Introduction
Ovarian cancer is one of the most common
gynecological malignancies with a high mortality rate (1). In 2017, 22,440 new cases and 14,080
deaths of ovarian cancer were estimated to occur in the United
States (2). At the time of diagnosis,
the majority of ovarian cancer patients have already progressed to
advanced disease, due to the lack of early clinical symptoms, which
results in high mortality (3).
However, the molecular mechanisms underlying the tumor metastasis
and chemoresistance in ovarian cancer are not well understood. The
epithelial-mesenchymal transition (EMT) contributes to the tumor
metastasis and chemoresistance (4).
EMT is a biological process accompanied by the loss of cell
adherence junctions and apical-basal polarity, and acquisition of
the mesenchymal phenotype to increase cell motility and
invasiveness (5). EMT marker gene
expression is altered during the EMT process with downregulation of
epithelial markers such as E-cadherin and upregulation of
mesenchymal markers, including Snai1 and 2, N-cadherin, vimentin,
Zeb 1/2, and Twist1/2 (6,7).
ASAP1 (ArfGAP with SH3 Domain, Ankyrin Repeat and PH
Domain 1) protein has N-terminal BAR, PH, ARF GAP, ankyrin repeat,
proline-rich, and C-terminal SH3 domains (8). ASAP1 expression is upregulated in a
variety of cancers as compared to normal tissue, and correlates
with poor survival and prognosis in colorectal, and neck and head
cancer patients (9,10). In breast cancer ASAP1expression
contributes to tumor invasion and metastasis (11). ASAP1 is upregulated in ovarian cancer
and associated with poor patient survival and prognosis (12). However, the function of ASAP1 in
ovarian cancer has not been investigated.
In the present study, we investigated the role of
ASAP1 in ovarian cancer cells using lentiviral vector mediated
overexpression. We demonstrated that ASAP1 was highly expressed in
ovarian cancer and associated with patient poor survival based on
analysis of expression of ASAP1 in clinical specimen from database.
To define the function of ASAP1 in ovarian cancer cells, we
overexpressed ASAP1 in both ovarian cancer SKOV3 and OVCAR3 cells
and found that overexpression of ASAP1 promoted cell proliferation,
migration, invasion and colony formation, and reduces sensitivity
to chemotherapy drug paclitaxel. Moreover, overexpression of ASAP1
promoted EMT, suggesting that ASAP1 plays an oncogenic role in
ovarian cancer cells and potentially contributes to ovarian tumor
metastasis.
Materials and methods
Cell culture
The ovarian cancer cell lines (SKOV3 and OVCAR3)
were obtained from ATCC and cultured in Dulbecco's Modified Eagle
Medium (DMEM) supplemented with 10% FBS (GE Healthcare Life
Sciences, Logan, UT, USA), 100 U/ml penicillin, and 100 µg/ml
streptomycin (Invitrogen; Thermo Fisher Scientific, Inc., Waltham,
MA, USA). HEK293FT cells were purchased from Invitrogen and
cultured in DMEM medium with 10% FBS, 100 U/ml penicillin, 100
µg/ml streptomycin, 1% glutamine, 1% nonessential amino acid, and 1
µg/ml geneticin.
Lentiviral vector production
The lentiviral ASAP1 expression vector was purchased
from Applied Biological Materials Inc., (Richmond, Canada). ASAP1
cDNA was driven by EF1α promoter, while luciferase cDNA driven by
EF1α promoter in the same lentiviral vector served as a control.
Lentivirus was packaged in HEK293FT cells as we published
previously (13). Stable cell lines
were generated by transducing the SKOV3 and OVCAR3 cells with the
lentiviral ASAP1 or control vector (without insert), and selected
with 3 µg/ml puromycin.
Cell migration assay
The transwell migration assays were performed using
a modified chamber system of BDFalcon™; BD Biosciences (San Jose,
CA, USA). These chambers were inserted into a 24-well plate with
3×104 of cells in 300 µl serum-free DMEM added to the
upper chamber. DMEM supplemented with 10% FBS was added into the
lower chamber as a chemoattractant, and cells were incubated for 24
h. The medium and non-migrated cells in the upper chamber were
removed, whereas the migrated cells on the lower side of the
membranes were fixed with methanol and stained with
hematoxylin-eosin (H&E). Pictures were taken at 10×
magnification, and transmigrated cells from at least three
different fields were counted.
Cell invasion assay
SKOV3 and OVCAR3 cells (5×105 per well)
transduced with ASAP1 and control lentiviral vectors were seeded in
serum-free DMEM onto inserts pre-coated with Matrigel (BDBioCoat™;
BD BioSciences). DMEM supplemented with 10% FBS was added to the
bottom chamber, and cells were incubated for 24 h. The upper
chamber containing the non-invasive cells were removed, and the
invasive cells on the lower side of the membranes were fixed with
methanol and stained with H&E. Images were taken at 10×
magnification, and invasive cells from at least three different
fields were counted.
Cell proliferation assay
Cell proliferation at different time points was
examined using MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl
tetrazolium bromide) Assay kit purchased from ATCC (Manassas, VA,
USA) following manufacturer's instructions. Ovarian cancer SKOV3 or
OVCAR3 cells were plated (8×103 cells/well) in 96-well
plates. 10 µl of MTT reagent (10 mg/ml) was added to each well and
incubated for 4 h. The reaction was terminated by adding 100 µl
lysis reagent, and incubated at 22°C in the dark for 2 h. The
absorbance was measured at 570 nm wavelength on a Bio-Rad
Laboratories, Inc., (Hercules, CA, USA) plate reader.
Cell clonogenic assay
SKOV3 and OVCAR3 cells transduced with lentiviral
ASAP1 or control vector were plated into 6-well plates (200
cells/well) in triplicate and then stained with 0.1% Crystal Violet
following culture for two weeks. Cell colonies were counted as
described previously (14).
Soft agar colony formation assay
SKOV3 (1×104) and OVCAR3
(5×104) cells were trypsinized and resuspended in growth
medium containing 0.35% agarose (BD Biosciences), and then plated
onto a solid layer of 0.7% agarose in growth medium in 6-well
plates. Fresh medium (500 µl) was added on alternate days for two
weeks. After 12 h incubation with 20 µl of MTT reagent (10 mg/ml),
stained cell colonies were imaged using a Nikon Ti inverted
microscope (Nikon Corporation, Tokyo, Japan).
Cell apoptosis
Stable SKOV3 and OVCAR3 ovarian cancer cell lines
were established with lentiviral ASAP1 expression and control
vectors. Cell apoptosis was measured using a caspase3/7 activity
assay kit (Promega Corporation, Madison, WI, USA) following
treatment with the chemotherapy drug paclitaxel at the indicated
doses for 24 h. Cell apoptosis was also detected in both SKOV3 and
OVCAR3 cells using western blot by examining cleaved-PARP and
cleaved-caspase3.
Immunofluorescence
To detect ASAP1 expression in ovarian cancer tissue,
sections from three different paraffin embedded human serous
carcinoma specimens were antigen-retrieved and incubated with
blocking buffer (5% normal goat serum, 3% bovine serum albumin, and
0.1% Triton-X 100 in PBS) for 1 h. Primary antibodies to ASAP1
(1:200; Rockland; Atlanta, GA, USA), and PCNA (1:200; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) were incubated with the
sections at 4°C overnight. After three rinsing for 5 min with PBST,
Alexa 594 or 488 conjugated goat anti-rabbit or mouse antibodies
were applied (1:200 dilution; Thermo Fisher Scientific, Inc.) for 1
h at room temperature. Cell nuclei were counterstained with DAPI
(Vector Laboratories, Inc., Burlingame, CA, USA). Images were taken
using a Nikon Ti inverted fluorescence microscope (Nikon
Corporation).
Western blot analysis
Ovarian cancer cells were collected in RIPA buffer
(Thermo Fisher Scientific, Inc.) containing 1% Halt Proteinase
Inhibitor Cocktail (Thermo Fisher Scientific, Inc.). An equal
amount of protein (80 µg/lane) was loaded onto 10% SDS-PAGE gels
and transferred onto nitrocellulose membranes. The membranes were
blocked with 5% non-fat milk for 1 h, and incubated with primary
antibodies against ASAP1 (Rockland; Atlanta, GA, USA), E-cadherin,
N-cadherin, β-catenin, vimentin, Snai2, Cleaved-PARP,
Cleaved-caspase3 (Cell Signaling Technology, Inc., Danvers, MA,
USA) and GAPDH (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
overnight at 4°C. After three rinses with PBST buffer for 5 min,
the membranes were incubated with secondary antibodies at room
temperature for 1 h. The membranes were incubated with
chemiluminescence solution for 2 mins and exposed on X-ray
film.
Statistical analysis
Significant differences were determined in at least
two independent experiments performed in triplicate and presented
as means ± SD using Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
ASAP1 was upregulated in ovarian
carcinoma and associated with poor patient survival and
prognosis
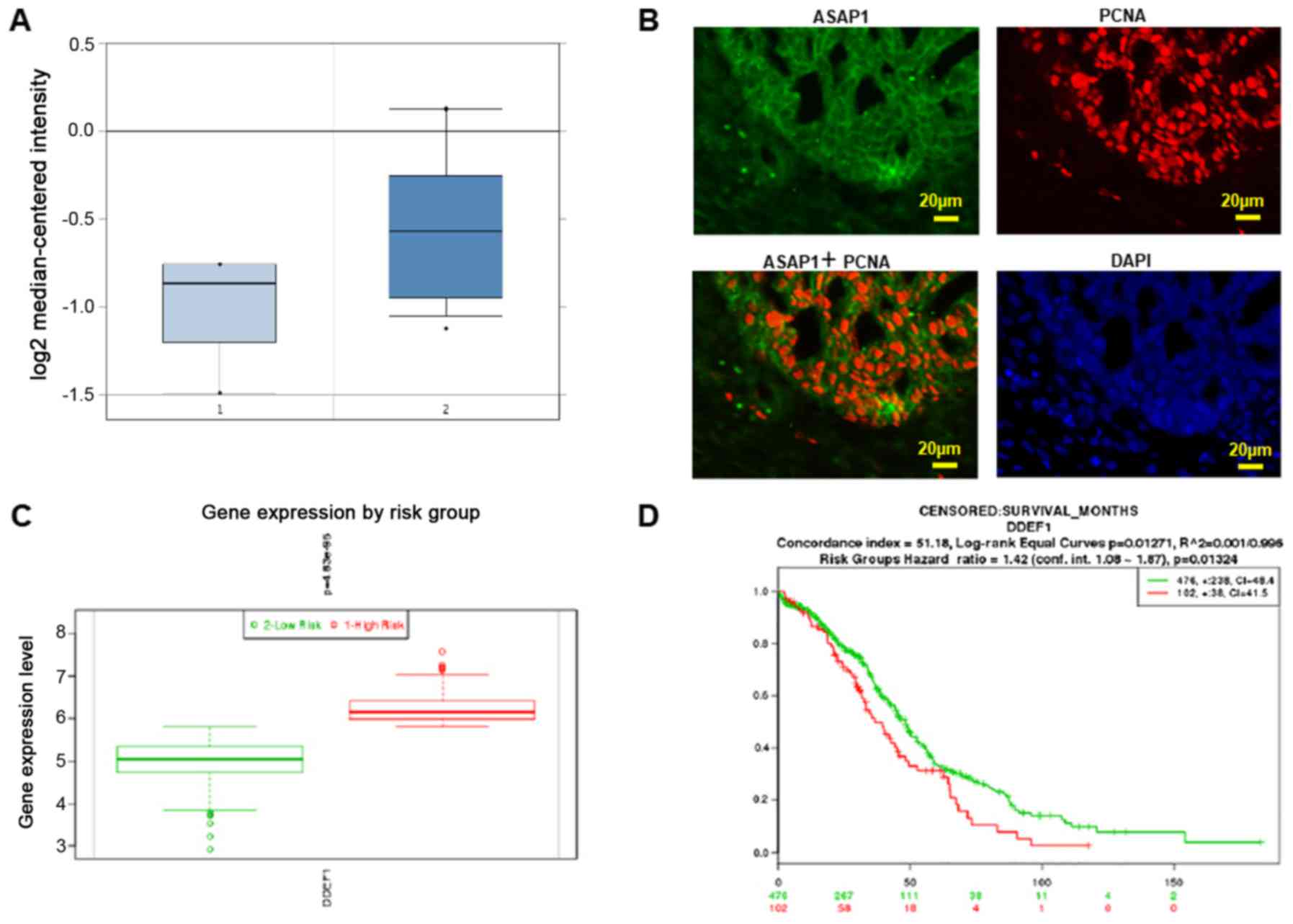
To examine ASAP1 expression in ovarian cancer, we
analyzed TCGA dataset on human ovarian serous carcinoma using the
Oncomine database. ASAP1 (DDEF1) expression was significantly
upregulated in 586 tumor specimens compared to expression in 8
normal tissue specimens (P=4.49e-4) (Fig.
1A). ASAP1 expression was examined by immunofluorescent
staining of sections from three different paraffin embedded ovarian
serous carcinoma specimen and ASAP1 were strongly stained in the
cytoplasm and membrane. The cell proliferation marker PCNA was
stained in the nuclei of tumor cells as shown in Fig. 1B. ASAP1 expression was then analyzed
in 568 tumor and 8 normal ovaries in TCGA database according to
risk groups using SurvExpress program. ASAP1 expression was found
to be significantly higher in the high risk group as compared to
the low risk group (P=4.83e-95) (Fig.
1C). Moreover, ovarian cancer patients in the high risk group
had significantly poorer overall survival compared to the low risk
group (P=0.01324) (Fig. 1D).
Overexpression of ASAP1 promoted EMT
in ovarian cancer cells
Since ASAP1 was highly expressed in ovarian cancer
and associated with poor patient survival, we hypothesized that
ASAP1 may promote tumorigenesis in ovarian cancer cells. To test
this hypothesis, we overexpressed ASAP1 in ovarian cancer SKOV3 and
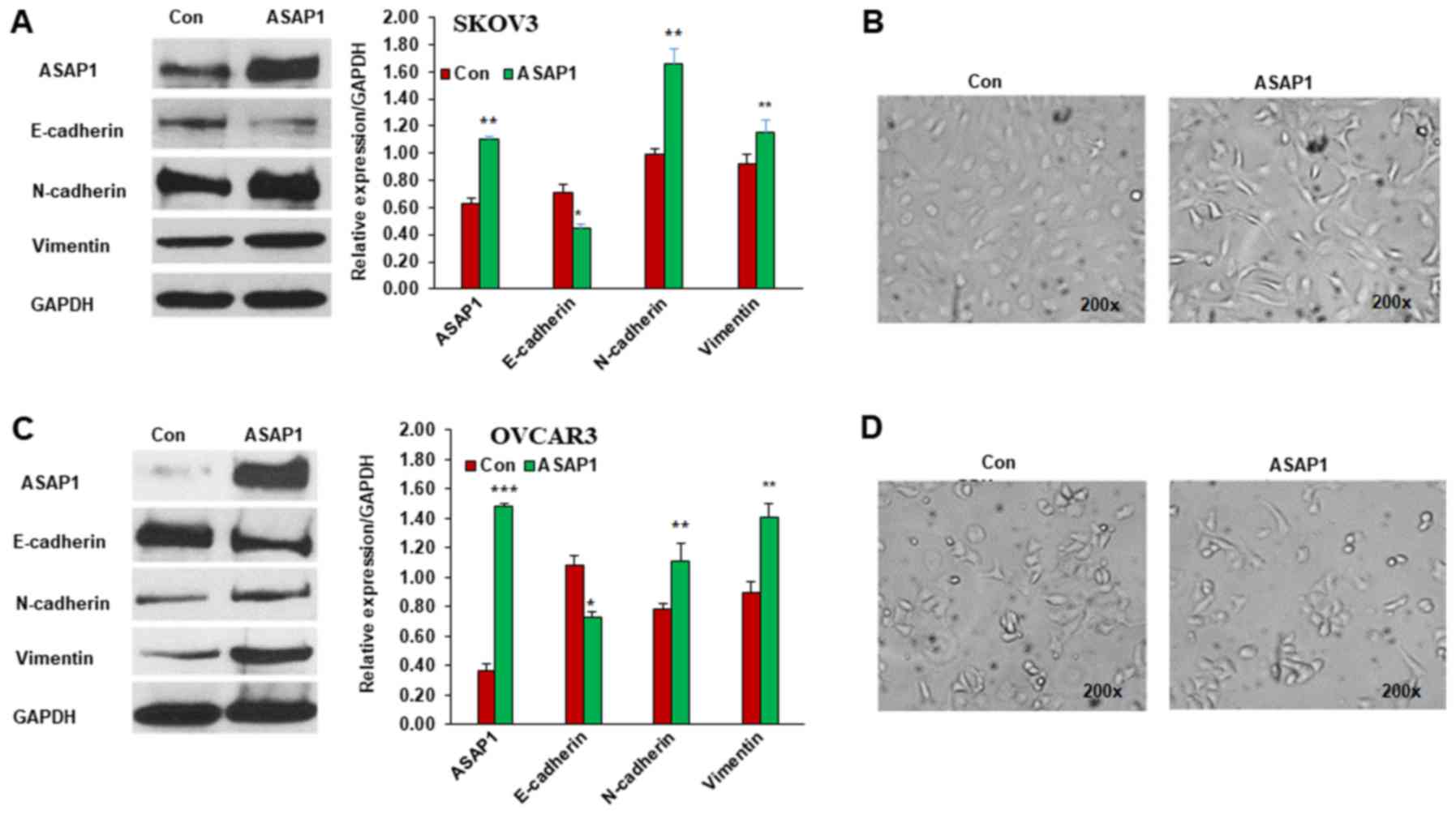
OVCAR3 cells by lentiviral transduction. The expression level of
ASAP1 was increased by approximately two-fold in SKOV3 cells
compared to control as detected by western blotting. Since ASAP1
was found to be associated with tumor metastasis and
chemoresistance in other types of cancer, we examined the effect of
ASAP1 overexpression on expression of EMT markers in ovarian cancer
cells. Overexpression of ASAP1 increased the expression of
mesenchymal cell markers including N-cadherin and Vimentin, while
decreasing epithelia cell marker E-cadherin expression in SKOV3
cells (Fig. 2A). Moreover, SKOV3
cells expressing ASAP1 had a mesenchymal phenotype with a
spindle-like fibroblast morphology, while control cells had a
cobblestone-like appearance of epithelial cells (Fig. 2B). Similarly, overexpression of ASAP1
promoted EMT in OVCAR3 cells compared to empty vector transduced
control cells (Fig. 2C). However,
overexpression of ASAP1 only led to a modest mesenchymal phenotype
in OVCAR3 cells compared to control cells transduced with empty
vector (Fig. 2D). Our results
indicated that overexpression of ASAP1 promoted EMT in both ovarian
cancer SKOV3 and OVCAR3 cells.
Overexpression of ASAP1 promoted cell
migration and invasion in ovarian cancer cells
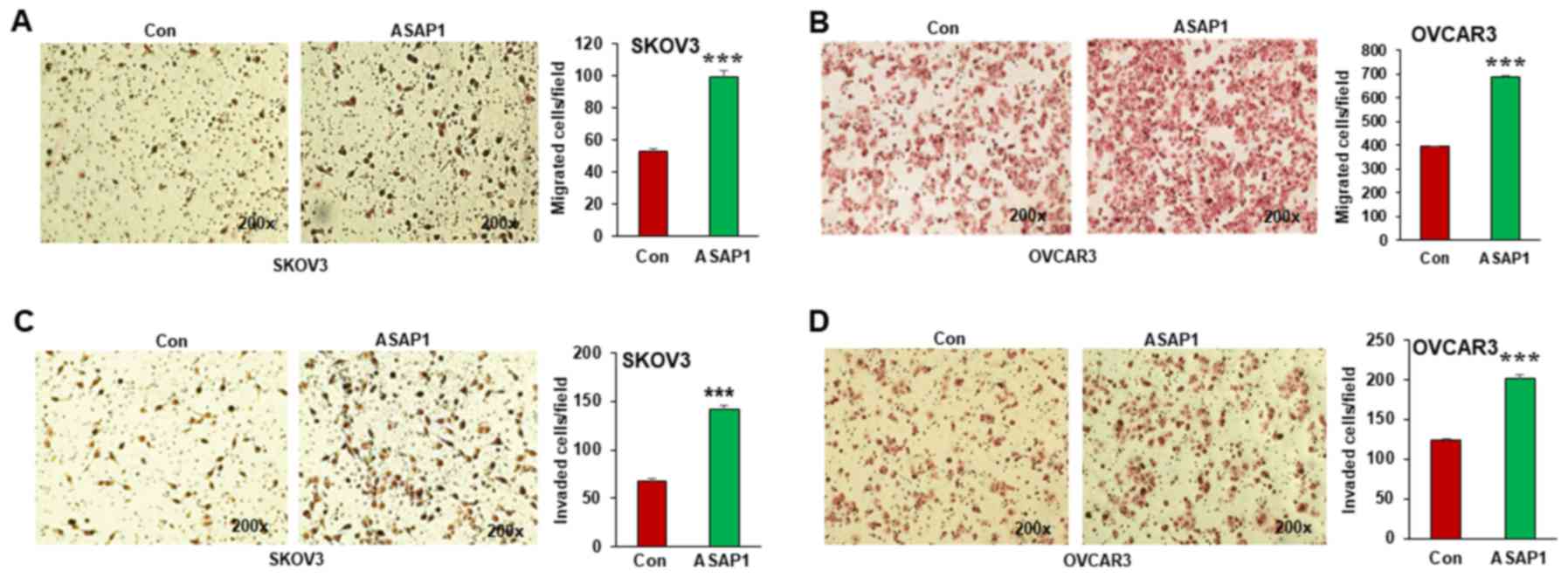
ASAP1 expression promoted EMT in ovarian cancer
cells, suggesting that ASAP1 may contribute to tumor metastasis.
Therefore, we examined the effect of ASAP1 expression on cell
migration and invasion in both SKOV3 and OVCAR3 cells. As shown in
Fig. 3A and B, ASAP1 expression
significantly promoted cell migration in both SKOV3 and OVCAR3
cells on transwell plates. We also examined cell invasion of
ovarian cancer cells using matrigel coated transwell plates.
Overexpression of ASAP1 significantly promoted cell invasion in
both SKOV3 (Fig. 3C) and OVCAR3
(Fig. 3D) cells.
Overexpression of ASAP1 promoted
ovarian cancer cell proliferation and survival
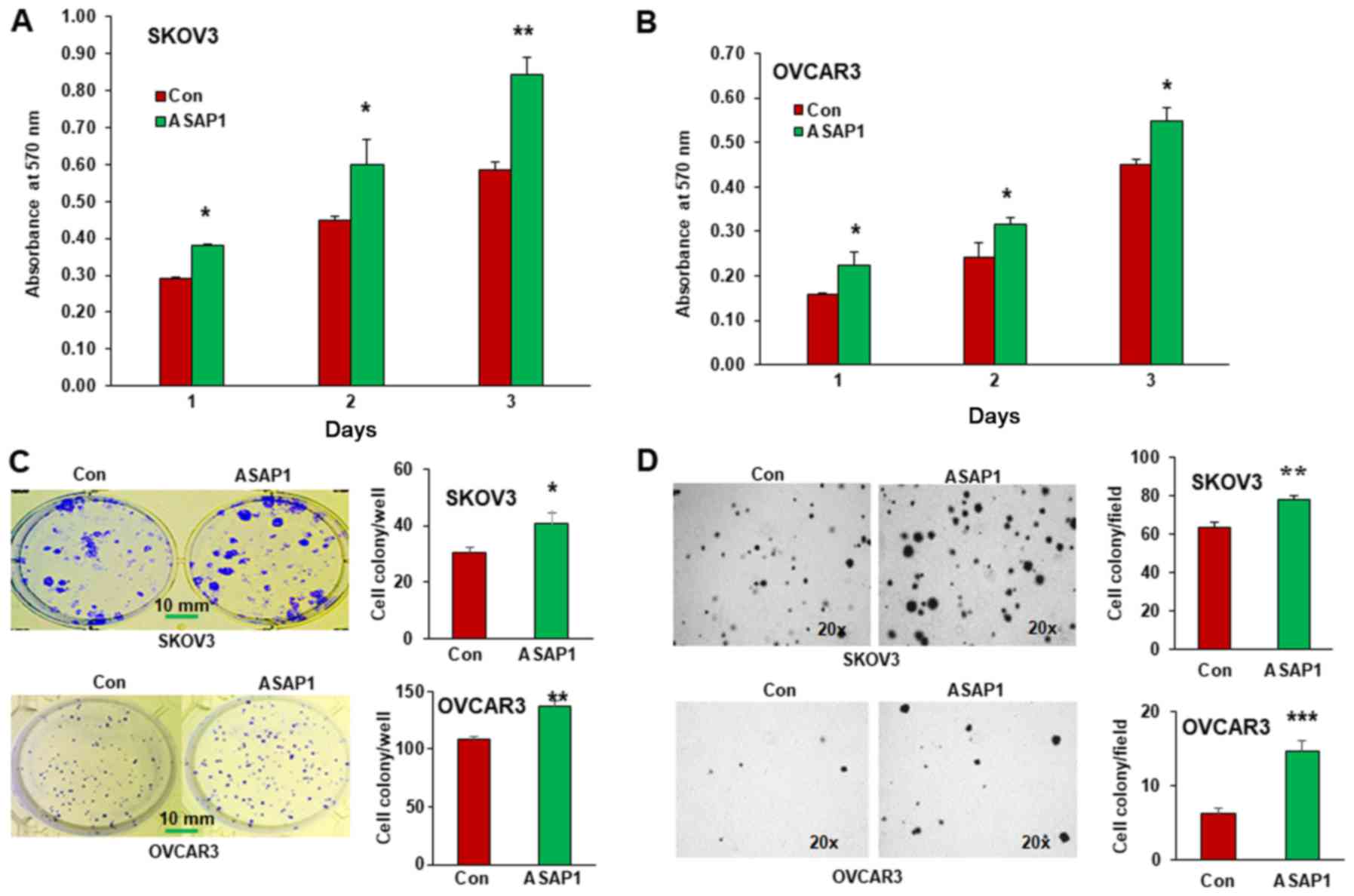
To determine whether ASAP1 plays a role on cell
proliferation, we performed MTT assays on ASAP1 expressing and
control ovarian cancer cells at different time points (1, 2 and 3
days). Overexpressing ASAP1 significantly increased cell
proliferation in both SKOV3 and OVCAR3 cells (Fig. 4A and B). To determine cell survival,
we performed cell clonogenic assays and counted cell colonies in
ASAP1 expressing and control SKOV3 and OVCAR3 cells. Overexpression
of ASAP1 significantly promoted cell survival in both cell lines
(Fig. 4C). In addition, to examine
the role of ASAP1 in tumorigenesis, we examined the
anchorage-independent cell growth by soft agar assays, which is an
in vitro method to detect tumor cell malignancy.
Overexpression of ASAP1 significantly increased the number of
colonies formed in soft agar as compared to control cells (Fig. 4D).
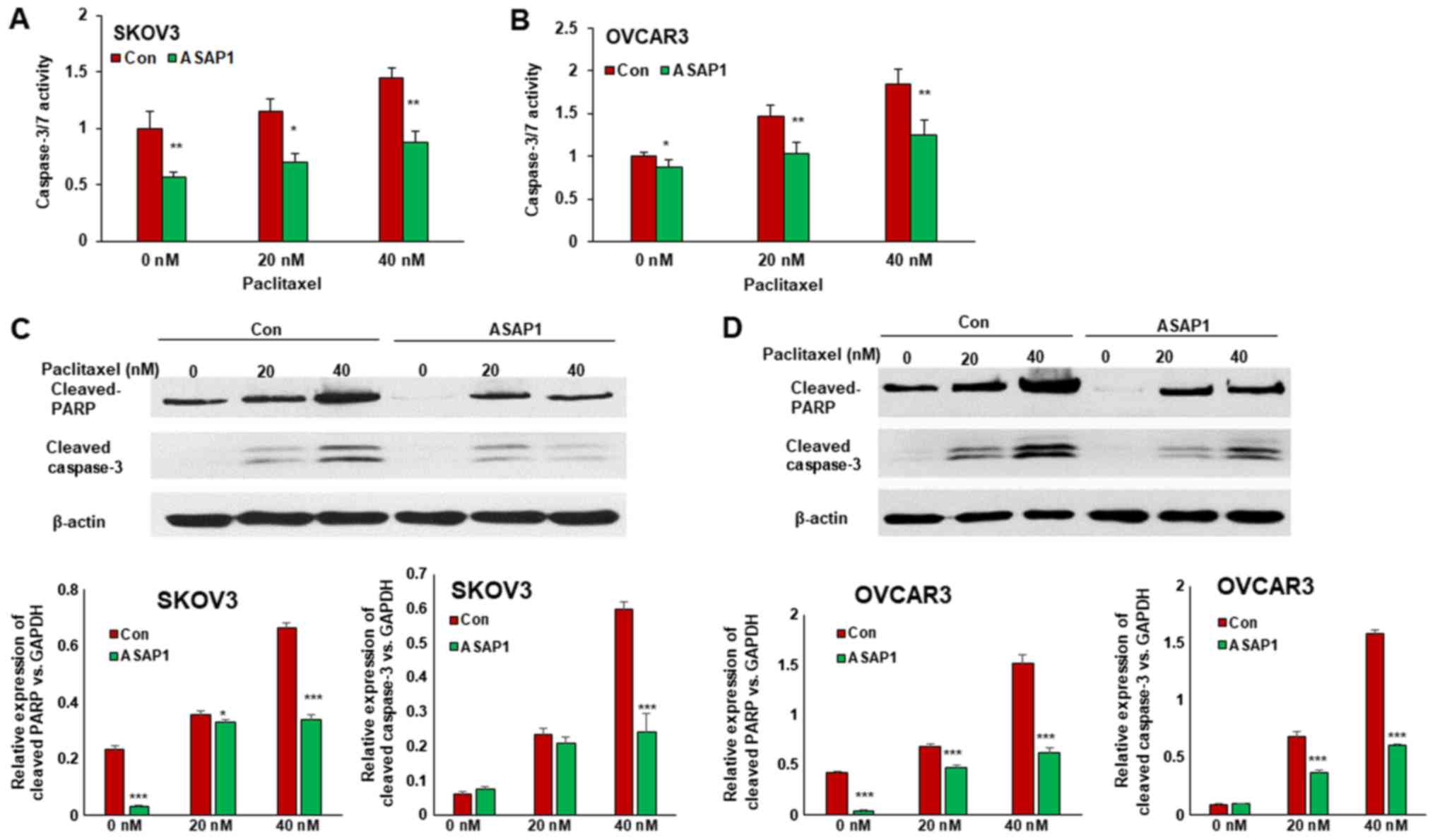
Overexpression of ASAP1 inhibited
apoptosis induced by chemotherapy drug paclitaxel in ovarian cancer
cells
To determine the role of ASAP1 in cell apoptosis, we
treated ovarian cancer cells with different doses of the
chemotherapy drug paclitaxel, and cell apoptosis was examined by
measuring Caspase3/7 activity. Overexpression of ASAP1 led to
1.7-fold decrease of apoptosis in ASAP1 expressing SKOV3 cells
compared to control (Fig. 5A).
Furthermore, ASAP1 expression inhibited cell apoptosis induced by
paclitaxel by 1.6-fold when cells were treated with 20 and 40 nM
paclitaxel, respectively (Fig. 5A).
Overexpression of ASAP1 in OVCAR3 cells led to 1.4-fold decrease in
apoptosis when cells were treated with 20 or 40 nM paclitaxel
(Fig. 5B). Cell apoptosis was also
examined by detecting cleaved-PARP and cleaved-caspase3 using
western blot, ASAP1 expression decreased paclitaxel induced cell
apoptosis in both SKOV3 (Fig. 5C) and
OVCAR3 cells (Fig. 5D). These data
indicate that ASAP1 expression in ovarian cancer cells promoted
chemoresistance.
Discussion
In the present study, for the first time we showed
that endogenous ASAP1 was highly expressed in ovarian cancer
compared to normal ovaries, and ASAP1 expression was associated
with overall poor patient survival by analyzing TCGA database,
suggesting that ASAP1 is a potential biomarker for diagnosis and
prognosis of ovarian cancer patients. Our finding was consistent
with previous studies that ASAP1 was also highly expressed in
several other cancers including melanoma (15), colorectal cancer (9), head and neck carcinoma (10), and breast cancer (11). Interestingly, ASAP1 expression was
shown to correlate with the poor prognosis of ovarian cancer
patients (12). ASAP1 was
endogenously expressed in both SKOV3 and OVCAR3, although we are
not able to compare it with normal human ovarian epithelial cells.
However, ASAP1 was not detectable in normal mammary epithelial
cells (11). Our studies indicated
that ASAP1 expression was associated with poor survival and
prognosis in ovarian cancer patients. However, further studies are
required to understand how ASAP1 expression is correlated with the
different types of ovarian cancer, as well as disease grade and
stage.
Although we used a more invasive SKOV3 and less
invasive OVCAR3 cells for our studies, morphologically, SKOV3
showed a mesenchymal while OVCAR3 displayed an epithelial
phenotype. We used gain of function approach to define the role of
ASAP1 in ovarian cancer cells. For the first time we have shown
that ASAP1 functions as an oncogene by promoting EMT in ovarian
cancer cells. Overexpression of ASAP1 significantly altered EMT
marker gene expression in both SKOV3 and OVCAR3 cells compared to
control cells transduced with empty vector (Fig. 2A and C). In OVCAR3 cells, we did not
observe a robust EMT phenotype switch following ASAP1 expression as
we observed in SKOV3 cells. This was not surprise since OVCAR3
cells are well-differentiated epithelial cells and non-invasive
(16). Overexpression of ASAP1 in
OVCAR3 cells was not sufficient to fully convert epithelial to
mesenchymal phenotypes, but only a partial phenotypic switch. In
this case, it may require additional factors for a full phenotypic
switch.
Extensive studies demonstrated that EMT was
associated with tumor metastasis and chemoresistance in various
human cancers. We showed that overexpression of ASAP1 promoted EMT
in both SKOV3 and OVCAR3 cells, indicating that ASAP1 may
contribute to tumor metastasis and chemoresistance in ovarian
cancer cells. Although it is not clear how ASAP1 contributes to EMT
in ovarian cancer cells, it was previously found that GEP100
activated an Arf6 pathway with ASAP1 serving as an effector via
receptor tyrosine kinases (RTKs), thus activated beta1 integrins
and disrupted E-cadherin-based adhesion and promoted EMT in breast
cancer (17). Arf6-ASAP1-EPB41L5 axis
is another pathway to contribute to ASAP1-mediated EMT (18). ASAP1 was also shown to bind the SH3
domain of several members of the src family through its
proline-rich domain and contributed to tumor metastasis in
colorectal cancer (9). In addition,
the non-coding endogenous small miRNAs also play a role in
regulating EMT by directly targeting EMT marker genes or through
indirect regulation. We previously showed that miR-203 inhibited
EMT by targeting mesenchymal marker Snail in ovarian cancer cells
(19). Interestingly, ASAP1 was
identified as a target gene of miR-203 and overexpression of
miR-203 led to inhibition of EMT in prostate cancer cells (20). Although we did not test whether
miR-203 directly regulated ASAP1 and thereby inhibited EMT in
ovarian cancer cells, it was highly possible that ASAP1 mediated
EMT was also regulated by miR-203 in ovarian cancer cells. ASAP1
expression promoted EMT in ovarian cancer cells, which may lead to
tumor metastasis and chemoresistance. We showed that overexpression
of ASAP1 promoted cell proliferation, migration and invasion in
both SKOV3 and OVCAR3 cells. Although the overexpression level of
ASAP1 in OVCAR3 cells is higher than that in SKOV3 cells, the
migration and invasion were similar in both cell lines (Fig. 3), which may be caused by more invasive
SKOV3 compared with less invasive OVCAR3 cells (16).
Therefore, ASAP1 may promote ovarian tumor
metastasis. ASAP1 was shown to promote tumor metastasis in breast
cancer (11), colorectal (9) and prostate cancer (21). The role of ASAP1 on ovarian tumor
metastasis is presently being tested in our lab using an orthotopic
ovarian cancer mouse model. Moreover, ASAP1 expression can be
inhibited with a small inhibitor UCS15A by disrupting ASAP1 binding
with cortactin, thus blocking breast tumor metastasis (11). ASAP1 expression inhibited apoptosis in
both SKOV3 and OVCAR3 cells induced by the chemotherapy drug
paclitaxel as we showed by two different assays including
luciferase reporter gene assay (Fig. 5A
and B) and western blot (Fig. 5C and
D). The luciferase reporter gene assay for apoptosis was more
sensitive than western blot (22),
thus we observed the significant difference of apoptosis at 20 nM
by measuring caspase3/7 activity. Thus, inhibition of ASAP1
expression using small molecule inhibitor like UCS15A may provide a
novel approach in treating ovarian cancer patients by inhibiting
cell invasion and enhancing the efficacy of chemotherapy drugs.
In summary, our studies revealed that the high ASAP1
expression was associated with poor ovarian patient survival and
prognosis. Overexpression of ASAP1 promoted ovarian cancer cell
proliferation, migration and invasion, and enhanced chemoresistance
by promoting EMT. This study is a part of our long-term work to
define the molecular mechanisms underlying ASAP1function in ovarian
cancer. We have already generated ASAP1 knockout cell lines using
CRISPR/Cas9 and they will be used to address its function in
ovarian tumor metastasis as our further studies. Targeting ASAP1
using small molecule inhibitors may inhibit tumor metastasis and
enhance the efficacy of chemotherapy drugs by inhibiting EMT, thus
provide a novel target for ovarian cancer therapy.
Acknowledgements
This study was supported by a West Cancer Center
grant to J. Yue.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kosary CL: FIGO stage, histology,
histologic grade, age and race as prognostic factors in determining
survival for cancers of the female gynecological system: An
analysis of 1973-87 SEER cases of cancers of the endometrium,
cervix, ovary, vulva, and vagina. Semin Surg Oncol. 10:31–46. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yeung KT and Yang J:
Epithelial-mesenchymal transition in tumor metastasis. Mol Oncol.
11:28–39. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nieto MA: Epithelial plasticity: A common
theme in embryonic and cancer cells. Science. 342:12348502013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sánchez-Tilló E, Fanlo L, Siles L,
Montes-Moreno S, Moros A, Chiva-Blanch G, Estruch R, Martinez A,
Colomer D, Győrffy B, et al: The EMT activator ZEB1 promotes tumor
growth and determines differential response to chemotherapy in
mantle cell lymphoma. Cell Death Differ. 21:247–257. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pinho AV, Rooman I and Real FX:
p53-dependent regulation of growth, epithelial-mesenchymal
transition and stemness in normal pancreatic epithelial cells. Cell
Cycle. 10:1312–1321. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kahn RA and Lambright DG: A PH domain with
dual phospholipid binding sites regulates the ARF GAP, ASAP1.
Structure. 23:1971–1973. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Müller T, Stein U, Poletti A, Garzia L,
Rothley M, Plaumann D, Thiele W, Bauer M, Galasso A, Schlag P, et
al: ASAP1 promotes tumor cell motility and invasiveness, stimulates
metastasis formation in vivo and correlates with poor survival in
colorectal cancer patients. Oncogene. 29:2393–2403. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sato H, Hatanaka KC, Hatanaka Y,
Hatakeyama H, Hashimoto A, Matsuno Y, Fukuda S and Sabe H: High
level expression of AMAP1 protein correlates with poor prognosis
and survival after surgery of head and neck squamous cell carcinoma
patients. Cell Commun Signal. 12:172014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hashimoto S, Hirose M, Hashimoto A,
Morishige M, Yamada A, Hosaka H, Akagi K, Ogawa E, Oneyama C,
Agatsuma T, et al: Targeting AMAP1 and cortactin binding bearing an
atypical src homology 3/proline interface for prevention of breast
cancer invasion and metastasis. Proc Natl Acad Sci USA. 103:pp.
7036–7041. 2006; View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hou T, Yang C, Tong C, Zhang H, Xiao J and
Li J: Overexpression of ASAP1 is associated with poor prognosis in
epithelial ovarian cancer. Int J Clin Exp Pathol. 7:280–287.
2013.PubMed/NCBI
|
|
13
|
Yue J, Sheng Y, Ren A and Penmatsa S: A
miR-21 hairpin structure-based gene knockdown vector. Biochem
Biophys Res Commun. 394:667–672. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo X, Zhang R, Wang J and Lu R: Antiviral
RNA silencing initiated in the absence of RDE-4, a double-stranded
RNA binding protein, in Caenorhabditis elegans. J Virol.
87:10721–10729. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ehlers JP, Worley L, Onken MD and Harbour
JW: DDEF1 is located in an amplified region of chromosome 8q and is
overexpressed in uveal melanoma. Clin Cancer Res. 11:3609–3613.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang L, Moss T, Mangala LS, Marini J, Zhao
H, Wahlig S, Armaiz-Pena G, Jiang D, Achreja A, Win J, et al:
Metabolic shifts toward glutamine regulate tumor growth, invasion
and bioenergetics in ovarian cancer. Mol Syst Biol. 10:7282014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kinoshita R, Nam JM, Ito YM, Hatanaka KC,
Hashimoto A, Handa H, Otsuka Y, Hashimoto S, Onodera Y, Hosoda M,
et al: Co-overexpression of GEP100 and AMAP1 proteins correlates
with rapid local recurrence after breast conservative therapy. PLoS
One. 8:e767912013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Handa H, Hashimoto A, Hashimoto S and Sabe
H: Arf6 and its ZEB1-EPB41L5 mesenchymal axis are required for both
mesenchymal- and amoeboid-type invasion of cancer cells. Small
GTPases. 1–7. 2016.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao G, Guo Y, Chen Z, Wang Y, Yang C,
Dudas A, Du Z, Liu W, Zou Y, Szabo E, et al: miR-203 functions as a
tumor suppressor by inhibiting epithelial to mesenchymal transition
in ovarian cancer. J Cancer Sci Ther. 7:34–43. 2015.PubMed/NCBI
|
|
20
|
Viticchiè G, Lena AM, Latina A, Formosa A,
Gregersen LH, Lund AH, Bernardini S, Mauriello A, Miano R, Spagnoli
LG, et al: MiR-203 controls proliferation, migration and invasive
potential of prostate cancer cell lines. Cell Cycle. 10:1121–1131.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lin D, Watahiki A, Bayani J, Zhang F, Liu
L, Ling V, Sadar MD, English J, Fazli L, So A, et al: ASAP1, a gene
at 8q24, is associated with prostate cancer metastasis. Cancer Res.
68:4352–4359. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alvero AB, Montagna MK and Mor G:
Correlation of caspase activity and in vitro chemo-response in
epithelial ovarian cancer cell lines. Methods Mol Biol. 414:79–82.
2008.PubMed/NCBI
|