Introduction
Gastric cancer (GC) is a multifactorial disease
(1), which occurs frequently
worldwide and is ranked as the second most common cause of
cancer-associated mortality (13.3%, 2012) (2). Tumorigenesis and tumor development
depend, in part, on the reprogramming of tumor metabolism (3,4). Targeting
tumor metabolism may be a potential therapeutic strategy in
directing treatment for GC (5–7).
Aerobic glycolysis is a phenotypic characteristic of
cancer metabolism, which is known as the ‘Warburg effect’. Pyruvate
kinase (PK) is the key enzyme which catalyzes the dephosphorylation
of phosphoenolpyruvate (PEP) to pyruvate at the final step of
glycolysis to release energy (8). A
total of 4 isoenzymes of pyruvate kinases: PKM1; PKM2; PKL; and
PKR, have been identified in mammals (9). Among them, PKM1 is present in the
majority of adult tissues, and as a splice variant of PKM1, PKM2
has been identified in fetal tissues, adult stem cells and various
cancer cells (10–12). PKM2, which controls the final
rate-limiting step of glycolysis, is crucial for aerobic
glycolysis. Numerous studies have revealed that tumor cells
exclusively express PKM2, including hepatocellular carcinoma
(13), human glioblastoma (14), prostate cancer (15), breast cancer (16), cholangiocarcinoma (17) and gastric cancer (18–20).
Additionally, increased expression levels of PKM2 result in
metastasis and poor prognosis for patients with cancer (21–23).
Thereby, PKM2 is important in the cancer progression as a key
regulator of Aerobic glycolysis. Hence, knockdown PKM2 inhibited
cell proliferation, glucose metabolism and suppressed the growth of
xenografts (24). Targeting PKM2 is a
possible mechanism for reducing the ‘Warburg effect’ of GC and
influencing the tumor microenvironment, which may facilitate the
potential development of PKM2-targeted therapy for GC.
PKM2, is a critical downstream mediator of the
phosphatidylinositol-3-kinase/protein kinase B/mechanistic target
of rapamycin (PI3K/Akt/mTOR) signaling pathway (25). Multiple studies have demonstrated that
the P13K/Akt/mTOR signaling pathway may be associated with cell
proliferation and survival, with the induction of apoptosis and
with tumor glucose metabolism (26,27). The
present study aimed to investigate whether LY294002, a specific
P13K inhibitor, could inhibit proliferation, induce apoptosis and
inhibit the Warburg effect in GC cells, potentially by
downregulating PKM2.
Materials and methods
Cell lines and culture
Various human GC cell lines, including SGC-7901
(moderately differentiated), BGC-823 (poorly differentiated) and
immortalized normal gastric epithelium cells (GES-1), and the human
cervical cancer HeLa cell line were provided by the Department of
Oncology, The Affiliated Drum Tower Hospital of Nanjing University,
Medical School, (Nanjing City, Jiangsu Province). HeLa cell lines
were used as a positive control for the present study. Cells were
cultured in RPMI1640 (Hyclone, Logan, UT, USA) supplemented with
10% fetal bovine serum (Hangzhou Sijiqing Biological Engineering
Materials Co., Ltd., Hangzhou, China) and 100 units/ml penicillin
and 100 µg/ml streptomycin in a humidified air with 5%
CO2 at 37°C (Thermo Fisher Scientific, Inc., Waltham,
MA, USA).
Cell transfection
A pU6 plasmid, synthesized by Shanghai GeneChem Co.,
Ltd., (Shanghai, China), containing siRNA targeting PKM2 mRNA and
empty plasmids as the negative control were transfected into the
SGC-7901 cells using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. Cells were cultured and selected in medium containing 400
mg/ml G418 (Santa Cruz Biotechnology, Inc., Dallas, TX, USA).
Cell viability assay
Cell survival rate was assessed by using a Cell
Counting Kit-8 (CCK-8) assay (KeyGen Biotech Co., Ltd., Jiangsu
Province, P.R. China), according to the manufacturer's protocol.
BGC-823 cells were plated at a density of 1×104
cells/well in 100 µl RPMI-1640 (Hyclone) into 96-well plates and
cultured for 24 h (~80% confluent). Cells were then treated with
LY294002 at various concentrations (0, 10, 20, 50, 100 µmol/l), or
with DMSO (0.2%) as a control, for 24 and 48 h. SGC-7901 cells from
the non-transfected, negative control (empty plasmid) and
PKM2-siRNA groups were plated at a density of 1×104
cells/well in 96-well plates and cultured at 37°C for 24–48 h. The
absorbance was measured at 450 nm using a Cell Counting Kit-8 assay
(Nanjing KeyGen Biotech Co., Ltd., Nanjing, China).
Annexin V-fluorescein isothiocyanate
(FITC) apoptosis assay
BGC-823 cells were plated into 6-well plates at a
density of 1×106 cells/well. Following treatment with
the indicated concentrations of LY294002 (0, 10, 20, 50, 100
µmol/l), or DMSO (0.2%) for 48 h, the cells were dual-stained using
an Annexin V-FITC kit (Nanjing KeyGen Biotech Co., Ltd.). SGC-7901
cells from the non-transfected, negative control and PKM2-siRNA
groups were seeded at a density of 1×106 cells/well in
6-well plates and cultured to 80% confluence for 48 h, and then the
aforementioned dual staining was performed. Apoptotic cells were
detected by flow cytometry using BD FACSCalibur flow cytometer (BD
Biosciences, Franklin Lakes, NJ, U.S.A.). BD FACSDiva software was
used to analyze data (version 7.0, BD Biosciences, U.S.A.).
Western blotting analysis
The protein expression level of PKM2 was evaluated
in each cell line (SGC-7901, BGC-823, GES-1 and HeLa cells) by
western blot analysis. The protein levels of p-Akt, p-mTOR, hypoxia
inducible factor-1α (HIF-1α) and PKM2 were assessed in BGC-823
cells, and PKM2, Glut-1 and LDHA expression levels in SGC-7901
cells of the non-transfected, negative control and PKM2-siRNA
groups were evaluated by western blot analysis. Total cell extracts
were prepared on ice for 30 min in lysis buffer (150 mM NaCl, 50 mM
Tris-HCl, pH 8.0, 0.1% SDS, 0.2% EDTA, 1% Triton X-100, 1% sodium
deoxycholate and 0.01% PMSF) and supplemented with protease
inhibitors (aprotinin, leupeptin, phenylmethylsulfonyl fluoride,
sodium orthovanadate; Roche Diagnostics, Basel, Switzerland), and
centrifuged at 9,660 × g in 4°C (cat. no. 5804R; Eppendorf,
Hamburg, Germany) for 15 min to remove nuclei and cell debris.
Additionally, tumor samples were subjected to homogenate ahead of
total cell extracts. Protein concentration was quantified using the
bicinchoninic assay kit (Nanjing KeyGen Biotech Co., Ltd., China),
according to the manufacturer's protocol. In total, 50 µg of each
protein sample loaded onto SDS-PAGE (10%) and transferred on to
polyvinylidene fluoride membranes (Immobilon®-P;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) using a semi-dry
transfer system (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Non-specific binding was blocked by incubating the membranes in 1X
TBS containing 0.05% Tween-20) supplemented with 5% non-fat dry
milk for 1 h. Blots were incubated at 4°C overnight with monoclonal
rabbit antibodies against PKM2 (cat. no. 4053, dilution, 1:1,000;
Cell Signaling Technology, Inc., Danvers, MA, U.S.A.), p-Akt (cat.
no. 13038, dilution, 1:1,000; Cell Signaling Technology, Inc.),
p-mTOR (cat. no. 5536, dilution, 1:1,000; Cell Signaling
Technology, Inc.), HIF-1α (cat. no. 36169, dilution, 1:1,000; Cell
Signaling Technology, Inc.) glucose transporter-1 (cat. no. 12939,
dilution, 1:1,000; Cell Signaling Technology, Inc.) and lactate
dehydrogenase A (cat. no. 3582, dilution, 1:1,000; Cell Signaling
Technology, Inc.) and a monoclonal mouse antibody to β-actin (cat.
no. sc-130300, 1:3,000; Santa Cruz Biotechnology, Inc., Dallas, TX,
U.S.A.) was used as an internal control. Blots were then incubated
with horseradish peroxidase-conjugated goat anti-mouse antibody
(No. 074-1806, dilution, 1:1,000; KPL, Inc., Gaithersburg, MD, USA)
and/or goat anti-rabbit antibody (cat. no. L3012, dilution,
1:1,000; KPL, Inc.). Antibody staining was visualized using
enhanced chemiluminescence Western Blot Substrate (Pierce; Thermo
Fisher Scientific, Inc.) The images of western blot products were
collected and analyzed using Quantity One V4.31 (Bio-Rad
Laboratories, Inc.).
Immunofluorescence analysis
BGC-823 cells were seeded at a density of
1×106 cells/well into 6-well plates, prior to treatment
with LY294002 (0, 50 or 100 µmol/l) or with DMSO (0.2%) (2 µl/well)
for 48 h at 37°C. Subsequent to washing with PBS 3 times, the cells
were fixed with cold acetone for 10 min at 4°C. Next, the cells
were blocked with 10% normal goat serum (Wuhan Boster Biological
Technology, Ltd., Wuhan, China) for 30 min and probed with an
antibody against PKM2 (cat. no. 4053, dilution, 1:100, Cell
Signaling Technology, Inc.) at 4°C overnight. Alexa Fluor dye
conjugated secondary antibodies (2 mg/ml goat anti-rabbit IgG (H+L)
highly cross-absorbed) (cat. no. R37117, Alexa Fluor 594; dilution,
1:1,000; Invitrogen; Thermo Fisher Scientific, Inc.) were incubated
with cells for 1 h at 20°C to enable the samples to be visualized
under a fluorescent microscope (Axio Imager A1; Carl Zeiss AG,
Oberkochen, Germany). The nuclei were stained using DAPI (2 µg/ml;
Invitrogen; Thermo Fisher Scientific, Inc.) for 15 min under dark
conditions at 20°C.
RNA isolation and reverse
transcription polymerase chain reaction (RT-qPCR) assay
Total RNA from SGC-7901 cells of the
non-transfected, negative control and PKM2-siRNA groups were
extracted using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacture's protocol. The
first-strand cDNA was synthesized using High-Capacity cDNA Reverse
Transcription kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.). RT-primers of LAT-1 mRNAs were designed and synthesized as
follows: Forward, 5′-AGTACCATGCGGGACCATC-3′ and reverse,
5′-GCGTTATCCAGCGTGATTTT-3′ (Invitrogen; Thermo Fisher Scientific,
Inc.). Real-time quantitative polymerase chain reaction (qRT-PCR)
was performed according to the TaqMan Gene Expression Assays
protocol (Applied Biosystems; Thermo Fisher Scientific, Inc.)
(28). The PCR program was as
follows: 95°C for 10 min, followed by 32 cycles of 95°C for 15 sec,
60°C for 30 sec and 72°C for 45 sec. Fold-induction was calculated
using the formula 2−ΔΔCt (29).
LDH activity and lactate assay
BGC-823 cells were seeded at a density of
1×104 cells/well into 96-well plates and subsequently
treated with 0 or 50 µmol/l LY294002 for 48 h. SGC-7901 cells of
the non-transfected, negative control and PKM2-siRNA groups were
cultured for 48 h at 37°C. LDH activity was determined using an LDH
Cytotoxicity Assay kit (Beyotime Institute of Biotechnology,
Haimen, China). Lactate in the culture medium was determined using
a Lactate Assay kit (Nanjing Jiancheng Bioengineering Institute,
Nanjing, China).
Statistical analysis
The data are expressed as the mean ± standard
deviation, and analyzed by using Least Significant Difference and
Student Newman-Keuls methods following analysis of variance and the
Student's t-test method. The data were analyzed using SPSS v.19.0
software (IBM Corp., Armonk, NY, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression of PKM2 is upregulated in
GC cell lines
Western blot analysis demonstrated that, PKM2
expression was significantly increased in GC cell lines (SGC-7901,
BGC-823) and HeLa cells, compared with in the GES-1 cells (Fig. 1A; P<0.05).
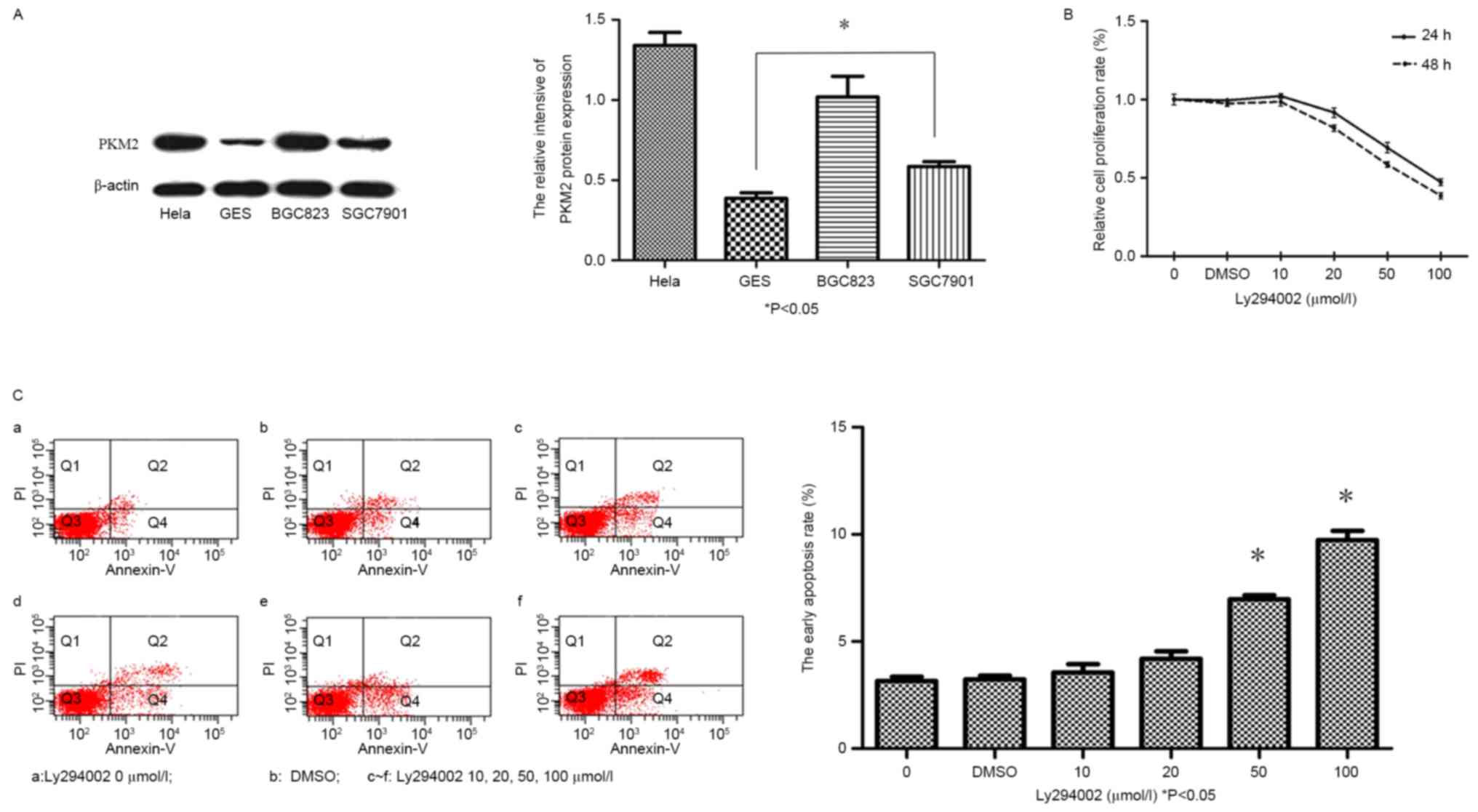 | Figure 1.Expression of PKM2 in GC lines was
significantly upregulated. LY294002 inhibits the viability of and
induces early apoptosis in GC cells. (A) PKM2 protein expression in
GC cell lines (SGC-7901, BGC-823 and HeLa), as detected via western
blot analysis, was significantly upregulated compared with in GES-1
cells. (B) Evaluation of BGC-823 cell viability, following
treatment with LY294002 (0–100 mM) or DMSO for 24 and 48 h. (C)
Evaluation of the early apoptotic rate of BGC823 cells following
treatment with LY294002 (0, 10, 20, 50 or 100 µmol/l) or DMSO for
48 h. *P<0.05 vs. control. PKM2, pyruvate kinase M2; GC, gastric
cancer; LY294002, PKM inhibitor; DMSO, dimethyl sulfoxide. |
Effects of LY294002 on proliferation
and apoptosis in BGC-823 cells
Cell viability of BGC-823 cells, following LY294002
inhibitor treatment was assessed using a CCK-8 assay. As
illustrated in Fig. 1B, LY294002 was
able to inhibit the proliferation of BGC-823 cells in vitro.
Cell viability in the 20 µmol/l LY294002 group was significantly
decreased (91.64±3.06% vs. 100% for the control group; P<0.05),
compared with the control group. This was observed as a dose- and
time-dependent decrease in the viability of BGC-823 cells following
LY294002 treatment. Furthermore, it was demonstrated that, in
response to LY294002 treatment, the early apoptotic rate was
significantly increased in the BGC-823 cells compared with the
control group, in a dose-dependent manner (50 and 100 µmol/l
LY294002; Fig. 1C; P<0.05).
Effects of LY294002 on p-Akt, p-mTOR,
HIF-1α and PKM2 in the BGC-823 cells
To evaluate whether LY294002 can inhibit p-AKT,
p-mTOR, HIF-1α and PKM2 expression, BGC-823 cells treated with 0,
10, 20, 50 or 100 µmol/l LY294002 and DMSO for 48 h were assessed
via western blotting and immunofluorescence. Following 10 µmol/l
LY294002 treatment for 48 h, p-AKT and p-mTOR expression were
decreased when compared with in the control group (P<0.05).
Furthermore, HIF-1α expression was reduced following treatment with
20 µmol/l LY294002 for 48 h; and PKM2 expression decreased
following treatment with 50 µmol/l LY294002 at 48 h. A
dose-dependent association was revealed between the LY294002
concentration and the expression levels of p-AKT, p-mTOR, HIF-1α
and PKM2 (Fig. 2A). In addition, the
intracellular distribution of PKM2 in BGC-823 cells was detected,
revealing that PKM2 was distributed around the nucleus and the
expression decreased following treatment with LY294002 (Fig. 2B).
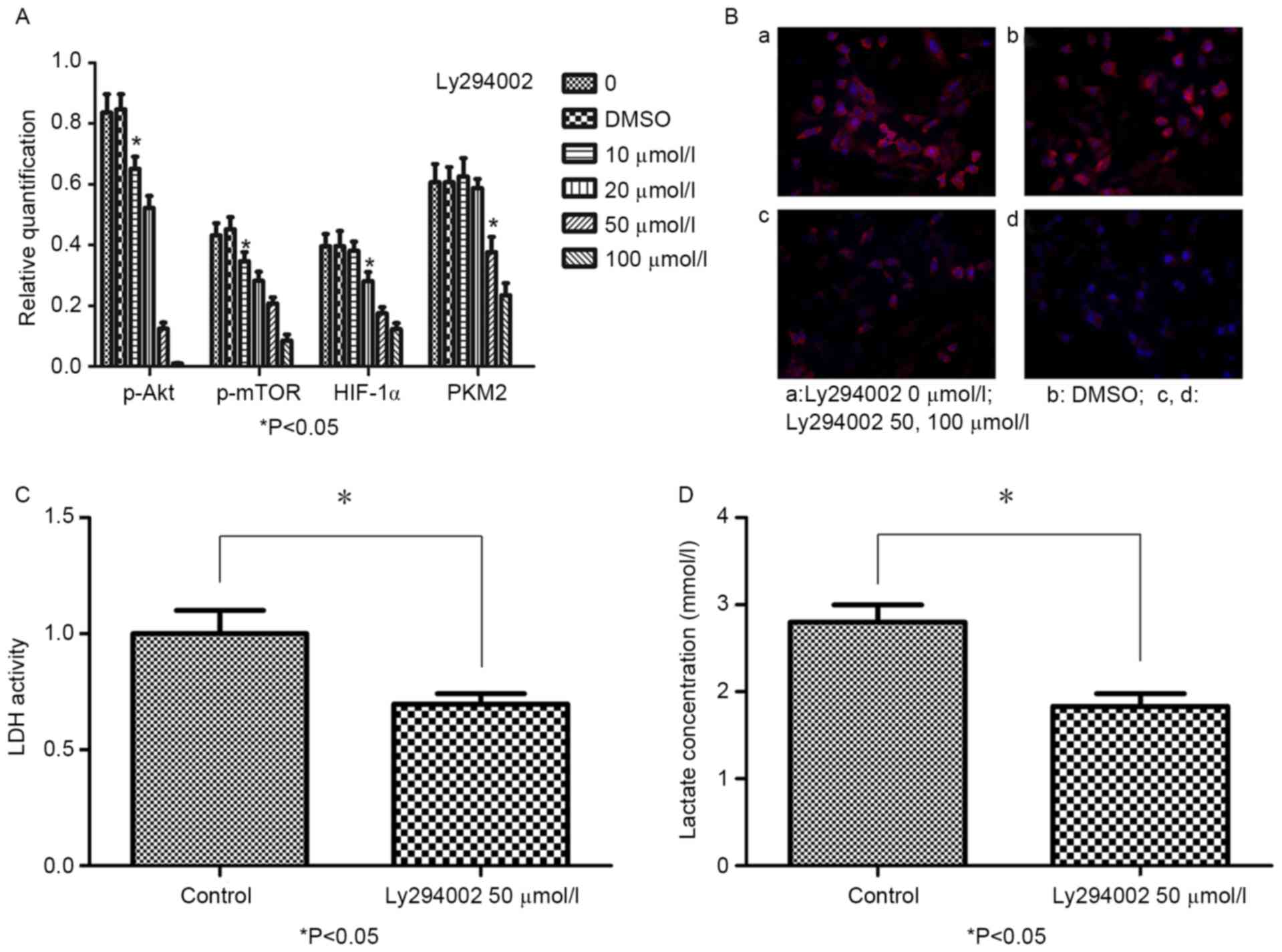 | Figure 2.LY294002 reduces p-AKT, p-mTOR,
HIF-1α and PKM2 protein expression, and inhibits LDH activity and
lactate production in BGC-823 cells. (A) Protein expression of
p-mTOR, HIF-1α and PKM2 in BGC-823 cells treated with LY294002 (0,
10, 20, 50 or 100 µmol/l) or DMSO for 48 h, as detected by western
blot analysis (B) Double immunofluorescence staining revealed the
intracellular distribution of PKM2 in BGC-823 cells prior to and
following LY294002 treatment, at ×200 magnification. (B, a)
Intracellular distribution of PKM2 in BGC-823 cells without
LY294002 treatment. (B, b) Intracellular distribution of PKM2 in
BGC-823 cells with DMSO. (B, c) Intracellular distribution of PKM2
in BGC-823 cells treated with 50 µmol/l LY294002 for 48 h. (B, d)
Intracellular distribution of PKM2 in BGC-823 cells treated with
100 µmol/l LY294002 for 48 h. (C) LDH activity significantly
decreased after 48 h of treatment with 50 µmol/l LY294002. (D)
Lactate production significantly decreased after 48 h of treatment
with 50 µmol/l LY294002. *P<0.05 vs. control. p-AKT,
phospho-protein kinase B; p-mTOR, phospho-mechanistic target of
rapamycin; HIF-1α, hypoxia inducible factor-1α; PKM2, pyruvate
kinase M2; DMSO, dimethyl sulfoxide. |
Effects of LY294002 on LDH activity
and lactate production in BGC-823 cells
To determine the effects of LY294002 on LDH activity
and lactate production in BGC-823 cells, LDH activity and lactate
production were measured. LDH activity (Fig. 2C) and lactate production (Fig. 2D) in the 50 µmol/l LY294002 group were
significantly decreased compared with in the control group
(P<0.05).
Knockdown of PKM2 suppresses
proliferation, induces apoptosis and inhibits the Warburg effect in
SGC7901 cells
PKM2 vectors downregulating the plasmid or control
plasmid (non-transfected group and empty plasmid group) were
transfected into the SGC-7901 cells according to the protocol
aforementioned. The expression levels of PKM2 were analyzed by
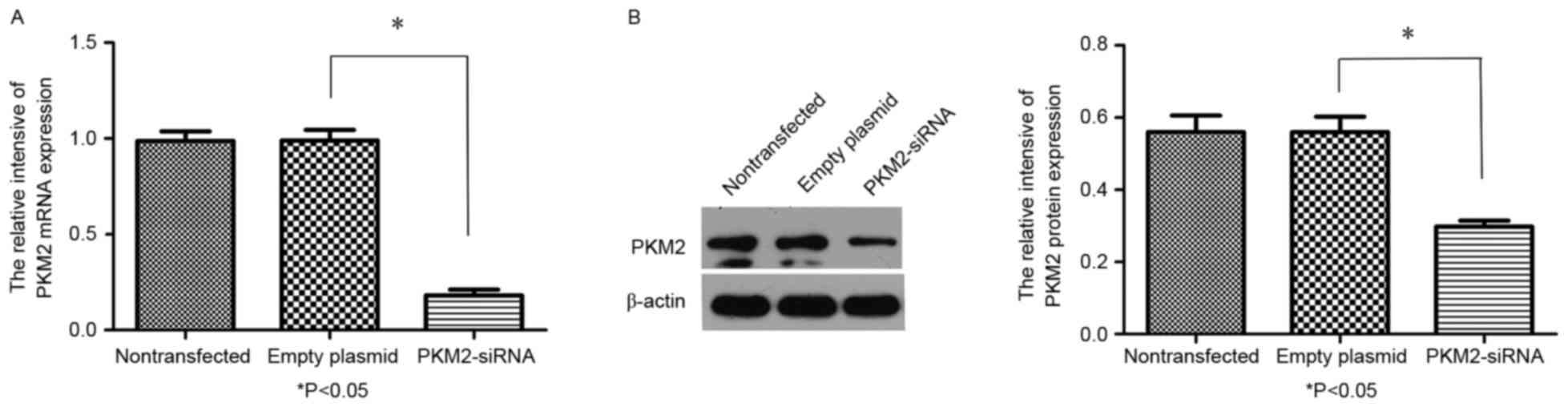
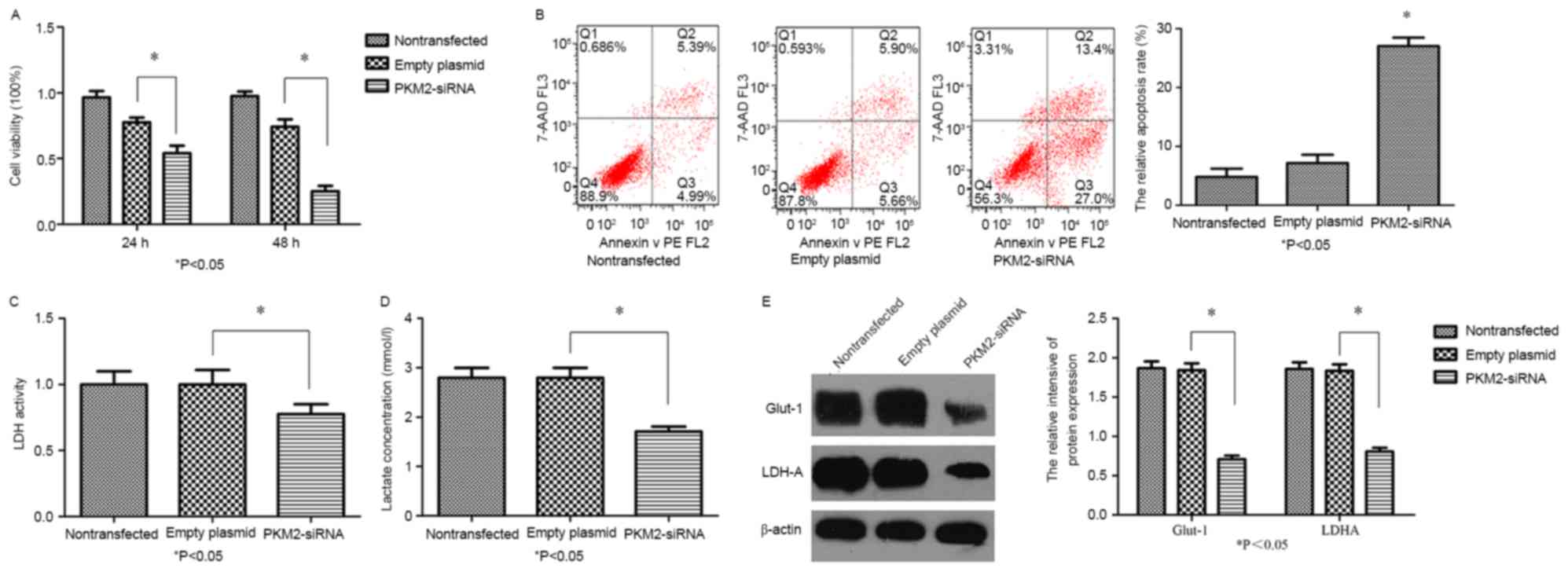
RT-qPCR and immunoblotting. PKM2-siRNA decreased the mRNA (Fig. 3A) and protein (Fig. 3B) levels of PKM2 (P<0.05). A CCK-8
assay demonstrated significant time-dependent inhibition of
SGC-7901 cell proliferation following the knockdown of PKM2
(Fig. 4A; P<0.05). It was further
revealed that the early apoptosis rate of SGC-7901/PKM2-siRNA cells
was increased compared with that of the control cells (Fig. 4B; P<0.05). These results indicated
that PKM2 was able to promote the proliferation and suppress the
apoptosis of GC cells. To determine whether PKM2 knockdown exerts
any influence on the Warburg effect in SGC7901 cells, LDH activity
and lactate production were measured. Compared with the control
groups, LDH activity (Fig. 4C) and
lactate production (Fig. 4D) in
SGC7901 cells of the siRNA-PKM2 group was significantly decreased
(P<0.05). Western blotting demonstrated that the knockdown of
PKM2 significantly decreased Glut-1 and LDHA protein expression
levels (Fig. 4E), compared with in
the non-transfected or empty plasmid groups. In combination, these
outcomes indicate that PKM2-knockdown in SGC-7901 cells was
associated with the Warburg effect.
Discussion
The P13K/AKT/mTOR signaling pathway serves a crucial
role in transduction, which regulates cell proliferation, survival,
migration and metabolism (26,27,30,31).
Accumulating evidence has demonstrated that activation of the
P13K/Akt/mTOR signaling pathway may be involved in the initiation
and progression of numerous types of cancer, including GC (32–38).
Therefore, inhibition of this signaling pathway is widely
considered as an attractive anticancer therapeutic strategy. In the
present study, we demonstrated that a specific P13K inhibitor,
LY294002, inhibited the proliferation, and increased the early
apoptotic rate, of BGC-823 cells in vitro. Furthermore, it
was observed that LDH activity and lactate production was inhibited
by LY294002 in BGC-823 cells. Therefore, targeting the
P13K/Akt/mTOR pathway may suppress cell proliferation and induce
apoptosis, and thus reduce metastasis.
Targeting of the aerobic glycolysis (Warburg effect)
pathway is considered to be a potential promising anticancer
strategy (39). PKM2 is an essential
glycolytic enzyme in this pathway, which alters the Warburg effect
of tumor cells. PKM2 is overexpressed in diverse types of cancer
cells (13–20). Accumulating data has demonstrated that
increased PKM2 expression may contribute to the rapid growth of
tumors, and induce apoptosis in multiple types of human cancer,
including GC (36–45). PKM2 is a critical downstream target of
P13K/Akt/mTOR signaling pathway (46,47). In
the present study, we observed significantly increased PKM2
expression in GC cell lines (SGC-7901, BGC-823), which is
indicative of the importance of PKM2 to GC progression. The present
study also revealed that LY294002 reduced p-Akt, p-mTOR and HIF-1α
protein expression in BGC-823 cells, and inhibited PKM2 protein
expression, following high concentration (50 µmol/l) treatment.
These data suggest that LY294002 may reduce tumor cell
proliferation and aerobic glycolysis by inhibiting the
P13K/Akt/mTOR/PKM2 signaling pathway.
To verify that PKM2 mediates the effects of LY294002
on GC cell proliferation, apoptosis and metastasis, we transfected
a PKM2-knockdown plasmid into SGC-7901 cells. It was confirmed that
cell proliferation was suppressed in the cells transfected with
this siRNA-PKM2. We further demonstrated that PKM2-knockdown
increased the early apoptosis rate of SGC-7901 cells. Furthermore,
the results from the present study also demonstrated that
downregulation of PKM2 significantly decreased LDH activity and
lactate production; and that it reduced Glut-1 and LDHA protein
expression levels. These data suggest that LY294002 was able to
inhibit GC cell proliferation, induce apoptosis and reduce the
Warburg effect, partly by inhibiting PKM2.
In conclusion, the present study demonstrated
significantly increased PKM2 expression in GC cell lines, and
revealed that LY294002 markedly inhibited the proliferation,
induced the apoptosis and inhibited the Warburg effect of GC cells.
Therefore, the anticancer activities of LY294002 may be partly
linked to the downregulation of PKM2.
Acknowledgements
The present study was supported by the Outstanding
Youth Project of Nanjing City (grant no. JQX14005) and the National
Science Foundation (grant nos. 81272742, 81401974, 81400306 and
81401977).
References
|
1
|
Wadhwa R, Song S, Lee JS, Yao Y, Wei Q and
Ajani JA: Gastric cancer-molecular and clinical dimensions. Nat Rev
Clin Oncol. 10:643–655. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Masoudi-Nejad A and Asgari Y: Metabolic
cancer biology: Structural-based analysis of cancer as a metabolic
disease, new sights and opportunities for disease treatment. Semin
Cancer Biol. 30:21–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pavlova NN and Thompson CB: The emerging
hallmarks of cancer metabolism. Cell Metab. 23:27–47. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shen CT, Wei WJ, Qiu ZL, Song HJ and Luo
QY: Afamin promotes glucose metabolism in papillary thyroid
carcinoma. Mol Cell Endocrinol. 434:108–115. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jan CI, Tsai MH, Chiu CF, Huang YP, Liu CJ
and Chang NW: Fenofibrate suppresses oral tumorigenesis via
reprogramming metabolic processes: Potential drug repurposing for
oral cancer. Int J Biol Sci. 12:786–798. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Navarro P, Bueno MJ, Zagorac I, Mondejar
T, Sanchez J, Mourón S, Muñoz J, Gómez-López G, Jimenez-Renard V,
Mulero F, et al: Targeting tumor mitochondrial metabolism overcomes
resistance to antiangiogenics. Cell Rep. 15:2705–2718. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li Z, Yang P and Li Z: The multifaceted
regulation and functions of PKM2 in tumor progression. Biochim
Biophys Acta. 1846:285–296. 2014.PubMed/NCBI
|
|
9
|
Jurica MS, Mesecar A, Heath PJ, Shi W,
Nowak T and Stoddard BL: The allosteric regulation of pyruvate
kinase by fructose-1,6-bisphosphate. Structure. 6:195–210. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tamada M, Suematsu M and Saya H: Pyruvate
kinase M2: Multiple faces for conferring benefits on cancer cells.
Clin Cancer Res. 18:5554–5561. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Christofk HR, Vander Heiden MG, Harris MH,
Ramanathan A, Gerszten RE, Wei R, Fleming MD, Schreiber SL and
Cantley LC: The M2 splice isoform of pyruvate kinase is important
for cancer metabolism and tumour growth. Nature. 452:230–233. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Christofk HR, Vander Heiden MG, Wu N,
Asara JM and Cantley LC: Pyruvate kinase M2 is a
phosphotyrosine-binding protein. Nature. 452:181–186. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu WR, Tian MX, Yang LX, Lin YL, Jin L,
Ding ZB, Shen YH, Peng YF, Gao DM, Zhou J, et al: PKM2 promotes
metastasis by recruiting myeloid-derived suppressor cells and
indicates poor prognosis for hepatocellular carcinoma. Oncotarget.
6:846–861. 2015.PubMed/NCBI
|
|
14
|
Mukherjee J, Ohba S, See WL, Phillips JJ,
Molinaro AM and Pieper RO: PKM2 uses control of HuR localization to
regulate p27 and cell cycle progression in human glioblastoma
cells. Int J Cancer. 139:99–111. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wong N, Yan J, Ojo D, De Melo J, Cutz JC
and Tang D: Changes in PKM2 associate with prostate cancer
progression. Cancer Invest. 32:330–338. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shashni B, Sakharkar KR, Nagasaki Y and
Sakharkar MK: Glycolytic enzymes PGK1 and PKM2 as novel
transcriptional targets of PPARγ in breast cancer pathophysiology.
J Drug Target. 21:161–174. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu G, Yu W, Jin G, Xu D, Chen Y, Xia T, Yu
A, Fang W, Zhang X, Li Z and Xie K: PKM2 regulates neural invasion
of and predicts poor prognosis for human hilar cholangiocarcinoma.
Mol Cancer. 14:1932015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen G, Feng W, Zhang S, Bian K, Yang Y,
Fang C, Chen M, Yang J and Zou X: Metformin inhibits gastric cancer
via the inhibition of HIF1α/PKM2 signaling. Am J Cancer Res.
5:1423–1434. 2015.PubMed/NCBI
|
|
19
|
Gao Y, Xu D, Yu G and Liang J:
Overexpression of metabolic markers HK1 and PKM2 contributes to
lymphatic metastasis and adverse prognosis in Chinese gastric
cancer. Int J Clin Exp Pathol. 8:9264–9271. 2015.PubMed/NCBI
|
|
20
|
Tang R, Yang C, Ma X, Wang Y, Luo D, Huang
C, Xu Z, Liu P and Yang L: MiR-let-7a inhibits cell proliferation,
migration, and invasion by down-regulating PKM2 in gastric cancer.
Oncotarget. 7:5972–5984. 2016.PubMed/NCBI
|
|
21
|
Luo W and Semenza GL: Emerging roles of
PKM2 in cell metabolism and cancer progression. Trends Endocrinol
Metab. 23:560–566. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Panasyuk G, Espeillac C, Chauvin C,
Pradelli LA, Horie Y, Suzuki A, Annicotte JS, Fajas L, Foretz M,
Verdeguer F, et al: PPARγ contributes to PKM2 and HK2 expression in
fatty liver. Nat Commun. 3:6722012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen L, Shi Y, Liu S, Cao Y, Wang X and
Tao Y: PKM2: The thread linking energy metabolism reprogramming
with epigenetics in cancer. Int J Mol Sci. 15:11435–11445. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shuch B, Linehan WM and Srinivasan R:
Aerobic glycolysis: A novel target in kidney cancer. Expert Rev
Anticancer Ther. 13:711–719. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun Q, Chen X, Ma J, Peng H, Wang F, Zha
X, Wang Y, Jing Y, Yang H, Chen R, et al: Mammalian target of
rapamycin up-regulation of pyruvate kinase isoenzyme type M2 is
critical for aerobic glycolysis and tumor growth. Proc Natl Acad
Sci USA. 108:pp. 4129–4134. 2011; View Article : Google Scholar : PubMed/NCBI
|
|
26
|
García-Carracedo D, Villaronga MÁ,
Álvarez-Teijeiro S, Hermida-Prado F, Santamaría I, Allonca E,
Suárez-Fernández L, Gonzalez MV, Balbín M, Astudillo A, et al:
Impact of PI3K/AKT/mTOR pathway activation on the prognosis of
patients with head and neck squamous cell carcinomas. Oncotarget.
7:29780–29793. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen S, Fisher RC, Signs S, Molina LA,
Shenoy AK, Lopez MC, Baker HV, Koomen JM, Chen Y, Gittleman H, et
al: Inhibition of PI3K/Akt/mTOR signaling in PI3KR2-overexpressing
colon cancer stem cells reduces tumor growth due to apoptosis.
Oncotarget. 8:50476–50488. 2016.PubMed/NCBI
|
|
28
|
Petrov A, Beer M and Blome S: Development
and validation of a harmonized TaqMan-based triplex real-time
RT-PCR protocol for the quantitative detection of normalized gene
expression profiles of seven porcine cytokines. PLoS One.
9:e1089102014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bauer TM, Patel MR and Infante JR:
Targeting P13Kinase in cancer. Pharmacol Ther. 146:53–60. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ying J, Xu Q, Liu B, Zhang G, Chen L and
Pan H: The expression of the PI3K/AKT/mTOR pathway in gastric
cancer and its role in gastric cancer prognosis. Onco Targets Ther.
8:2427–2433. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhu L, Derijard B, Chakrabandhu K, Wang
BS, Chen HZ and Hueber AO: Synergism of PI3K/Akt inhibition and Fas
activation on colon cancer cell death. Cancer Lett. 354:355–364.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Denoyelle C, Lambert B, Meryet-Figuière M,
Vigneron N, Brotin E, Lecerf C, Abeilard E, Giffard F, Louis MH,
Gauduchon P, et al: miR-491-5p-induced apoptosis in ovarian
carcinoma depends on the direct inhibition of both BCL-XL and EGFR
leading to BIM activation. Cell Death Dis. 5:e14452014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Houédé N and Pourquier P: Targeting the
genetic alterations of the PI3K-AKT-mTOR pathway: Its potential use
in the treatment of bladder cancers. Pharmacol Ther. 145:1–18.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang L, Wu J, Lu J, Ma R, Sun D and Tang
J: Regulation of the cell cycle and PI3K/Akt/mTOR signaling pathway
by tanshinone I in human breast cancer cell lines. Mol Med Rep.
11:931–939. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li Y, Zhang Z, Zhang X, Lin Y, Luo T, Xiao
Z and Zhou Q: A dual PI3K/AKT/mTOR signaling inhibitor miR-99a
suppresses endometrial carcinoma. Am J Transl Res. 8:719–731.
2016.PubMed/NCBI
|
|
37
|
Su CC and Chiu TL: Tanshinone IIA
decreases the protein expression of EGFR and IGFR blocking the
PI3K/Akt/mTOR pathway in gastric carcinoma AGS cells both in vitro
and in vivo. Oncol Rep. 36:1173–1179. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Riquelme I, Tapia O, Espinoza JA, Leal P,
Buchegger K, Sandoval A, Bizama C, Araya JC, Peek RM and Roa JC:
The gene expression status of the PI3K/AKT/mTOR pathway in gastric
cancer tissues and cell lines. Pathol Oncol Res. 22:797–805. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yu L, Chen X, Wang L and Chen S: The sweet
trap in tumors: Aerobic glycolysis and potential targets for
therapy. Oncotarget. 7:38908–38926. 2016.PubMed/NCBI
|
|
40
|
Chu B, Wang J, Wang Y and Yang G:
Knockdown of PKM2 induces apoptosis and autophagy in human A549
alveolar adenocarcinoma cells. Mol Med Rep. 12:4358–4363. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sun H, Zhu A, Zhang L, Zhang J, Zhong Z
and Wang F: Knockdown of PKM2 suppresses tumor growth and invasion
in lung adenocarcinoma. Int J Mol Sci. 16:24574–24587. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
He Y, Wang Y, Liu H, Xu X, He S, Tang J,
Huang Y, Miao X, Wu Y, Wang Q and Cheng C: Pyruvate kinase isoform
M2 (PKM2). participates in multiple myeloma cell proliferation,
adhesion and chemoresistance. Leuk Res. 39:1428–1436. 2015.
|
|
43
|
Liu Q, Liang M, Liu T, Vuitton L, Zheng S,
Gao X, Lu M, Li X, Sheyhidin I and Lu X: M2 isoform of pyruvate
kinase (PKM2) is upregulated in Kazakh's ESCC and promotes
proliferation and migration of ESCC cells. Tumour Biol.
37:2665–2672. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mohammad GH, Olde Damink SW, Malago M,
Dhar DK and Pereira SP: Pyruvate kinase M2 and lactate
dehydrogenase a are overexpressed in pancreatic cancer and
correlate with poor outcome. PLoS One. 11:e01516352016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lu W, Cao Y, Zhang Y, Li S, Gao J, Wang
XA, Mu J, Hu YP, Jiang L, Dong P, et al: Up-regulation of PKM2
promote malignancy and related to adverse prognostic risk factor in
human gallbladder cancer. Sci Rep. 6:263512016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Nemazanyy I, Espeillac C, Pende M and
Panasyuk G: Role of PI3K, mTOR and Akt2 signalling in hepatic
tumorigenesis via the control of PKM2 expression. Biochem Soc
Trans. 41:917–922. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yang L, Hou Y, Yuan J, Tang S, Zhang H,
Zhu Q, Du YE, Zhou M, Wen S, Xu L, et al: Twist promotes
reprogramming of glucose metabolism in breast cancer cells through
PI3K/AKT and p53 signaling pathways. Oncotarget. 6:25755–25769.
2015.PubMed/NCBI
|