Introduction
Worldwide, breast cancer occurs in the epithelial
cells of the mammary gland and is the most common type of invasive
malignancy in females (1). Every
year, ~450,000 females succumb as a consequence of breast cancer,
which is the second leading cause of cancer-associated mortality in
females following lung cancer (1).
The incidence rate of breast cancer is increasing around the world
since the 1970s and has become a major public health problem
(2). The breast is a non-vital organ
so, theoretically, breast cancer should not be fatal; however, due
to the loose connection between cells, breast cancer cells are
released early from cancer nests via the blood or lymphatic
vessels, leading to distant metastases and life-threatening disease
for patients (3).
In recent years, comprehensive treatment models,
focusing on local and systemic treatments, have become more popular
in breast cancer therapy. Surgical intervention and radiation are
the predominant local treatment options, whereas systemic
treatments are on the basis of drug intervention (4). The classification of the type of cancer
determines the treatment strategy and outcome. For example, hormone
receptor positive breast cancer responds well to an endocrine
therapy (5), whereas human epidermal
growth factor (HER)2-targeting drugs, including trastuzumab
(Herceptin) and pertuzumab (Perjeta), may result in an improved
outcome for HER2+ breast cancer (6). Triple-negative breast cancer type has a
poor prognosis, compared with the other types of breast cancer, due
to the lack of targeted drug treatments (7).
The development of breast cancer cells is controlled
by complex signaling networks. The major signaling pathways
involved in mammary gland involution, signal transducer and
activator of transcription (STAT3), nuclear factor-kappa B (NF-κB),
transforming growth factor beta (TGF-β), and retinoid acid
receptors (RARs)/retinoid X receptors (RXRs), are reviewed as part
of the complex network of signaling pathways that crosstalk in a
contextual-dependent manner. These factors, also involved in breast
cancer development, and are important regulatory nodes for
signaling amplification following weaning (8). A previous study demonstrated that the
application of a single molecule is unlikely to suppress the
cross-talk between cancerous cells (9). Therefore, multi-drug combination
treatments have become the principal treatment strategy. At
present, the most common types of therapeutics used for breast
cancer treatment are chemotherapeutic agents, including
Doxorubicin, Paclitaxel, Docetaxel, Thioridazine, Disulfiram and
Camptothecin. Multi-drug combination treatments also include
hormone blockers and monoclonal antibodies (10). Patient compliance is notably difficult
with chemotherapeutic drugs, due to their severe toxicity for the
human body (11). Therefore,
exploring the development of novel cancer drugs with reduced side
effects is important.
Tetramethylpyrazine (TMP), an effective component of
the traditional Chinese medicine Chuanxiong, is the active
ingredient of Umbelliferae plant root extracts and has primarily
been used in the treatment of various neurovascular, including
cerebral blood deficiency, cerebral thrombosis or cerebral
infarction caused by cerebral embolism, and cardiovascular
diseases, including angina and coronary heart disease (12,13).
Furthermore, TMP has been demonstrated to exhibit beneficial
effects in a number of types of epithelial malignant cancer
including lung (14), ovarian
(15) and hepatocellular cancer
(16). Previous studies have
validated that TMP exhibited the ability to reduce the resistance
of breast cancer cells to chemotherapy (17,18).
However, the detailed function and underlying molecular mechanism
of TMP in breast cancer therapy remain unknown. Therefore, in the
present study, the effect and mechanism of TMP on cell viability,
apoptosis and migration was investigated.
Materials and methods
Cell culture
The breast cancer cell line MDA-MB-231 was purchased
from the Institute of Biochemistry and Cell Biology at the Chinese
Academy of Sciences (Shanghai, China). MDA-MB-231 cells were
maintained in Dulbecco's modified Eagle's medium (DMEM; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 100 µg/ml streptomycin (HyClone; GE Healthcare,
Logan, UT, USA). Cells were cultured at 37°C in a humidified
incubator containing 5% CO2. The culture medium was
changed every 3 days. Only cells in the exponential growth phase
were included in the present study.
Cell viability analysis
A Cell Counting kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was used to analyze the
viability of breast cancer cells following TMP (Dalian Meilun
Biotech Co., Ltd, Dalian, China) treatment for 24, 48 and 72 h at
37°C. A total of 5×103 MDA-MB-231 cells/well were seeded
(5,000) on each film and transferred to 96-well plates. DMEM was
removed after 8 h, and DMEM containing 0, 800, 1,600 and 3,200 µM
TMP was used to treat the cells for 24, 48 and 72 h. Furthermore,
the 0 µM TMP group was used as the control. The conditioned culture
medium was removed prior to CCK-8 examination. Subsequently, 100 µl
DMEM and 10 µl CCK-8 solution were added to each well, followed by
CCK-8 incubation at 37°C for 2.5 h. The optical density at 450 nm
was determined using a microplate reader (BioTek Instruments, Inc.,
Winooski, VT, USA).
Cell apoptosis analysis
A total of 2×105 MDA-MB-231 cells/well
were seeded onto each film and placed in 6-well plates. Cells used
in this experiment were sub-confluent. Cells were collected after
72 h TMP treatment at 37°C. The concentrations of TMP were 0, 800,
1,600 and 3,200 µM, the 0 µM TMP group was used as the control.
Then, cells were treated in accordance with the protocol of the
Vybrant® Apoptosis Assay kit (Thermo Fisher Scientific,
Inc.). In detail, ice-cold PBS was used to wash the cells three
times. Subsequently, cells were centrifuged at 300 × g for 5 min at
room temperature and re-suspended in 1X Annexin-Binding Buffer. The
apoptosis rate was determined by staining with Annexin
V-allophycocyanin and propidium iodide. All cells were analyzed by
FACScan flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
Data were acquired using CellQuest™ software (version 5.1; BD
Biosciences) to reveal the impact of TMP on cell apoptosis.
Cell migration and invasion
analysis
A total of 2×105 MDA-MB-231 cells/well
were seeded into the upper chamber of an 8.0-µm pore Transwell
apparatus (EMD Millipore, Billerica, MA, USA) and maintained in
DMEM containing 0.2% bovine serum albumin (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany). Conditioned DMEM with different
concentrations (0, 800, 1,600 and 3,200 µM) of TMP was added to the
lower chamber. Furthermore, the 0 µM TMP group was used as the
control. After 1 h incubation at 37°C in an atmosphere containing
5% CO2, the upper chamber was washed with PBS and cells
on the top surface of the insert were removed with a cotton swab.
The invasion assay procedure was similar to that of the cell
migration assay, except that the Transwell membrane was coated with
Matrigel diluted 1:3 (BD Biosciences) and the cells were incubated
for 12 h at 37°C. Cells that migrated to the bottom surface of the
insert were fixed with 4% paraformaldehyde for 30 min at room
temperature and stained with 0.1% crystal violet for 12 h at room
temperature for subsequent observations by using a light microscope
(cat. no. IX71; Olympus Corporation, Tokyo, Japan). Images were
taken randomly at magnification, ×200 and the total cell count was
calculated by counting the number of cells in five
randomly-selected observation fields.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from MDA-MB-231 cells using
the AxyPrep™ Multisource Total RNA Miniprep kit (Axygen Scientific,
Inc., Union City, CA, USA) according to the manufacturer's protocol
in an environment of 4°C. cDNA was synthesized using the
PrimeScript™ RT reagent kit (Takara Bio, Inc., Otsu, Japan). qPCR
was performed using an ABI 7500 Sequencing Detection System and
SYBR® Premix Ex Taq™ (Takara Bio, Inc.). Cycling
conditions were as follows: 40 cycles of 95°C for 5 sec; and 60°C
for 34 sec. The comparative 2−∆∆Cq method (19) was used to calculate the relative
expression level of each target gene with β-actin as the control
gene. All primers used to amplify target genes are listed in
Table I.
 | Table I.Sequences of the primers used in the
quantitative polymerase chain reaction. |
Table I.
Sequences of the primers used in the
quantitative polymerase chain reaction.
| Gene | Sequence (5′-3′) |
|---|
| Akt1 |
|
|
Forward |
ATGAGCGACGTGGCTATTGTGAAG |
|
Reverse |
GAGGCCGTCAGCCACAGTCTGGATG |
| Akt2 |
|
|
Forward |
ATGAATGAGGTGTCTGTCATCAAAGAA |
|
Reverse |
GGCTGCTTGAGGCTGTTGGCGACC |
| Akt3 |
|
|
Forward |
CAGTCTGTCTGCTACAGCCTGGATA |
|
Reverse |
ATGAGCGATGTTACCATTGT |
| β-actin |
|
|
Forward |
CCAACCGCGAGAAGATGA |
|
Reverse |
CCAGAGGCGTACAGGGATAG |
Western blot analysis
A total of 2×105 MDA-MB-231 cells/well
were seeded onto each film and placed in 6-well plates. After 48 h
incubation with 0, 800, 1,600 and 3,200 µM TMP, cells were washed
with PBS, detached from the well using 0.25% trypsin and
centrifuged at 1,000 × g for 5 min at room temperature.
Furthermore, the 0 µM TMP group was used as the control.
Cytoplasmic proteins were extracted using NE-PER®
Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Protein concentration was detected in accordance with the
instructions of the BCA protein assay kit (Thermo Fisher
Scientific, Inc.). SDS-PAGE (10% gel) was used to separate
total-protein kinase B (t-Akt; cat. no. 4685), phosphorylated-Akt
(p-Akt; cat. no. 4060), total-caspase-3 (t-casp3; cat. no. 9662)
and β-actin (dilution of all antibodies, 1:1,000; cat. no. 4970;
Cell Signaling Technology, Inc., Danvers, MA, USA), and SDS-PAGE
(15% gel) was used to separate cleaved-caspase-3 (cleaved-casp3;
dilution 1:1,000; cat. no. 9662; Cell Signaling Technology, Inc.).
Cell homogenates containing equal amounts of protein (30 µg) were
subjected to SDS-PAGE and transferred to 0.22 µm polyvinylidene
difluoride membranes, which were subsequently blocked with 5% fat
free milk at room temperature for 1 h. All primary antibodies were
purchased from Cell Signaling Technology, Inc. and the membranes
were incubated with these antibodies overnight at 4°C. The
following day, the membranes were washed three times with
Tris-buffered saline containing Tween-20 (TBST) and the secondary
horseradish peroxidase-conjugated anti-rabbit immunoglobulin G
antibody (dilution, 1:5,000; cat. no. 7074; Cell Signaling
Technology, Inc.) was applied to the membranes for 1 h at room
temperature. Following three washes in TBST, the membranes were
incubated in enhanced chemiluminescence (ECL) solution according to
the protocol of the ECL detection kit (GE Healthcare) at room
temperature. Positive immunoreactive bands were quantified and
normalized to β-actin.
Statistical analysis
Each sample was analyzed in triplicate and the
experiments were repeated three times. Data from all experiments
are expressed as the mean ± standard error of the mean. The
differences between experimental groups and controls were assessed
using Student's t-test or one-way analysis of variance with post
hoc differences via the Student-Newman-Keuls test. P<0.05 was
considered to indicate a statistically significant difference.
Results
TMP inhibits the viability of
MDA-MB-231 cells
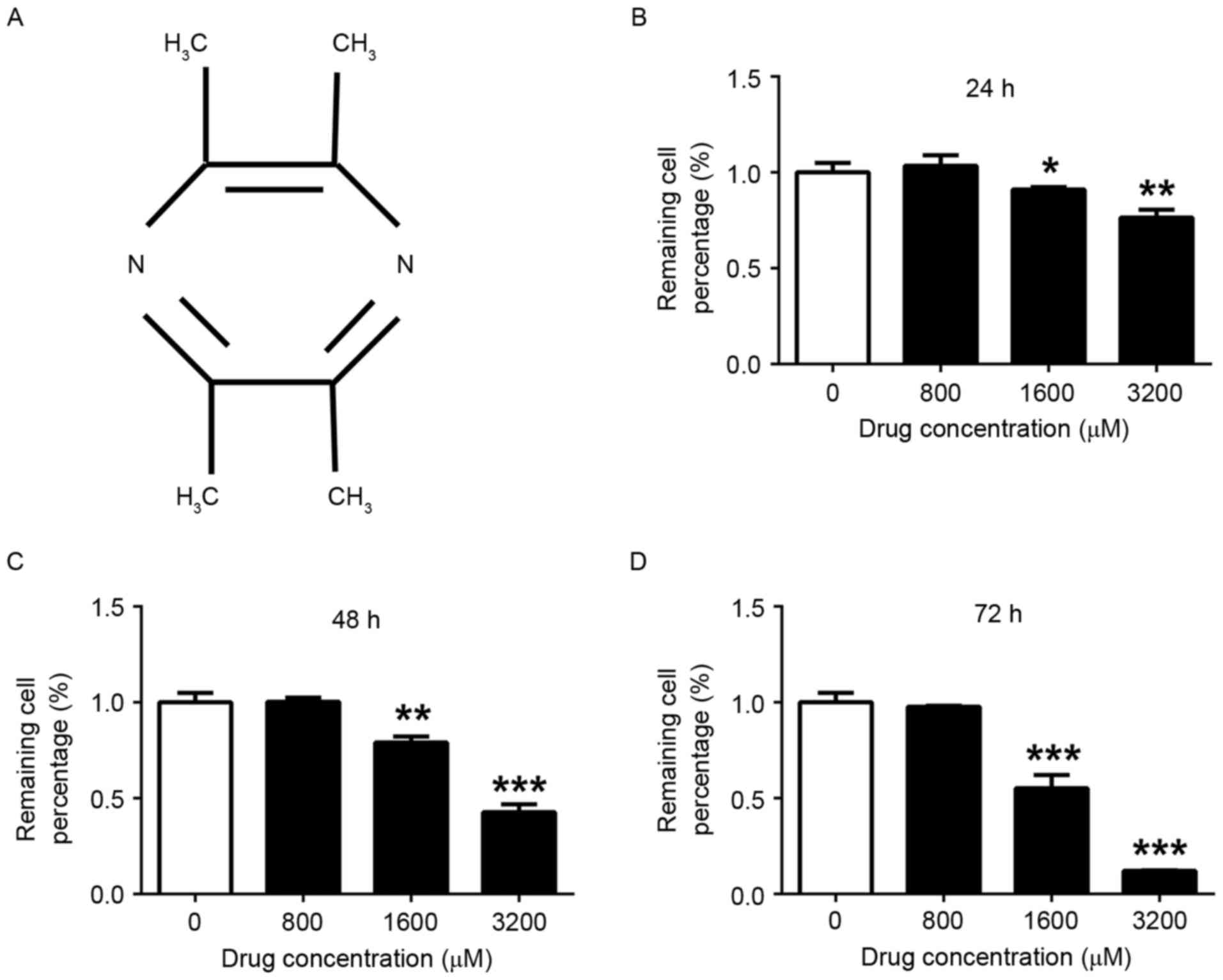
The chemical structure of TMP was depicted in
Fig. 1A. Following treatment with TMP
(0, 800, 1,600 and 3,200 µM) for 24, 48 and 72 h, the viability of
MDA-MB-231 cells was determined using the CCK8 assay. Fig. 1B-D demonstrates that TMP at 1,600 and
3,200 µM significantly inhibited the viability of MDA-MB-231 cells
after 24 (1,600 µM group, P<0.05; 3,200 µM group, P<0.01), 48
(1,600 µM group, P<0.01; 3,200 µM group, P<0.001) and 72 h
(1,600 µM group, P<0.001; 3,200 µM group, P<0.001), compared
with the viability of control cells. Furthermore, the suppression
of viability occurred in a dose-depended manner. However, there was
no significant effect on cell viability when the concentration of
TMP was <800 µM.
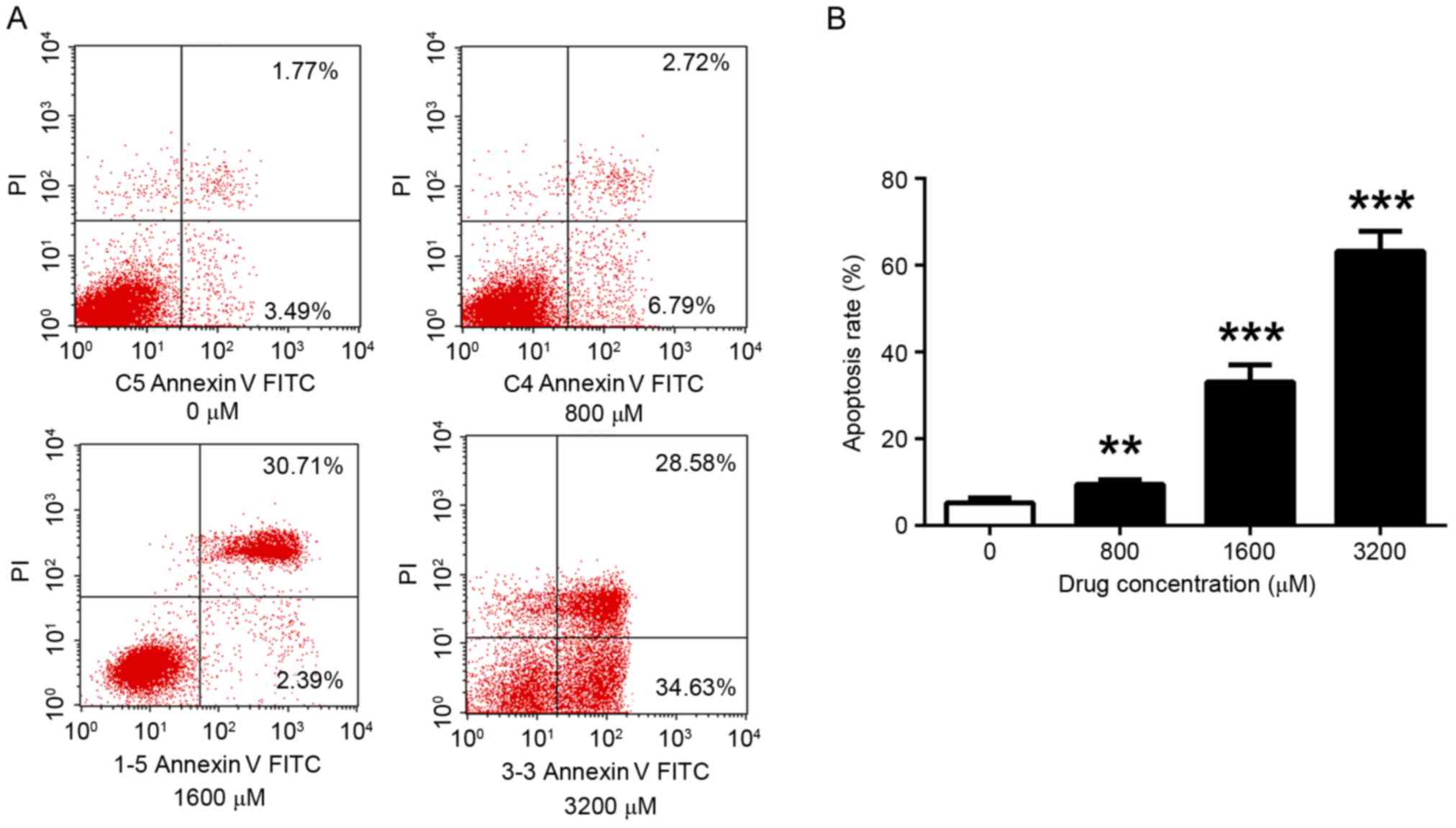
TMP enhances the apoptosis of
MDA-MB-231 cells
The effect of TMP on the apoptosis of MDA-MB-231
cells was examined using flow cytometry. Consistent with effect
observed in the viability assay, apoptosis was altered in a
dose-dependent manner. The level of apoptosis significantly
increased with increasing TMP concentrations. After 72 h
incubation, the results indicated that 800, 1,600 and 3,200 µM TMP
significantly increased the apoptosis rate (9.51, 33.10 and 63.21%,
respectively), compared with the control (5.26%; P<0.01,
P<0.001 and P<0.001, respectively) (Fig. 2).
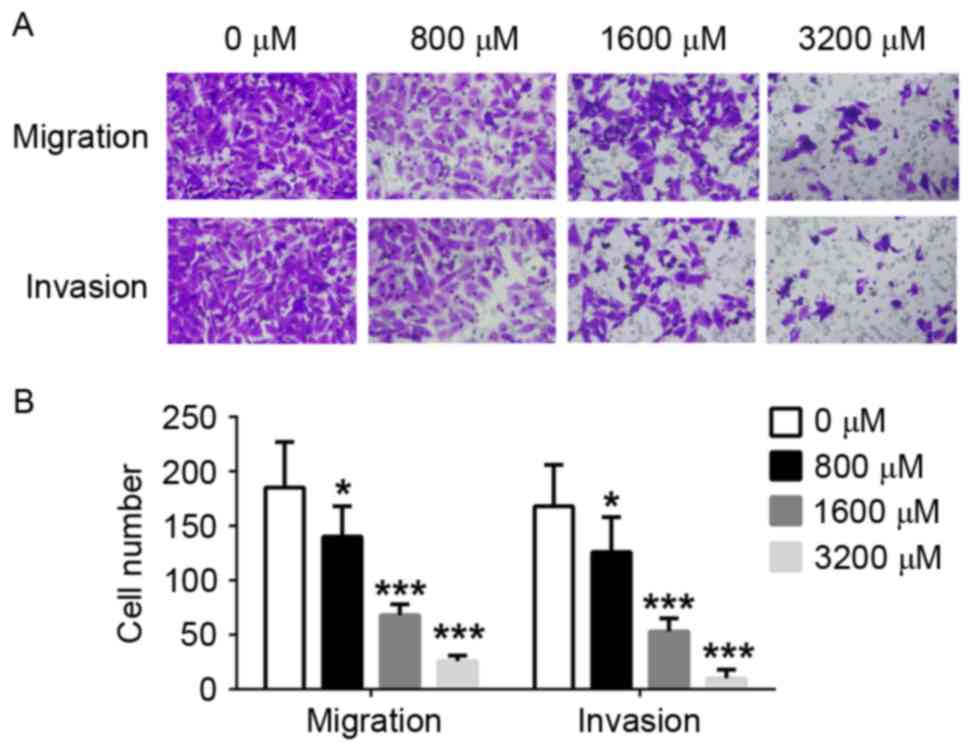
TMP impairs the migration and invasion
of MDA-MB-231 cells
Migration and invasion of MDA-MB-231 cells was
significantly inhibited by 800, 1,600 and 3,200 µM TMP, compared
with the control (P<0.05, P<0.001 and P<0.001,
respectively). The inhibition efficiency was positively associated
with drug concentration. Furthermore, the migratory and invasive
capabilities of the cells was decreased by >50%, compared with
the control, when TMP concentration >,1600 µM (Fig. 3). The results of the present study
revealed that TMP may prevent the migration and invasion of breast
cancer cells.
TMP may regulate the viability,
migration, invasion and apoptosis of breast cancer cells by
decreasing the expression or activity of Akt and caspase-3
To additionally explore the results of the present
study, the alterations in the expression and activity of Akt and
caspase-3 were investigated. The results demonstrated that 1,600
and 3,200 µM TMP significantly inhibited the gene expression of
Akt1 (P<0.01 and P<0.001, respectively), Akt2 (P<0.01 and
P<0.001, respectively) and Akt3 (P<0.05 and P<0.01,
respectively; Fig. 4A), compared with
the control. Furthermore, TMP downregulated the activity of Akt and
caspase-3, the relative expression of p-Akt to t-Akt significantly
decreased (1,600 µM group: P<0.01; 3,200 µM group: P<0.01),
and the relative expression of cleaved-casp3 to t-casp3
significantly increased (800 µM group: P<0.05; 1,600 µM group:
P<0.001; 3,200 µM group: P<0.001) (Fig. 4B and C). These results indicate that
Akt and caspase-3 may serve important roles in cell viability,
migration, invasion, and apoptosis.
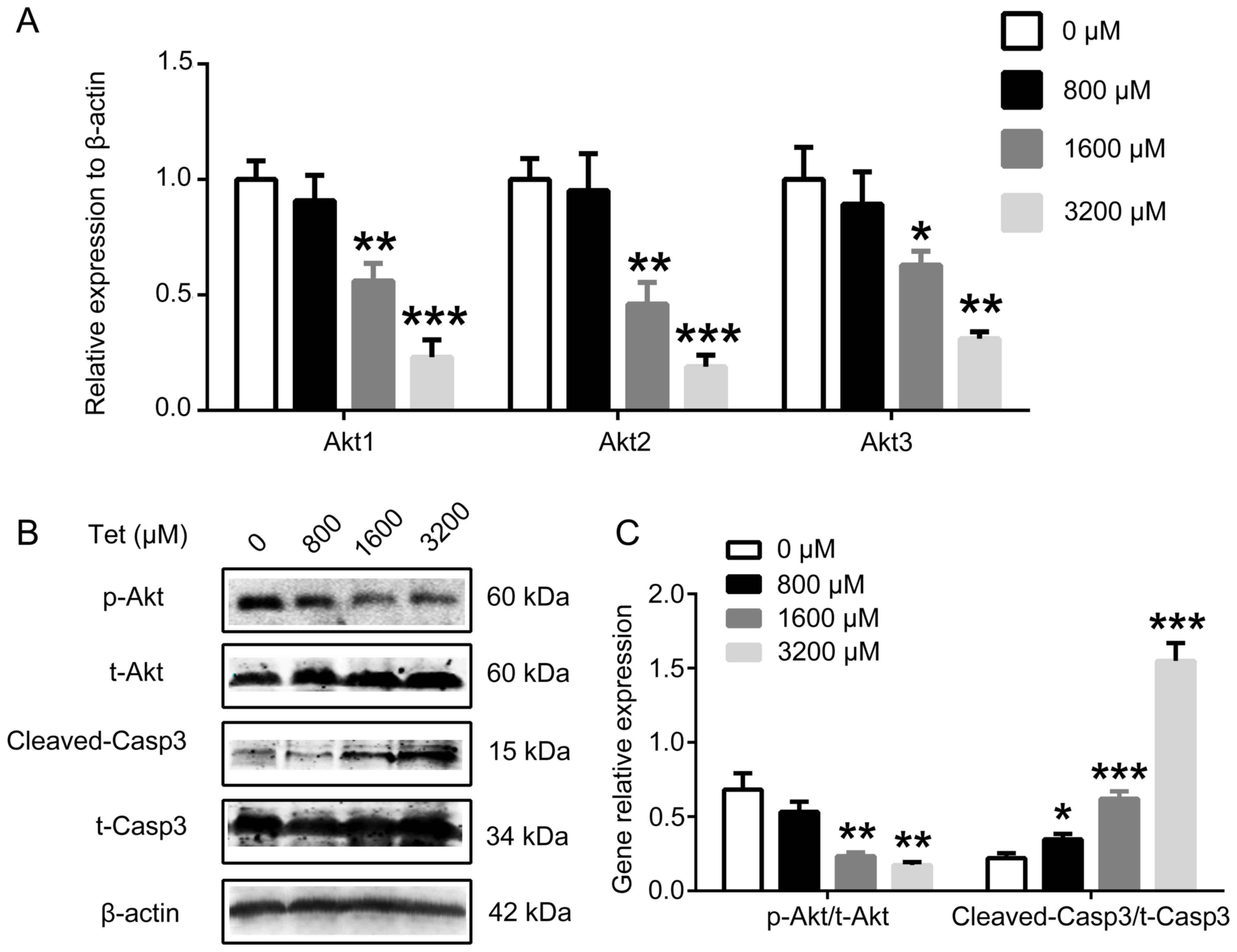 | Figure 4.TMP decreases the gene expression and
protein activity of the Akt signaling pathway and increases
caspase-3 activity. (A) The reverse transcription-quantitative
polymerase chain reaction revealed that TMP significantly
downregulated the gene expression of Akt1, Akt2 and Akt3 at 1,600
and 3,200 µM. (B) Western blot analysis indicated that TMP
decreased the activity of the Akt signaling pathway and increased
the activity of caspase-3. β-actin was the loading control. (C)
Quantification of western blot analysis results which validated
that 1,600 and 3,200 µM significantly decreased the relative
expression of p-Akt to t-Akt and increased the relative expression
of cleaved-casp3 to t-casp3. *P<0.05, **P<0.01, ***P<0.001
vs. control. TMP, tetramethylpyrazine; Akt, protein kinase B; p-,
phosphorylated; t-, total; casp3, caspase-3. |
Discussion
The majority of types of malignant tumor, including
breast cancer, are characterized by continuous cell division and
viability, suppression of the initiation of apoptosis, the ability
to metastasize, and potential recurrence. It is difficult to remove
all cancer cells using surgical techniques due to their ability to
spread to other tissues via the blood stream or lymphatic system.
Therefore, drug interventions to clear cancer cells from the blood
or lymphatic systems are important. Due to the occurrence of severe
side effects and the possibility of drug resistance against common
chemotherapeutic therapies, there is a requirement to identify less
toxic and more efficacious treatment alternatives. Consequently,
natural alternative products have received growing attention.
TMP was extracted, isolated and purified from the
traditional Chinese medicine Chuanxiong Hort. Although the content
of TMP in Chuanxiong Hort is abundant, the extraction process is
time- and energy-consuming (12).
Therefore, the majority of TMP is artificially synthesized
(20). Previous studies have focused
on identifying the underlying molecular mechanisms of TMP activity
due to its well established antitumor effect and its ability to
reverse resistance to chemotherapy treatments, while causing less
adverse reactions (16,17). For example, TMP was able to inhibit
the growth and migration of glioma by regulating calcium influxes
(21) and additionally, TMP was
revealed to decrease the metastases of melanoma by suppressing
vascular endothelial growth factor activity (22). Furthermore, TMP has been identified to
serve a function in the reversal of multidrug resistance in a
number of types of malignant tumor, including reversing the
multi-drug resistance in hepatocellular carcinoma via inhibiting
P-gp, MRP2, MRP3 and MRP5 (16); TMP
could effectively reverse multi-drug resistance of bladder cancer
cells and its mechanisms may be associated with the alteration of
MRP1, GST, BCL-2 and TOPO-II (23);
TMP as a salvage agent for patients with relapsed or refractory
non-Hodgkin's lymphoma may also be associated with its effect on
the expression of P-gp (24).
However, the effect of TMP was different in various types of
tumors. Previous studies have demonstrated that 200 µM TMP
significantly inhibited hepatocellular carcinoma cell proliferation
(25) and that 300 µg/ml TMP
significantly inhibited the viability of acute lymphocytic leukemia
cell lines (26). These results were
similar to the results of the present study where it was observed
that breast cancer cells were inhibited by 1,600 µM TMP. However,
only a limited number of studies have investigated the effect and
mode of action of TMP on breast cancer cells. A recent study
revealed that the combination treatment of the tetramethylpyrazine
piperazine derivative DLJ14 and adriamycin inhibited the
progression of resistant breast cancer (27); however, the function of DLJ14 alone
remains unknown. In fact, DLJ14 is a tetramethylpyrazine piperazine
derivative; therefore, tetramethylpyrazine and DLJ14 are different
drugs (27). It has been demonstrated
that TMP causes apoptotic death and tumor regression in human
breast cancer cells in in vitro, and in vivo models
(28). However, there were no further
studies evaluating the effect of TMP on the migration and invasion
abilities of breast cancer, and the underlying molecular mechanisms
following TMP treatment remain unknown. The results of the present
study identified that TMP may regulate breast cancer cell
migration, invasion and apoptosis by affecting the activity of Akt,
and caspase-3, which is distinct from the results of the
aforementioned studies (18,28). Our study not only confirmed the effect
of TMP on the migration and invasion of breast cancer in addition
to its role of apoptosis, but further found it possible to directly
targets in breast cancer cells.
Excessive viability and apoptosis disorders are the
two primary reasons for the genesis and development of malignant
tumors. The Akt signaling pathway has been identified to serve
functions in a number of types of disease, particularly in
malignancies (29). The Akt signaling
pathway principally regulates the activity of cancer cells
(30,31). Previous studies have indicated that
the Akt signaling pathway is a key factor in the viability and
migration of a number of types of tumor including colorectal
(32), and prostate cancer (33), and glioblastoma (34) and osteosarcoma (35). Additionally, the Akt signaling pathway
is an important regulator of breast cancer (36–38).
Apoptosis disorders are an additional cause for the occurrence of
breast cancer. The initiation of apoptosis is typically triggered
via caspase-3 (39), which is being
used to treat cancer; for example, a number of chemotherapeutic
drugs, including melatonin, doxorubicin and cisplatin, induce
cancer cell apoptosis by upregulating the activity of caspase-3
(40,41). Therefore, novel drugs, which repress
Akt signaling or increase caspase-3 activity, may be effective
tools to improve breast cancer prognosis.
The present study focused on understanding the
effect and mode of action of TMP on the viability, migration,
invasion and apoptosis of breast cancer cells. The results of the
present study demonstrated that TMP is effective against a number
of cancer cell characteristics. In addition, TMP was able to
modulate the activity of the Akt signaling pathway and caspase-3,
up to a concentration of 1,600 µM. However, the viability,
migration, invasion and apoptosis of breast cancer cells were not
significantly inhibited following treatment with 800 µM TMP.
Additionally, the influence of TMP on the activity of Akt and
caspase-3 was more significant than the effect on their expression.
In order to understand the mode of action of TMP on Akt and
caspase-3 signaling pathways, and the effect of TMP in vivo,
additional studies are required.
The results of the present study revealed that TMP
was effective against the viability, migration, invasion and
apoptosis of breast cancer cells. It is hypothesized that the
molecular mechanisms underlying these actions involve the Akt
signaling pathway and caspase-3. The results of the present study
suggest that TMP is a novel drug candidate for the regulation of
breast cancer genesis and development; however, additional in
vivo studies are required.
Acknowledgements
The present study was supported by the Health System
‘Outstanding Young Talent’ Cultivation Plan of Shanghai Jinshan
District (grant no. JSYQ201620), the Science and Technology
Innovation Fund Projects of Shanghai Jinshan District (grant no.
2015-3-24) and the Medical Subject Construction Fund Project of
Shanghai Jinshan District (grant no. JSZK2015B06).
References
|
1
|
Kamangar F, Dores GM and Anderson WF:
Patterns of cancer incidence, mortality and prevalence across five
continents: Defining priorities to reduce cancer disparities in
different geographic regions of the world. J Clin Oncol.
24:2137–2150. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sim X, Ali RA, Wedren S, Goh DL, Tan CS,
Reilly M, Hall P and Chia KS: Ethnic differences in the time trend
of female breast cancer incidence: Singapore, 1968–2002. BMC
Cancer. 6:2612006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gujam FJ, Going JJ, Edwards J, Mohammed ZM
and McMillan DC: The role of lymphatic and blood vessel invasion in
predicting survival and methods of detection in patients with
primary operable breast cancer. Crit Rev Oncol Hematol. 89:231–241.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hack CC, Voiß P, Lange S, Paul AE, Conrad
S, Dobos GJ, Beckmann MW and Kümmel S: Local and systemic therapies
for breast cancer patients: Reducing Short-term symptoms with the
methods of integrative medicine. Geburtshilfe Frauenheilkd.
75:675–682. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nagaraj G, Ellis MJ and Ma CX: The natural
history of hormone receptor-positive breast cancer: Attempting to
decipher an intriguing concept. Oncology (Williston Park).
26:696–700. 2012.PubMed/NCBI
|
|
6
|
Tang Y, Wang Y, Kiani MF and Wang B:
Classification, treatment strategy and associated drug resistance
in breast cancer. Clin Breast Cancer. 16:335–343. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rybárová S, Hodorová I, Hajduková M,
Schmidtová K, Mojzis J, Kajo K, Kviatkovská Z, Plank L, Benický M,
Mirossay A, et al: Expression of MDR proteins in breast cancer and
its correlation with some clinical and pathological parameters.
Neoplasma. 53:128–135. 2006.PubMed/NCBI
|
|
8
|
Zaragozá R, García-Trevijano ER, Lluch A,
Ribas G and Viña JR: Involvement of different networks in mammary
gland involution after the pregnancy/lactation cycle: Implications
in breast cancer. IUBMB Life. 67:227–238. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Espinoza-Fonseca LM: Targeting MDM2 by the
small molecule RITA: Towards the development of new multi-target
drugs against cancer. Theor Biol Med Model. 2:382005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Núñez C, Capelo JL, Igrejas G, Alfonso A,
Botana LM and Lodeiro C: An overview of the effective combination
therapies for the treatment of breast cancer. Biomaterials.
97:34–50. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma P and Mumper RJ: Paclitaxel
nano-delivery systems: A comprehensive review. J Nanomed
Nanotechnol. 4:10001642013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang Y, Fu Q and Zhao W:
Tetramethylpyrazine inhibits osteosarcoma cell proliferation via
downregulation of NF-κB in vitro and in vivo. Mol Med Rep.
8:984–988. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li WM, Liu HT, Li XY, Wu JY, Xu G, Teng
YZ, Ding ST and Yu C: The effect of tetramethylpyrazine on hydrogen
peroxide-induced oxidative damage in human umbilical vein
endothelial cells. Basic Clin Pharmacol Toxicol. 106:45–52.
2010.PubMed/NCBI
|
|
14
|
Zheng CY, Xiao W, Zhu MX, Pan XJ, Yang ZH
and Zhou SY: Inhibition of cyclooxygenase-2 by tetramethylpyrazine
and its effects on A549 cell invasion and metastasis. Int J Oncol.
40:2029–2037. 2012.PubMed/NCBI
|
|
15
|
Yin J, Yu C, Yang Z, He JL, Chen WJ, Liu
HZ, Li WM, Liu HT and Wang YX: Tetramethylpyrazine inhibits
migration of SKOV3 human ovarian carcinoma cells and decreases the
expression of interleukin-8 via the ERK1/2, p38 and AP-1 signaling
pathways. Oncol Rep. 26:671–679. 2011.PubMed/NCBI
|
|
16
|
Wang XB, Wang SS, Zhang QF, Liu M, Li HL,
Liu Y, Wang JN, Zheng F, Guo LY and Xiang JZ: Inhibition of
tetramethylpyrazine on P-gp, MRP2, MRP3 and MRP5 in multidrug
resistant human hepatocellular carcinoma cells. Oncol Rep.
23:211–215. 2010.PubMed/NCBI
|
|
17
|
Zhang Y, Liu X, Zuo T, Liu Y and Zhang JH:
Tetramethylpyrazine reverses multidrug resistance in breast cancer
cells through regulating the expression and function of
P-glycoprotein. Med Oncol. 29:534–538. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang P, Zheng BB, Wang HY, Chen JH, Liu
XY and Guo XL: DLJ14, a novel chemo-sensitization agent, enhances
therapeutic effects of adriamycin against MCF-7/A cells both in
vitro and in vivo. J Pharm Pharmacol. 66:398–407. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang ZY, Wang SL and Lin Y: Progress in
the pharmacokinetics and clinical pharmacodynamics of
tetramethylpyrazine in nervous system. Chin J Clin Pharmacol.
26:535–539. 2010.
|
|
21
|
Fu YS, Lin YY, Chou SC, Tsai TH, Kao LS,
Hsu SY, Cheng FC, Shih YH, Cheng H, Fu YY and Wang JY:
Tetramethylpyrazine inhibits activities of glioma cells and
glutamate neuro-excitotoxicity: Potential therapeutic application
for treatment of gliomas. Neuro Oncol. 10:139–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen L, Lu Y, Wu JM, Xu B, Zhang LJ, Gao
M, Zheng SZ, Wang AY, Zhang CB, Zhang WW and Lei N: Ligustrazine
inhibits B16F10 melanoma metastasis and suppresses angiogenesis
induced by vascular endothelial growth factor. Biochem Biophys Res
Commun. 386:374–379. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang S, Lei T and Zhang M: The reversal
effect and its mechanisms of tetramethylpyrazine on multidrug
resistance in human bladder cancer. PLoS One. 11:e01577592016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang XG and Jiang C: Ligustrazine as a
salvage agent for patients with relapsed or refractory
non-Hodgkin's lymphoma. Chin Med J (Engl). 123:3206–3211.
2010.PubMed/NCBI
|
|
25
|
Cao J, Miao Q, Miao S, Bi L, Zhang S, Yang
Q, Zhou X, Zhang M, Xie Y, Zhang J and Wang S: Tetramethylpyrazine
(TMP) exerts antitumor effects by inducing apoptosis and autophagy
in hepatocellular carcinoma. Int Immunopharmacol. 26:212–220. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang XJ, Xu YH, Yang GC, Chen HX and Zhang
P: Tetramethylpyrazine inhibits the proliferation of acute
lymphocytic leukemia cell lines via decrease in GSK-3β. Oncol Rep.
33:2368–2374. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen J, Wang W, Wang H, Liu X and Guo X:
Combination treatment of ligustrazine piperazine derivate DLJ14 and
adriamycin inhibits progression of resistant breast cancer through
inhibition of the EGFR/PI3K/Akt survival pathway and induction of
apoptosis. Drug Discov Ther. 8:33–41. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pan J, Shang JF, Jiang GQ and Yang ZX:
Ligustrazine induces apoptosis of breast cancer cells in vitro and
in vivo. J Cancer Res Ther. 11:454–458. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mundi PS, Sachdev J, McCourt C and
Kalinsky K: AKT in cancer: New molecular insights and advances in
drug development. Br J Clin Pharmacol. 82:943–956. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Matson DR, Hardin H, Buehler D and Lloyd
RV: AKT activity is elevated in aggressive thyroid neoplasms where
it promotes proliferation and invasion. Exp Mol Pathol.
103:288–293. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Robert M, Frenel JS, Bourbouloux E, Berton
Rigaud D, Patsouris A, Augereau P, Gourmelon C and Campone M:
Efficacy of buparlisib in treating breast cancer. Expert Opin
Pharmacother. 18:2007–2016. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhu L, Tian G, Yang Q, De G, Zhang Z, Wang
Y, Nie H, Zhang Y, Yang X and Li J: Thyroid hormone receptor β1
suppresses proliferation and migration by inhibiting PI3K/Akt
signaling in human colorectal cancer cells. Oncol Rep.
36:1419–1426. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lim W, Jeong M, Bazer FW and Song G:
Coumestrol inhibits proliferation and migration of prostate cancer
cells by regulating AKT, ERK1/2 and JNK MAPK cell signaling
cascades. J Cell Physiol. 232:862–871. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Clark PA, Bhattacharya S, Elmayan A,
Darjatmoko SR, Thuro BA, Yan MB, van Ginkel PR, Polans AS and Kuo
JS: Resveratrol targeting of AKT and p53 in glioblastoma and
glioblastoma stem-like cells to suppress growth and infiltration. J
Neurosurg. 126:1448–1460. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Han XG, Du L, Qiao H, Tu B, Wang YG, Qin
A, Dai KR, Fan QM and Tang TT: CXCR1 knockdown improves the
sensitivity of osteosarcoma to cisplatin. Cancer Lett. 369:405–415.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yamaguchi M, Osuka S, Weitzmann MN, Shoji
M and Murata T: Increased regucalcin gene expression extends
survival in breast cancer patients: Overexpression of regucalcin
suppresses the proliferation and metastatic bone activity in
MDA-MB-231 human breast cancer cells in vitro. Int J Oncol.
49:812–822. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang Q, Pan Y, Cheng Y and Li H, Liu D
and Li H: Lunasin suppresses the migration and invasion of breast
cancer cells by inhibiting matrix metalloproteinase-2/-9 via the
FAK/Akt/ERK and NF-κB signaling pathways. Oncol Rep. 36:253–262.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang SX, Polley E and Lipkowitz S: New
insights on PI3K/AKT pathway alterations and clinical outcomes in
breast cancer. Cancer Treat Rev. 45:87–96. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ried SJ and Shi Y: Molecular mechanisms of
caspase regulation during apoptosis. Nat Rev Mol Cell Biol.
5:897–907. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fan LL, Sun GP, Wei W, Wang ZG, Ge L, Fu
WZ and Wang H: Melatonin and doxorubicin synergistically induce
cell apoptosis in human hepatoma cell lines. World J Gastroenterol.
16:1473–1481. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang L, Jin F, Qin A, Hao Y, Dong Y, Ge S
and Dai K: Targeting Notch1 signaling pathway positively affects
the sensitivity of osteosarcoma to cisplatin by regulating the
expression and/or activity of Caspase family. Mol Cancer.
13:1392014. View Article : Google Scholar : PubMed/NCBI
|