Introduction
Glioblastoma multiforme (GBM) is the most aggressive
astrocytic brain tumor, with the highest degree of histological
abnormality (1,2). Patients with GBM are typically
associated with a poor prognosis, with a median survival of 12–14
months, according to a statistical report of nervous system tumors
diagnosed in the United States between 2006 and 2010 (3,4). Current
therapy consists of maximal safe resection of the tumor mass
followed by radio- and chemotherapy (5). The drug of choice is temozolomide (TMZ),
but the treatment yields a median survival benefit of only 2.5
months. In addition, tumor recurrence and resistance to TMZ often
occur (6,7).
Cancer cells commonly present an increased metabolic
activity, which results in oxidative stress (8,9). The
precise role of oxidative stress in tumor biology and its
implication in cancer therapy remains a complex matter. The
excessive levels of reactive oxygen species (ROS) may result in
cell damage and apoptosis (10).
However, ROS production, which includes nitric oxide (NO), may
improve the survival, growth and neoplastic phenotype of various
cancer cells, including astrocytic brain tumors (11,12).
The neuronal nitric oxide synthase (nNOS) enzyme
synthesizes the largest amount of NO in the body, exerting an
important role in homeostasis (13).
However, nNOS and NO are also involved in brain diseases and cancer
pathogenesis (14,15). In general, brain tumors and
peritumoral areas express NOS, which improves the blood supply
required for cancer development (16,17). NO
formed by nNOS contributes to angiogenesis, vasodilation and
vascular permeability, thus serving a role in tumor growing and
malignancy (18). Also, the
non-selective NOS inhibitor N(ω)-nitro-L-arginine methyl ester
(L-NAME) controlled brain tumor growth in a rat model (19).
In summary, oxidative stress is a biochemical change
that affects tumor growth and cancer cells' response to
antineoplastic drugs. Identifying molecules that regulate oxidative
stress in TMZ-injured tumor cells will enrich our knowledge on
tumor biology and responses to anticancer therapy. The present
study evaluated whether NOS enzymes serve a role in the damage
caused by TMZ on astrocytic tumor cells. First, the effects of TMZ
(250 µM) on oxidative stress and cell viability were examined.
These effects were evaluated in astrocytoma (U251MG and U138MG) and
glioblastoma (ATCC U87MG) cell lines at 2, 48 and 72 h. Then, it
was investigated whether NOS enzymes would affect the cell damage
caused by TMZ. For that purpose, L-NAME (a nonspecific NOS
inhibitor) and 7-NI (an nNOS inhibitor) were used (20,21).
Finally, the strategy of RNAi was applied to explore the
involvement of the nNOS enzyme in such cell responses.
Materials and methods
Cell culture
The human likely glioblastoma cell line U87MG
[ATCC® HTB14™; American Type Culture
Collection (ATCC), Manassas, VA, USA], named ATCC U87MG for
simplicity, and the astrocytoma cell lines U251MG (European
Collection of Authenticated Cell Cultures, Salisbury, UK) and
U138MG (ATCC® HTB-16™), were maintained in Dulbecco's
modified Eagle's medium (DMEM)/F12 (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% (v/v)
heat-inactivated fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), 1% GlutaMAX™ (Gibco; Thermo Fisher
Scientific, Inc.), 1% penicillin/streptomycin solution and 250
ng/ml amphotericin B (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany). The cells were seeded into 25 cm3 culture
flasks and maintained at 37°C in a humidified atmosphere with 5%
CO2.
Determination of cell viability
Cells were seeded in 96-well plates at a
concentration of 0.5×104 cells/well. The cells were
incubated with 200 µM of L-NAME (Sigma-Aldrich; Merck KGaA) or 100
µM of 7-nitroindazole (7-NI; Sigma-Aldrich; Merck KGaA) 1 h prior
to the addition of 250 µM of TMZ (Orion Corporation, Espoo,
Finland). After 2, 48, or 72 h, cell viability was measured by a
quantitative colorimetric assay with MTT, which reveals the
mitochondrial activity of living cells. Briefly, 50 µl of the
MTT-labeling reagent (0.5 mg/ml) were added to each well, and the
plate was incubated for an additional 3-h period. The insoluble
formazan was dissolved with dimethyl sulfoxide, and MTT reduction
was measured at 595 nm in an absorbance plate reader
(SpectraMax® M2; Molecular Devices, LLC, Sunnyvale, CA,
USA). Experiments were carried out in triplicate in 8 independent
assays. Control cells without treatment were considered to exhibit
100% viability.
ROS test
The 2′,7′-dichlorofluorescein-diacetate (DCFH-DA)
(Sigma-Aldrich) assay was used to measure ROS production. Briefly,
cells were seeded in 96-well plates at a density of
0.5×104 cells/well. After 24 h, cells received L-NAME or
7-NI 1 h before TMZ (250 µM) exposition for 2, 48 or 72 h.
Subsequently, the medium was removed, and the cells were incubated
with DCFH-DA at a final concentration of 20 µM in DMEM/F12 for 30
min at 37°C, and next washed with Dulbecco's PBS. DCFH-DA levels
were measured in a microplate reader (SpectraMax® M2;
Molecular Devices, LLC) with excitation and emission wavelengths
set at 485 and 535 nm, respectively.
Synthetic siRNAs
The present study used a previously described siRNA
named siRNAnNOShum_4400, which targets an nNOS mRNA sequence
identified by the BIOPREDsi algorithm (22,23). This
sequence is present in exon 28 of all nNOS splicing variants α, β
and γ (5′-GCGAACGTACGAAGTGACCAA-3′; nt 4,898-4,918; NM_000620.2)
(23). siRNAnNOShum_4400 was
synthesized by Qiagen, Inc. as double-stranded RNA sequence with 21
nt. The present study also used the AllStars® Negative
Control siRNA (Qiagen, Inc.).
Cell transfection
Transfections of three different lineages (ATCC
U87MG, U251MG and U138MG) with siRNAnNOShum_4400 were carried out
with Lipofectamine® 2000 Transfection Reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) and
Opti-MEM® (Gibco; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. The effect of
siRNAnNOShum_4400 on nNOS mRNA content was determined by RT-qPCR as
described below. The present study also evaluated the effect of
siRNAnNOShum_4400 on the viability of ATCC U87MG cells injured by
TMZ. To investigate this, following siRNA transfection, the medium
was exchanged for DMEM/F12 (Thermo Fisher Scientific, Inc.),
followed by the addition of TMZ at 250 or 500 µM. After 48 h of
incubation at 37°C, cell viability was determined by MTT assay as
described above. Glioblastoma cells without any treatment comprised
the mock control group.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
First, total RNA was extracted with the
RNeasy® Mini kit (Qiagen GmbH, Hilden, Germany)
following the manufacturer's protocol, and quantified by
fluorometry (Qubit® 2.0 Fluorometer Firmware 3.11;
Thermo Fisher Scientific, Inc.). The purity was considered
acceptable for RNA/protein ratios above 1.8. RNA integrity was
analyzed by 1% agarose gel electrophoresis using ethidium bromide
(Invitrogen™; Thermo Fisher Scientific, Inc.). Complementary (c)DNA
synthesis was performed from 500 ng total RNA using random primers
(SuperScript® First-Strand Synthesis System for RT-PCR;
Invitrogen; Thermo Fisher Scientific, Inc.). RT-qPCR was carried
out in a 7500 Fast Real-Time PCR System (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The forward and reverse primers for nNOS
were 5′-GGTGGAGATCAATATCGCGGTT-3′ and 5′-CCGGCAGCGGTACTCATTCT-3′,
respectively (24). For the
housekeeping gene, the GC-rich promoter binding protein 1 primers
5′-TCACTTGAGGCAGAACACAGA-3′ and 5′-AGCACATGTTTCATCATTTTCAC-3′ were
used (25). Amplification products
were detected via intercalation of the fluorescent dye
SYBR® Green (Applied Biosystems; Thermo Fisher
Scientific, Inc.). Briefly, 10 µl reaction mixture contained 5.0 µl
Fast SYBR® Green Master Mix (Applied Biosystems; Thermo
Fisher Scientific, Inc.), 2.0 µl cDNA (diluted 1:10), as previously
described (23), and 0.4 µl each
sense and antisense primer (10 pmol/µl). The PCR program included
an initial denaturation at 95°C for 5 min, followed by 40 cycles of
amplification (95°C for 1 min and 60°C for 1 min). Each experiment
was carried out in triplicate, and the assay included non-template
negative RT controls. The 2−ΔΔCq relative quantification
method was used to express the RNA interference (RNAi) effects on
nNOS mRNA content (26).
Nuclear staining with Hoechst
33342
Hoechst 33342 (Sigma-Aldrich; Merck KGaA) staining
was used to detect injured cells. Cells were cultured in 24-well
plates and transfected with siRNAnNOShum_4400 as above described.
After 24 h, cells received TMZ for 48 h. Injured and
non-transfected cell groups were also included as positive
controls. Briefly, the cells were fixed in 4.0% paraformaldehyde
for 10 min prior to staining with Hoechst 33342 (5.0 µg/ml/well) in
the dark for 10 min at room temperature. Subsequently, the
coverslips were washed twice with PBS, air-dried, mounted onto
glass slides and observed under a fluorescence microscope (TCS SP5;
Leica Microsystems, GmbH, Wetzlar, Germany). The nuclear
condensation, fragmentation and bright staining of damaged cells
were identified by intense local staining in the nucleus, in
contrast to the diffused staining of DNA in healthy cells (27).
Statistical analysis
SPSS version 17 (SPSS, Inc., Chicago, IL, USA) was
used to analyze the data. All results were expressed as means ±
standard error of the mean. One-way analysis of variance with
Tukey's post hoc test was applied to evaluate inter-group results.
Differences between paired groups were analyzed by the Student's
t-test. P<0.05 was considered to indicate a statistically
significant difference.
Results
TMZ affects astrocytoma and
glioblastoma cell viability, and increases oxidative stress
All cell lines exposed to TMZ (250 µM) presented a
decreased cell viability with higher levels of ROS. These effects
varied in intensity according to the time points and cell lines
studied. As shown in Fig. 1A, TMZ
caused a progressive decline in cell viability at 2, 48 and 72 h in
the ATCC U87MG cell line. The values varied from 92% (2 h) to 52%
(72 h), and they were significantly different between 2, 48 and 72
h time points. The cells produced ROS at increased levels, reaching
the maximum peak at 48 h (P<0.05). TMZ also affected cell
viability and oxidative stress in astrocytoma cells. On U251MG
cells, TMZ caused the highest cell damage at 48 h (Fig. 1B). Compared with ATCC U87MG and U138MG
cell lines, the differences in cell viability among time points 2,
48 and 72 h were small (84–88%) and not statistically significant,
as determined by the MTT assay. The highest oxidative stress
response in U251MG cells was observed at 72 h post-TMZ. Finally,
TMZ caused a progressive reduction in U138MG cell viability, with a
parallel increase in ROS production (P<0.05; Fig. 1C). These two effects peaked at 72 h
post-TMZ.
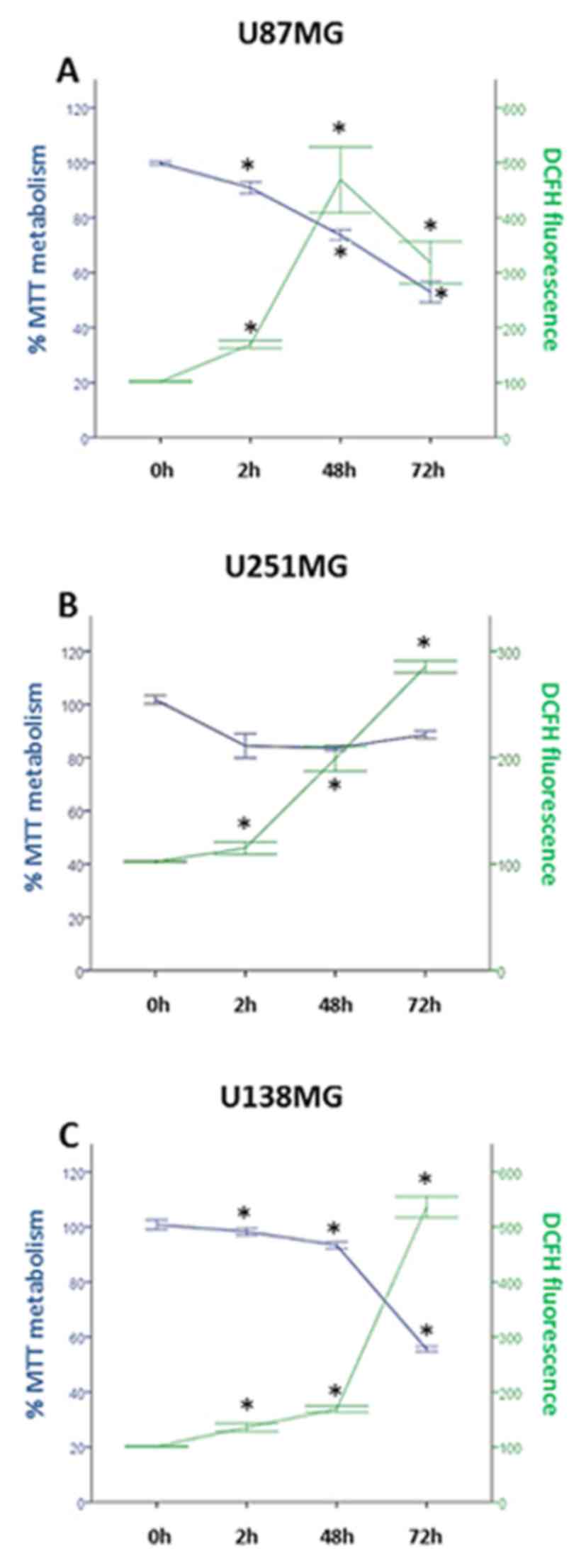 | Figure 1.TMZ reduces the viability of the (A)
American Type Culture Collection U87MG, (B) U251MG and (C) U138MG
cell lines, and increases their oxidative stress. Experiments were
carried out in triplicate in eight independent assays. Circles and
squares represent mean values ± standard error of the mean
regarding MTT and DCFH assay results, respectively, at each
time-point. Cells were injured by TMZ (250 µM) for 2, 48 or 72 h.
TMZ affected cell viability and the production of ROS, as
determined by MTT and DCFH assays, respectively. From 2 to 48 h
post-TMZ, all cell lines exhibited a significant reduction in cell
viability along with an increase in oxidative stress response.
Asterisks represent statistically significant differences
(*P<0.05, one-way analysis of variance followed by Tukey's post
hoc test). TMZ, temozolomide; DCFH, 2′,7′-dichlorofluorescein. |
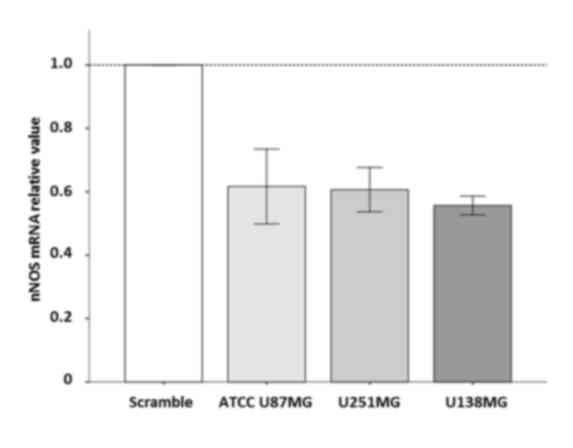
nNOS-targeted siRNA reduces nNOS mRNA
content in astrocytoma and glioblastoma cell lines
The present study evaluated whether the enzyme nNOS
could be silenced by RNAi. For that purpose, the effect of a
previously described siRNAnNOShum_4400 (37.5 nM) on nNOS mRNA
content was examined at 24 h after transfection. All transfected
cell lines (i.e. ATCC U87MG, U251MG and U138MG) presented their
nNOS mRNA content reduced to ~50% at 24 h (Fig. 2). This strategy used to suppress nNOS
expression was additionally employed in the present study to
examine the role of this enzyme in TMZ-injured glioblastoma
cells.
Effects of NOS inhibitors on the
viability and oxidative stress of ATCC U87MG cells exposed to TMZ
for 48 h
Inhibiting NOS enzymes causes effects on TMZ cell
damage at 48 h. As shown in Fig. 3A,
L-NAME and 7-NI decreased the viability of TMZ-damaged cells by 9.6
and 12.8%, respectively (P<0.05), compared to TMZ 250 group. In
parallel, these inhibitors also hampered the increase in ROS
production caused by TMZ (P<0.05; Fig.
3B), compared to TMZ 250 group. As noted, 7-NI was more
effective than L-NAME in improving the effects of TMZ, as well as
in controlling the oxidative stress response. These data highlight
a relevant role for nNOS in the actions of TMZ against glioblastoma
cells.
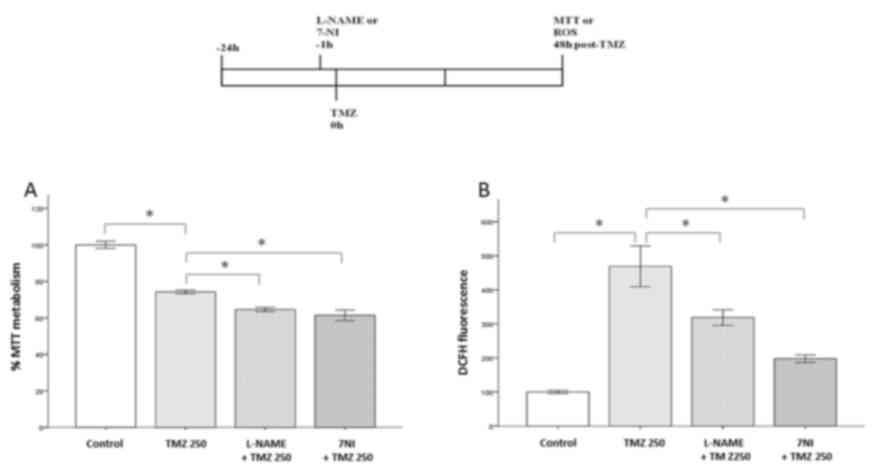 | Figure 3.Effects of NOS inhibition on American
Type Culture Collection U87MG cells exposed to TMZ for 48 h. Cell
viability and ROS production were measured by MTT and DCFH-DA
assays, respectively. Bars express mean ± standard error of the
mean regarding the results of MTT or DCFH-DA assays. Experiments
were carried out in triplicate in eight independent assays. A
schematic representation of the experimental procedure is included.
Briefly, either L-NAME or 7-NI was added to cell preparations 1 h
before TMZ treatment. Next, the cells received TMZ (250 µM) at the
time-point 0 h. The cells were then incubated for an additional
period of 48 h, prior to being subjected to the MTT and DCFH-DA
assays. Pretreatment with L-NAME (200 µM) or 7-NI (100 µM) in cells
injured by TMZ (A) reduced their viability and (B) decreased ROS
production. Asterisks represent statistically significant
differences (*P<0.05, one-way analysis of variance followed by
Tukey´s post hoc test). NOS, nitric oxide synthase; ROS, reactive
oxygen species; TMZ, temozolomide; DCFH-DA,
2′,7′-dichlorofluorescein-diacetate; L-NAME, N(ω)-nitro-L-arginine
methyl ester; 7-NI, 7-nitroindazole. |
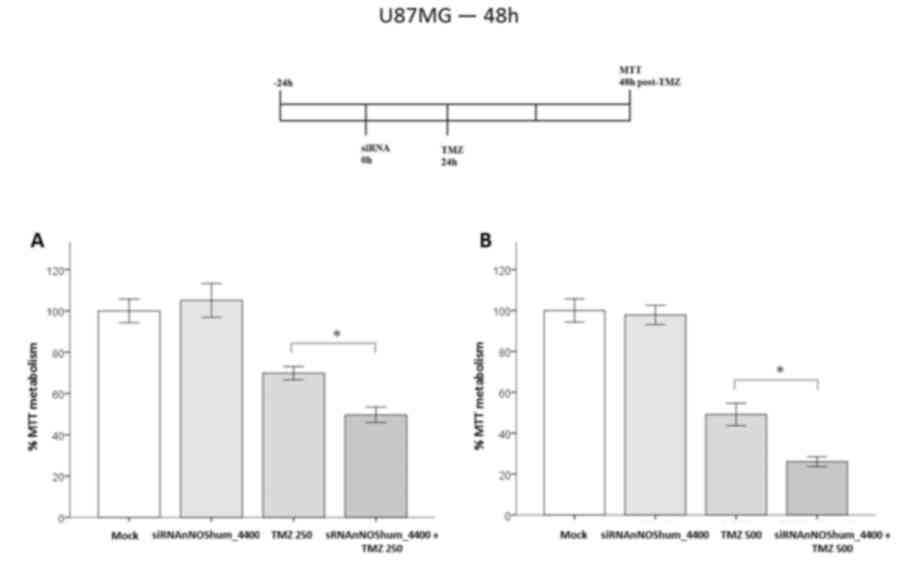
nNOS-targeted siRNA potentiates the
effects of TMZ
nNOS enzyme was knocked-down by using
siRNAnNOShum_4400, which was transfected 24 h before TMZ cell
damage. This siRNA alone caused no significant changes in cell
viability. However, siRNA-transfected cells were more susceptible
to the effects of TMZ at 48 h. They presented an additional 20%
decrease in cell viability compared with that of the injured
mock-transfected group (70 vs. 50%, respectively; P<0.05)
(Fig. 4A). Cells damaged by a higher
TMZ concentration (500 µM) also presented the same response (44 vs.
26%, respectively; P<0.05) (Fig.
4B).
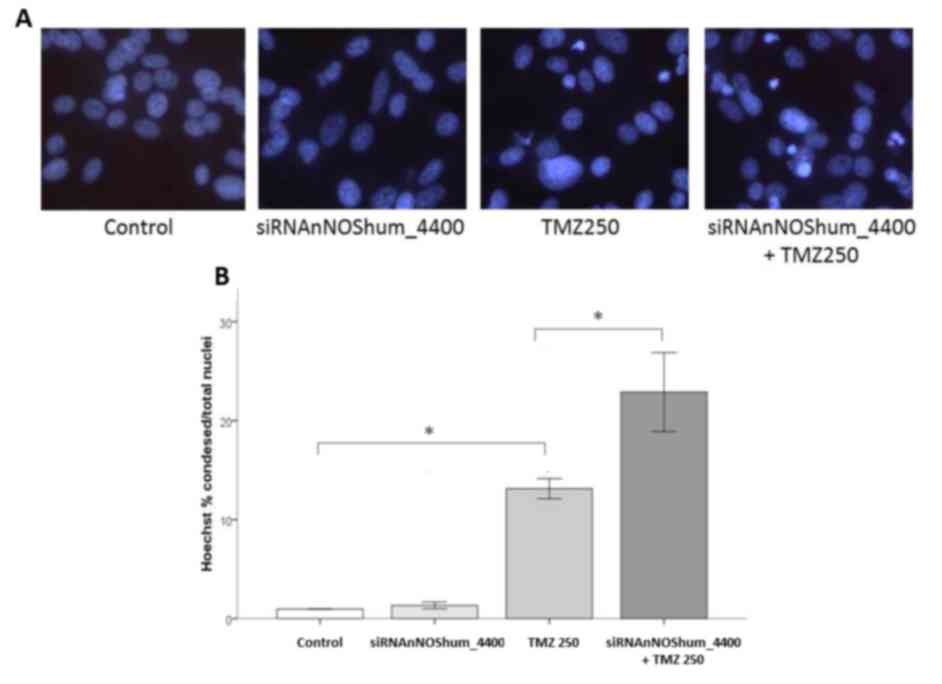
nNOS-targeted siRNA increases the
damage of cells caused by TMZ
The present study conducted a Hoechst 33342 analysis
of TMZ-injured cells. It was observed that TMZ (250 µM) alone
caused a marked damage on ATCC U87MG cells, as shown by the typical
nuclear Hoechst staining (Fig. 5A).
Silencing the nNOS enzyme with siRNAnNOShum_4400 caused no increase
in cell apoptosis. However, cells with silenced nNOS were more
susceptible to TMZ injury than control cells. As shown in Fig. 5B, the damage caused by TMZ was
significantly higher in the group transfected with nNOS-targeted
siRNA compared with the TMZ (250 µM) alone (P<0.05).
Discussion
Growing evidence has implicated NOS enzymes in the
biology of tumor cells (28–30). The present study examined whether the
nNOS enzyme affects astrocytoma and glioblastoma cell responses to
TMZ cell damage. TMZ acts by forming O6-methylguanine
nt, which mispairs with thymine during DNA replication. In
consequence, the drug causes cell cycle arrest in G2/M
phase in astrocytic tumor cells, which finally induces cell death
(31). TMZ also reduces the membrane
potential of the mitochondrion, releases cytochrome c from
this organelle, and increases activated caspases 3 and 9 cell
content (32). Oliva et al
(2011) suggested that U251MG cells demonstrated an increase in ROS
production at 2 and 4 h post-TMZ (33), which indicated that oxidative stress
contributed to the TMZ effects on astrocytoma tumor cells, as
confirmed by a protective effect obtained with the antioxidant
N-acetylcysteine. Indeed, TMZ at 300 µM caused a 10-fold increase
in ROS levels in TMZ-sensitive U251MG cells, but no effect occurred
on resistant cells (33). The present
study used TMZ at 250 µM. The drug increased oxidative stress in
ATCC U87MG, U251MG and U138MG cells. The peak of ROS concentration
occurred at 48 or 72 h, according to each cell line (Fig. 1). Corroborating previous studies, cell
groups with increased levels of ROS exhibited a lower cell
viability than those with reduced ROS levels (33,34).
In a recent study from 2016, Allen et al
(32) compared the original U87MG
cell line first identified at Uppsala University with the
commercial U87MG cell line used in the present study
(ATCC® HTB14™). Genotyping results based on short tandem
repeat (STR) analysis revealed that the ATCC and Uppsala U87MG
cells lines are from distinct origins (32). However, the STR profiling from the
ATCC U87MG cell line is identical to that previously published by
Bady et al in 2012 and the CLS Cell Line Services (35,36). In
addition, the commercial ATCC U87MG cell line was identified as a
cell line of central nervous system (CNS) tumor origin (Allen et
al 2016), by comparing the transcriptional profiles of ATCC
U87MG with those of 1,036 cell lines regarding 18 different tissue
derivation lineages of the Cancer Cell Line Encyclopedia (CCLE)
(37). In summary, ATCC U87MG can be
classified as a bonafide human glioblastoma cell line (35).
The World Health Organization recently updated the
classification of tumors of the CNS, including molecular parameters
in addition to conventional histology (38). With this regard, all three lineages
used in the present study are classified as astrocytic tumor cells,
termed astrocytoma (U251MG and U138MG) or glioblastoma (ATCC
U87MG). Glioblastoma is the most malignant astrocytic tumor (grade
IV), with the following histopathology features: Cellular
polymorphism, nuclear atypia, high mitotic activity, increased
abnormal growth of blood vessels around the tumor, vascular
thrombosis, microvascular proliferation and necrosis (1,2,39).
Although no animal model perfectly represents all
aspects of human glioblastomas, the model of choice must mimic the
features of the disease under investigation (40,41). The
lineage used in our study, i.e. ATCC U87MG, exhibits numerous
aspects of this primary brain tumor in rodent models. Intracerebral
implantation of ATCC U87MG cells in nude mice resulted in the
growth of tumors whose volume increased by ~50 mm3 in 39
days, presenting an infiltrative pattern with a marked
neovascularization and large necrotic center areas (42,43), which
indicates that the model has ‘face’ validity. The ATCC U87MG cell
line has also been used for refining diagnostic techniques on
histopathology and imaging of glioblastoma tumors (44–46).
Regarding the ‘predictive’ validity of the model, ATCC U87MG tumors
can be treated with TMZ, the drug of choice for GBM, which was
employed in the present study (47,48).
Besides TMZ, the ATCC U87MG cell line has also been used for
pre-clinical testing of different therapeutic agents against
glioblastoma (49–53). In summary, the scientific data
available suggest that the ATCC U87MG cell lineage provides a
glioblastoma model with invasiveness, tumor-induced necrosis and
vascular alterations that mimics human glioblastomas, indicating
that it is a useful experimental model for studies on tumor biology
and therapeutics. Thus, the genetic differences between the ATCC
U87MG cell line and the original Uppsala U87MG lineage are unlikely
to affect the conclusions of the present study.
The gas NO contributes to oxidative stress in
astrocytic tumors (11). NO is often
referred to as a toxic and reactive molecule. However, it inhibits
the apoptosis of cells associated with caspase 3-like enzymes via
S-nitrosylation or cyclic guanosine monophosphate-dependent
pathways (54). At mM concentrations,
it also inhibits catalase and cytochrome P-450 enzymes (55). NO is released during the synthesis of
L-citrulline, which is catalyzed by NOS enzymes (12). This gaseous molecule will form two
metabolites: Nitrite and nitrate (13). Nitrite damages DNA strands, leading to
various biochemical features observed in cancer cells such as
changes in p53 activity and epidermal growth factor signaling
(56–58). The increased NOS activity reported in
glioma cells also contributes to oxidative stress (11). Similarly, inhibiting NOS enzymes by
L-NAME resulted in a lower nitrite concentration in rat glioma
tumor tissues with a decrease in tumor volume, which reinforces the
benefits of reducing NOS activity in glioma cells (19,59). No
previous study, however, has addressed the involvement of NOS
enzymes in the damage of astrocytic tumor cells cells (astrocytoma
and glioblastoma) caused by TMZ. In the present study, cells were
pretreated with L-NAME to evaluate the effects of this drug on
oxidative stress and viability. It was observed that NOS enzymes
affect both responses. L-NAME sensitized ATCC U87MG cells to 250 µM
of TMZ. The viability of glioblastoma cells and their oxidative
stress levels were decreased at 48 h. Altogether, these data reveal
that NOS enzymes contribute to oxidative stress in glioblastoma
cells injured by TMZ. It is possible to speculate that inhibition
of NOS enzymes during TMZ cell damage may converge to the same
apoptotic pathway described above, thus improving the damage of
glioblastoma cells.
nNOS is highly expressed in astrocytic tumors. The
expression of nNOS occurs in both the tumor and peritumoral areas
of brain tumors (16). Indeed, tumors
with high histological grades also present increased levels of nNOS
expression (18,60). Although these findings suggest a
possible role for nNOS in gliomagenesis, no previous study has
addressed its involvement in astrocytic tumor cells injured by TMZ.
The present study evaluated the effects of the nNOS inhibitor 7-NI
and a synthetic siRNA targeting nNOS. At 48 h, 7-NI decreased the
viability of ATCC U87MG cells, restraining their oxidative stress
response to 250 µM of TMZ (Fig. 3).
The role of the nNOS enzyme in glioblastoma cell responses to TMZ
was confirmed by RNAi experiments. ATCC U87MG cells subjected to
nNOS silencing for 24 h were more vulnerable to TMZ compared to
cells exposed to TMZ alone. They exhibited a decreased viability
with an increased rate of injury (Figs.
4 and 5). The results from nNOS
knock-down experiments reinforce those obtained with the nNOS
inhibitor 7-NI, suggesting that this enzyme serves at least a
partial role in glioblastoma cell defenses against TMZ.
Understanding the biology of astrocytic cells is a
challenging effort for studies that use animal models of brain
tumors (61–63). Beyond modeling tumor pathogenesis, the
results from animal models have also identified potential targets
for glioma chemotherapy (64–67). The present study revealed that
suppressing the nNOS enzyme improves the effects of TMZ on
glioblastoma cells. siRNAs designed to silence other enzymes also
increase TMZ injury in astrocytic cells, as observed for DNA
methyltransferases and kinases (68–70). RNAi
with non-enzymatic targets was also observed to be valuable in
sensitizing cells to TMZ: In a previous study, it was noted that
silencing the voltage-gated potassium channel Eag1 makes
glioblastoma cells more vulnerable to TMZ (71). The same effect occurred for drug
resistance proteins, heat-shock proteins 90, 27 and 72, and other
targets involved in cell signaling (72–77).
The studies mentioned above stand that siRNAs hold
the potential to be RNAi-based drugs associated with TMZ. However,
exploitation of RNAi in the clinic will depend on improvements in
oligonucleotides chemical structure for stability
(phosphorothioated backbones that avoid ribonuclease attack) and
specificity (i.e. locked nucleic acids), as well as the development
of novel carriers for siRNA delivery (78). The oligonucleotide siRNAnNOShum_4400
used in the present study has no phosphorothioate backbones and
locked nucleic acids (23). Thus, we
recommend adopting these chemical changes in future studies with
nNOS-targeted siRNAs for improvements in the specificity and
duration of silencing effects.
A significant limitation to treat brain tumors is
the location where such tumors grow, i.e. the CNS, since the
blood-brain barrier offers an obstacle for drug distribution
(79). The introduction of siRNAs
directly into brain tumor tissues by the convection-enhanced
delivery (CED) technique is a viable alternative to circumvent this
obstacle (80). Combining CED with
the use of nanoparticles for carrying siRNAs is a promising
strategy to treat glioma tumors by RNAi (81–83).
Finally, a sustained delivery of siRNAs implanted in tumor tissues
would be of value to control aggressive cancer types. In a previous
study, biodegradable poly (lactic-co-glycolic acid) devices
implanted into pancreatic tumors provided a 4-month delivery of
siRNAs with significant antitumor results (84). Implantation of such delivery system
following brain tumors ablation would be a promising strategy to
prevent tumor recurrence.
Regarding clinical trials, at least ten RNAi-based
drugs are currently in phase II and will become approved treatments
in the following years (78). Of
these, three clinical trials address cancer diseases, including
pancreatic tumors, hepatocellular carcinomas and neuroendocrine
tumors (78). It may be possible that
certain RNAi-based drugs successfully tested in animal models of
brain tumors could be evaluated in clinical trials.
To conclude, the present study revealed a new target
for RNAi, the nNOS enzyme, which also improved the anticancer
effects of TMZ on glioblastoma cells. The synthetic duplex for
silencing nNOS, siRNAnNOShum_4400, merits further studies to
explore its potential use for brain tumor anticancer therapy.
Acknowledgements
The present study received financial support from
CAPES [Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior (Programa Nacional de Pós-doutorado; grant no.,
3731–37/2010)], CNPq [Conselho Nacional de Desenvolvimento
Científico e Tecnológico (grant no., 467467/2014-5)] and FAP-DF
[Fundação de Apoio à Pesquisa do Distrito Federal (grant
no., 2010/00302-9)]. The nNOS sequence used for designing
siRNAnNOShum_4400 is under patent registration at the Brazilian
Institute for Industrial Property-Instituto Nacional da Propriedade
Industrial (patent no., INPI, BR 10 2012 032844 5).
References
|
1
|
Lassman AB: Molecular biology of gliomas.
Curr Neurol Neurosci Rep. 4:228–233. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maher EA, Furnari FB, Bachoo RM, Rowitch
DH, Louis DN, Cavenee WK and DePinho RA: Malignant glioma: Genetics
and biology of a grave matter. Genes Dev. 15:1311–1333. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Henson JW: Treatment of glioblastoma
multiforme: A new standard. Arch Neurol. 63:337–341. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ostrom QT, Gittleman H, Farah P, Ondracek
A, Chen Y, Wolinsky Y, Stroup NE, Kruchko C and Barnholtz-Sloan JS:
CBTRUS statistical report: Primary brain and central nervous system
tumors diagnosed in the United States in 2006–2010. Neuro Oncol. 15
Suppl 2:ii1–ii56. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stupp R, Hegi ME, Gilbert MR and
Chakravarti A: Chemoradiotherapy in malignant glioma: Standard of
care and future directions. J Clin Oncol. 25:4127–4136. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pedretti M, Verpelli C, Mårlind J, Bertani
G, Sala C, Neri D and Bello L: Combination of temozolomide with
immunocytokine F16-IL2 for the treatment of glioblastoma. Br J
Cancer. 103:827–836. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sathornsumetee S and Rich JN: New
treatment strategies for malignant gliomas. Expert Rev Anticancer
Ther. 6:1087–1104. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cairns RA, Harris IS and Mak TW:
Regulation of cancer cell metabolism. Nat Rev Cancer. 11:85–95.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kardeh S, Ashkani-Esfahani S and Alizadeh
AM: Paradoxical action of reactive oxygen species in creation and
therapy of cancer. Eur J Pharmacol. 735:150–168. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nogueira V and Hay N: Molecular pathways:
Reactive oxygen species homeostasis in cancer cells and
implications for cancer therapy. Clin Cancer Res. 19:4309–4314.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Conti A, Guli C, La Torre D, Tomasello C,
Angileri FF and Aguennouz M: Role of inflammation and oxidative
stress mediators in gliomas. Cancers (Basel). 2:693–712. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim SH, Kwon CH and Nakano I:
Detoxification of oxidative stress in glioma stem cells: Mechanism,
clinical relevance, and therapeutic development. J Neurosci Res.
92:1419–1424. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Moncada S and Bolaños JP: Nitric oxide,
cell bioenergetics and neurodegeneration. J Neurochem.
97:1676–1689. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luo CX and Zhu DY: Research progress on
neurobiology of neuronal nitric oxide synthase. Neurosci Bull.
27:23–35. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Thomsen LL and Miles DW: Role of nitric
oxide in tumour progression: Lessons from human tumours. Cancer
Metastasis Rev. 17:107–118. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bakshi A, Nag TC, Wadhwa S, Mahapatra AK
and Sarkar C: The expression of nitric oxide synthases in human
brain tumours and peritumoral areas. J Neurol Sci. 155:196–203.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fukumura D and Jain RK: Role of nitric
oxide in angiogenesis and microcirculation in tumors. Cancer
Metastasis Rev. 17:77–89. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tanriover N, Ulu MO, Isler C, Durak H, Oz
B, Uzan M and Akar Z: Neuronal nitric oxide synthase expression in
glial tumors: Correlation with malignancy and tumor proliferation.
Neurol Res. 30:940–944. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Swaroop GR, Kelly PA, Bell HS, Shinoda J,
Yamaguchi S and Whittle IR: The effects of chronic nitric oxide
synthase suppression on glioma pathophysiology. Br J Neurosurg.
14:543–548. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Roche AK, Cook M, Wilcox GL and Kajander
KC: A nitric oxide synthesis inhibitor (L-NAME) reduces licking
behavior and Fos-labeling in the spinal cord of rats during
formalin-induced inflammation. Pain. 66:331–341. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Southan GJ and Szabó C: Selective
pharmacological inhibition of distinct nitric oxide synthase
isoforms. Biochem Pharmacol. 51:383–394. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huesken D, Lange J, Mickanin C, Weiler J,
Asselbergs F, Warner J, Meloon B, Engel S, Rosenberg A, Cohen D, et
al: Design of a genome-wide siRNA library using an artificial
neural network. Nat Biotechnol. 23:995–1001. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Titze-de-Almeida SS, Lustosa CF, Horst CH,
Bel ED and Titze-de-Almeida R: Interferon Gamma potentiates the
injury caused by MPP(+) on SH-SY5Y cells, which is attenuated by
the nitric oxide synthases inhibition. Neurochem Res. 39:2452–2464.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dotsch J, Harmjanz A, Christiansen H,
Hänze J, Lampert F and Rascher W: Gene expression of neuronal
nitric oxide synthase and adrenomedullin in human neuroblastoma
using real-time PCR. Int J Cancer. 88:172–175. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kwon MJ, Oh E, Lee S, Roh MR, Kim SE, Lee
Y, Choi YL, In YH, Park T, Koh SS and Shin YK: Identification of
novel reference genes using multiplatform expression data and their
validation for quantitative gene expression analysis. PLoS One.
4:e61622009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sandhu LC, Warters RL and Dethlefsen LA:
Fluorescence studies of Hoechst 33342 with supercoiled and relaxed
plasmid pBR322 DNA. Cytometry. 6:191–194. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jia W, Jackson-Cook C and Graf MR:
Tumor-infiltrating, myeloid-derived suppressor cells inhibit T cell
activity by nitric oxide production in an intracranial rat glioma +
vaccination model. J Neuroimmunol. 223:20–30. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Muntané J and La Mata MD: Nitric oxide and
cancer. World J Hepatol. 2:337–344. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sikora AG, Gelbard A, Davies MA, Sano D,
Ekmekcioglu S, Kwon J, Hailemichael Y, Jayaraman P, Myers JN, Grimm
EA and Overwijk WW: Targeted inhibition of inducible nitric oxide
synthase inhibits growth of human melanoma in vivo and synergizes
with chemotherapy. Clin Cancer Res. 16:1834–1844. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sang DP, Li RJ and Lan Q: Quercetin
sensitizes human glioblastoma cells to temozolomide in vitro via
inhibition of Hsp27. Acta Pharmacol Sin. 35:832–838. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jakubowicz-Gil J, Langner E, Badziul D,
Wertel I and Rzeski W: Apoptosis induction in human glioblastoma
multiforme T98G cells upon temozolomide and quercetin treatment.
Tumour Biol. 34:2367–2378. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Oliva CR, Moellering DR, Gillespie GY and
Griguer CE: Acquisition of chemoresistance in gliomas is associated
with increased mitochondrial coupling and decreased ROS production.
PLoS One. 6:e246652011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang WB, Wang Z, Shu F, Jin YH, Liu HY,
Wang QJ and Yang Y: Activation of AMP-activated protein kinase by
temozolomide contributes to apoptosis in glioblastoma cells via p53
activation and mTORC1 inhibition. J Biol Chem. 285:40461–40471.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Allen M, Bjerke M, Edlund H, Nelander S
and Westermark B: Origin of the U87MG glioma cell line: Good news
and bad news. Sci Transl Med. 8:354re32016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bady P, Diserens AC, Castella V, Kalt S,
Heinimann K, Hamou MF, Delorenzi M and Hegi ME: DNA fingerprinting
of glioma cell lines and considerations on similarity measurements.
Neuro Oncol. 14:701–711. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Barretina J, Caponigro G, Stransky N,
Venkatesan K, Margolin AA, Kim S, Wilson CJ, Lehár J, Kryukov GV,
Sonkin D, et al: The cancer cell line encyclopedia enables
predictive modelling of anticancer drug sensitivity. Nature.
483:603–607. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Louis DN, Perry A, Reifenberger G, von
Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD,
Kleihues P and Ellison DW: The 2016 world health organization
classification of tumors of the central nervous system: A summary.
Acta Neuropathol. 131:803–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Reni M, Mazza E, Zanon S, Gatta G and
Vecht CJ: Central nervous system gliomas. Crit Rev Oncol Hematol.
113:213–234. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Goldbrunner RH, Wagner S, Roosen K and
Tonn JC: Models for assessment of angiogenesis in gliomas. J
Neurooncol. 50:53–62. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Stylli SS, Luwor RB, Ware TM, Tan F and
Kaye AH: Mouse models of glioma. J Clin Neurosci. 22:619–626. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cheng SY, Huang HJ, Nagane M, Ji XD, Wang
D, Shih CC, Arap W, Huang CM and Cavenee WK: Suppression of
glioblastoma angiogenicity and tumorigenicity by inhibition of
endogenous expression of vascular endothelial growth factor. Proc
Natl Acad Sci USA. 93:pp. 8502–8507. 1996; View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Doblas S, He T, Saunders D, Pearson J,
Hoyle J, Smith N, Lerner M and Towner RA: Glioma morphology and
tumor-induced vascular alterations revealed in seven rodent glioma
models by in vivo magnetic resonance imaging and angiography. J
Magn Reson Imaging. 32:267–275. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kirschner S, Murle B, Felix M, Arns A,
Groden C, Wenz F, Hug A, Glatting G, Kramer M, Giordano FA and
Brockmann MA: Imaging of orthotopic glioblastoma xenografts in mice
using a clinical CT scanner: Comparison with Micro-CT and
histology. PLoS One. 11:e01659942016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu X, Dong C, Shi J, Ma T, Jin Z, Jia B,
Liu Z, Shen L and Wang F: Radiolabeled novel mAb 4G1 for
immunoSPECT imaging of EGFRvIII expression in preclinical
glioblastoma xenografts. Oncotarget. 8:6364–6375. 2017.PubMed/NCBI
|
|
46
|
Rogers S, Hii H, Huang J, Ancliffe M,
Gottardo NG, Dallas P, Lee S and Endersby R: A novel technique of
serial biopsy in mouse brain tumour models. PLoS One.
12:e01751692017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Arcella A, Oliva MA, Staffieri S, Aalberti
S, Grillea G, Madonna M, Bartolo M, Pavone L, Giangaspero F,
Cantore G and Frati A: In vitro and in vivo effect of human
lactoferrin on glioblastoma growth. J Neurosurg. 123:1026–1035.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Nitta Y, Shimizu S, Shishido-Hara Y,
Suzuki K, Shiokawa Y and Nagane M: Nimotuzumab enhances
temozolomide-induced growth suppression of glioma cells expressing
mutant EGFR in vivo. Cancer Med. 5:486–499. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gromeier M, Lachmann S, Rosenfeld MR,
Gutin PH and Wimmer E: Intergeneric poliovirus recombinants for the
treatment of malignant glioma. Proc Natl Acad Sci USA. 97:pp.
6803–6808. 2000; View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kang KB, Wang TT, Woon CT, Cheah ES, Moore
XL, Zhu C and Wong MC: Enhancement of glioblastoma radioresponse by
a selective COX-2 inhibitor celecoxib: Inhibition of tumor
angiogenesis with extensive tumor necrosis. Int J Radiat Oncol Biol
Phys. 67:888–896. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jin J, Choi SH, Lee JE, Joo JD, Han JH,
Park SY and Kim CY: Antitumor activity of 7-O-succinyl macrolactin
A tromethamine salt in the mouse glioma model. Oncol Lett.
13:3767–3773. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gravina GL, Mancini A, Marampon F,
Colapietro A, Delle Monache S, Sferra R, Vitale F, Richardson PJ,
Patient L, Burbidge S and Festuccia C: The brain-penetrating CXCR4
antagonist, PRX177561, increases the antitumor effects of
bevacizumab and sunitinib in preclinical models of human
glioblastoma. J Hematol Oncol. 10:52017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhong X, Zhao H, Liang S, Zhou D, Zhang W
and Yuan L: Gene delivery of apoptin-derived peptide using an
adeno-associated virus vector inhibits glioma and prolongs animal
survival. Biochem Biophys Res Commun. 482:506–513. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Blaise GA, Gauvin D, Gangal M and Authier
S: Nitric oxide, cell signaling and cell death. Toxicology.
208:177–192. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Brunelli L, Yermilov V and Beckman JS:
Modulation of catalase peroxidatic and catalatic activity by nitric
oxide. Free Radic Biol Med. 30:709–714. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cobbs CS, Whisenhunt TR, Wesemann DR,
Harkins LE, Van Meir EG and Samanta M: Inactivation of wild-type
p53 protein function by reactive oxygen and nitrogen species in
malignant glioma cells. Cancer Res. 63:8670–8673. 2003.PubMed/NCBI
|
|
57
|
Xu W, Liu LZ, Loizidou M, Ahmed M and
Charles IG: The role of nitric oxide in cancer. Cell Res.
12:311–320. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang P, Wang YZ, Kagan E and Bonner JC:
Peroxynitrite targets the epidermal growth factor receptor, Raf-1,
and MEK independently to activate MAPK. J Biol Chem.
275:22479–22486. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Oyoshi T, Nomoto M, Hirano H and Kuratsu
J: Pathodynamics of nitric oxide production within implanted glioma
studied with an in vivo microdialysis technique and
immunohistochemistry. J Pharmacol Sci. 91:15–22. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Broholm H, Rubin I, Kruse A, Braendstrup
O, Schmidt K, Skriver EB and Lauritzen M: Nitric oxide synthase
expression and enzymatic activity in human brain tumors. Clin
Neuropathol. 22:273–281. 2003.PubMed/NCBI
|
|
61
|
Agnihotri S, Burrell KE, Wolf A, Jalali S,
Hawkins C, Rutka JT and Zadeh G: Glioblastoma, a brief review of
history, molecular genetics, animal models and novel therapeutic
strategies. Arch Immunol Ther Exp (Warsz). 61:25–41. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Lenting K, Verhaak R, Ter Laan M,
Wesseling P and Leenders W: Glioma: Experimental models and
reality. Acta Neuropathol. 133:263–282. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Resende FF, Bai X, Del Bel EA, Kirchhoff
F, Scheller A and Titze-de-Almeida R: Evaluation of
TgH(CX3CR1-EGFP) mice implanted with mCherry-GL261 cells as an in
vivo model for morphometrical analysis of glioma-microglia
interaction. BMC Cancer. 16:722016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Chen J, McKay RM and Parada LF: Malignant
glioma: Lessons from genomics, mouse models, and stem cells. Cell.
149:36–47. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Cloughesy TF, Cavenee WK and Mischel PS:
Glioblastoma: From molecular pathology to targeted treatment. Annu
Rev Pathol. 9:1–25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kegelman TP, Hu B, Emdad L, Das SK, Sarkar
D and Fisher PB: In vivo modeling of malignant glioma: The road to
effective therapy. Adv Cancer Res. 121:261–330. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Wang Y and Jiang T: Understanding high
grade glioma: Molecular mechanism, therapy and comprehensive
management. Cancer Lett. 331:139–146. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kato T, Natsume A, Toda H, Iwamizu H,
Sugita T, Hachisu R, Watanabe R, Yuki K, Motomura K, Bankiewicz K
and Wakabayashi T: Efficient delivery of liposome-mediated
MGMT-siRNA reinforces the cytotoxity of temozolomide in
GBM-initiating cells. Gene Ther. 17:1363–1371. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Shervington A and Patel R: Silencing DNA
methyltransferase (DNMT) enhances glioma chemosensitivity.
Oligonucleotides. 18:365–374. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wen X, Huang A, Liu Z, Liu Y, Hu J, Liu J
and Shuai X: Downregulation of ROCK2 through nanocomplex sensitizes
the cytotoxic effect of temozolomide in U251 glioma cells. PLoS
One. 9:e920502014. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Sales TT, Resende FF, Chaves NL,
Titze-De-Almeida SS, Báo SN, Brettas ML and Titze-De-Almeida R:
Suppression of the Eag1 potassium channel sensitizes glioblastoma
cells to injury caused by temozolomide. Oncol Lett. 12:2581–2589.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Cruickshanks N, Shervington L, Patel R,
Munje C, Thakkar D and Shervington A: Can hsp90alpha-targeted siRNA
combined with TMZ be a future therapy for glioma? Cancer Invest.
28:608–614. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Jakubowicz-Gil J, Langner E, Badziul D,
Wertel I and Rzeski W: Silencing of Hsp27 and Hsp72 in glioma cells
as a tool for programmed cell death induction upon temozolomide and
quercetin treatment. Toxicol Appl Pharmacol. 273:580–589. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Paul-Samojedny M, Pudelko A, Kowalczyk M,
Fila-Daniłow A, Suchanek-Raif R, Borkowska P and Kowalski J:
Combination therapy with AKT3 and PI3KCA siRNA enhances the
antitumor effect of temozolomide and carmustine in T98G
glioblastoma multiforme cells. BioDrugs. 30:129–144. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Qian C, Li P, Yan W, Shi L, Zhang J, Wang
Y, Liu H and You Y: Downregulation of osteopontin enhances the
sensitivity of glioma U251 cells to temozolomide and cisplatin by
targeting the NF-κB/Bcl-2 pathway. Mol Med Rep. 11:1951–1955. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Tivnan A, Zakaria Z, O'Leary C, Kögel D,
Pokorny JL, Sarkaria JN and Prehn JH: Inhibition of multidrug
resistance protein 1 (MRP1) improves chemotherapy drug response in
primary and recurrent glioblastoma multiforme. Front Neurosci.
9:2182015. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Wang Q, Du J, Xu B, Xu L, Wang X, Liu J
and Wang J: Silence of bFGF enhances chemosensitivity of glioma
cells to temozolomide through the MAPK signal pathway. Acta Biochim
Biophys Sin (Shanghai). 48:501–508. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Titze-de-Almeida R, David C and
Titze-de-Almeida SS: The race of 10 synthetic RNAi-based drugs to
the pharmaceutical market. Pharm Res. 34:1339–1363. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
de Boer AG and Gaillard PJ: Drug targeting
to the brain. Annu Rev Pharmacol Toxicol. 47:323–355. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Lonser RR, Sarntinoranont M, Morrison PF
and Oldfield EH: Convection-enhanced delivery to the central
nervous system. J Neurosurg. 122:697–706. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Cohen ZR, Ramishetti S, Peshes-Yaloz N,
Goldsmith M, Wohl A, Zibly Z and Peer D: Localized RNAi
therapeutics of chemoresistant grade IV glioma using
hyaluronan-grafted lipid-based nanoparticles. ACS Nano.
9:1581–1591. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Danhier F, Messaoudi K, Lemaire L, Benoit
JP and Lagarce F: Combined anti-Galectin-1 and anti-EGFR
siRNA-loaded chitosan-lipid nanocapsules decrease temozolomide
resistance in glioblastoma: In vivo evaluation. Int J Pharm.
481:154–161. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Tsujiuchi T, Natsume A, Motomura K, Kondo
G, Ranjit M, Hachisu R, Sugimura I, Tomita S, Takehara I, Woolley
M, et al: Preclinical evaluation of an O(6)-methylguanine-DNA
methyltransferase-siRNA/liposome complex administered by
convection-enhanced delivery to rat and porcine brains. Am J Transl
Res. 6:169–178. 2014.PubMed/NCBI
|
|
84
|
Golan T, Khvalevsky EZ, Hubert A, Gabai
RM, Hen N, Segal A, Domb A, Harari G, David EB, Raskin S, et al:
RNAi therapy targeting KRAS in combination with chemotherapy for
locally advanced pancreatic cancer patients. Oncotarget.
6:24560–24570. 2015. View Article : Google Scholar : PubMed/NCBI
|