targeted
proteins and internal control of β-actin.
MG-63 cell proliferation assay
To detect the proliferation activity of MG-63 cells
transfected with the miR-499a-5p mimic or its NC, the MTT assay was
performed using Cell Proliferation kit I (Sigma-Aldrich; Merck
KGaA) on 96-well plates at a density of 4×103
cells/well, according to the manufacturer's protocol. After 24, 48
and 72 h of incubation, MTT-formazan (Dissolved in chloroform: 10
mg/ml clear, dark violet) production was estimated by VersaMax
(Molecular Devices, LLC, Sunnyvale, CA, USA) at 570 nm to evaluate
the proliferation rate. The index was determined at 48 and 72 h
normalized to that at 24 h. Subsequently, the images of the
transfected cells were captured using a microscope (model no. DP72;
Olympus Corporation, Tokyo, Japan) at ×100 magnification.
Statistical analysis
Data are presented as the mean ± standard error of
the mean. All statistical analyses were performed using SPSS 17.0
software (SPSS, Inc., Chicago, IL, USA). The differences between
two groups were compared using a paired Student's t-test and the
differences between three or more groups using one-way analysis of
variance followed by Tukey's post-hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Low expression of miR-499a-5p and high
expression of PPM1D mRNA are associated with OS
Between May 2007 and May 2016, 62 OS tissues and the
adjacent non-tumorous specimens were collected. Using miRNA
microarray analysis, it was determined that 29 miRNAs were
significantly upregulated, while 26 miRNAs were downregulated in
the OS tissues. Among the downregulated miRNAs, miR-499a-5p
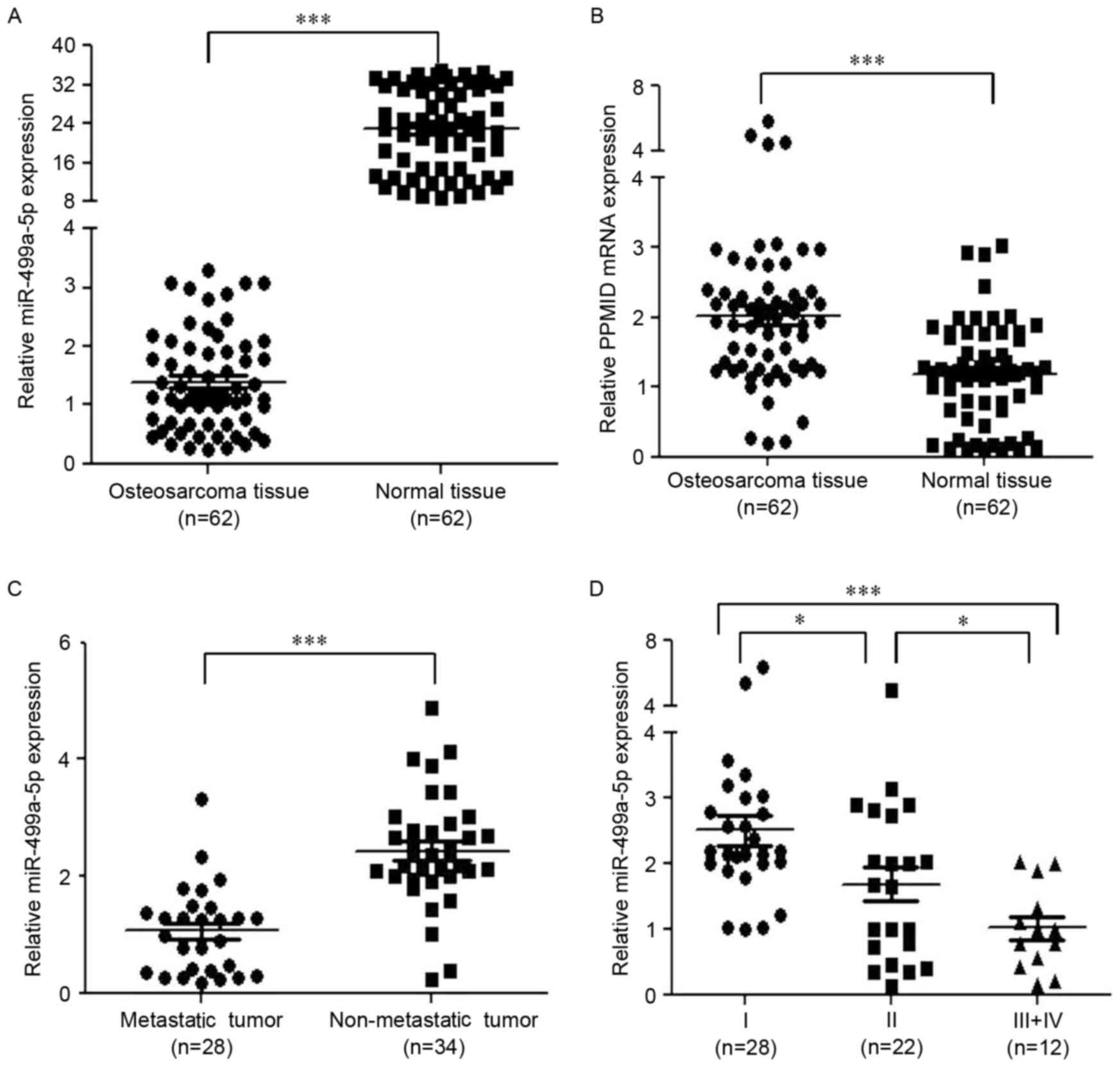
expression exhibited the largest decrease (Table I). Through RT-qPCR analyses, the
expression of miR-499a-5p was observed to be lower (Fig. 1A), while that of PPM1D was
higher (Fig. 1B) in cancerous tissues
compared with that in the healthy samples. Furthermore, there were
28 metastatic and 34 non-metastatic tumor patients among all the OS
patients, and the expression of miR-499a-5p was significantly
downregulated in the metastatic samples compared with that in the
non-metastatic samples (Fig. 1C).
Additionally, with regard to TNM staging (15), tumors in stages III+IV exhibited the
lowest expression of miR-499a-5p, while those in stage I, exhibited
the highest expression of miR-499a-5p (Fig. 1D).
 | Table I.miRNA microarray analyses of
differentially expressed miRNAs in OS tissues and adjacent normal
tissues (OS tissues/normal tissues). |
Table I.
miRNA microarray analyses of
differentially expressed miRNAs in OS tissues and adjacent normal
tissues (OS tissues/normal tissues).
| miRNAs | Fold-change | P-value |
|---|
| Upregulated |
|
|
|
hsa-miR-351-3p | 20.18 | 0.0107 |
|
hsa-miR-202-3p | 19.04 | 0.0178 |
|
hsa-miR-273-3p | 17.95 | 0.0117 |
|
hsa-miR-121-5p | 16.77 | 0.0275 |
|
hsa-miR-208-3p | 16.39 | 0.0208 |
|
hsa-miR-248 | 15.28 | 0.0099 |
|
hsa-miR-128-5p | 14.73 | 0.0047 |
|
hsa-miR-19-3p | 14.07 | 0.0137 |
|
hsa-miR-213 | 13.78 | 0.0472 |
|
hsa-miR-192 | 13.07 | 0.0429 |
|
hsa-miR-191 | 12.18 | 0.0378 |
|
hsa-miR-576-3p | 11.45 | 0.0118 |
|
hsa-miR-472-3p | 10.11 | 0.0251 |
|
hsa-miR-502a-3p | 9.19 | 0.0187 |
|
hsa-miR-521-3p | 8.37 | 0.0049 |
|
hsa-miR-541e | 8.01 | 0.0391 |
|
hsa-miR-131-5p | 6.89 | 0.0172 |
|
hsa-miR-9a-5p | 6.12 | 0.0394 |
|
hsa-miR-493-3p | 5.99 | 0.0187 |
|
hsa-miR-702-5p | 5.08 | 0.0292 |
|
hsa-miR-198 | 4.77 | 0.0031 |
|
hsa-miR-130 | 4.54 | 0.0093 |
|
hsa-miR-122-5p | 3.93 | 0.0172 |
|
hsa-miR-122 | 3.01 | 0.0364 |
|
hsa-miR-174 | 2.91 | 0.0208 |
|
hsa-miR-215 | 2.66 | 0.0407 |
|
hsa-miR-274 | 2.27 | 0.0099 |
|
hsa-miR-176 | 2.11 | 0.0152 |
|
hsa-miR-18 | 1.93 | 0.0381 |
| Downregulated |
|
|
|
hsa-miR-197-3p | 0.99 | 0.0113 |
|
hsa-miR-415-5p | 0.97 | 0.0309 |
|
hsa-miR-1406 | 0.87 | 0.0293 |
|
hsa-miR-13a-5p | 0.82 | 0.0183 |
|
hsa-miR-311a | 0.78 | 0.0211 |
|
hsa-miR-307b | 0.72 | 0.0191 |
|
hsa-miR-51-5p | 0.69 | 0.0313 |
|
hsa-miR-25b-5p | 0.62 | 0.0091 |
|
hsa-miR-25a-5p | 0.59 | 0.0049 |
|
hsa-miR-99a-3p | 0.53 | 0.0412 |
|
hsa-miR-110 | 0.51 | 0.0279 |
|
hsa-miR-238-3p | 0.45 | 0.0019 |
|
hsa-miR-47-5p | 0.44 | 0.0373 |
|
hsa-miR-29-3p | 0.42 | 0.0147 |
|
hsa-miR-37-5p | 0.39 | 0.0045 |
|
hsa-miR-215 | 0.37 | 0.0078 |
|
hsa-miR-27-3p | 0.35 | 0.0128 |
|
hsa-miR-90 | 0.29 | 0.0327 |
|
hsa-miR-77-5p | 0.27 | 0.0401 |
|
hsa-miR-398 | 0.16 | 0.0019 |
|
hsa-miR-187 | 0.11 | 0.0108 |
|
hsa-miR-150 | 0.09 | 0.0071 |
|
hsa-miR-381-3p | 0.07 | 0.0131 |
|
hsa-miR-294 | 0.06 | 0.0307 |
|
hsa-miR-201-5p | 0.05 | 0.0287 |
|
hsa-miR-499a-5p | 0.04 | 0.0175 |
PPM1D is targeted by miR-499a-5p in
MG-63 cells
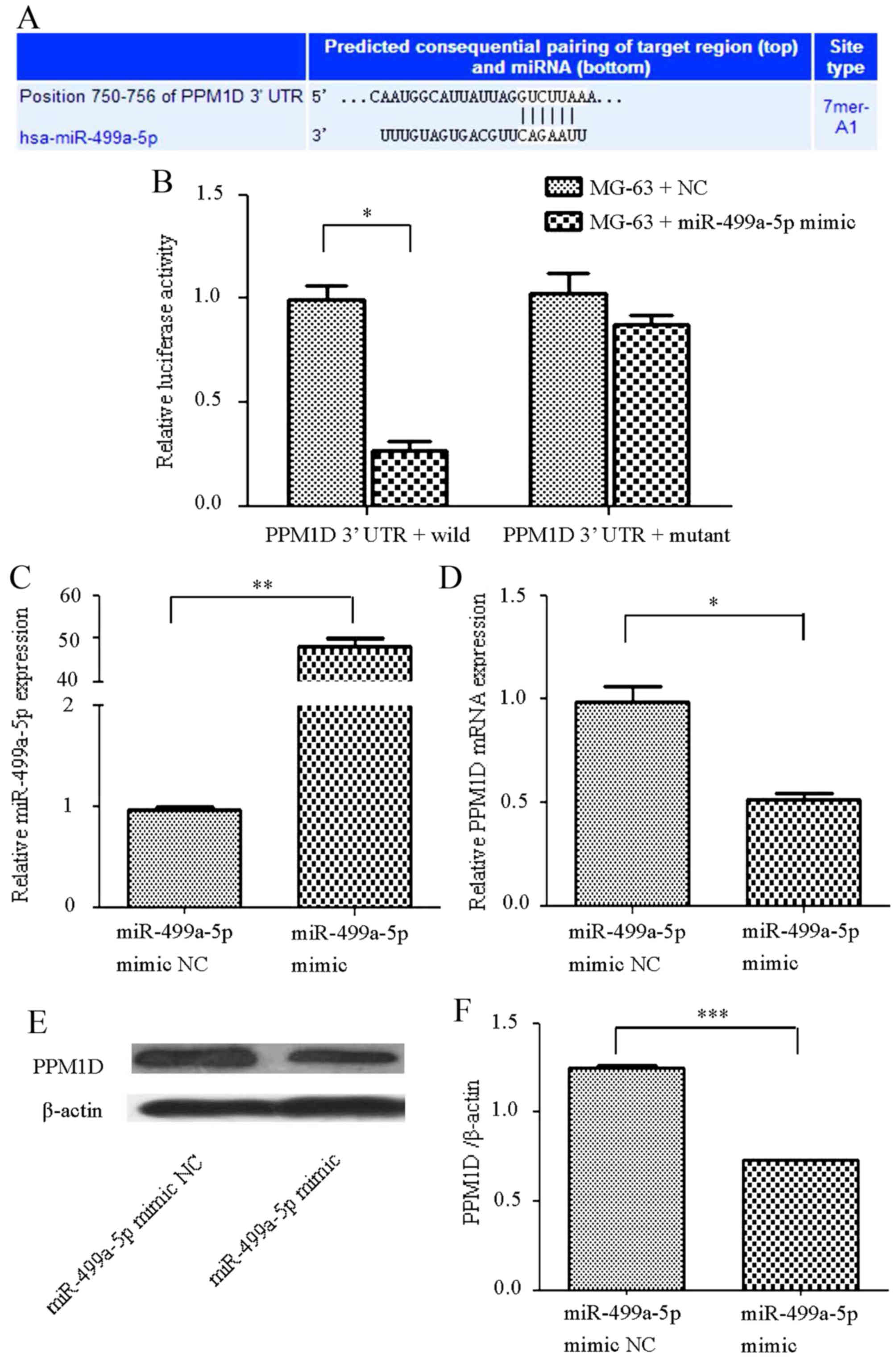
In order to confirm that PPM1D is targeted by
miR-499a-5p, the potential targets of miR-499a-5p were examined
using Targetscan software and the conserved predicted consequential
pairing of the PPM1D 3′-UTR of miR-499a-5p was observed
(Fig. 2A). In addition, human OS
MG-63 cells were transfected with an miR-499a-5p mimic or an NC,
and the luciferase activity was notably suppressed in the
PPM1D 3′-UTR wild-type group. However, no statistically
significant differences were observed in the mutant group (Fig. 2B). When cells were transfected with
the mimic, miR-499a-5p expression was elevated (Fig. 2C), while PPM1D mRNA and protein
expression was significantly decreased compared with that of the NC
(Fig. 2D-F).
miR-499a-5p inversely regulates MG-63
cell growth and proliferation by targeting PPM1D through Akt/GSK-3β
signaling
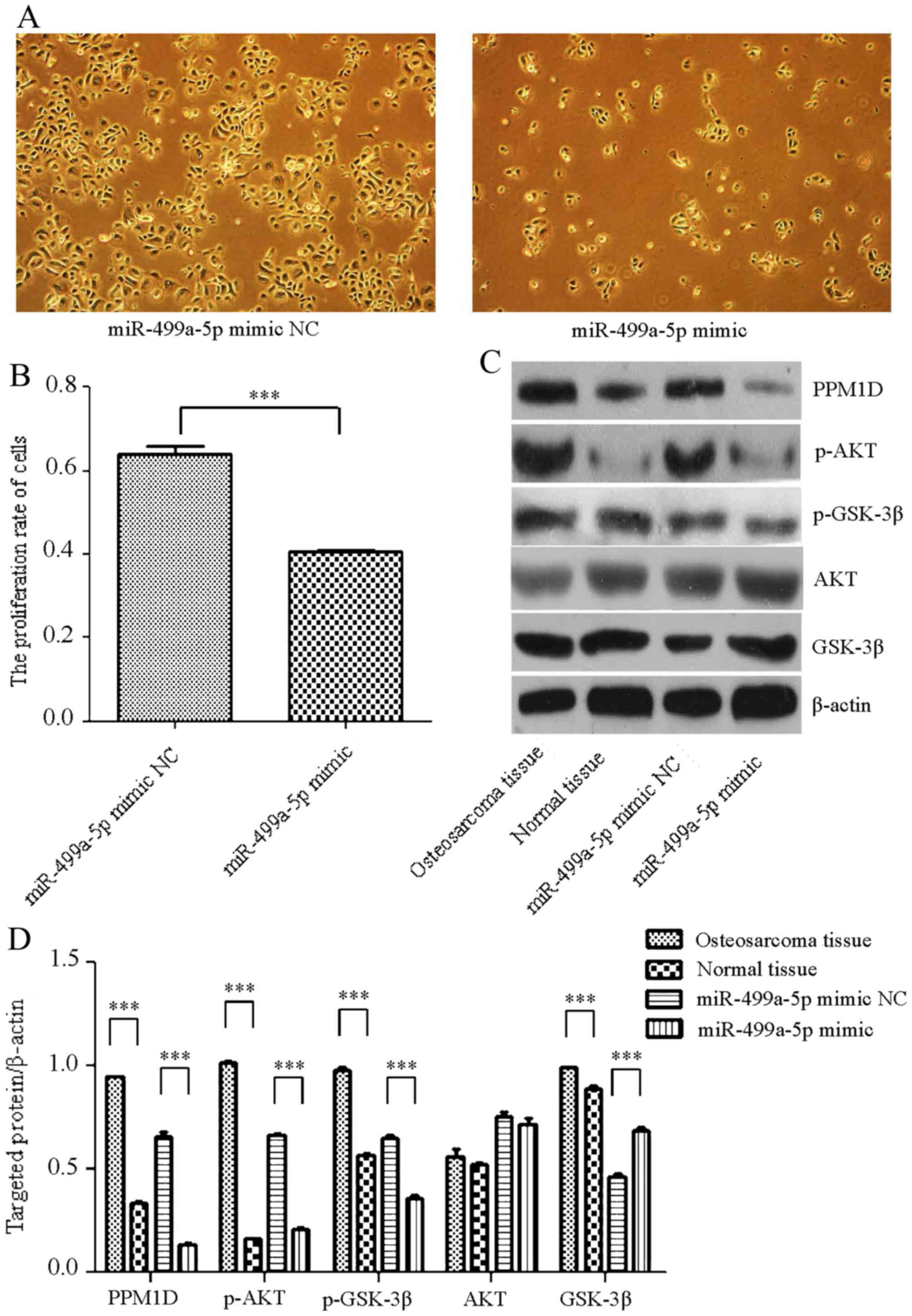
Following transfection of MG-63 cells with an
miR-499a-5p mimic, fewer cells were observed in the transfection
group compared with the number in the NC group (Fig. 3A). Using the Cell Proliferation kit,
it was observed that the proliferation rate of the cells
transfected with the miR-499a-5p mimic was significantly decreased
(Fig. 3B). In addition, total protein
was extracted from the tumorous tissues, the non-tumorous tissues
and the transfected cells. Western blot analysis indicated that the
expression of PPM1D, as well as that of the downstream proteins of
p-AKT and p-GSK-3β, was upregulated in OS tissues and downregulated
in cells transfected with the miR-499a-5p mimic compared with
expression in the normal tissues and the miR-499a-5p mimic NC
group, respectively (Fig. 3C). Image
J software analyses indicated that the expression of PPM1D, p-AKT
and p-GSK-3β was significantly altered (Fig. 3D).
Discussion
A large volume of evidence has demonstrated that
miRNAs may regulate human tumor development and may progress as
oncogenes or cancer suppressors through mediating the expression of
their targets (17,18). Therefore, in order to fully understand
the precise mechanisms associated with pathological and
physiological processes and associated diseases, miRNAs may provide
a novel solution for the diagnosis and therapy of various types of
disease, particularly cancer (19).
Numerous studies have revealed that miRNAs are involved in the
development and progression of OS. Certain aspects of these studies
have focused on the role of miRNAs in promoting the development of
OS. For example, increased expression of miRNA-148a in OS promotes
cancer cell growth by targeting phosphatase and tensin homolog
(20), miRNA-181a promotes
proliferation and inhibits apoptosis by suppressing component of
the cleavage factor Im 25 in OS (21), and miR-26a induces OS cell growth and
metastasis via the Wnt/β-catenin pathway (22). However, the majority of studies have
illustrated that high levels of miRNAs exert an inhibitory effect
on OS. For instance, miRNA-101 inhibited OS cell migration and
invasion through regulation of the enhancer of zeste homolog 2
(23), elevated expression of
miRNA-646 suppressed OS cell proliferation, migration and invasion
by inversely targeting fibroblast growth factor-2 (24), and miRNA-145 inhibited OS cell growth
by suppressing the expression of ρ-associated,
coiled-coil-containing protein kinase 1 (25). In addition, transfecting OS cells with
a plasmid with upregulated miRNA-410 suppressed cell proliferation
or promoted cell apoptosis by targeting vascular endothelial growth
factor (26). Furthermore, miR-497
has been revealed to be strongly associated with a human OS cell
line and its downregulation is believed to contribute toward tumor
cell growth and proliferation (19,27).
However, no studies have demonstrated an association between the
expression of miR-499a-5p and OS. In the present study, 62 patient
OS tissues and the adjacent non-tumorous tissues were collected
between May 2007 and May 2016. miRNA microarray analysis
demonstrated that 29 miRNAs were significantly upregulated, while
26 were downregulated in the OS tissues. Among the upregulated
miRNAs, miR-351-3p was elevated ~20-fold and was the most promoted.
In addition, among the downregulated miRNAs, miR-499a-5p was
decreased 25-fold and was the most declined. Therefore, miR-499a-5p
was selected for the subsequent experiments. It was identified that
in the metastasis group, miR-499a-5p expression was markedly lower
than that in the non-metastasis group. Furthermore, as the patients
were further divided into stage I, II or III+IV groups according to
the TNM stage classification system (15), miR-499a-5p expression was notably
declined in the stage III+IV tumors compared with that in stage I.
These data indicate that miR-499a-5p was inversely associated with
tumor tissue differentiation and metastasis.
PPM1D is overexpressed in a number of human
cancer types, including breast carcinomas, neuroblastomas,
pancreatic adenocarcinomas, gastric carcinomas and medulloblastomas
(28–32). However, in certain types of cancer, it
has been reported that PPM1D negatively regulates neutrophil
migration and inflammation (33) and
suppresses ovarian cancer metastasis (12). The function of PPM1D in cancer
metastasis appears to be type-specific. In the present study, not
only was an inverse association between miR-499a-5p expression and
OS observed, but protein and mRNA levels of PPM1D were
revealed to be upregulated in OS tissues. Additionally, in MG-43
cells transfected with miR-499a-5p, the mRNA and protein expression
of PPM1D was downregulated. The results of the Targetscan
predictive software and transfection of MG-63 cells with an
miR-499a-5p mimic demonstrated that PPM1D was targeted by
miR-499a-5p. In the human OS SAOS-2 cell line, the downregulation
of miR-497 contributes toward cell growth and cisplatin resistance
through the Akt pathway (18), and in
ovarian cancer lines, Wip1 suppresses ovarian cancer
metastasis through the ataxia telangiectasia-mutated
protein/Akt/Snail-mediated signaling pathway (12). In the present study, the proliferation
rate of MG-63 cells was significantly decreased following
transfection with an miR-499a-5p mimic which promoted the
expression of miR-499a-5p compared with its NC group. In the OS
tissues and the MG-63 cells, expression of PPM1D and
downstream proteins p-Akt and p-GSK-3β were all inversely regulated
by miR-499a-5p.
In conclusion, the present study provides novel and
significant basic evidence demonstrating that miR-499a-5p may
target-regulate OS tissue and cell proliferation and
differentiation via the PPM1D/Akt/GSK-3β signaling pathways.
However, the exact mechanism underpinning this may require further
investigation in the future.
References
|
1
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zheng H and Min J: Role of long noncoding
RNA HOTAIR in the growth and apoptosis of osteosarcoma cell MG-63.
Biomed Res Int. 2016:57576412016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Angiero F, Moltrasio F, Cattoretti G and
Valente MG: Clinical and histopathological profile of primary or
secondary osteosarcoma of the jaws. Anticancer Res. 31:4485–4489.
2011.PubMed/NCBI
|
|
4
|
Jemal A, Siegel R, Ward E, Murray T, Xu J
and Thun MJ: Cancer statistics, 2007. CA Cancer J Clin. 57:43–66.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Picci P: Osteosarcoma (osteogenic
sarcoma). Orphanet J Rare Dis. 2:62007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Desandes E: Survival from adolescent
cancer. Cancer Treat Rev. 33:609–615. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Maeda Y, Farina NH, Matzelle MM, Fanning
PJ, Lian JB and Gravallese EM: Synovium-derived MicroRNAs regulate
bone pathways in rheumatoid arthritis. J Bone Miner Res.
32:461–472. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou W, Zou B, Liu L, Cui K, Gao J, Yuan S
and Cong N: MicroRNA-98 acts as a tumor suppressor in
hepatocellular carcinoma via targeting SALL4. Oncotarget.
7:74059–74073. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Choi E, Choi E and Hwang KC: MicroRNAs as
novel regulators of stem cell fate. World J Stem Cells. 5:172–187.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hou YY, Lee JH, Chen HC, Yang CM, Huang
SJ, Liou HH, Chi CC, Tsai KW and Ger LP: The association between
miR-499a polymorphism and oral squamous cell carcinoma progression.
Oral Dis. 21:195–206. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao M, Zhang H, Zhu G, Liang J, Chen N,
Yang Y, Liang X, Cai H and Liu W: Association between
overexpression of Wip1 and prognosis of patients with non-small
cell lung cancer. Oncol Lett. 11:2365–2370. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yin S, Wang P, Yang L, Liu Y, Wang Y, Liu
M, Qi Z, Meng J, Shi TY, Yang G and Zang R: Wip1 suppresses ovarian
cancer metastasis through the ATM/AKT/Snail mediated signaling.
Oncotarget. 7:29359–29370. 2016.PubMed/NCBI
|
|
13
|
Zhang X, Wan G, Mlotshwa S, Vance V,
Berger FG, Chen H and Lu X: Oncogenic Wip1 phosphatase is inhibited
by miR-16 in the DNA damage signaling pathway. Cancer Res.
70:7176–7186. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu S, Qi L, Han W, Wan X, Jiang S, Li Y,
Xie Y, Liu L, Zeng F, Liu Z and Zu X: Overexpression of wip1 is
associated with biologic behavior in human clear cell renal cell
carcinoma. PLoS One. 9:e1102182014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Deng J, Zhang R, Pan Y, Wang B, Wu L, Jiao
X, Bao T, Hao X and Liang H: Comparison of the staging of regional
lymph nodes using the sixth and seventh editions of the
tumor-node-metastasis (TNM) classification system for the
evaluation of overall survival in gastric cancer patients: Findings
of a case-control analysis involving a single institution in China.
Surgery. 156:64–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dang X, Ma A, Yang L, Hu H, Zhu B, Shang
D, Chen T and Luo Y: MicroRNA-26a regulates tumorigenic properties
of EZH2 in human lung carcinoma cells. Cancer Genet. 205:113–123.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qi P, Cheng SQ, Wang H, Li N, Chen YF and
Gao CF: Serum microRNAs as biomarkers for hepatocellular carcinoma
in Chinese patients with chronic hepatitis B virus infection. PLoS
One. 6:e284862011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shao XJ, Miao MH, Xue J, Xue J, Ji XQ and
Zhu H: The down-regulation of MicroRNA-497 contributes to cell
growth and cisplatin resistance through PI3K/Akt pathway in
osteosarcoma. Cell Physiol Biochem. 36:2051–2062. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang H, Wang Y, Xu T, Li C, Wu J, He Q,
Wang G, Ding C, Liu K, Tang H and Ji F: Increased expression of
microRNA-148a in osteosarcoma promotes cancer cell growth by
targeting PTEN. Oncol Lett. 12:3208–3214. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Geng S, Gu L, Ju F, Zhang H, Wang Y, Tang
H, Bi Z and Yang C: MicroRNA-224 promotes the sensitivity of
osteosarcoma cells to cisplatin by targeting Rac1. J Cell Mol Med.
20:1611–1619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qu F, Li CB, Yuan BT, Qi W, Li HL, Shen
XZ, Zhao G, Wang JT and Liu YJ: MicroRNA-26a induces osteosarcoma
cell growth and metastasis via the Wnt/β-catenin pathway. Oncol
Lett. 11:1592–1596. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang K, Zhang Y, Ren K, Zhao G, Yan K and
Ma B: MicroRNA-101 inhibits the metastasis of osteosarcoma cells by
downregulation of EZH2 expression. Oncol Rep. 32:2143–2149. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun XH, Geng XL, Zhang J and Zhang C:
miRNA-646 suppresses osteosarcoma cell metastasis by downregulating
fibroblast growth factor 2 (FGF2). Tumour Biol. 36:2127–2134. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li E, Zhang J, Yuan T and Ma B: MiR-145
inhibits osteosarcoma cells proliferation and invasion by targeting
ROCK1. Tumour Biol. 35:7645–7650. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao D, Jia P, Wang W and Zhang G:
VEGF-mediated suppression of cell proliferation and invasion by
miR-410 in osteosarcoma. Mol Cell Biochem. 400:87–95. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ruan WD, Wang P, Feng S, Xue Y and Zhang
B: MicroRNA-497 inhibits cell proliferation, migration, and
invasion by targeting AMOT in human osteosarcoma cells. Onco
Targets Ther. 9:303–313. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li J, Yang Y, Peng Y, Austin RJ, van
Eyndhoven WG, Nguyen KC, Gabriele T, McCurrach ME, Marks JR, Hoey
T, et al: Oncogenic properties of PPM1D located within a breast
cancer amplification epicenter at 17q23. Nat Genet. 31:133–134.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hirasawa A, Saito-Ohara F, Inoue J, Aoki
D, Susumu N, Yokoyama T, Nozawa S, Inazawa J and Imoto I:
Association of 17q21-q24 gain in ovarian clear cell adenocarcinomas
with poor prognosis and identification of PPM1D and APPBP2 as
likely amplification targets. Clin Cancer Res. 9:1995–2004.
2003.PubMed/NCBI
|
|
30
|
Loukopoulos P, Shibata T, Katoh H, Kokubu
A, Sakamoto M, Yamazaki K, Kosuge T, Kanai Y, Hosoda F, Imoto I, et
al: Genome-wide array-based comparative genomic hybridization
analysis of pancreatic adenocarcinoma: Identification of genetic
indicators that predict patient outcome. Cancer Sci. 98:392–400.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ehrbrecht A, Müller U, Wolter M, Hoischen
A, Koch A, Radlwimmer B, Actor B, Mincheva A, Pietsch T, Lichter P,
et al: Comprehensive genomic analysis of desmoplastic
medulloblastomas: Identification of novel amplified genes and
separate evaluation of the different histological components. J
Pathol. 208:554–563. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fuku T, Semba S, Yutori H and Yokozaki H:
Increased wild-type p53-induced phosphatase 1 (Wip1 or PPM1D)
expression correlated with downregulation of checkpoint kinase 2 in
human gastric carcinoma. Pathol Int. 57:566–571. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sun B, Hu X, Liu G, Ma B, Xu Y, Yang T,
Shi J, Yang F, Li H, Zhang L and Zhao Y: Phosphatase Wip1
negatively regulates neutrophil migration and inflammation. J
Immunol. 192:1184–1195. 2014. View Article : Google Scholar : PubMed/NCBI
|