Introduction
Pituitary adenomas are one of the most common
intracranial tumors; having the third highest morbidity rate of all
intracranial tumors following brain gliomas and meningiomas
(1). Pituitary adenomas are divided
into different classifications through tumor size and hormone
secretion function (2). According to
tumor size, pituitary adenomas are divided into microadenomas and
macroadenomas by an arbitrary cutoff size of 10 mm (2). According to hormone secretion, the
tumors are divided into hormone-secreting pituitary adenomas and
nonfunctional adenomas (2). Although
the majority of pituitary adenomas are benign and rarely progress
to malignancies, they are usually aggressive due to hormone
excessive or insufficiency (1). Due
to the partial pressure of the tumor and disordered pituitary
hormone secretion, pituitary adenomas are characterized by a series
of clinical symptoms which seriously affect the quality of life of
the patients. The aim of the present study was to identify
objective molecular markers to predict the aggressive potential of
tumors, and guide strategies for surgery and medical treatment.
In our previous study, gene arrays were used to
screen for differentially expressed genes in aggressive pituitary
adenomas and a series of upregulated genes, including galectin-3
(Gal-3) were identified, indicating that Gal-3 may be involved in
the invasive development of pituitary adenomas (3,4). Gal-3, a
β-galactoside-binding protein, is one of the most frequently
investigated members of the galectin family. Gal-3 is involved in
normal biological processes including cell growth, differentiation
and cell adhesion, but is also involved in the progression of a
number of types of cancer, including pituitary, thyroid, colon and
breast cancer (5–8). However, the mechanisms underlying the
involvement of Gal-3 in the tumorigenesis and development of
pituitary adenomas remain unknown. In the present study, Gal-3
expression was detected in three different rat pituitary tumor cell
lines (GH3, GH4C1 and RC-4B/C). Expression and knockdown vectors of
Gal-3 were utilized to investigate the involvement of Gal-3 in the
tumorigenesis and progression of pituitary tumors, and provide a
theoretical basis for treatment of pituitary tumors.
Materials and methods
Cell culture
Three kinds of rat pituitary tumor cell lines GH3
was obtained from BeNa Culture Collection (Kunshan, China), and
GH4C1 and RC-4B/C were obtained from American Type Culture
Collection (ATCC; Manassas, VA, USA). Cells were maintained in
Dulbecco's modified Eagle's medium (DMEM; Gibco, Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 15% horse
serum (Gibco, Thermo Fisher Scientific, Inc.), 2.5% fetal bovine
serum (FBS; Gibco, Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 100 µg/ml streptomycin, and were maintained at 37°C
in a 5% CO2 humidified environment.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis of Gal-3
Total cellular RNA was extracted from pituitary
tumor cells using TRIzol reagent (Omega Bio-Tek, Inc., Norcross,
GA, USA) according to the manufacturer's instructions. Then, RNA
was reverse-transcribed into cDNA using PrimeScript™ RT
reagent kit with gDNA Eraser, and cDNA amplification was performed
using a SYBR Green PCR assay (both from Takara Bio, Inc., Dalian,
Japan). The transcriptional levels of genes were calculated using
the 2−ΔΔCq method (9). The
PCR reaction (20 µl total volume) contained 10 µl 2X SYBR Premix Ex
Taq™ (Takara Bio, Inc., Otsu, Japan), 0.40 µmol/l each
primer and 0.2±0.02 µg cDNA template. The following three-step
RT-qPCR reaction was performed: Pre-denaturation at 95°C for 30
sec, followed by 40 cycles of denaturation at 95°C for 5 sec,
annealing at 56–60°C for 20 sec and elongation at 72°C for 20 sec.
The sequences for the primers are as follows: Gal-3 sense,
5′-CCCCGCTTCAATGAGA-3′ and antisense, 5′-GAATGGTTTGCCGCTC-3′;
β-actin sense, 5′-CGTTGACATCCGTAAAGAC-3′ and antisense,
5′-TAGGAGCCAGGGCAGTA-3′.
Construction of expression and
interference vectors and cell transfection
The Gal-3 gene was amplified using primers (Gal-3
forward, 5′-ATAGGATCCATGGCAGACGGCTTCTCACTT-3′ and reverse,
5′-ATCGAAGACTTAGATCATGGCGTGGGAAG-3′) and was inserted into the
corresponding site of the pGPU6/GFP/Neo vector (GenePharma Co.,
Ltd., Shanghai, China) according to the manufacturer's instructions
to construct the overexpression vector pGPU6/GFP/Neo-Gal-3-Homo-1.
Negative control pGPU6/GFP/Neo-shNC (target sequence,
5′-GTTCTCCGAACGTGTCACGT-3′), and Gal-3 interference vectors
pGPU6/GFP/Neo-shGal-3-Homo-A (target sequence,
5′-AGCCTTCCCAGGGCAACCTGG-3′) were purchased from GenePharma Co.,
Ltd. Cells were seeded in cell plates with DMEM and cultured until
the cells reached 70–80% confluence, and then were transfected with
800 ng/well expression plasmids or interference plasmids using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. The untransfected
cells served as the control group.
MTT assay
Cell proliferation was measured using an MTT assay.
GH3 and GH4C1 cells were seeded in 96-well plates and transfected
with interference vectors, expression vectors or empty control
vectors. At the indicated time-points following transfection (24,
48 and 72 h), 20 µl MTT (5 mg/ml; Bioswamp; Wuhan Beinglay
Biological Technology Co., Wuhan, China) was added to each well.
Following incubation for 4 h, the medium was removed and 150 µl
DMSO was added to each well. Following shaking at low speed for 10
min, the absorbance of converted the dye was measured at a
wavelength of 490 nm.
Cell apoptosis assay
Apoptosis analysis was performed using Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) flow
cytometry kit (BD Biosciences, Franklin Lakes, CA, USA) according
to the manufacturer's instructions. GH3 and GH4C1 cells were
transfected with Gal-3 interference vectors, empty vectors or
expression vectors using Lipofectamine 2000. Cells were washed 48 h
after this three times with ice-cold PBS and resuspended in 200 µl
of binding buffer at a concentration of 1×106 cells/ml.
Annexin V-FITC (10 µl) and PI (10 µl) were added and cells were
incubated for 30 min at 4°C in the dark. Finally, 300 µl binding
buffer was added and the cells were analyzed using flow cytometry
(Cytomics FC 500) within 1 h and the results analyzed using CXP 2.1
Software (both from Beckman Coulter, Brea, CA, USA).
Cell migration assay
Cell migration and invasion were assessed by
Transwell assays. GH3 and GH4C1 cells from different groups were
starved with serum-free DMEM medium for 24 h. Cells were diluted to
1×105 cells/ml and 200 µl cell suspension was added in
the upper Transwell chambers (Corning Inc., Corning, NY, USA), and
500 µl DMEM medium supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) in the lower chambers, this was repeated three
times for every group. Following incubation for 48 h in 24-well
plates at 37°C, cells were fixed for 10 min using 1 ml/well 4%
paraformaldehyde (Bioswamp; Wuhan Beinglay Biological Technology
Co.), stained by 0.5% crystal violet (Bioswamp; Wuhan Beinglay
Biological Technology Co.) for 30 min at room temperature and
washed 3 times using 1X PBS. Finally, the Transwell plates were
wiped carefully with a cotton swab to remove non-migrated cells,
and viewed under an optical microscope (Nikon Corporation, Tokyo,
Japan) at magnification of ×200 to count the number of cells.
Western blot analysis
Antibodies against Gal-3 (cat. no. ab2785; dilution
1:1,000), BCL2 associated X apoptosis regulator (Bax; cat. no.
ab32503; dilution 1:2,000), caspase-3 (cat. no. ab90437; dilution
1:1,000), matrix metalloproteinase 7 (MMP7; cat. no. ab5706;
dilution 1:1,000) and β-actin (cat. no. ab8227; dilution 1:2,000)
were all purchased from Abcam (Cambridge, UK). Western blot
analysis was performed as previously described (10). Briefly, cells were washed with PBS and
lysed in lysis buffer (20 mmol/l Tris-HCL pH 7.4, 150 mmol/l NaCl,
0.5% Nonidet P-40, 1 mmol/l EDTA, 50 µg/ml leupeptin, 30 µg/ml
aprotinin, and 1 mmol/l PMSF). The protein concentration was
determined by BCA kit (Bioswamp; Wuhan Beinglay Biological
Technology Co.) Equal amounts of protein (30 µg) were separated by
10% SDS-PAGE and then transferred onto PVDF membrane (EMD
Millipore, Billerica, MA, USA). The membranes were blocked for 2 h
at room temperature with 5% skim milk in TBS (20 mmol/l Tris, 500
mmol/l NaCl, and 0.05% Tween-20). Subsequently, the membrane was
incubated with primary antibodies overnight at 4°C. Then, the
membranes were washed twice with TBS and incubated in biotinylated
goat anti-rabbit IgG secondary antibody (cat. no. W4011; 1:3,000
dilution; Promega Corporation, Madison, WI, USA) for 2 h at room
temperature. Immunoreactivity was visualized by colorimetric
reaction using ECL substrate buffer (EMD Millipore). Membranes were
scanned with Gel Doz EZ imager (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Statistical analysis
Data were expressed as the mean ± standard
deviation. Data were analyzed by one-way analysis of variance using
SPSS 19.0 software (IBM Corp, Armonk, NY, USA), followed by
Dunnett's or Duncan's tests for multiple comparisons. P<0.05,
P<0.01 and P<0.001 were considered to indicate a
statistically significant difference.
Results
Expression of Gal-3 in rat pituitary
tumor cells
To explore the expression levels of Gal-3 in rat
pituitary tumor cells, three cell lines were selected to undergo
RT-qPCR, and the results revealed that Gal-3 expression levels were
increased in GH3 and GH4C1 cells compared with RC-4B/C cells
(Fig. 1A). As presented in Fig. 1B and C, the interference vector
(shGal-3) and expression vector (pGal-3) were successfully
expressed in GH3 and GH4C1 cells. Then, the expression efficiency
of the vectors was assessed with RT-qPCR, which revealed that
shGal-3 significantly reduced the expression of Gal-3 and pGal-3
markedly increased the expression level of Gal-3 compared with
their respective controls (Fig. 1D and
E).
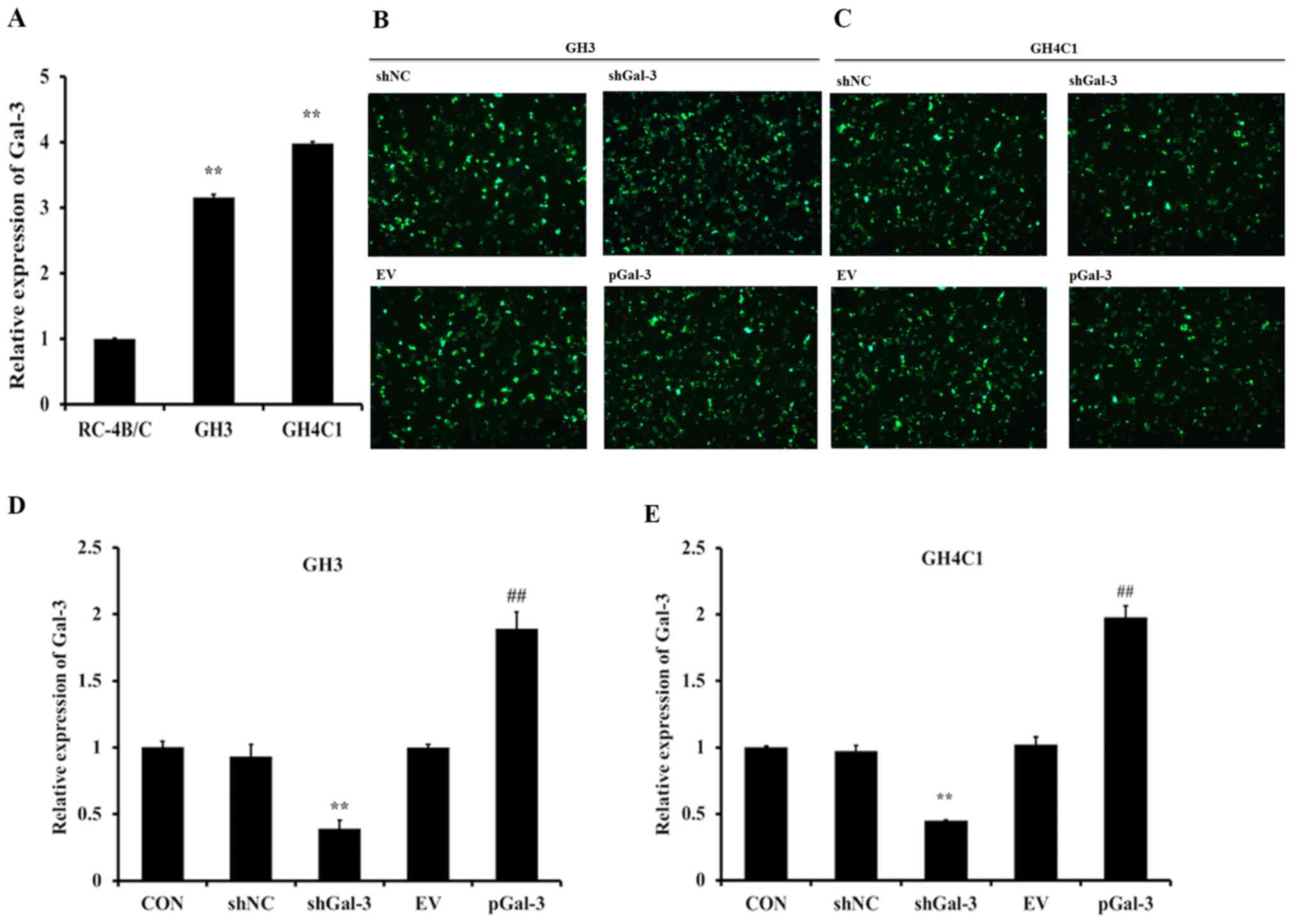 | Figure 1.Expression of Gal-3 in rat pituitary
tumor cell lines. (A) The mRNA levels of Gal-3 in three different
cell lines (GH3, GH4C1 and RC-4B/C) were detected by RT-qPCR. (B)
GH3 cells and (C) GH4C1 cells transfected with shNC, shGal-3, EV
and pGal-3 and detected by fluorescence microscopy (magnification,
×200). The mRNA expression levels of Gal-3 in (D) GH3 cells and (E)
GH4C1 cells transfected with shNC, shGal-3, EV and pGal-3
**P<0.01 vs. RC-4B/C or shNC; ##P<0.01 vs.
shGal-3. Gal-3, galectin-3; shNC, negative control interference
vector; shGal-3, galectin-3 interference vector; EV, empty vector
control; pGal-3, galectin-3 overexpression vector; CON,
control. |
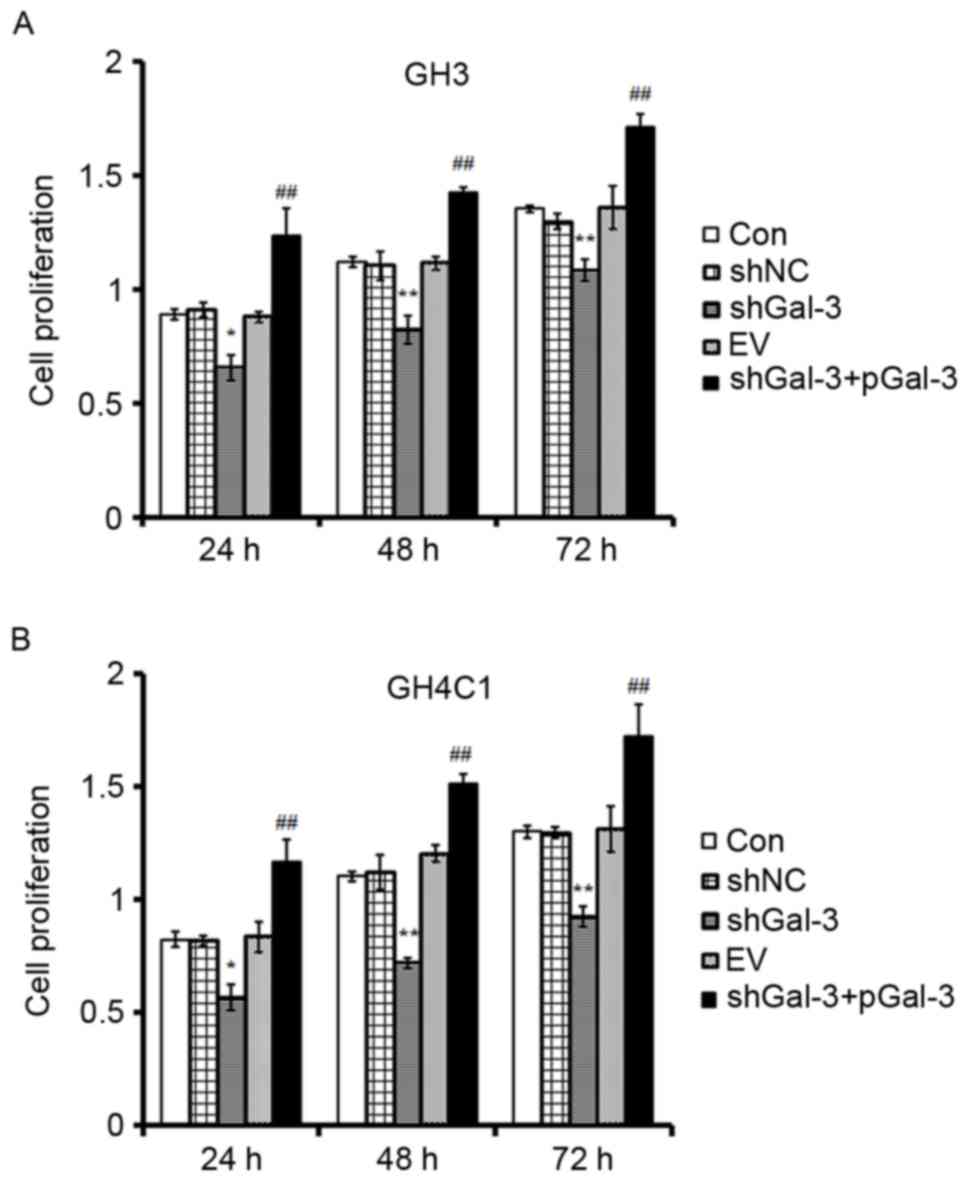
The effect of Gal-3 on rat pituitary
tumor cell proliferation
Cell proliferation was measured by MTT. There were
five groups: CON (control), shNC (cells transfected with the
negative control interference vector), shGal-3 (cells transfected
with shGal-3), EV (cells transfected with an empty vector) and
shGal-3+pGal-3 (cells transfected with shGal-3 initially, and then
the Gal-3 expression vector). As presented in Fig. 2, cell proliferation in the shGal-3
group was decreased compared with the shNC group at 24, 48 and 72
h. Cell proliferation in the shGal-3+pGal-3 group was also
significantly increased in a time-dependent manner compared with
the shGal-3 group (Fig. 2). The
results indicated that Gal-3 promoted the proliferation of GH3 and
GH4C1 cells.
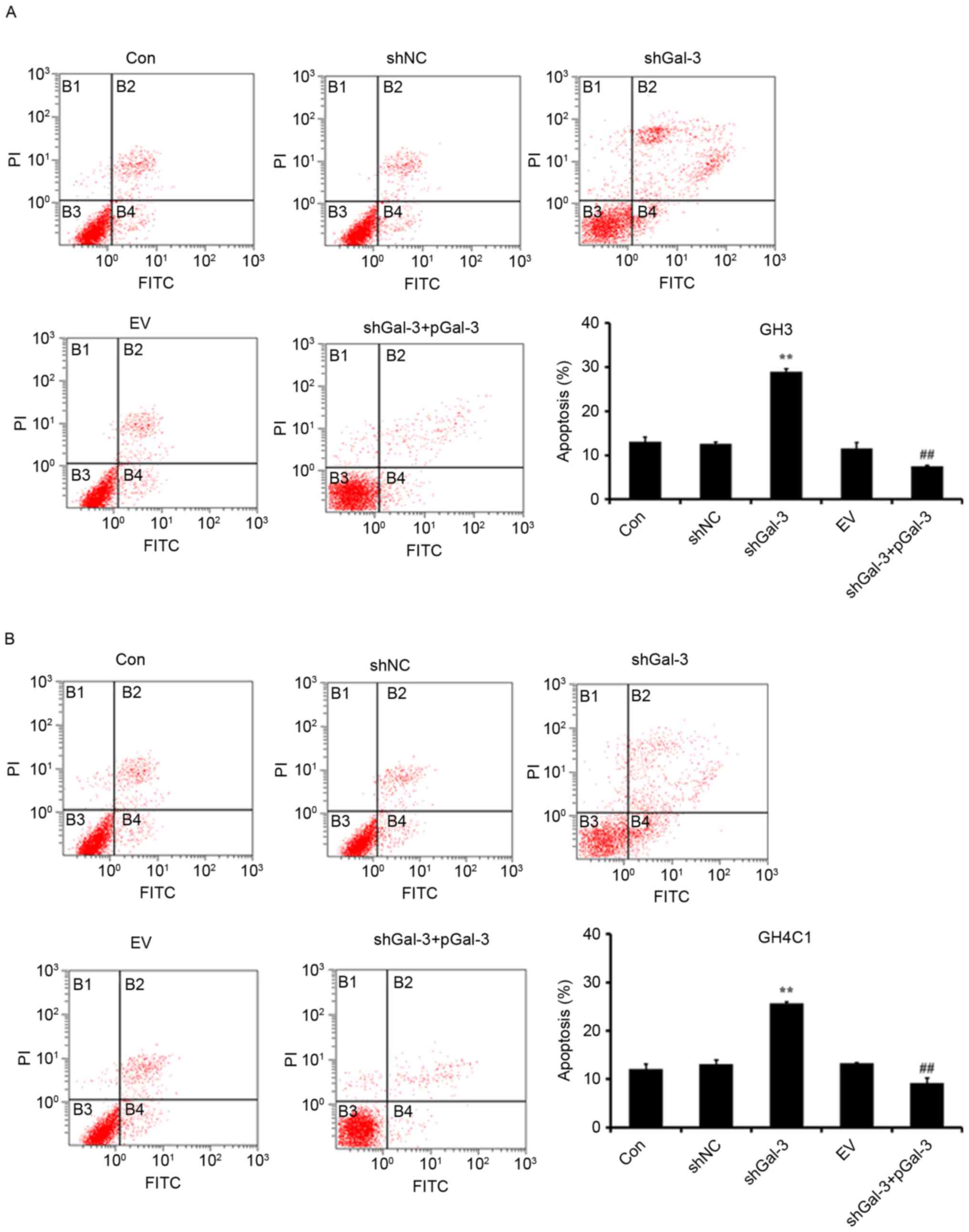
Effect of Gal-3 on rat pituitary tumor
cell apoptosis
Cell apoptosis was measured by Annexin V-FITC/PI
flow cytometry. As presented in Fig.
3, transfection with shGal-3 significantly increased the
percentage of apoptotic cells compared with the shNC group, while
the shGal-3+pGal-3 group had decreased apoptosis compared with the
shGal-3 group. These results indicated that Gal-3 inhibited the
apoptosis of GH3 and GH4C1 cells.
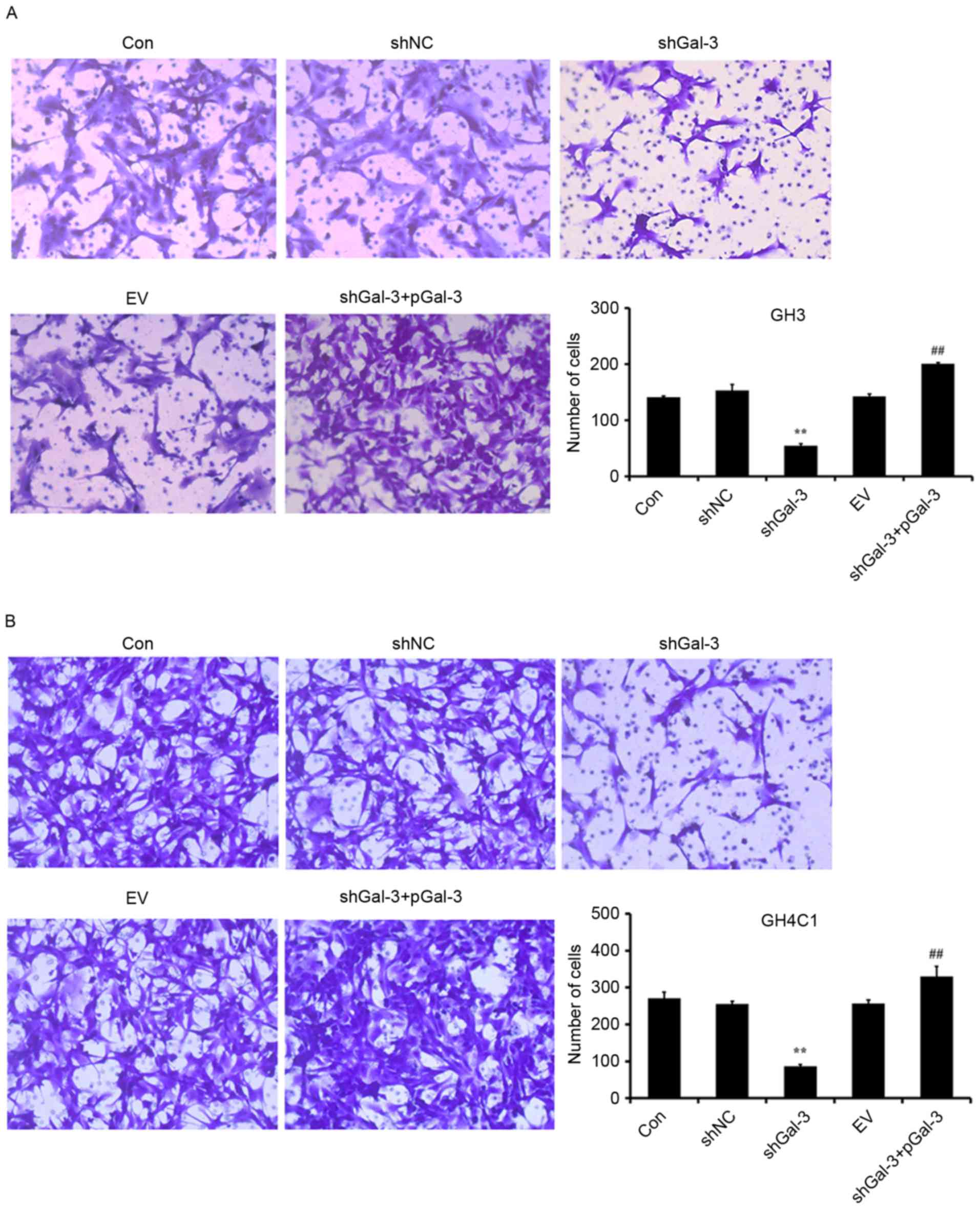
Effect of Gal-3 on rat pituitary tumor
cell migration
Cell migration was assessed using a Transwell test.
As presented in Fig. 4, transfection
with shGal-3 significantly decreased the number of migrating cells
compared with the shNC group, while shGal-3+pGal-3 groups
significantly increased the migration of cells compared with
shGal-3. These results indicated that Gal-3 increased the migration
of GH3 and GH4C1 cells.
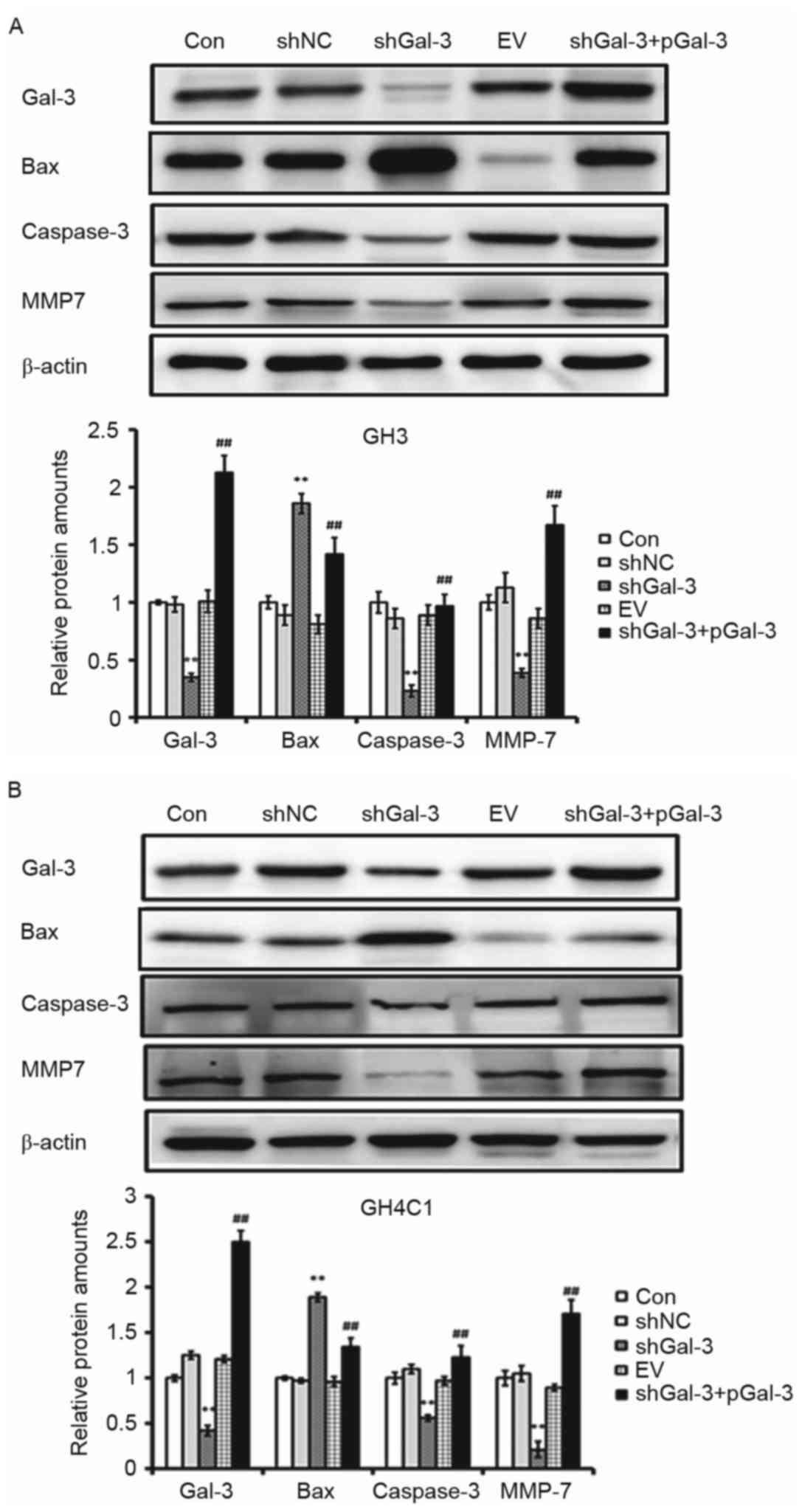
Effect of Gal-3 on
apoptosis-associated genes and oncogenes
In order to further understand the effect of Gal-3
on apoptosis-associated genes and oncogenes, the expression of Bax,
caspase-3 and MMP7 were detected by Western blot. As presented in
Fig. 5, in GH3 and GH4C1 cells, the
protein expression levels of caspase-3 and MMP7 were decreased
while Bax expression was increased in the shGal-3 group compared
with the controls. In addition, caspase-3 and MMP7 protein
expression were increased following overexpression of Gal-3, while
Bax expression was decreased (Fig.
5).
Discussion
Galectins are a family of animal lectins with
diverse biological functions. They are involved in extracellular
and intracellular processes: Extracellularly, by interacting with
cell-surface and extracellular matrix glycoproteins and
glycolipids, and intracellularly, by interacting with cytoplasmic
and nuclear proteins to regulate signaling pathways (11). Previous research has demonstrated that
galectins serve vital functions in cancer and that they contribute
to neoplastic transformation, the maintenance of transformed
phenotypes, tumor cell adhesion and survival, angiogenesis and
tumor metastasis (12). Among the
different members of the galectin family, Gal-3 has been reported
to be involved in various biological processes, including cell
proliferation and differentiation, tumor cell adhesion, apoptosis,
tumor progression and metastasis.
In the present study, Gal-3 expression levels in GH3
and GH4C1 cells were increased compared with RC-4B/C cells. Then,
two pituitary tumor cell lines were selected for further study
concerning the role of Gal-3 on cell proliferation, cell apoptosis,
cell migration and the expression of apoptosis-associated genes and
oncogenes. In a previous study, inhibition of Gal-3 gene expression
by RNA interference decreased HP75 cell proliferation and increased
apoptosis (13). The present study
demonstrated that inhibition of Gal-3 expression following
transfection with RNA interference vectors decreased cell
proliferation, migration and the expression of caspase-3 and MMP7
in GH3 and GH4C1 cells, but increased cell apoptosis and expression
of the apoptosis-associated gene Bax. Furthermore, following
interference with Gal-3 expression, overexpression of Gal-3
repaired the reduction of cell proliferation, migration and
expression of caspase-3 and MMP7, and inhibited cell apoptosis and
Bax protein expression. These results indicated that Gal-3
contributes to the tumorigenesis and progression of pituitary
tumors.
Gal-3 is expressed in a number of types of tumor,
and the intensity of its expression and localization depend on
tumor progression, invasiveness and metastatic potential (14). Previous evidence suggests that Gal-3
may be a useful biomarker for differential diagnosis of brain
tumors, including various glioneuronal tumors, pituitary adenomas,
meningiomas and schwannomas (15).
The present study is focused on the involvement of Gal-3 on
pituitary adenomas. Given the function of Gal-3 in pituitary tumor
cell proliferation and apoptosis, Gal-3 may be a potential target
for the treatment of aggressive pituitary tumors. However, the
precise mechanisms by which Gal-3 promotes the tumorigenesis and
progression of pituitary tumors require further study.
Acknowledgements
The present study was supported by grants from
Project of Health and Family Planning Commission of Hubei Province
(grant nos. WJ2015MA012 and WJ2015MB118).
References
|
1
|
Jiang X and Zhang X: The molecular
pathogenesis of pituitary adenomas: An update. Endocrinol Metab
(Seoul). 28:245–254. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chatzellis E, Alexandraki KI, Androulakis
II and Kaltsas G: Aggressive pituitary tumors. Neuroendocrinology.
101:87–104. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gong J, Diao BO, Yao GJ, Liu Y and Xu GZ:
Analysis of regulatory networks constructed based on gene
coexpression in pituitary adenoma. J Genet. 92:489–497. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Huang CX, Hou YH and Liu YS: Expression of
galectin-3 correlates with apoptosis in pituitary adenoma cells.
Neurosci Bull. 24:34–38. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Arfaoui-Toumi A, Kria-Ben Mahmoud L, Ben
Hmida M, Khalfallah MT, Regaya-Mzabi S and Bouraoui S: Implication
of the galectin-3 in colorectal cancer development (about 325
tunisian patients). Bull Cancer. 97:E1–E8. 2010.PubMed/NCBI
|
|
6
|
Manivannan P, Siddaraju N, Jatiya L and
Verma SK: Role of pro-angiogenic marker galectin-3 in follicular
neoplasms of thyroid. Indian J Biochem Biophys. 49:392–394.
2012.PubMed/NCBI
|
|
7
|
Yamaki S, Fujii T, Yajima R, Hirakata T,
Yamaguchi S, Fujisawa T, Tsutsumi S, Asao T, Yanagita Y, Iijima M
and Kuwano H: Clinicopathological significance of decreased
galectin-3 expression and the long-term prognosis in patients with
breast cancer. Surg Today. 43:901–905. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang CX, Zhao JN, Zou WH, Li JJ, Wang PC,
Liu CH and Wang YB: Reduction of galectin-3 expression reduces
pituitary tumor cell progression. Genet Mol Res. 13:6892–6898.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Na YJ, Jeon YJ, Suh JH, Kang JS, Yang KH
and Kim HM: Suppression of IL-8 gene expression by radicicol is
mediated through the inhibition of ERK1/2 and p38 signaling and
negative regulation of NF-kappaB and AP-1. Int Immunopharmacol.
1:1877–1887. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu FT and Rabinovich GA: Galectins as
modulators of tumour progression. Nat Rev Cancer. 5:29–41. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Newlaczyl AU and Yu LG: Galectin-3-a
jack-of-all-trades in cancer. Cancer Lett. 313:123–128. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Riss D, Jin L, Qian X, Bayliss J,
Scheithauer BW, Young WF Jr, Vidal S, Kovacs K, Raz A and Lloyd RV:
Differential expression of galectin-3 in pituitary tumors. Cancer
Res. 63:2251–2255. 2003.PubMed/NCBI
|
|
14
|
Righi A, Jin L, Zhang S, Stilling G,
Scheithauer BW, Kovacs K and Lloyd RV: Identification and
consequences of galectin-3 expression in pituitary tumors. Mol Cell
Endocrinol. 326:8–14. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Park SH, Min HS, Kim B, Myung J and Paek
SH: Galectin-3: A useful biomarker for differential diagnosis of
brain tumors. Neuropathology. 28:497–506. 2008. View Article : Google Scholar : PubMed/NCBI
|