Introduction
Primary liver cancer has the fifth-highest incidence
of malignant tumors and is the third-leading cause of
cancer-associated mortality, 90% of cases are hepatocellular
carcinoma (HCC) (1). In certain areas
of Asia and the Middle East, HCC ranks as the leading cause of
cancer-associated mortality (2).
Surgical resection, which is the first-choice treatment modality
for HCC, can significantly improve the prognosis and prolong the
survival time of patients. However, surgery inevitably induces
neuroendocrine, metabolic and inflammatory responses (3,4), directly
affecting post-operative recovery and the survival time of patients
(5,6).
Consequently, the induction of this immune response can be
injurious to long-term prognosis during the perioperative period.
Therefore, the perioperative period is a critical period for
long-term prognosis. The primary method of improving the long-term
prognosis of tumor patients is to understand the mechanisms of
tumor metastasis and corresponding immune responses. Recently,
substantial attention has been paid to the association between
methods of anesthesia and long-term prognosis (7). As an important role in the whole
perioperative period, the responsibility of the anesthetists to
improve the long-term prognosis of cancer patients has been
increasingly appreciated in recent years (8). Metastasis is one of the most important
characteristics of malignant tumors, serving an important role in
treatment efficacy and quality of life of patients (9). The metastasis of tumors comprises of a
complex series of events and can be divided into a number of
sequential steps: i) Release from the primary tumor and invasion
into the surrounding tissues; ii) intravasation; iii) transition in
the blood circulation; iv) arrest in the capillary bed; v)
extravasation; and vi) growth at preferred sites (10). In this cascade, angiogenesis, the
formation of a new blood supply from pre-existing vasculature, is a
critical step of tumor metastasis (11). Angiogenesis is a complex multistage
process that is regulated by the balance between angiogenic and
angiostatic factors (12–14). Among these associated factors,
vascular endothelial growth factor (VEGF) has been demonstrated to
be one of the most important angiogenic mediators (15). VEGF is the most commonly studied
angiogenic mediator and has been well established as a molecular
target in clinical studies (16,17) In
addition to its role in stimulating angiogenesis, VEGF induces
vascular permeability to circulating macromolecules (16). Previous studies (18,19) have
shown that signal transducer and activator of transcription 3
(STAT3) regulates angiogenesis primarily by modulating VEGF
expression, although several other candidates affected by STAT3
have been reported to show distinct effects on angiogenesis.
General anesthesia (GA) and GA combined with
epidural block (GEA) are the most commonly anesthetic methods in
hepatectomy. GEA can prevent or attenuate the surgical stress
response by blocking afferent neural transmission, which prevents
harmful afferent input from reaching the central nervous system
(20). Previous studies (21,22) have
shown that the choice of anesthesia methods was only associated
with perioperative stress and immune function for a short time.
However, animal experiments and clinical studies (23,24) in
recent years have reported that different anesthetic methods serve
important roles in cancer recurrence or metastasis. Compared with
GA, GEA significantly reduces the dependency on opioids (25). Opioids, including morphine, one of the
most commonly used, can inhibit cellular and humoral immune
function (26,27). An animal study (28) strongly indicated that opioids,
including morphine and fentanyl, can promote tumor metastasis and
recurrence. Clinical trials (29,30) have
demonstrated that morphine can accelerate the growth of breast
tumors. Consequently, paravertebral anesthesia and analgesia
combined with GA led to a lower incidence of cancer recurrence or
metastasis. Recent research indicates that the association of
opioids with tumor behavior is complex and remains unclear
(31).
The present study compared the anti-metastatic
effect of GA and GEA on tumor-bearing rats with hepatectomy. The
results revealed that the anti-metastatic effect of GEA is
significant superior to GA. Furthermore, the mechanism of the two
anesthesia methods on tumor metastasis and angiogenesis was
investigated. As the results of the present study demonstrate, one
possible mechanism by which GEA suppresses tumor metastasis and
angiogenesis is the improvement of the expression of interferon-γ
(IFN-γ) and the inhibition of transforming growth factor-α (TGF-α),
VEGF and p-STAT3.
Materials and methods
Materials
The rat hepatic cancer cell line Morris hepatoma
3924A was purchased from American Type Culture Collection
(Manassas, VA, USA) Morphine, bupivacaine and fentanyl were
purchased from Shanghai Sangon Biotechnology Co., Ltd. (Shanghai,
China). The cell line was cultured at 37°C in a humidified
atmosphere containing 5% CO2 in Dulbecco's modified
Eagle's medium with 10% fetal bovine serum (FBS), 100 U
ml−1 penicillin and 100 mg ml−1 streptomycin
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 30 male
American Certification Institute (ACI) rats (age, 12–18 weeks;
weight, 200–250 g) were purchased from the Experimental Animal
Center of Changchun Biological Institute (Changchun, China), and
they were housed at a temperature of 20–22°C, relative humidity of
50–60% and 12/12 h light/dark cycles. They were also provided with
free access to food and water. The 7-0 injury-free operative
sutures were purchased from Shanghai Surgical Suture Company. ELISA
kit and antibodies used in the present study were purchased from
Invitrogen (Thermo Fisher Scientific, Inc.). Details were as
follows: IFN-γ ELISA kit (cat no. BMS621); TGF-α ELISA kit (cat no.
EHTGFA); VEGF ELISA kit (cat no. ERVEGFACL); phospho-STAT3 (Tyr705)
polyclonal antibody (cat no. 44-380G; dilution, 1:1,000); and
phospho-EGFR (Tyr1086) polyclonal antibody (cat no. 36-9700;
dilution, 1:500). The anti-rabbit IgG secondary antibody,
horseradish peroxidase (cat no. P7899; dilution, 1:100,000) was
provided by Sigma-Aldrich (Merck KGaA; Darmstadt, Germany).
Animals and the tumor xenograft
model
All animal experiments were performed in compliance
with the Animal Management Rules of the Ministry of Health of the
People's Republic of China (http://www.medste.gd.cn/Html/sciedu/sysgl/Class3141/24961720080910094400.html)
and approved by the Animal Care and Use Committee of Jilin
University (Changchun, China). All animals were pathogen-free and
allowed ad libitum access to food and water. To produce the tumor
xenograft model, MH3924A cells in the logarithmic phase were
injected subcutaneously into the upper axillary fossa of the male
ACI rats (12–18 weeks, n=10 per group). After 14 days, subcutaneous
tumors were surgically removed. Fresh tumor tissues were chosen and
divided into l-mm3 tumor pieces. The male rats were
fixed on a frame prior to anesthesia, then the abdominal body hair
was removed and the animals were disinfected with iodine. A 5-ml
syringe was placed at the back of the fixed rats to raise the
abdomen of the rats for surgical convenience. Next, when the size
of incision in upper abdomen reached ~0.5 cm, the skin,
subcutaneous tissue and peritoneal tissue was consecutively cut and
then the rat liver was exposed. The left lobe was cleaned with wet
cotton and the liver capsule was punctured with ophthalmic
scissors, making a further long cut under the capsule. Next, the
as-prepared tumor tissue was inserted into the long cut, which was
closed with 7-0 microscopic sutures. Prior to abdominal closure, it
should be confirmed that there was no blister tissue out and no
bleeding.
GEA
When the tumor volume reached ~300 mm3,
these tumor-bearing rats were randomly divided into three treatment
groups: One group was treated with hepatectomy and general
anesthesia combined with epidural block (group G + E), one group
was treated with hepatectomy and general anesthesia (group G) and
one group, the control group, did not undergo any treatment (group
C). There were 10 rats included in each group.
GA
A simple tumor resection was performed under general
anesthesia. Specifically, sevoflurane together with oxygen was
injected into a self-made anesthesia box. Next, the retraction
response of rat toes to stimulus was continuously assessed to
determine the degree of anesthesia. When the retraction response of
rat toes to stimulus disappeared, tracheal incubation was conducted
for the rat by transmission light intubation. Specifically, the
50-W illuminant was placed 5 cm away from the neck of the rat. In
this way, the light could pass through the neck skin and trachea
wall in sequence. A mini sized spatula with a 145° angle were used
to push away the tongue for clear observation of the epiglottis and
vocal cord movement under the transmitted light, thus an 18 G
cannula was inserted under a bright view. The regular movement of
the rat thorax was observed following connection with the
ventilator with mechanical ventilation. The parameters of the
ventilator were set as follows: Breathing rate, 55/min; tidal
volume, 3 ml/100 g; inspiration rate: expiration rate, 1.5:1;
sevoflurane concentration, 2–3% (v/v). To maintain the anesthesia
effect, morphine (0.2 ml/100 g, 10% solution) was injected
intraperitoneally. When the rats woke following surgery, the
endotracheal tube was removed.
Epidural anesthesia
Perioperative epidural analgesia consisted of a
bolus of 6–9 ml of 0.25% bupivacaine followed by a postoperative
infusion of bupivacaine (0.1–0.2%) with or without 2 µg/ml of
fentanyl at a rate of 6–9 ml/h. Specifically, the limbs and head of
the rat were fixed following the success of tracheal incubation
under general anesthesia. A self-made sterile PVC epidural catheter
with diameter of 0.3 mm was inserted via removing the eighth or
ninth thoracic spinous process using a vertical needling and was
advanced 1 cm cephalad into the epidural space. Subsequently, 0.25%
of bupivacaine was micro-injected via the epidural catheter.
Hepatectomy
According to observations, the tumor size was
suitable for hepatectomy and no distant metastasis was observed at
day 14 following inoculation with tumor cells. Hepatectomy was
conducted following successful anesthesia with the assistance of
undergraduate students majoring in liver surgery (Departments of
General Surgery, China-Japan Union Hospital Jilin University,
Changchun, China). Specifically, the skin, subcutaneous tissue and
peritoneal tissue was consecutively removed from the original upper
abdomen incision. Next, the left lobe of the liver was exposed and
the megascopic in situ tumor tissue was removed, as was 5 mm
normal liver tissue around the edge of tumor. The 7-0 microscopic
suture was used to close the abdomen following the confirmation of
no bleeding.
Blood sample collection and ELISA
assay
Venous blood samples were collected from orbits of
rats immediately prior to surgery and at 2, 7 and 30 days following
surgery. The blood samples were left to clot at room temperature
and then stored overnight in a refrigerator to contract the clot.
The clot was removed and the remaining material was centrifuged at
4,000 × g for 5 min and the resulting serum was stored at −80°C
until analysis. Plasma levels of IFN-γ, TGF-α and VEGF were
measured by an ELISA. Plasma levels of each cytokine were measured
in duplicate from one aliquot of each sample and the mean value
from these two measurements was used as the final concentration.
Quantification was achieved using an automated microplate reader.
Concentrations of all cytokines were reported in pg/ml. The rats
were sacrificed 30 days following surgery. The lungs were removed
and collected for the following experiments. The metastatic nodes
in the lungs of mice were visualized by fixing them in Bouin's
solution (saturated picric acid: Formalin: Acetic acid, 15:5:1) at
room temperature for 24 h and photographed using a high-definition
digital camera (IXUS 110, Canon Inc., Tokyo, Japan) to evaluate the
anti-metastatic effects of general anesthetic and general
anesthetic with epidural block.
Western blot analysis
For each group, 0.1 g of the lung tissue was
collected for protein extraction and lysed using a Dounce
homogenizer (20 strokes) in 400 µl of buffer containing 50 mM tris
(hydroxymethyl) aminomethane hydrochloride (Tris-HCl), pH 7.6, 42
mM KCl, 5 mM MgCl2, 1 mM dithiothreitol (DTT), 0.5%
3-[(3-cholamidopropyl) dimethylammonio]-1-propanesulfonate hydrate,
1 mM phenylmethylsulfonyl-fluoride and 1 µg/ml leupeptin
(Sigma-Aldrich; Merck KGaA), and then centrifuged at 15,000 × g for
20 min at 4°C. The protein concentrations of the lysates were
determined by BCA protein assay. Equal amounts (50 µg) of protein
were separated on a 10% SDS-PAGE and transferred onto
polyvinylidene difluoride membranes. The membranes were blocked
with 5% non-fat milk in Tris-buffered saline/0.1% Tween-20 for 2 h,
subsequently blotted with respective primary antibodies overnight
at 4°C as described previously in the materials section, and then
blotted with horseradish peroxidase-conjugated secondary antibody
at 37°C for 1 h. The protein bands were visualized using an
enhanced chemiluminescent agent (Fuzhou Maixin Biotech Co., Ltd.,
Fuzhou, China). Protein levels were quantified by density analysis
using ImageJ software version 1.62 (National Institutes of Health,
Bethesda, MD, USA), and expressed as the interest protein/internal
control.
Statistical analysis
Data from ≥3 groups were compared using one-way
analysis of variance followed by Dunnett's post-hoc test using the
SPSS software package (version 13.0; SPSS Inc., Chicago, IL, USA).
The experiments were performed in triplicate and the data are
presented as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
The anti-metastatic effects of GA and
GEA
The evaluation on the in vivo anti-metastatic
activity of general anesthesia and epidural anesthesia was
performed. In the in vivo experiment, the rats were
sacrificed 30 days after surgery and the lungs were dissected. The
average numbers of metastatic nodules on the lung surface in
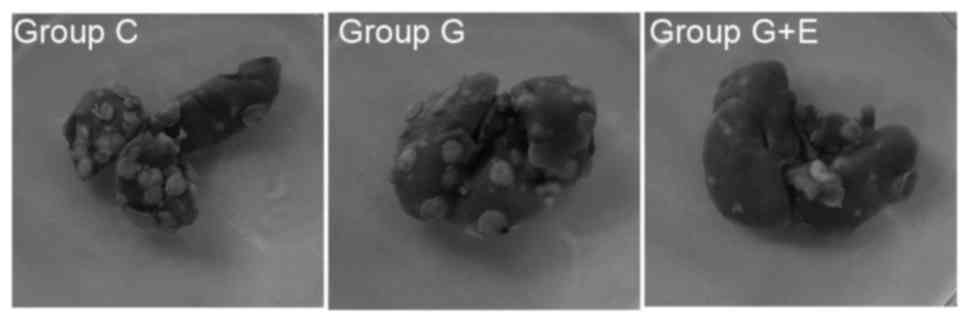
different groups were counted. Representative photos of the lungs
with the metastatic nudes are presented in Fig. 1. As depicted in Table I, the number of metastasis nodules in
the G + E group was 18±6, which was significantly (P<0.05) less
than that in group C (40±6) or group G (38±5). This result
indicated that GEA could effectively inhibit lung metastasis
compared with GA following hepatectomy. The conditions of abdominal
lymph node metastasis, the rate of malignant ascites and number of
abdominal wall planting nodules also validated this conclusion. As
presented in Table I, the number of
animals for which there was a presence of abdominal lymph node
metastasis was significantly lower for group G + E (1/10) than for
group C (7/10) or group G (6/10). For group C and group G, the
rates of malignant ascites were higher than that of group G + E,
which demonstrated the low metastatic rate following surgery with
epidural anesthesia. Abdominal wall-implanted nodules represented
another important evaluation index for metastatic condition.
According to Table I, the number of
rats with abdominal wall-implanted nodules in group G + E was 2/10,
which was significantly lower than that for group C (8/10) or group
G (5/10). Hepatectomy with different anesthetic techniques revealed
different rates of tumor metastasis. Notably, GEA could effectively
inhibit tumor metastasis compared to GA following hepatectomy.
 | Table I.Quantified data of metastases
analysis (n=10). |
Table I.
Quantified data of metastases
analysis (n=10).
| Groups | Lung
metastasesa, n | Abdominal lymph
node metastasis, n | Rate of malignant
ascites, n | Abdominal
wall-implanted nodules, n | Volumes of lung
with all metastasesa,
cm3 |
|---|
| Group C |
40±6 | 7/10 | 8/10 | 8/10 |
14.85±1.66 |
| Group G + E |
18±6b,c | 1/10b,c | 2/10b,c | 2/10b,c |
12.93±1.52 |
| Group G |
38±5 | 6/10 | 7/10 | 5/10 |
13.06±2.08 |
Changes in cytokine concentration in
the serum of rats that underwent different treatments determined by
ELISA
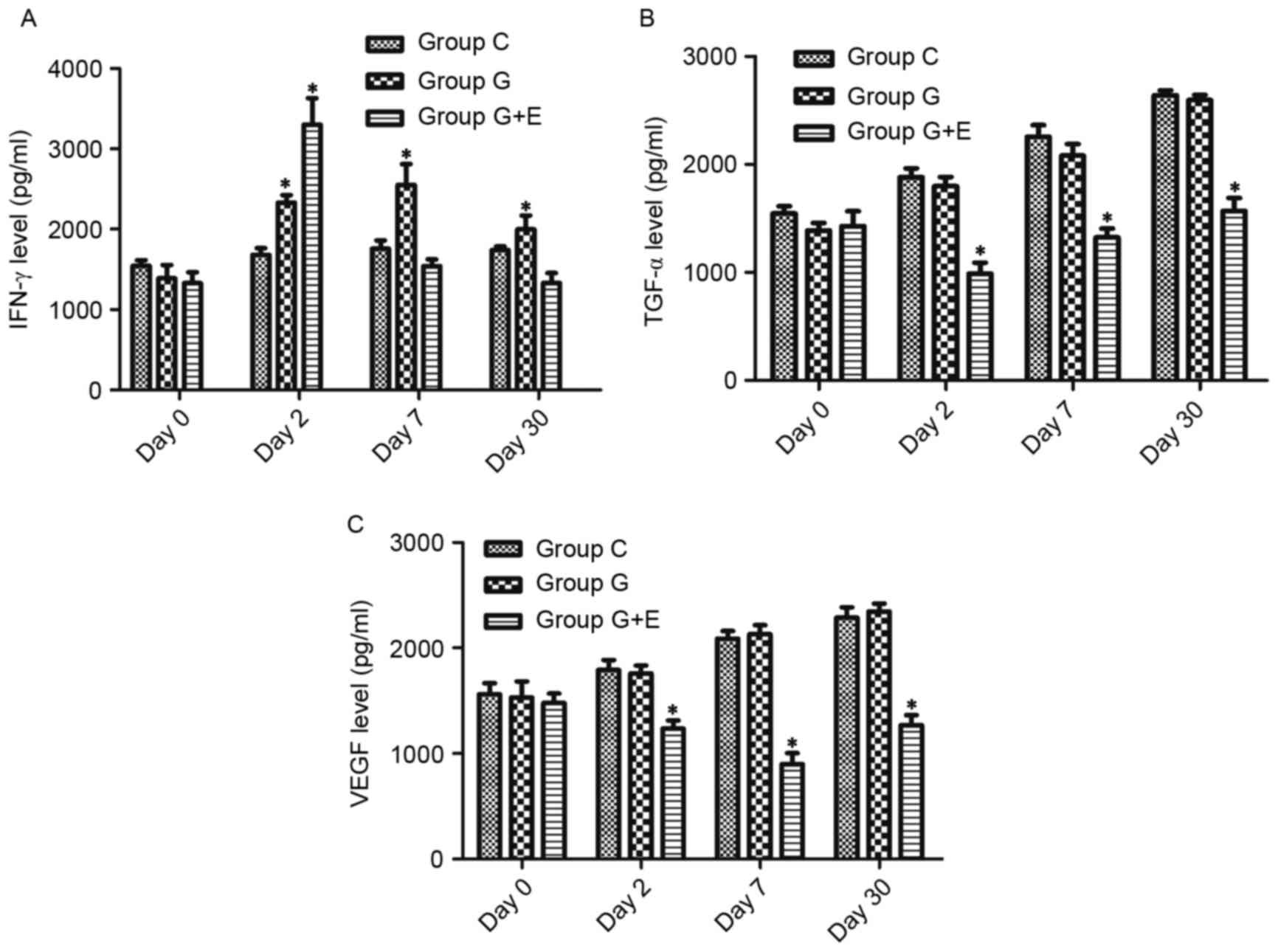
As shown in Fig. 2A,
there was no significant difference in IFN-γ levels between groups
C, G and G + E on day 0. On day 2, the concentration of IFN-γ in
group G + E (3,411±232 pg/ml) was significantly higher than that in
group C (1,497±146 pg/ml) and G (2,578±165 pg/ml). On day 7 and day
30, there was no significant difference between group C and G, but
these concentrations were significantly higher than those in group
G + E. For group G + E, the concentration of IFN-γ at day 7 and day
30 was slightly lower than that on day 2. Similarly, there was no
significant difference in regard to concentrations of TGF-α and
VEGF between group C, G and G + E on day 0 (Fig. 2B and C). On the following days,
concentrations of TGF-α and VEGF were significantly lower in group
G + E than in groups C and G (P<0.05). Notably, the
concentration of TGF-α increased slowly over time following
surgery. However, the concentration of VEGF was at its minimum on
day 7. Groups C and G exhibited increases in the concentration of
TGF-α and VEGF, whereas the concentration in group G + E rats
remained lower than those in groups C and G.
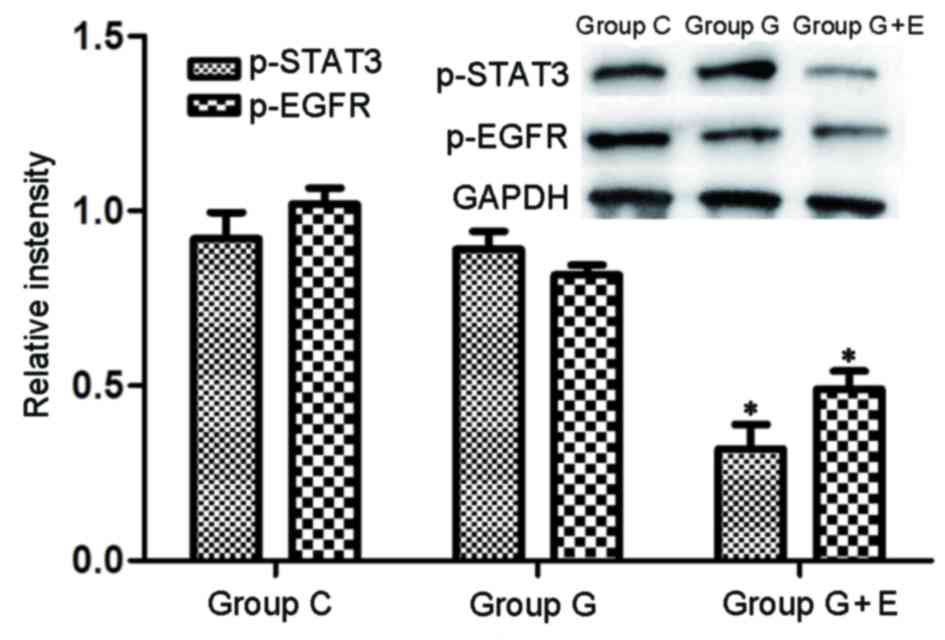
Levels of phosphorylated STAT3
(p-STAT3) and p-VEGF in lung metastasis tissues determined by
western blot analysis
To investigate the mechanism of the
anesthesia-induced anti-metastatic effect further, levels of
p-STAT3 and p-EGFR lung metastatic tissues were determined by
western blot analysis (Fig. 3). The
levels of p-STAT3 and p-EGFR in group G were slightly lower than
those in group C. It was shown that there were significantly
(P<0.05) lower levels of p-STAT3 and p-EGFR in group G + E than
in groups C and G. Evidence indicates that aberrant STAT3 signaling
promotes the initiation and progression of human cancer by either
inhibiting apoptosis or inducing cell proliferation, angiogenesis,
invasion and metastasis (32–34). Additionally, expression of EGFR is
altered in a variety of human cancer types, including carcinoma of
the lung, breast, head and neck, ovary and bladder, as well as in
glioma (35–37). STAT3 and EGFR serve notable roles in
tumor development and metastasis (34,38). The
results of the present study indicated that GEA significantly
inhibited tumor metastasis by reducing levels of p-STAT3 and
p-EGFR.
Discussion
The perioperative period is a critical period of
damage to the body's defense mechanism and has a substantial
influence on the long-term prognosis of tumor patients. The role of
anesthesia in improving the long-term prognosis of patients
following surgery has been known and taken seriously in recent
years.
It has been reported that there are four major
mechanisms that surgery could contribute to an increase in tumor
metastasis: i) The operation and touch of the tumor during surgery
increased the chance of tumor cells releasing into the bloodstream
(39); ii) Tumors in situ
(TIS) serve a role in inhibiting angiogenesis: In surgery, the
resection of TIS disrupts this line of defense, with the residual
lesions promoting diseases and inducing the growth of blood vessels
to increase the risk of tumor metastasis (40); iii) Perioperative immune suppression:
Levels of epinephrine and norepinephrine during the perioperative
period are substantially increased, which is considered to be the
key to the connection between stress response and cancer
progression (41); iv) The increase
in local and systemic growth factor (EGF and VEGF) release in
surgery promotes the recurrence of local and distant tumors
(42). Furthermore, anti-angiogenic
factors, including angiostatin and endostatin, decrease during
surgery (43). Catecholamine promotes
growth in a variety of tumor types by activating the STAT3
signaling pathway to increase the level of VEGF and inhibit the
cellular immune response (5).
In the present study, the levels of IFN-γ in the
plasma of rats in group G + E were higher than those in the two
other groups, whereas the levels of VEGF and TGF-α in group G + E
were lower than those in other two groups, as was the degree of
STAT3 activation. The present study indicates that compared with
GA, GEA can reduce the incidence of liver cancer metastasis
following surgery. A possible mechanism may include the inhibition
of STAT3 and relevant cytokines, including IFN-γ and TGF-α.
IFN-γ, which is secreted by T cells, can suppress
tumor cell proliferation, upregulate the expression of surface
major histocompatibility complex antigens and tumor necrosis factor
and prevent tumor growth via inhibition of tumor angiogenesis and
other mechanisms (44). A previous
study demonstrated that IFN-γ can decrease the defense of tumors
cells escaping immune system by regulating expression of the Fas
cell surface death receptor/Fas ligand and enhancing sensitivity to
Fas-mediated apoptosis, thus inhibiting the malignant proliferation
of tumor cells (45).
EGFR is a membrane-bound receptor tyrosine kinase
that belongs to the ErbB subfamily (46). On ligand binding, EGFR initiates the
activation of a series of cellular signal transduction pathways
that regulate cell proliferation and survival (47). The primary ligands for EGFR include
EGF and TGF-α in liver cells (48).
TGF-α produced by liver cells can bind with EGFR to promote the
proliferation of liver cells by autocrine, or increase the
proliferation of the vascular endothelium by paracrine (6). EGFR is activated by ligand binding,
which causes it to homodimerize, activating downstream pathways,
including the RAS proto-oncogene, GTPase/Raf proto-oncogene,
serine/threonine kinase mitogen-activated protein kinase pathway,
the Janus kinase (JAK)/STAT pathway and the phosphoinositide
3-kinase/RAC serine/threonine-protein kinase pathway (49). Two possible pathways for STAT3
activation exist, a JAK-dependent mechanism and a JAK-independent
one, which initiates dimer formation of STAT3 in the cytoplasm.
Thereafter, activated STAT3 transfers into the nucleus, binds to
specific DNA sequences and induces transcription of target genes
(47,50). This sequential process promotes tumor
cell proliferation and inhibits apoptosis by regulating expression
of the apoptosis suppressor genes, such as MCL1, BCL2 family
apoptosis regulator and BCL2-like 1, and cell cycle regulatory
factors, such as cyclin D1/D2 and MYC proto-oncogene, BHLH
transcription factor (3).
In the present study, the expression levels of VEGF
and TGF-α in group C were significantly higher than those in group
G; however, those in group G were higher than those in group G + E.
Overexpression of EGFR and its relevant ligands or the sequential
activation of downstream signal pathways is observed in numerous
tumor types (37,38). The formation of tumors is promoted by
the following methods: Overexpression of EGFR triggers the
enhancement of downstream signal transduction (51); the mutation of EGFR or overexpression
of its ligands results in continuous EGFR activation (52); the function of autocrine loop is
enhanced (53), mechanisms of
downregulation of oncogenic receptors are lost (54); and abnormal signal transduction
pathways are activated (55). In the
present study, the increase in the expression of EGFR and its
ligand demonstrated that the EGFR signal transduction pathway
alters the formation of liver cancer metastasis via at least two
mechanisms: The overexpression of TGF-α and EGFR, leading to the
formation of an autocrine loop and increased downstream signal
transduction, respectively. STAT3 is an oncogene that is closely
associated with tumor development, evolvement, differentiation and
immunity (18,19). The sustained activation or
overexpression of STAT3 is frequently observed in skin, lung and
breast cancer (56). To the best of
our knowledge, mutations to STAT3 have not yet been observed, so we
hypothesize that the sustained activation of STAT3 is triggered by
enhancement of the intensity of upstream signal transduction
pathways.
Song and Grandis (57)
demonstrated that cell lines overexpressing TGF-α exhibited
sustained activation of STAT3, and the addition of exogenous TGF-α
could enhance the activation of squamous cell carcinoma of the head
and neck. For certain tumors, the sustained activation of STAT3
resulted in the enhancement of activation of EGFR signal
transduction pathways (57). STAT3
binds to activated EGFR via SH2 domains; stimulation of EGFR
induces Tyr705 phosphorylation of STAT3, which is a prerequisite
for dimerization of STATs, which again occurs via the SH2 domains
(58). The dimerization of
tyrosine-phosphorylated STATs leads to their nuclear accumulation,
DNA binding at specific sites and the transcriptional activation of
target genes (57). Research on
breast cancer, squamous cell carcinoma of the head and neck and
epidermoid cancer has revealed that sustained activation of STAT3
is relevant to the formation of the EGFR/TGF-α autocrine loop
(59–61).
The present study revealed that the metastasis of
liver cancer was decreased in rats that underwent surgery with GEA
compared with those that underwent GA. The results demonstrated
that the mechanism by which this occurred may be associated with
the inhibition of STAT3 and certain relevant cytokines. However,
these results were obtained from animal experiments, which require
careful consideration and further confirmation in clinical
trials.
References
|
1
|
Zhu RX, Seto WK, Lai CL and Yuen MF:
Epidemiology of hepatocellular carcinoma in the Asia-pacific
region. Gut Liver. 10:332–339. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Llovet JM: Updated treatment approach to
hepatocellular carcinoma. J Gastroenterol. 40:225–235. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Buggy DJ and Smith G: Epidural anesthesia
and analgesia: Better outcome after major surgery? Growing evidence
suggests so. BMJ. 319:530–531. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ogawa K, Hirai M, Katsube T, Murayama M,
Hamaguchi K, Shimakawa T, Naritake Y, Hosokawa T and Kajiwara T:
Suppression of cellular immunity by surgical stress. Surgery.
127:329–336. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ben-Eliyahu S: The price of anticancer
intervention. Does surgery promote metastasis? Lancet Oncol.
3:578–579. 2002.PubMed/NCBI
|
|
6
|
Ben-Eliyahu S, Page GG, Yirmiya R and
Shakhar G: Evidence that stress and surgical interventions promote
tumor development by suppressing natural killer cell activity. Int
J Cancer. 80:880–888. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Deiner S and Silverstein JH: Long-term
outcomes in elderly surgical patients. Mt Sinai J Med. 79:95–106.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Deflandre E and Lacroix S: It is time for
anesthetists to act as perioperative physicians. Minerva
Anestesiol. 82:257–258. 2016.PubMed/NCBI
|
|
9
|
Martin TA, Ye L, Sanders AJ, Lane J and
Jiang WG: Cancer invasion and metastasis: Molecular and cellular
perspectiveMetastatic Cancer Clinical Biological Perspectives 2013.
Jandial R: Landes Bioscience; Austin, TX: pp. 135–168. 2013
|
|
10
|
Tsuchiya Y, Sawada S, Yoshioka I, Ohashi
Y, Matsuo M, Harimaya Y, Tsukada K and Saiki I: Increased surgical
stress promotes tumor metastasis. Surgery. 133:547–555. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ucuzian AA, Gassman AA, East AT and
Greisler HP: Molecular mediators of angiogenesis. J Burn Care Res.
31:158–175. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Distler JH, Hirth A, Kurowska-Stolarska M,
Gay RE, Gay S and Distler O: Angiogenic and angiostatic factors in
the molecular control of angiogenesis. Q J Nucl Med. 47:149–161.
2003.PubMed/NCBI
|
|
13
|
Simons M: Integrative signaling in
angiogenesis. Mol Cell Biochem. 264:99–102. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen Z and Han ZC: STAT3: A critical
transcription activator in angiogenesis. Med Res Rev. 28:185–200.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hoeben A, Landuyt B, Highley MS, Wildiers
H, Van Oosterom AT and De Bruijn EA: Vascular endothelial growth
factor and angiogenesis. Pharmacol Rev. 56:549–580. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Senger DR, Galli SJ, Dvorak AM, Perruzzi
CA, Harvey VS and Dvorak HF: Tumor cells secrete a vascular
permeability factor that promotes accumulation of ascites fluid.
Science. 219:983–985. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Leung DW, Cachianes G, Kuang WJ, Goeddel
DV and Ferrara N: Vascular endothelial growth factor is a secreted
angiogenic mitogen. Science. 246:1306–1309. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xie TX, Wei D, Liu M, Gao AC, Ali-Osman F,
Sawaya R and Huang S: Stat3 activation regulates the expression of
matrix metalloproteinase-2 and tumor invasion and metastasis.
Oncogene. 23:3550–3560. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xie TX, Huang FJ, Aldape KD, Kang SH, Liu
M, Gershenwald JE, Xie K, Sawaya R and Huang S: Activation of stat3
in human melanoma promotes brain metastasis. Cancer Res.
66:3188–3196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Exadaktylos AK, Buggy DJ, Moriarty DC,
Mascha E and Sessler DI: Can anesthetic technique for primary
breast cancer surgery affect recurrence or metastasis?
Anesthesiology. 105:660–664. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bauer M, Rensing H and Ziegenfuss T:
Anesthesia and perioperative immune function. Anaesthesist.
47:538–556. 1998.(In German). View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Borsook D, George E, Kussman B and Becerra
L: Anesthesia and perioperative stress: Consequences on neural
networks and postoperative behaviors. Prog Neurobiol. 92:601–612.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Marshall L, Khan AH and Buggy DJ: Can
anaesthetic and analgesic techniques for cancer surgery affect
cancer recurrence and metastasis? Curr Anesthesiology Rep.
5:190–202. 2015. View Article : Google Scholar
|
|
24
|
Vahabi S and Eatemadi A: Effects of
anesthetic and analgesic techniques on cancer metastasis. Biomed
Pharmacother. 87:1–7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ogren SO and Holm AC: Test-specific
effects of the 5-HT reuptake inhibitors alaproclate and zimelidine
on pain sensitivity and morphine analgesia. J Neural Transm.
47:253–271. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sacerdote P, Bianchi M, Gaspani L,
Manfredi B, Maucione A, Terno G, Ammatuna M and Panerai AE: The
effects of tramadol and morphine on immune responses and pain after
surgery in cancer patients. Anesth Analg. 90:1411–1414. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yeager MP, Colacchio TA, Yu CT,
Hildebrandt L, Howell AL, Weiss J and Guyre PM: Morphine inhibits
spontaneous and cytokine-enhanced natural killer cell cytotoxicity
in volunteers. Anesthesiology. 83:500–508. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sasamura T, Nakamura S, Iida Y, Fujii H,
Murata J, Saiki I, Nojima H and Kuraishi Y: Morphine analgesia
suppresses tumor growth and metastasis in a mouse model of cancer
pain produced by orthotopic tumor inoculation. Eur J Pharmacol.
441:185–191. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ash SA and Buggy DJ: Does regional
anaesthesia and analgesia or opioid analgesia influence recurrence
after primary cancer surgery? An update of available evidence. Best
Pract Res Clin Anaesthesiol. 27:441–456. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gupta K, Kshirsagar S, Chang L, Schwartz
R, Law PY, Yee D and Hebbel RP: Morphine stimulates angiogenesis by
activating proangiogenic and survival-promoting signaling and
promotes breast tumor growth. Cancer Res. 62:4491–4498.
2002.PubMed/NCBI
|
|
31
|
Cao LH, Li HT, Lin WQ, Tan HY, Xie L,
Zhong ZJ and Zhou JH: Morphine, a potential antagonist of cisplatin
cytotoxicity, inhibits cisplatin-induced apoptosis and suppression
of tumor growth in nasopharyngeal carcinoma xenografts. Sci Rep.
6:187062016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rahaman SO, Vogelbaum MA and Haque SJ:
Aberrant Stat3 signaling by interleukin-4 in malignant glioma
cells: Involvement of IL-13Ralpha2. Cancer Res. 65:2956–2963. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lamprecht B, Kreher S, Anagnostopoulos I,
Jöhrens K, Monteleone G, Jundt F, Stein H, Janz M, Dörken B and
Mathas S: Aberrant expression of the Th2 cytokine IL-21 in Hodgkin
lymphoma cells regulates STAT3 signaling and attracts Treg cells
via regulation of MIP-3alpha. Blood. 112:3339–3347. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yue P, Zhang X, Paladino D, Sengupta B,
Ahmad S, Holloway RW, Ingersoll SB and Turkson J: Hyperactive EGF
receptor, Jaks and Stat3 signaling promote enhanced colony-forming
ability, motility and migration of cisplatin-resistant ovarian
cancer cells. Oncogene. 31:2309–2322. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sordella R, Bell DW, Haber DA and
Settleman J: Gefitinib-sensitizing EGFR mutations in lung cancer
activate anti-apoptotic pathways. Science. 305:1163–1167. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ali SM, Wang K, Johnson A, Rodriguez AA,
Elvin JA, Vergilio JA, Suh JH, Chumsri S, Morosini D, Yelensky R,
et al: Abstract P6-03-02: EGFR genomic alterations in 5,605 cases
of refractory and metastatic breast cancer. Cancer Res. 76 Suppl
4:P6–03-02. 2016. View Article : Google Scholar
|
|
37
|
Milagre CS, Gopinathan G, Everitt G,
Thompson RG, Kulbe H, Zhong H, Hollingsworth RE, Grose R, Bowtell
DD, Hochhauser D and Balkwill FR: Adaptive upregulation of EGFR
limits attenuation of tumor growth by neutralizing IL6 antibodies,
with implications for combined therapy in ovarian cancer. Cancer
Res. 75:1255–1264. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Falchook GS, Naing A, Wheler JJ,
Tsimberidou AM, Zinner R, Hong DS, Fu S, Piha-Paul SA, Janku F,
Hess KR, et al: Dual EGFR inhibition in combination with anti-VEGF
treatment in colorectal cancer. Oncoscience. 1:540–549. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Engilbertsson H, Aaltonen KE, Björnsson S,
Kristmundsson T, Patschan O, Rydén L and Gudjonsson S:
Transurethral bladder tumor resection can cause seeding of cancer
cells into the bloodstream. J Urol. 193:53–57. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang W, Liu JN and Tan XY: Vaccination
with xenogeneic tumor endothelial proteins isolated in situ
inhibits tumor angiogenesis and spontaneous metastasis. Int J
Cancer. 125:124–132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ramírez MF, Huitink JM and Cata JP:
Perioperative clinical interventions that modify the immune
response in cancer patients. Open J Anesthesiology. 3:133–139.
2013. View Article : Google Scholar
|
|
42
|
Wang X, Cao W, Mo M, Wang W, Wu H and Wang
J: VEGF and cortactin expression are independent predictors of
tumor recurrence following curative resection of gastric cancer. J
Surg Oncol. 102:325–330. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chavira E, Wilson M, Assaf SA, Ingles SA,
Llanes A and Chmait R: Anti-angiogenic state in twin-twin
transfusion syndrome ameliorated after laser surgery. Ame J Obst
Gynecology. 206:S52. 2012. View Article : Google Scholar
|
|
44
|
Zielinski CE, Mele F, Aschenbrenner D,
Jarrossay D, Ronchi F, Gattorno M, Monticelli S, Lanzavecchia A and
Sallusto F: Pathogen-induced human TH17 cells produce IFN-γ or
IL-10 and are regulated by IL-1β. Nature. 484:514–518. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
O'Connell J, Bennett MW, O'Sullivan GC,
Collins JK and Shanahan F: The Fas counterattack: Cancer as a site
of immune privilege. Immunol Today. 20:46–52. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Müller CB, De Bastiani MA, Becker M,
França FS, Branco MA, Castro MA and Klamt F: Potential crosstalk
between cofilin-1 and EGFR pathways in cisplatin resistance of
non-small-cell lung cancer. Oncotarget. 6:3531–3539. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gao SP, Mark KG, Leslie K, Pao W, Motoi N,
Gerald WL, Travis WD, Bornmann W, Veach D, Clarkson B and Bromberg
JF: Mutations in the EGFR kinase domain mediate STAT3 activation
via IL-6 production in human lung adenocarcinomas. J Clin Invest.
117:3846–3856. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Abbott BD, Buckalew AR, DeVito MJ, Ross D,
Bryant PL and Schmid JE: EGF and TGF-alpha expression influence the
developmental toxicity of TCDD: Dose response and AhR phenotype in
EGF, TGF-alpha, and EGF + TGF-alpha knockout mice. Toxicol Sci.
71:84–95. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Weaver CT, Harrington LE, Mangan PR,
Gavrieli M and Murphy KM: Thl7: An effector CD4 T cell lineage with
regulatory T cell ties. Immunity. 24:677–688. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sen B, Saigal B, Parikh N, Gallick G and
Johnson FM: Sustained src inhibition results in signal transducer
and activator of transcription 3 (STAT3) activation and cancer cell
survival via altered Janus-activated kinase-STAT3 binding. Cancer
Res. 69:1958–1965. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Juergens RA and Brahmer JR: Adjuvant
treatment in non-small cell lung cancer: Where are we now? J Natl
Compr Canc Netw. 4:595–600. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Smith RA and Glynn TJ: Epidemiology of
lung cancer. Radiol Clin North Am. 38:453–470. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Fossella FV, DeVore R, Kerr RN, Crawford
J, Natale RR, Dunphy F, Kalman L, Miller V, Lee JS, Moore M, et al:
Randomized phase III trial of docetaxel versus vinorelbine or
ifosfamide in patients with advanced non-small-cell lung cancer
previously treated with platinum-containing chemotherapy regimens.
The TAX 320 non-small cell lung cancer study group. J Clin Oncol.
18:2354–2362. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Herrera B, van Dinther M, Ten Dijke P and
Inman GJ: Autocrine bone morphogenetic protein-9 signals through
activin receptor-like kinase-2/Smad1/Smad4 to promote ovarian
cancer cell proliferation. Cancer Res. 69:9254–9262. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Jean GW and Shah SR: Epidermal growth
factor receptor monoclonal antibodies for the treatment of
metastatic colorectal cancer. Pharmacotherapy. 28:742–754. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Siveen KS, Sikka S, Surana R, Dai X, Zhang
J, Kumar AP, Tan BK, Sethi G and Bishayee A: Targeting the STAT3
signaling pathway in cancer: Role of synthetic and natural
inhibitors. Biochim Biophys Acta. 1845:136–154. 2014.PubMed/NCBI
|
|
57
|
Song JI and Grandis JR: STAT signaling in
head and neck cancer. Oncogene. 19:2489–2495. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Morlacchi P, Robertson FM, Klostergaard J
and McMurray JS: Targeting SH2 domains in breast cancer. Future Med
Chem. 6:1909–1926. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lapeire L, Hendrix A, Lambein K, Van
Bockstal M, Braems G, Van Den Broecke R, Limame R, Mestdagh P,
Vandesompele J, Vanhove C, et al: Cancer-associated adipose tissue
promotes breast cancer progression by paracrine oncostatin M and
Jak/STAT3 signaling. Cancer Res. 74:6806–6819. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Penuel E, Kapp AV, Do A, Tam R, Sumiyoshi
T, Marathe C, Sa S, Peale F, Lackner M, Holden S, et al: Biomarker
evaluation in a randomized phase 2 study of MEHD7945A (MEHD) versus
cetuximab (Cet) in ≥2 line recurrent/metastatic (R/M) squamous cell
carcinomas of the head and neck (SCCHN) [MEHGAN]. Cancer Res. 75
Suppl 15:15532015. View Article : Google Scholar
|
|
61
|
Luwor RB, Baradaran B, Taylor LE, Iaria J,
Nheu TV, Amiry N, Hovens CM, Wang B, Kaye AH and Zhu HJ: Targeting
Stat3 and Smad7 to restore TGF-β cytostatic regulation of tumor
cells in vitro and in vivo. Oncogene. 32:2433–2441.
2013. View Article : Google Scholar : PubMed/NCBI
|