Introduction
Robust angiogenesis and aberrant blood vessel
formation are common characteristics of tumors and are considered
promising therapeutic targets for numerous types of cancer
(1,2).
Owing to the aberrant tumor vasculature, intratumoral hemorrhage is
a common feature of malignancy. The intensity of hemorrhage has
been identified to be associated with the histological grade and
stage of tumors (3). The leakiness of
tumor vessels promotes the extravasation of blood cells and plasma
proteins, as well as metastasis via the bloodstream (4–7).
Therefore, intratumoral hemorrhage serves a critical role in
prognosis.
Regarding the underlying molecular mechanism of
intratumoral hemorrhage, it has been suggested that the
interstitial blood may result from the capillary sprouting, termed
angiogenesis, which is stimulated by vascular endothelial growth
factor A (VEGFA) secreted by tumor cells. Blood escaping the
microvasculature may also be a normal feature resembling the
granulation tissue (5). Intratumoral
hemorrhage has also been attributed to the defective endothelial
lining of tumor vessels, which may be disorganized, loosely
connected, branched, overlapping or sprouting. Opening between
these abnormal endothelial cells may permit blood leakage (5). The intratumoral hemorrhage may range
from scattered blood cells to blood lakes, which are relatively
large regions of extravasated erythrocytes (8–11).
Pancreatic islets are richly vascularized
compartments with a dense network of capillaries. Although they
only account for 1% of the pancreatic mass, they receive between 7
and 10% of the total pancreatic blood flow (12). The endocrine cells release VEGFA,
angiopoietin-2 and insulin, promoting their proliferation (13). It has been documented that endocrine
tumors are highly vascularized (14),
particularly the well-differentiated pancreatic neuroendocrine
tumors (PNETs) (15). Therefore,
computed tomography angiography is used for diagnosing and
localizing small PNETs (16) and
drugs targeting VEGF signaling are used for PNET treatment.
Sunitinib has been approved for the treatment of unresectable or
metastatic progressive well-differentiated PNETs (17). However, the process of tumor
vascularization in PNETs remains unclear. Interestingly, the
microvascular density has been identified to be negatively
associated with tumor progression and the proliferation index of
endothelial cells (15). These
observations suggest a unique mechanism for PNET
vascularization.
In the present study, hemorrhage was investigated in
132 PNET clinical samples. In ~30% of these samples, regardless of
the tumor subtype, a special type of hemorrhagic region with a
smooth boundary unlined by endothelial cells, was observed. On the
basis of observations in clinical samples and the INS-1 xenograft
tumor model, the unique formation steps of these blood-filled
caverns were predicted and the relevant implications are
discussed.
Materials and methods
Clinical samples
The present study was approved by the Scientific
Ethics Committee of the Peking Union Medical College Hospital
(Beijing, China). Participants provided written informed consent
prior to participation. In total, 55 males and 77 females, aged
from 14 to 74 (mean age, 45.09), were recruited from January 1998
to December 2010 in the present study. Pathological sections of
PNETs with hematoxylin and eosin (H&E) staining were collected
and preserved in the Peking Union Medical College Hospital.
Patients were followed up every year and recurrence or metastasis
was recorded.
Cell culture
Rat insulinoma INS-1 cells were kindly provided by
Dr. Haiyan Wang from Division of Clinical Biochemistry, Geneva
University Medical Center (Geneva, Switzerland). INS-1 cells were
cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) containing 11.2 mM glucose and supplemented
with 10% fetal calf serum (FCS; Biological Industries Israel
BeitHaemek, Kibbutz-BeitHaemek, Israel), 50 µM 2-mercaptoethanol,
105 U/l penicillin and 100 mg/l streptomycin at 37°C
with 5% CO2.
Xenograft tumor models
The Animal Studies Committee of the China-Japan
Friendship Hospital (Beijing, China) approved the animal studies,
which were conducted according to the Principles of Laboratory
Animal Care. A total of 6 6–8-week-old female nude mice (Charles
River Laboratories, Beijing, China) weighing ~20 g were used in the
model. Mice were maintained under specific-pathogen-free conditions
at room temperature (between 20–24°C), with humidity between
35–55%, in a 12/12 h light/dark cycle, with food and water ad
libitum. INS-1 cells (1×106 in 100 µl FCS-free
culture medium) were subcutaneously injected into the back of the
mice and the glycemic level was monitored. Following 2–3 weeks,
when the glycemic level was <3.9, each mouse were euthanized by
the intraperitoneal injection of 0.1 ml of 200 mg/ml pentobarbitone
sodium and tumors with diameters between 0.53 and 0.77 cm were
collected for histological examination.
H&E, periodic acid-Schiff (PAS)
and Picro Sirius red staining
INS-1 tumors were fixed in 4% paraformaldehyde
(PFA)/PBS at 4°C overnight and embedded in paraffin. The slides
with 5 µm in thickness were rehydrated and stained with Harris's H
for 10 min and then with an EY solution for an additional 5 min.
Polysaccharide staining was performed using the PAS staining kit
(BasoDiagnostics Inc., Zhuhai, China). For connective tissue
staining, the slides were rehydrated and stained with Harris's H
for 10 min, washed in tap water, stained with Picro Sirius red
(Sigma Aldrich; Merck KGaA, Darmstadt, Germany) for 1 h and finally
washed in acidified water. The slides were observed under a light
microscope at magnifications of ×40-×400.
Immunohistochemistry/immunofluorescence
INS-1 tumors were fixed in 4% PFA/PBS for 2 h at
4°C, cryoprotected in a 30% sucrose/PBS solution at 4°C
overnightand then embedded in Tissue-Tek® O.C.T.
compound (version E, Sakura Finetek USA Inc., Radnor, PA, USA) and
preserved at −80°C. For frozen sections, the samples were
pre-warmed in −20°C for 30 min, and then cut into 8 µm sections.
The primary antibodies used were: Anti-mouse cluster of
differentiation 31 (CD31) (rat, 1:50, cat. no., 553370, BD
Biosciences, Franklin Lakes, NJ, USA), anti-epithelial (E-)
cadherin (mouse, 1:100, cat. no., 610181, BD Biosciences),
anti-β-catenin (mouse, 1:100, cat. no., 610154, BD Biosciences),
anti-insulin (rabbit, 1:200, cat. no., sc-9168, Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) and anti-laminin (rabbit,
1:100, cat. no., AB-19012, EMD Millipore, Billerica, MA, USA). The
samples were incubated with the primary antibodies at 4°C
overnight. For immunohistochemistry, anti-rat (cat. no., PV-6004,
ready to use, OriGene Technologies, Inc., Beijing, China) and
anti-rabbit (cat. no., PV-6001, ready to use, OriGene Technologies,
Inc.) kits were applied for the secondary antibody incubation and
detection procedures, according to the protocols of the
manufacturer. The samples were incubated with the secondary
antibodies for 30 min at room temperature (RT). For
immunofluorescence, Alex Fluor® 555 donkey anti-mouse
(1:400, cat. no., A-31570, Invitrogen; Thermo Fisher Scientific,
Inc.) and Alex Fluor® 488 donkey anti-rabbit (1:400,
cat. no., A-21206, Invitrogen) were used as secondary antibodies
and applied to the samples for 2 h at RT. Fluorescence images were
captured by Leica DM4000B fluorescence microscope with
magnification, ×200.
Statistical analysis
To determine the diameter of the blood-filled
caverns unlined by endothelial cells, five typical blood-filled
caverns were randomly selected in each of the 16 clinical slides
and their diameter was determined using Photoshop®
version 6.0 (Adobe Systems Inc., San Jose, CA, USA). For the
association of the occurrence of these caverns with other
clinicopathological features a χ2 test was used to
examine the sampling distribution of the statistical hypothesis.
Additionally, a Student's t-test was performed to compare the onset
age between two different groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
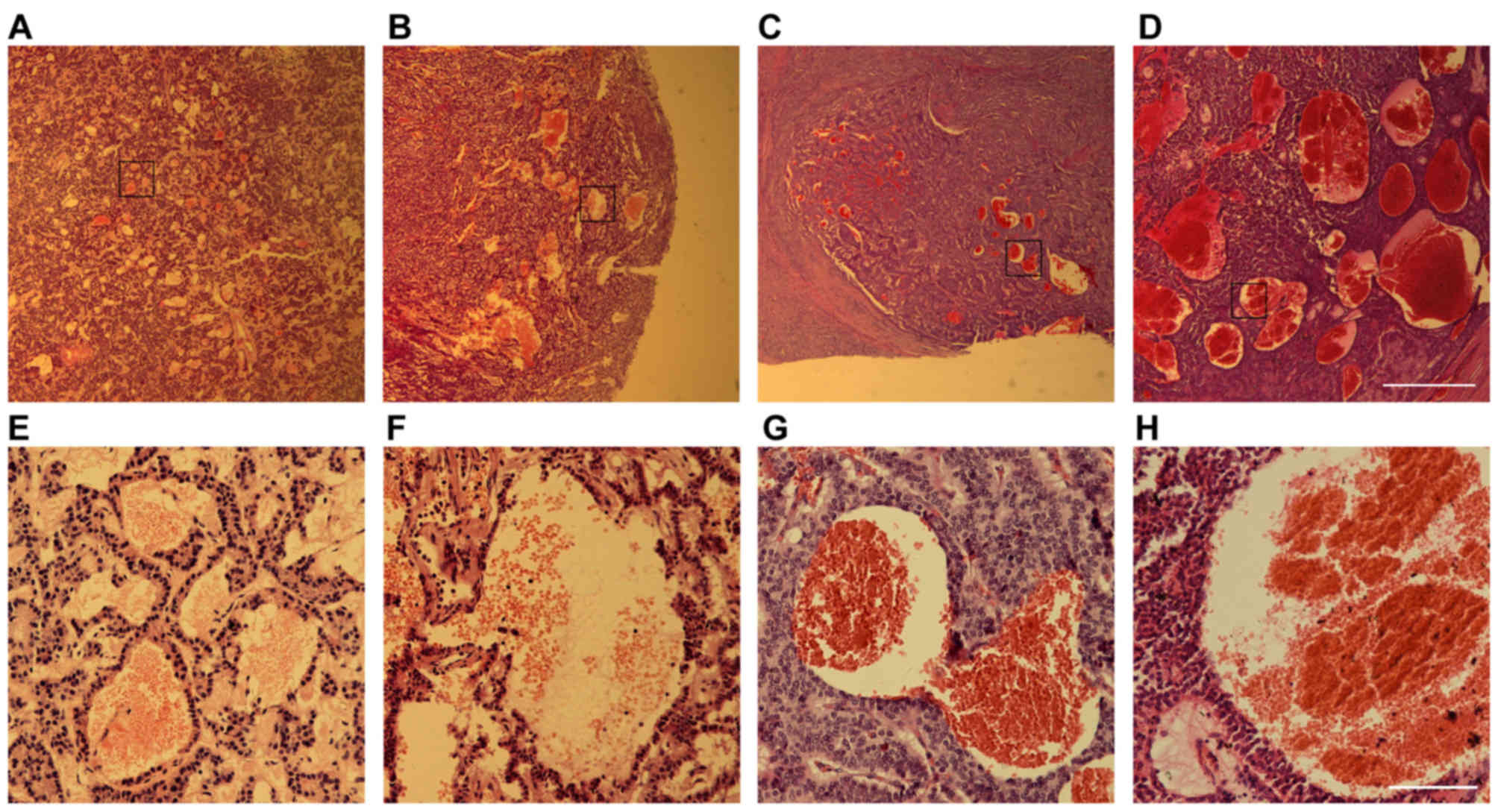
Blood-filled caverns unlined by
endothelial cells exist in ~30% of PNET clinical samples
A total of 132 PNET pathological samples, including
98 insulinoma cases, 3 glucagonoma, 9 gastrinoma and 22
non-functional PNETs, were examined. In addition to dilated blood
vessels and scattered hemorrhagic regions, which are usually common
in other types of tumors as well, a special type of hemorrhage was
observed in ~30% of the samples tested, regardless of the tumor
subtype (Fig. 1). These hemorrhagic
regions were presented as blood-filled caverns with a smooth
boundary and were unlined by endothelial cells. Their size varied
from <100 µm to 1.32 mm as observed in a non-functional PNET
sample (Fig. 1D).
To investigate whether the blood-filled caverns had
any clinical significance, their association with several
clinicopathological features was examined. However, no
statistically significant association with the tumor subtype,
presence of metastasis, disease recurrence, patient sex or age of
onset was observed (Table I). This
suggests that blood-filled caverns unlined by endothelial cells may
present a common feature of PNETs.
 | Table I.PNETs with or without blood-filled
caverns unlined by endothelial cells. |
Table I.
PNETs with or without blood-filled
caverns unlined by endothelial cells.
|
|
| With blood-filled
caverns unlined by endothelial cells | Without blood-filled
caverns unlined by endothelial cells |
|---|
|
|
|
|
|
|---|
| Tumor type | Total n | n (%) | Malignancy (%) | Male (%) | Mean ± SD age,
years | n (%) | Malignancy (%) | Male (%) | Mean ± SD age,
years |
|---|
| INS | 98 | 26 (26.5) | 1 (3.8) | 17 (65.4) | 46.2±12.4 | 72 (73.5) | 3 (5.8) | 26 (36.1) | 45.8±16.1 |
| GLU | 3 | 2 (66.7) | 1 (50) | 1 (100) | 40 | 1 (33.3) | 1 (100) | 0 (0) | 51 |
| GAS | 9 | 4 (44.4) | 4 (100) | 1 (25) | 43.2±8.0 | 5 (55.6) | 5 (100) | 2 (40) | 47.4±13.6 |
| NF | 22 | 7 (31.8) | 3 (42.9) | 2 (28.6) | 38.6±16.1 | 15 (68.2) | 4 (26.7) | 6 (40) | 42.9±12.1 |
| F | 110 | 32 (29.1) | 6 (18.8) | 19 (59.4) | 45.5±11.6 | 78 (70.9) | 9 (11.5) | 28 (35.9) | 45.9±15.7 |
| Total | 132 | 39 (29.5) | 9 (23.1) | 21 (53.8) | 44.2±12.6 | 93 (70.5) | 13 (14.0) | 34 (36.6) | 45.5±15.2 |
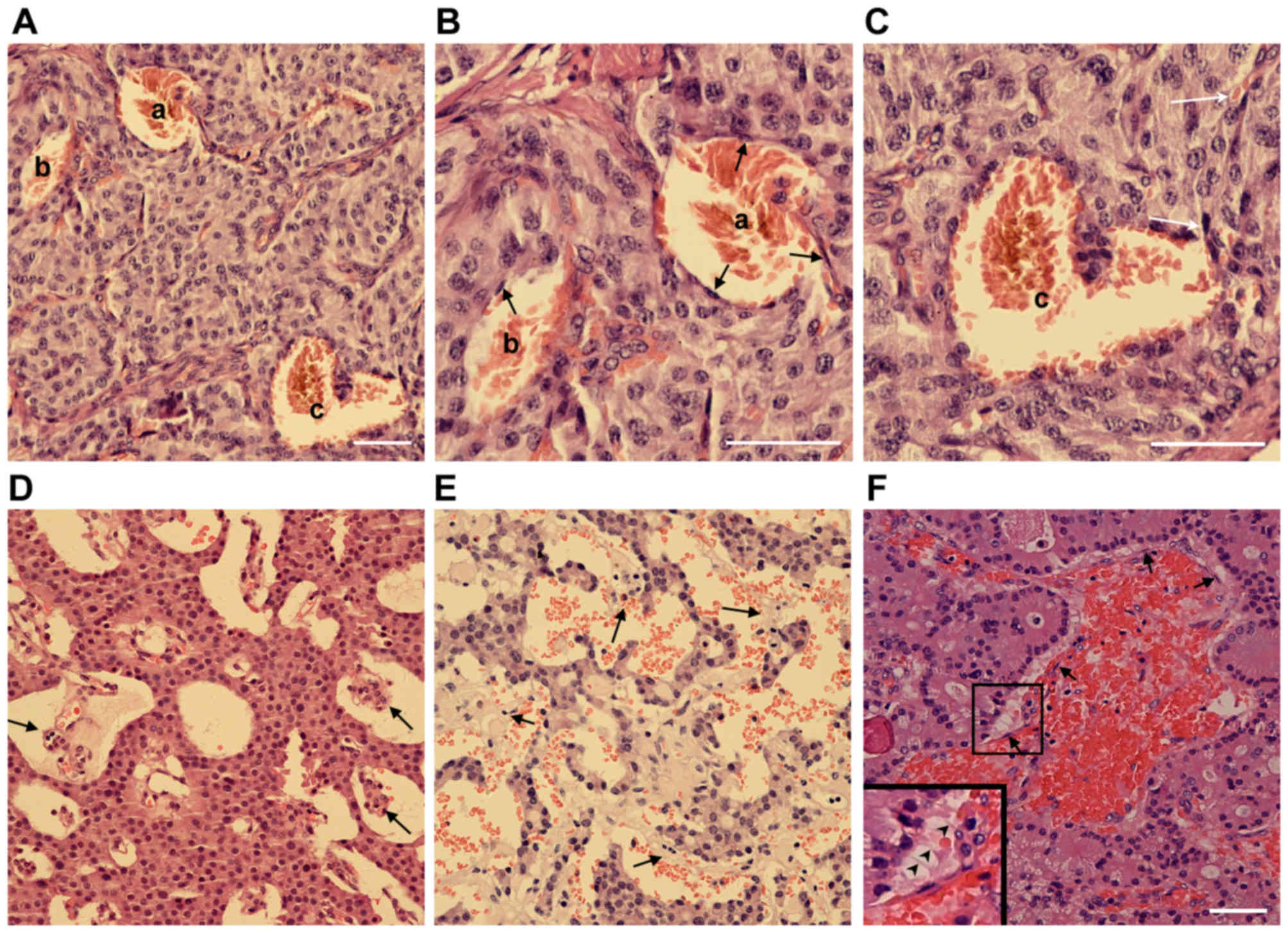
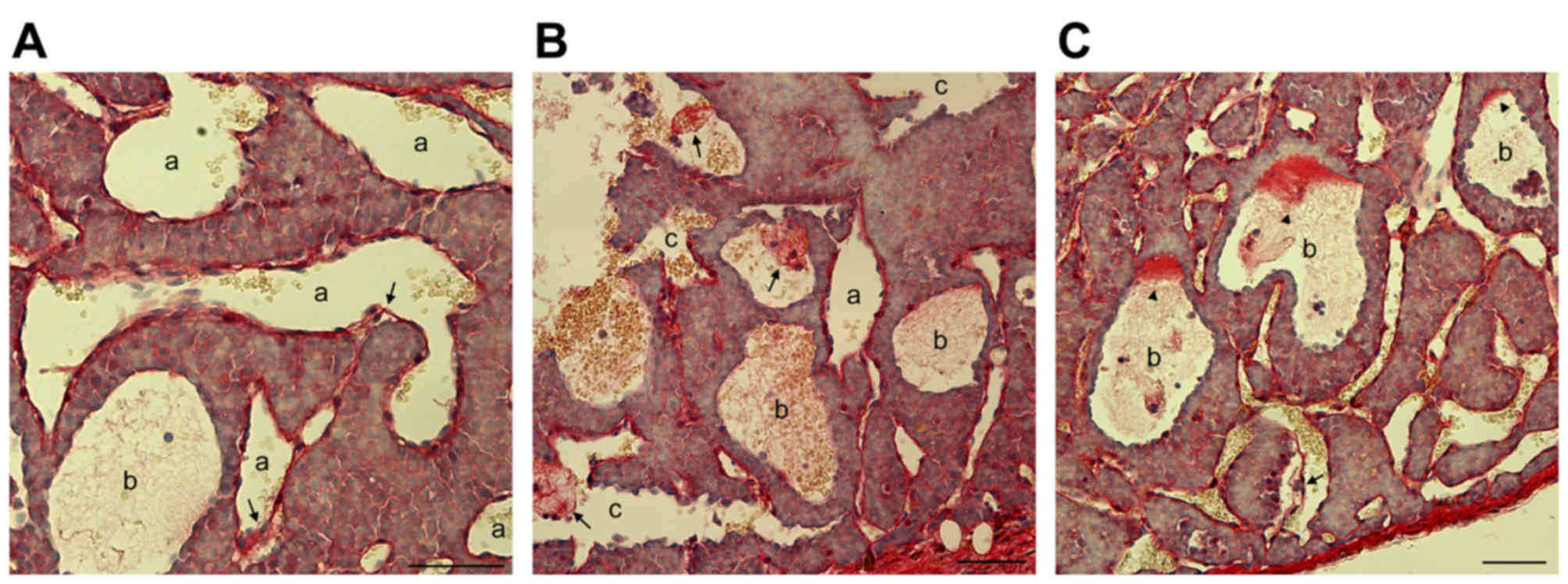
Extensive endothelial cell detachment
is present in clinical PNET samples
In certain PNET clinical samples, it was observed
that not every blood filled-cavern was unlined by endothelial
cells. By contrast, a number of them were partially lined by
endothelial cells. In certain cases, caverns unlined or (partially)
lined with endothelial cells were observed in the same tumor sample
(Fig. 2A-C). In addition, the blood
filled-caverns unlined by endothelial cells were associated with
blood vessels lined by endothelial cells (Fig. 2C). In certain blood-filled caverns,
the endothelial cells were detached from the peripheral tumor cells
(Fig. 2D-F). On the basis of these
observations, it was hypothesized that these blood-filled caverns,
which were unlined by endothelial cells, were generated by the
blind ends of dilated blood vessels lined by endothelial cells.
Further dilation of the endothelial cells may cause detachment from
the surrounding tumor cell layers and subsequent direct contact of
blood cells with the lined tumor tissue. Morphologically, as the
blood cells were mainly reserved within the original endothelial
caverns and not disseminated into the tumor tissue, their shape was
retained.
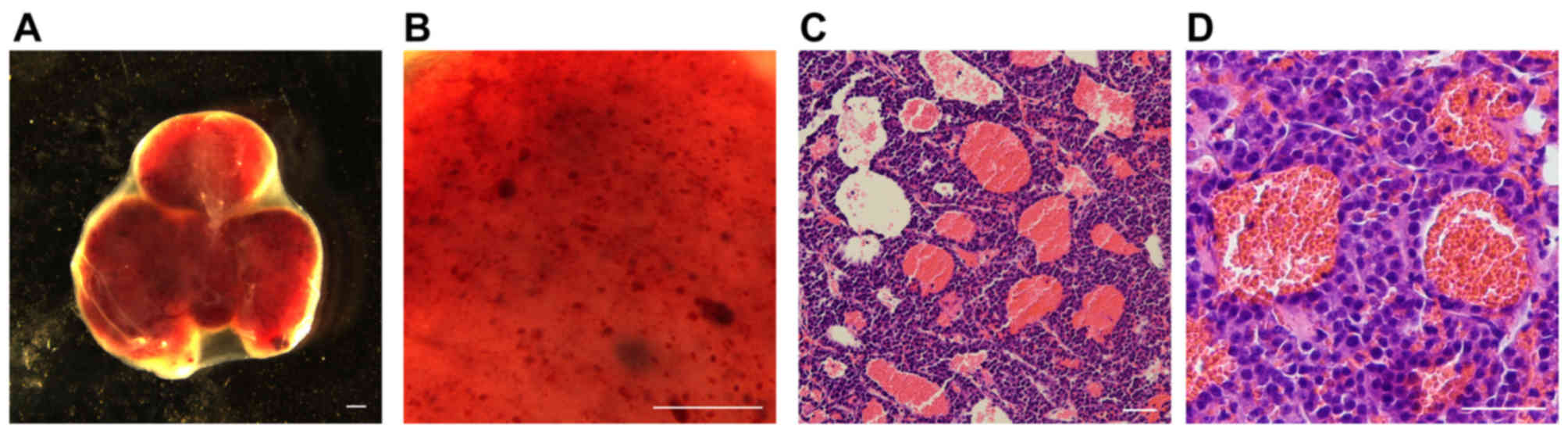
Blood-filled cavern formation in an
INS-1 xenograft mouse model
To further investigate the development of these
blood-filled caverns in PNETs, a xenograft tumor model was
generated using INS-1 cells derived from X-ray-induced rat
insulinoma (18). A total of
106 INS-1 cells were subcutaneously injected into nude
mice. Tumors were formed in the subsequent 2–3 weeks and the mice
developed hypoglycemia, which was lethal if no treatment was
provided. Abundant blood-filled caverns with a regular shape were
observed within the tumors (Fig. 3A).
The largest in size were able to be observed as blood spots under a
stereomicroscope (Fig. 3B). The
majority of the blood-filled caverns in the INS-1 tumors was
unlined by endothelial cells (Fig. 3C and
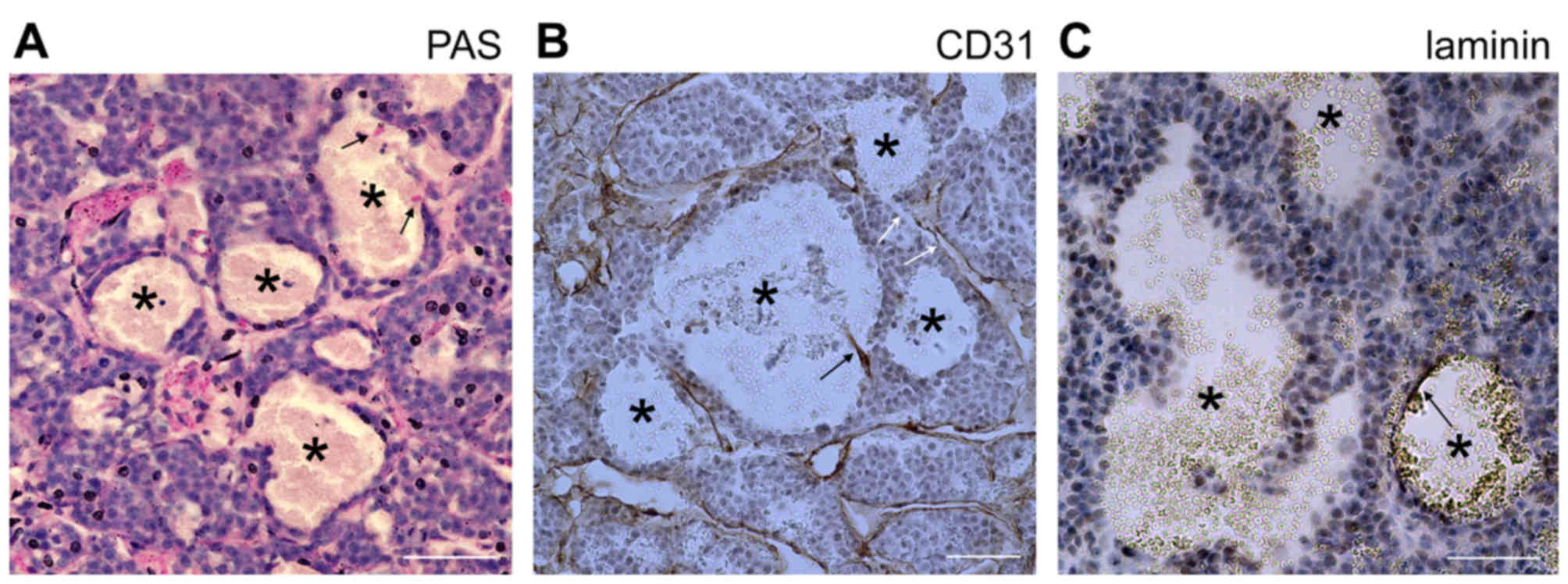
D) and stained negative for CD31, PAS and laminin (Fig. 4); indicating that they lack not only
the surrounding endothelial cells, but also the laminin basement
membrane which normally separates the endothelial from the tumor
cells. Furthermore, this result suggests that the surrounding tumor
cells that stained negative for PAS and laminin were distinct from
the endothelial-like cells, generated through vasculogenic
mimicry.
In certain endothelial-unlined blood-filled caverns,
CD31-positive cell debris (Fig. 4B)
or a fragment of laminin basement membrane was observed (Fig. 4C), suggesting the previous existence
of endothelial cells. In addition, certain blood-filled caverns
were directly linked to normal vasculature (Fig. 4B), suggesting that they were not
isolated hemorrhagic regions. Using Sirius red staining to detect
collagen expression in blood vessels, endothelial cells in
different formation stages of blood-filled caverns were observed.
In specific dilated blood vessels, the endothelial cells were
almost detached from the adjacent tumor cells, in other
blood-filled caverns the detached endothelial cells formed a
cluster and in blood-filled caverns unlined by endothelial cells
collagen deposition were only observed in specific sites of the
lakeside (Fig. 5). The results of the
INS-1 xenograft insulinoma model supported the hypothesis that the
blood-filled caverns unlined by endothelial cells were generated
from blood vessel dilation followed by endothelial detachment. This
process is distinct from the traditional ‘hemorrhage’, which is
defined as blood cell leakage from damaged vasculature resulting in
irregular and diffusive hemorrhage in tumor tissue. Therefore, it
is proposed that this specific type of hemorrhage is named
‘pseudo-hemorrhage’.
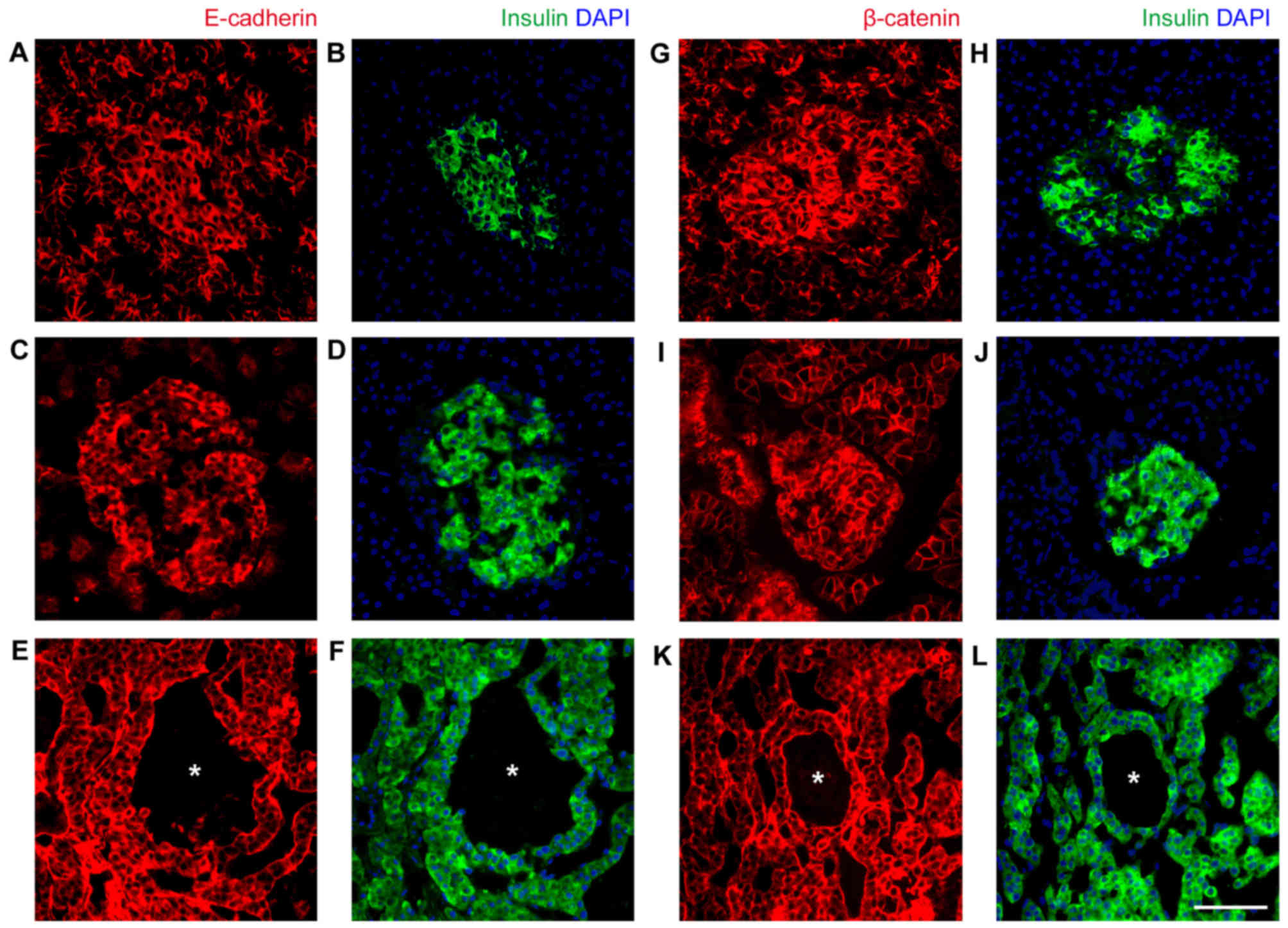
Intact cell-cell adhesion between
tumor cells may be responsible for the formation of
pseudo-hemorrhagic regions
The final question asked in the present study was
why the blood cells did not infiltrate into the tumor tissue, and
were contained within the originally formed caverns, resulting in
their shape remaining intact. Islet cells have an epithelial origin
and the cell-cell junctional protein content is associated with
β-cell maturation (19,20). As E-cadherin and β-catenin were highly
expressed in normal human and rat islets (Fig. 6), it was speculated that the cell-cell
connection of islet tumor cells prevents blood cell infiltration.
As anticipated, E-cadherin and β-catenin were highly expressed and
localized at cell-cell junctions of INS-1 tumor cells (Fig. 6). Therefore, even though no
association of pseudo-hemorrhage with clinical features was
identified, the underlining expression of E-cadherin and β-catenin
suggests that normal islet cell features are retained.
Discussion
In the present study, a special type of hemorrhagic
region was observed in ~30% of the PNET samples examined regardless
of the tumor subtype. These regions exhibited a smooth boundary,
were unlined by endothelial cells and were distinct from the
irregular hemorrhage as this is traditionally described. This type
of hemorrhage was also observed in an INS-1 xenograft tumor model.
On the basis of observations in clinical samples and INS-1 tumor
models, the unique formation steps of these hemorrhagic regions
were predicted. They involved dilation of blood vessels followed by
detachment of endothelial cells from the tumor tissue. This was
distinct from vasculogenic mimicry, or hemorrhage defined as blood
cell leakage from damaged vasculature. Therefore, this type of
hemorrhage was termed ‘pseudo-hemorrhage’.
Regarding the clinical significance of
pseudo-hemorrhage, no statistically significant association was
identified with tumor subtype, presence of metastasis, disease
recurrence, patient sex or age of onset. In previous studies with
genetically modified mice, including a RIP-TAG2 insulinoma model
(21) and aprohormone convertase
2-knockout glucagonoma model (22),
similar hemorrhagic regions resembling blood lakes have also been
reported. This suggests that this type of hemorrhagic region may be
a prominent feature of islet tumors. In a previous study, using
RIP-TAG2 mice injected with Monastral blue, it was demonstrated
that the blood lakes were not connected to the bloodstream
(21). In the present study, clear
morphological evidence was provided suggesting that the
pseudo-hemorrhagic regions, in clinical samples and INS-1 tumors,
are connected to the vascular channels. This is consistent with the
hypothesis that the pseudo-hemorrhagic regions are formed through
endothelial detachment.
The interaction between pancreatic islet cells and
endothelial cells is a major area of interest. In normal pancreatic
islets, the endocrine cells are connected to the capillary network,
and molecule exchange occurs between islet cells and the
bloodstream through fenestrae existing in endothelial cells
(23,24). These are pores ~100 nm covered by a
permeable diaphragm formed by the fusion of apical and basolateral
plasma membranes. It has been demonstrated that islets contain
~5-10-fold more fenestrae than exocrine tissue (24,25).
However, how the endocrine cells induce fenestrae and whether the
weak connection between endocrine and endothelial cells is
associated with fenestrae are poorly understood. It is worthy of
mention that endothelial detachment following islet transplantation
has been reported. It has been demonstrated that in isolated islets
6 weeks following transplantation, ~80% of the grafted microvessels
contained no endothelial cell lining (26). In the INS-1 xenograft tumor model and
PNET clinical samples used in the present study, extensive
endothelial detachment from tumor cells was observed while the
tumor cell-cell adhesion remained intact. It was therefore
hypothesized that the vulnerability of islet endothelial cells or
the relatively weak linkage between endothelial cells/basement
membranes and tumor cells was associated with the special features
of islet endocrine cells; and the underlying molecular mechanism
requires further investigation.
Liver metastasis occurs frequently in PNETs and
epithelial-mesenchymal transition (EMT) serves a critical role in
tumor cells entering the bloodstream, particularly when
intratumoral hemorrhage occurs (6,7,27). However, tumor cells tightly
surrounding the pseudo-hemorrhagic regions suggest a limited
possibility of EMT. The epithelial markers E-cadherin and β-catenin
serve an important role in the localization of tight junction
molecules. It has been demonstrated that E-cadherin deficiency in
stratified skin epithelium may result in failure of water retention
(28,29). In the present study, it was
demonstrated that E-cadherin and β-catenin were highly expressed
and localized at the cell-cell junctions of INS-1 tumor cells
surrounding the pseudo-hemorrhagic regions. This may prevent the
infiltration of blood cells into tumor tissue and retain them
within the original caverns. Therefore, although no statistically
significant association of pseudo-hemorrhage with any
clinicopathological feature was identified, the underlying
expression of E-cadherin and β-catenin suggests that islet cells
may have an epithelial origin.
Acknowledgements
The present study was supported by the National
Nature Science Foundation of China (grant nos. 81302334 and
81370873), the China-Japan Friendship Hospital Youth Science and
Technology Excellence Project (grant nos. 2015-QNYC-B-06 and
2014-QNYC-A-05) and the National Basic Research Program of China
(grant no. 2012CB966402). Dr Peigang Wang (Capital Medical
University, Beijing, China) is thanked for his contribution and
critical comments to the present study.
References
|
1
|
Ferrara N, Hillan KJ, Gerber HP and
Novotny W: Discovery and development of bevacizumab, an anti-VEGF
antibody for treating cancer. Nat Rev Drug Discov. 3:391–400. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arjaans M, Schröder CP, Oosting SF, Dafni
U, Kleibeuker JE and de Vries EG: VEGF pathway targeting agents,
vessel normalization and tumor drug uptake: From bench to bedside.
Oncotarget. 7:21247–2158. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Daldrup H, Shames DM, Wendland M, Okuhata
Y, Link TM, Rosenau W, Lu Y and Brasch RC: Correlation of dynamic
contrast-enhanced MR imaging with histologic tumor grade:
Comparison of macromolecular and small-molecular contrast media.
AJR Am J Roentgenol. 171:941–949. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dvorak HF, Nagy JA, Dvorak JT and Dvorak
AM: Identification and characterization of the blood vessels of
solid tumors that are leaky to circulating macromolecules. Am J
Pathol. 133:95–109. 1988.PubMed/NCBI
|
|
5
|
Van den Brenk HA, Crowe M, Kelly H and
Stone MG: The significance of free blood in liquid and solid
tumours. Br J Exp Pathol. 58:147–159. 1977.PubMed/NCBI
|
|
6
|
Ellis LM and Fidler IJ: Angiogenesis and
metastasis. Eur J Cancer. 32A:1–2460. 1996.
|
|
7
|
Liotta LA: Cancer cell invasion and
metastasis. Sci Am. 266:54–59. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schechter JE, Felicio LS, Nelson JF and
Finch CE: Pituitary tumorigenesis in aging female C57BL/6J mice: A
light and electron microscopic study. Anat Rec. 199:423–432. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liwnicz BH, Wu SZ and Tew JM Jr: The
relationship between the capillary structure and hemorrhage in
gliomas. J Neurosurg. 66:536–541. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schechter J, Ahmad N, Elias K and Weiner
R: Estrogen-induced tumors: Changes in the vasculature in two
strains of rat. Am J Anat. 179:315–323. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Meis-Kindblom JM and Kindblom LG:
Angiosarcoma of soft tissue: A study of 80 cases. Am J Surg Pathol.
22:683–697. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jansson L and Carlsson PO: Graft vascular
function after transplantation of pancreatic islets. Diabetologia.
45:749–763. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cao Z and Wang X: The endocrine role
between β cells and intra-islet endothelial cells. Endocr J.
61:647–654. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Terris B, Scoazec JY, Rubbia L, Bregeaud
L, Pepper MS, Ruszniewski P, Belghiti J, Flèjou J and Degott C:
Expression of vascular endothelial growth factor in digestive
neuroendocrine tumours. Histopathology. 32:133–138. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Couvelard A, O'Toole D, Turley H, Leek R,
Sauvanet A, Degott C, Ruszniewski P, Belghiti J, Harris AL, Gatter
K and Pezzella F: Microvascular density and hypoxia-inducible
factor pathway in pancreatic endocrine tumours: Negative
correlation of microvascular density and VEGF expression with
tumour progression. Br J Cancer. 92:94–101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Katayama A, Iseda I, Tone A, Matsushita Y,
Inoue K, Tsukamoto K, Yamashita H, Yamadori I, Wada J and Hida K:
The usefulness of super-selective computed tomography angiography
(CTA) for diagnosing and localizing a small insulinoma. Intern Med.
49:1983–1986. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Raymond E, Dahan L, Raoul JL, Bang YJ,
Borbath I, Lombard-Bohas C, Valle J, Metrakos P, Smith D, Vinik A,
et al: Sunitinib malate for the treatment of pancreatic
neuroendocrine tumors. N Engl J Med. 364:501–513. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Asfari M, Janjic D, Meda P, Li G, Halban
PA and Wollheim CB: Establishment of 2-mercaptoethanol-dependent
differentiated insulin-secreting cell lines. Endocrinology.
130:167–178. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Santos-Silva JC, Carvalho CP, de Oliveira
RB, Boschero AC and Collares-Buzato CB: Cell-to-cell contact
dependence and junctional protein content are correlated with in
vivo maturation of pancreatic beta cells. Can J Physiol Pharmacol.
90:837–850. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Collares-Buzato CB, Carvalho CP, Furtado
AG and Boschero AC: Upregulation of the expression of tight and
adherens junction-associated proteins during maturation of neonatal
pancreatic islets in vitro. J Mol Histol. 35:811–822. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hashizume H, Baluk P, Morikawa S, McLean
JW, Thurston G, Roberge S, Jain RK and McDonald DM: Openings
between defective endothelial cells explain tumor vessel leakiness.
Am J Pathol. 156:1363–1380. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jones HB, Reens J, Brocklehurst SR, Betts
CJ, Bickerton S, Bigley AL, Jenkins RP, Whalley NM, Morgan D and
Smith DM: Islets of Langerhans from prohormone convertase-2
knockout mice show α-cell hyperplasia and tumorigenesis with
elevated alpha-cell neogenesis. Int J Exp Pathol. 95:29–48. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bearer EL and Orci L: Endothelial
fenestral diaphragms: A quick-freeze, deep-etch study. J Cell Biol.
100:418–428. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Henderson JR and Moss MC: A morphometric
study of the endocrine and exocrine capillaries of the pancreas. Q
J Exp Physiol. 70:347–356. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zanone MM, Favaro E and Camussi G: From
endothelial to beta cells: Insights into pancreatic islet
microendothelium. Curr Diabetes Rev. 4:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lukinius A, Jansson L and Korsgren O:
Ultrastructural evidence for blood microvessels devoid of an
endothelial cell lining in transplanted pancreatic islets. Am J
Pathol. 146:429–435. 1995.PubMed/NCBI
|
|
27
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tunggal JA, Helfrich I, Schmitz A, Schwarz
H, Gunzel D, Fromm M, Kemler R, Krieg T and Niessen CM: E-cadherin
is essential for in vivo epidermal barrier function by regulating
tight junctions. EMBO J. 24:1146–1156. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ligon LA, Karki S, Tokito M and Holzbaur
EL: Dynein binds to beta-catenin and may tether microtubules at
adherens junctions. Nat Cell Biol. 3:913–917. 2001. View Article : Google Scholar : PubMed/NCBI
|