Introduction
Hepatocellular carcinoma (HCC) is a major malignancy
worldwide and exhibits increased incidence of tumor recurrence and
metastasis (1,2). Hepatitis B virus (HBV)-encoded X antigen
(HBx) acts as a multifunctional regulator in HBV-associated HCC
development (3). HBx-transgenic mice
exhibited increased expression of HBx, which may lead to HCC
(4,5).
However, the mechanism of the direct function of long-term
expression of HBx in the development of HCC remains obscure.
3-Phosphoinositide-dependent protein kinase-1 (PDK1)
is a crucial kinase that functions as the downstream effector
molecular regulator of phosphoinositide 3-kinase (PI3K) and
activates members of the activated protein kinase-dependent, cyclic
guanosine monophosphate-dependent and protein kinase C family of
protein kinases, including protein kinase B (Akt), protein kinase
C, p70 ribosomal protein S6 kinases and serum- and
glucocorticoid-dependent kinase (6–9).
Hyperactivation of PDK1 was identified in several types of human
cancer (10–12). Downregulation of PDK1 inhibits
metastasis of human breast cancer cells (12). Clinical data suggested that PDK1
exhibit increased expression in liver cancer and was associated
with a significantly decreased postoperative overall survival rate
and increased recurrence rates in patients with HCC following
curative resection (13).
Nevertheless, the molecular mechanism by which PDK1 contributes to
carcinogenesis of liver cancer (particularly of HBV-associated
liver cancer) remains unclear.
With-no-lysine (K) kinases (WNKs) are a group of
serine-threonine protein kinases with an atypical placement of the
catalytic lysine residue compared with other protein kinases
(14,15). WNK1 is the main downstream effector of
the PDK1-associated pathway (16),
which serves an important function in cell proliferation and
migration (17). Previous studies
demonstrated that WNK1 may also be an important kinase involved in
various types of cancer (18–21).
The results of the present study indicated the
increased expression of phospho (p)-PDK1 in HBV-associated HCC.
Additionally, the activation of PDK1/WNK1 signaling was
investigated in stable hepatic cell lines expressing HBx. An
inhibitor of PDK1 significantly suppressed the viability and
metastasis of hepatic cells.
Materials and methods
Cell culture
Human hepatoma cells SK-Hep1 and hepatic cells LO2
(Cell Resource Center, Shanghai Institute of Life Sciences, Chinese
Academy of Sciences, Shanghai, China) were maintained in Dulbecco's
modified Eagle's medium (DMEM) supplemented with 10% fetal bovine
serum, 100 U/ml penicillin and 100 µg/ml streptomycin at 37°C in a
humidified atmosphere containing 5% CO2. All reagents
were obtained from Corning Incorporated (Corning, NY, USA).
Transient transfection and generation
of stable cell lines
pHAGE-HBx vector expressing full-length HBx was
constructed. The synthesized full-length HBx gene was inserted into
the pHAGE vector to generate pHAGE-HBx plasmid. The construct was
confirmed by DNA sequencing. LO2 and SK-Hep1 cells were transfected
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) using 2.5 µg plasmids in a
6-well plate, according to the manufacturer's protocol. For stable
transfection, LO2 and SK-Hep1 cells were transfected using
pHAGE-HBx (or pHAGE-vector) for 48 h and maintained in DMEM
containing 2 µg/ml puromycin. After 3–4 weeks of selection,
individual colonies were isolated and expanded. The increased
expression of target genes in these colonies was confirmed using
the reverse transcription-quantitative polymerase chain reaction
(RT-qPCR).
RT-qPCR
Total RNA was extracted from HBx transfected or the
control cells using an Ultrapure RNA kit (CW0586S; CW Biotech,
Beijing, China). A total of 400 ng RNA was transcribed into cDNA
using the PrimeScript RT Mastermix (cat.no. RR036Q; Takara
Biotechnology Co., Ltd., Dalian, China), according to the
manufacturer's protocol. The cDNA was amplified using the
SYBR-Green PCR Master Mix (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. The thermocycling
conditions were as follows: Initial denaturation of 95°C for 5 min
followed by 40 cycles of 94°C for 15 sec and final extension of
55°C for 1 min. The following primers were used: HBx forward,
5′-actctcagcaatgtcaacg-3′ and reverse, 5′-atttatgcctacagcctcc-3′;
GAPDH forward, 5′-tgaaggtcggagtcaacgga-3′ and reverse
5′-cctggaagatggtgatgggat-3′. Reactions were performed in
triplicate. Relative expression of HBx was determined using the
2−ΔΔCq method (22).
Western blot analysis
Protein lysates from tissues were prepared using a
1% protease and phosphatase inhibitor cocktail [Halt™ Protease and
Phosphatase Inhibitor Cocktail, EDTA-Free (100X); cat. no. 78441;
Thermo Fisher Scientific, Inc.] in a lysis buffer (4% SDS, 0.1 M
dithiothreitol and 0.1 M Tris/HCl, pH 7.6). Cells were lyzed in
radioimmunoprecipitation assay buffer (50 mM Tris/HCl, 1% Triton
X-100, 0.5% sodium deoxycholate, 150 mM NaCl, 1 mM EDTA and 0.1%
SDS) containing a protease and phosphatase inhibitor cocktail (as
above). The total protein concentration was measured by BCA assay
and samples containing equal amounts of total protein (30 µg) were
separated by 10% SDS-PAGE and transferred onto nitrocellulose
membranes. The membranes were then blocked with 5% non-fat milk in
Tris-buffered saline at room temperature for 1 h. Following
blocking, membranes were incubated overnight at 4°C with the
following primary antibodies: Rabbit anti-WNK1 antibody (1:500;
cat. no. 4979S; Cell Signaling Technology, Inc., Danvers, MA, USA),
anti-p-PDK1 (Ser241) antibody (1:500; cat. no. 3061S;
Cell Signaling Technology, Inc.), rabbit anti-PDK1 antibody (1:500;
cat. no. 3062S; Cell Signaling Technology, Inc.), anti-p-WNK1
(Thr60) antibody (1:200; cat. no. AF4720; R&D
Systems, Inc., Minneapolis, MN, USA) and anti-GAPDH antibody
(1:2,000; cat. no. TA08; OriGene Technologies, Inc., Beijing,
China). Membranes were then washed and incubated with secondary
antibodies Goat Anti-Mouse IgG, HRP Conjugated (1:5,000; cat. no.
CW0102; CW Biotech, Beijing, China) or Goat Anti-Rabbit IgG, HRP
Conjugated (1:5,000; cat. no. CW0103; CW Biotech) at room
temperature for 1 h. The protein bands were visualized using
enhanced chemiluminescence Western Blotting Substrate (cat. no.
32106; Pierce; Thermo Fisher Scientific, Inc.)
Cell viability assay
Hepatic cell viability was detected using a Cell
Counting kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). Cells (5×103 cells/well in 100 µl
medium) were seeded in a 96-well plate. Cells were treated with 0,
5 or 20 µM PDK1 inhibitor II (cat. no. 521276; Merck KGaA,
Darmstadt, Germany) in a humidified incubator (37°C, 5%
CO2). Following incubation for the indicated durations,
a 1/10 volume of the CCK-8 solution was added to each well and
cells were incubated for a further 1 h. Cell viability was
determined by measuring the absorbance at 450 nm using a plate
reader (Spectramax Paradigm, Molecular Devices, LLC, Sunnyvale, CA,
USA).
Transwell migration assay
For the Transwell migration assay, 2×104
cells [suspended in DMEM containing 0.5% fetal bovine serum (FBS)]
were trypsinized and plated in the upper chamber of
fibronectin-coated plates (8 µm pore filter, Corning Incorporated).
The cells were then treated with 0, 5 or 20 µM PDK1 inhibitor II.
The lower chamber of the Transwell contained DMEM supplemented with
2.5% FBS. At 20 h, the filters were removed and the cells located
on the membrane were fixed with methanol for 2 h. The cells that
had migrated to the underside of the membrane were stained using
0.5% crystal violet at room temperature for 1 h and examined under
a light microscope (Nikon Eclipse 80i; Nikon Corporation, Tokyo,
Japan).
Clinical HCC samples
Primary HCC tissue and the paired normal liver
tissue specimens were obtained from 21 patients with primary liver
cancer who underwent surgical resection at China-Japan Union
Hospital of Jilin University (Jilin, China). The samples were
collected between March 2012 and February 2015. Of the total 21
patients, 17 (81%) were male and 4 (19%) were female. The median
age was 57.8 years (range, 42–73 years). All participants provided
written informed consent. Ethical approval was obtained from the
research ethics committee of China-Japan Union Hospital of Jilin
University.
Statistical analysis
Data were analyzed using GraphPad Prism (version
6.0) software (GraphPad Software, Inc., La Jolla, CA, USA). The
relevant data are expressed as the mean ± standard deviation.
Statistical significance between two groups was determined using
Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
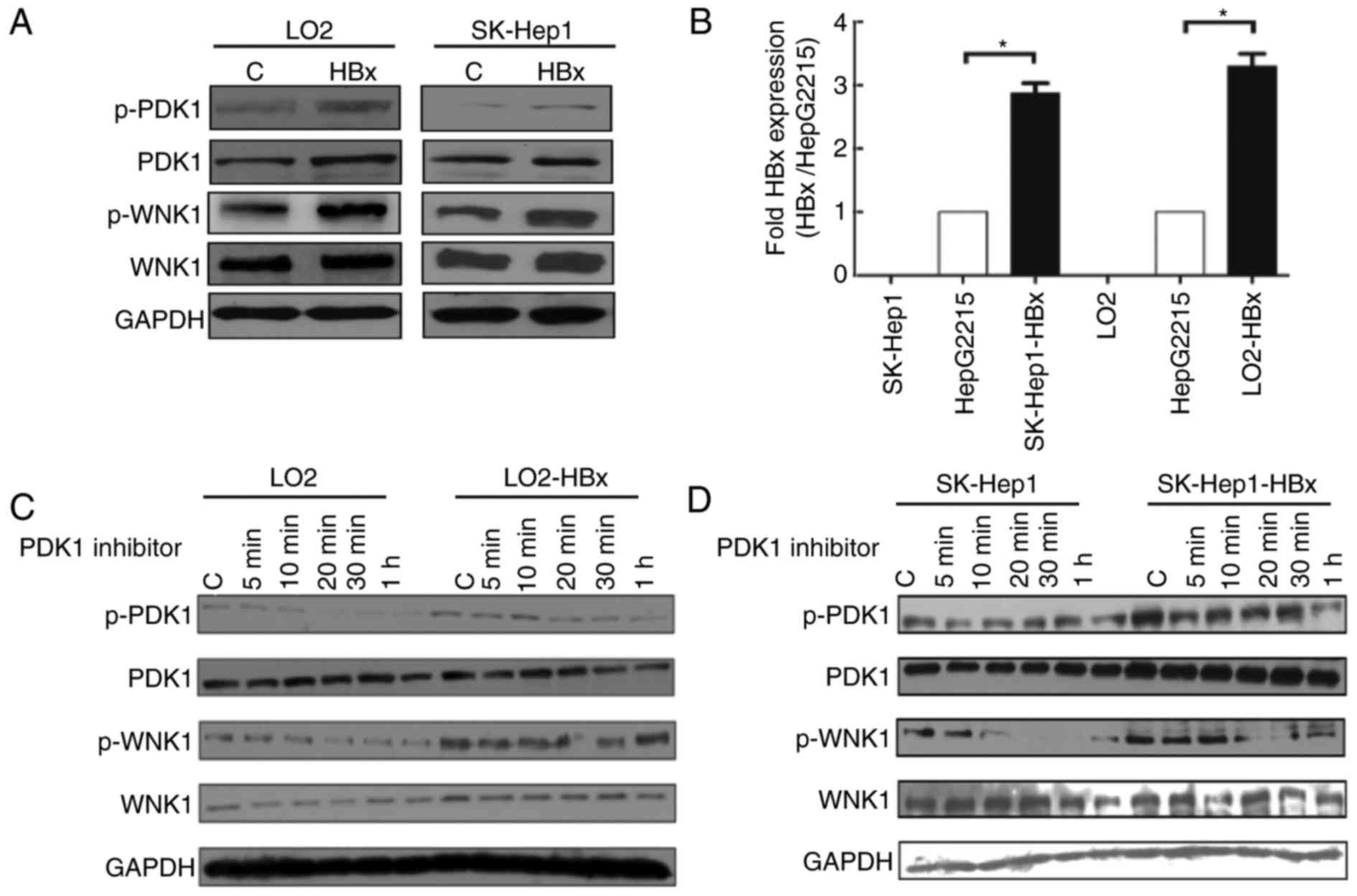
HBx affects the phosphorylation of
PDK1/WNK1 in hepatic cells
To evaluate the association between HBx and PDK1,
the expression of PDK1 and WNK1 was examined in HBx-overexpressing
hepatic cell lines. Hepatic cell lines SK-Hep1 and LO2 were
transfected with pHAGE-HBx or pHAGE-vector constructs and the
overexpression of HBx in the pHAGE-HBx construct was confirmed
using RT-qPCR (data not shown). Expression levels of p-PDK1
(Ser241) and p-WNK1 (Thr60) were markedly
upregulated in HBx-transfected hepatic cells compared with in the
control group (Fig. 1A). These
results indicate that HBx may affect the phosphorylation of PDK1
and WNK1 in hepatic cells.
PDK1/WNK1 pathway is suppressed in
response to the PDK1 inhibitor in vitro
SK-Hep1 and LO2 cell lines were employed to
establish HBx-stable expression cell lines in order to study the
mechanism underlying HBx-mediated regulation of PDK1. Stable
expression of HBx by gene transfection was confirmed using RT-qPCR.
HepG2.2.15 was used as an HBx-positive control cell line (Fig. 1B). Next, the expression of PDK1 and
WNK1 was evaluated in transfected or non-transfected SK-Hep1 and
LO2 cell lines treated with 20 µM PDK1 inhibitor for 0, 5, 10, 20,
30 and 60 min (Fig. 1C and D). The
phosphorylation of PDK1 and WNK1 were inhibited when SK-Hep1 and
LO2 cells were treated with a PDK1 inhibitor (Fig. 1C and D). The effects of the inhibitor
were decreased in stable cell lines expressing HBx compared with
the control (Fig. 1C and D). These
results demonstrate that WNK1 is one of the downstream effectors of
PDK1. Additionally, HBx may attenuate the effect of the PDK1
inhibitor on hepatic cells and thus regulate the PDK1/WNK1
signaling pathway.
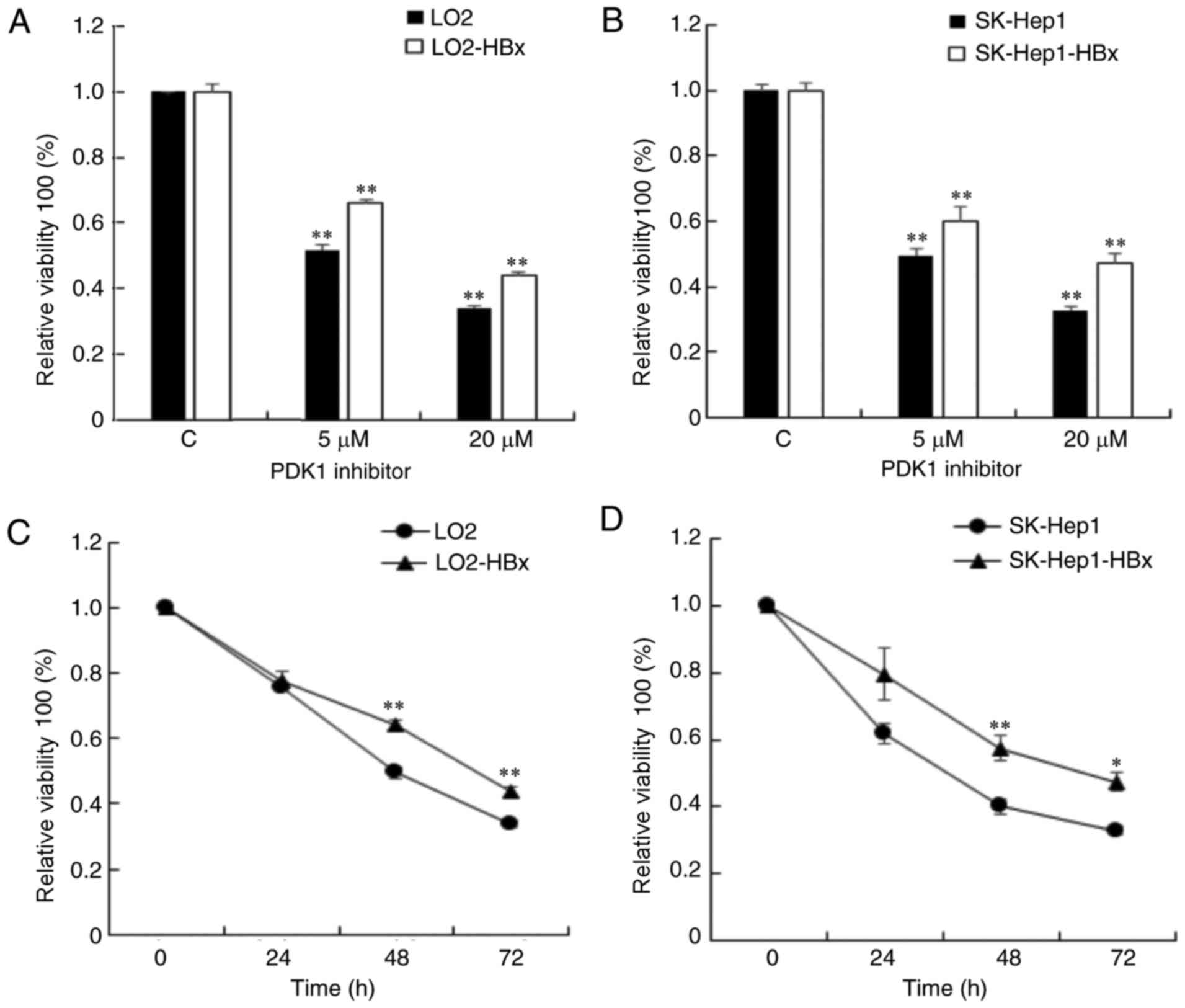
HBx inhibits the viability of hepatic
cells in response to the PDK1 inhibitor
The LO2-HBx cell line with stable expression of HBx
exhibited increased viability compared with the control (data not
shown). To investigate how HBx affects hepatic cell viability, a
CCK-8 assay was performed on LO2/LO2-HBx and SK-Hep1/SK-Hep1-HBx
cells. Hepatic cells were treated with 0, 5, 20 µM PDK1 inhibitor
for 72 h (Fig. 2A and B) and with 20
µM PDK1 inhibitor for 0, 24, 48 and 72 h (Fig. 2C and D). The viability of all cells
was significantly suppressed at 48 and 72 h, whereas the
suppression in the HBx stable-expression cells was weaker compared
with the control cells. Thus, the results of the present study
suggest that HBx may attenuate the effect of PDK1 inhibitor on the
viability of hepatic cells.
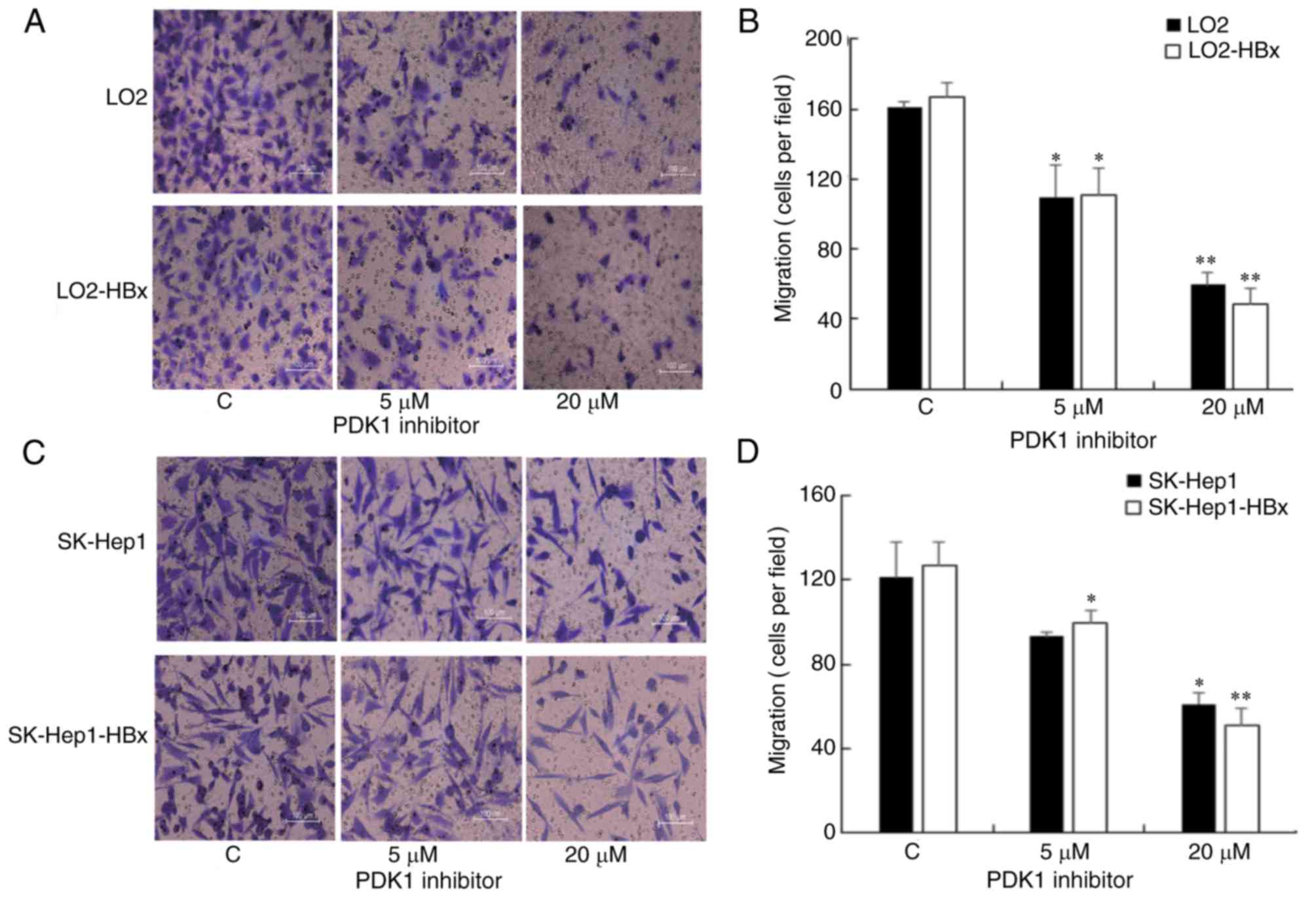
PDK1 inhibitor suppresses the
migration of hepatic cells
To investigate the function of the PDK1/WNK1
signaling pathway in the metastasis of hepatic cells, a Transwell
assay was employed to examine the migratory ability of LO2/LO2-HBx
(Fig. 3A and B) and
SK-Hep1/SK-Hep1-HBx cells (Fig. 3C and
D). The results indicated that the migration of cells was
gradually decreased in response to increasing doses of the PDK1
inhibitor in LO2/LO2-HBx (Fig. 3B)
and SK-Hep1/SK-Hep1-HBx cells (Fig.
3D). No significant differences in the migratory ability were
identified between the control and corresponding stable
HBx-expressing cells.
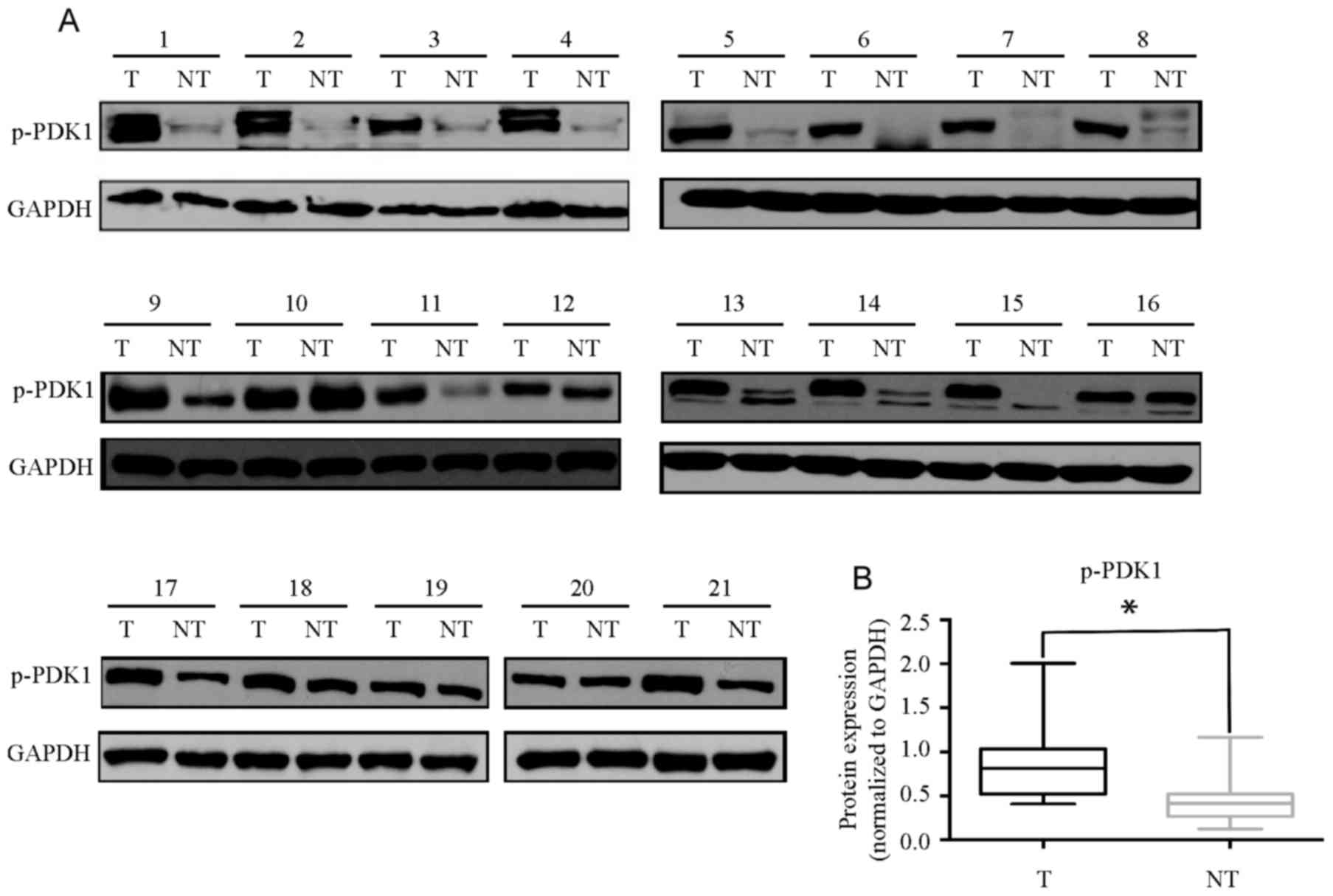
p-PDK1 is overexpressed in human HCC
tissues
Since activation of PDK1 is associated with
overexpression of HBx, p-PDK1 expression was evaluated in primary
HCC and paired normal liver tissue specimens (Fig. 4A). Increased expression of p-PDK1 was
identified in 71% (15/21) of the HCC samples compared with in the
corresponding non-tumor livers (Fig.
4B). These results indicate that p-PDK1 may be associated with
the development of HBV-associated HCC.
Discussion
Chronic infection in HBV is a major cause of liver
cancer, accounting for 55% of cases worldwide (23). The molecular mechanism underlying
HBV-induced HCC remains unclear. However, evidence suggests that
HBx protein serves a pivotal function in the development of HCC
(24). HBx may regulate several
signaling pathways. For example, HBx together with Akt may be
important for cell proliferation and tumorigenic transformation of
cells (25). Previous studies have
demonstrated that HBx activated Wnt/β-catenin signaling and induced
the Janus kinase/signal transducer and activator of transcription
signaling pathway (26–29). The molecular mechanisms involved in
the development of HBV-associated HCC are complex. Although a
number of molecules and signaling pathways have been identified,
further research is required.
The PI3K signaling pathway is one of the most
important pathways in cell proliferation and may be a suitable
target for cancer therapeutic intervention (30–32).
Previous studies demonstrated that downregulation of PDK1 may
suppress the progression of breast cancer (33,34). PDK1
is highly expressed in liver cancer and is associated with
prognosis (13). PDK1 may also be a
suitable target for HBV-induced cancer.
In the present study, the function of PDK1/WNK1
signaling in HBx-overexpressing hepatic cell lines was explored.
The results suggested an overexpression of p-PDK1 in the majority
of HCC tissues. Additionally, overexpression of p-PDK1 was
significantly associated with the expression of HBx which is an
important HBV-encoded protein involved in the genesis and
development of HCC (3). The results
of the present study demonstrated that upregulation of p-PDK1
occurs in HBV-associated HCC. In the present study, the expression
of HBx in human HCC tissues was not examined since all patients
were HBV-positive (indicating the expression of HBx). Nevertheless,
it has been demonstrated previously that the protein expression of
HBx is not always detectable in HBV-associated HCC (35), thus further investigation is
required.
The molecular mechanism underlying the upregulation
of p-PDK1 in HCC remains unclear. However, several studies
identified that PDK1 may be a potential therapeutic target to
improve melanoma and angiosarcoma treatment (10,11,36). The
results of the present study demonstrated that the inhibition of
the kinase activity of PDK1 may decrease the viability and
metastasis of hepatic cells. Additionally, no differences were
observed in the migration of control and stable HBx-expressing
cells, thus suggesting that the regulation of the PDK1/WNK1
signaling pathway by HBx may not be pivotal for liver cancer
metastasis. Evidence suggests that a PDK1 inhibitor may regulate
metastasis mainly through additional PDK1-mediated signaling
pathways (12).
The results of the present study demonstrated that
the expression of p-PDK1 and p-WNK1 was upregulated in
HBx-overexpressing HCC cells, which is in accordance with the
phosphoproteomic data of HCC samples (data not shown). Evidence
suggests that the PDK1/WNK1 signaling pathway is associated with
HBx-associated HCC. In the present study, it has been demonstrated
that the expression of p-PDK1 and p-WNK1 were upregulated in stable
HBx-expressing hepatic cells. Additionally, the upregulation of
p-PDK1 and p-WNK1 was inhibited in response to a PDK1 inhibitor in
LO2-HBx and SK-hep1-HBx cell lines in a dose- and time-dependent
manner. Stable expression of HBx reversed the effects of PDK1
inhibitor on cell viability compared with the control.
In conclusion, p-PDK1 is upregulated in human HCC
tissues and in stable HBx-expressing hepatic cell lines. The
overexpression of p-PDK1 serves an important function in
HBx-associated cell viability. PDK1 may be a potential target for
combating HBV-induced liver cancer.
Acknowledgements
The present study was supported by the 973 Program
of Chinese State Key Projects for Basic Research (grant nos.
2014CBA02001 and 2013CB910502), National Key Research and
Development Project (grant no. 2016YFC0902400), National Natural
Science Foundation of China (grant nos. 81123001, 81570526 and
81472209), Innovation Project (grant no. 16CXZ027), the 863 Program
of Chinese State High-Tech Program (grant nos. 2012AA020204 and
2014AA020906), the Program of International S&T Cooperation
(grant nos. 2014DFB30020 and 2014DFB30010), Natural Science
Foundation of Beijing (grant no. 7152036), Open Project Program of
the State Key Laboratory of Proteomics (Academy of Military Medical
Sciences; grant no. SKLP-O201509) and the China Postdoctoral
Science Foundation (grant no. 2015M582851).
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tang ZY: Hepatocellular carcinoma-cause,
treatment and metastasis. World J Gastroenterol. 7:445–454. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang XD, Wang Y and Ye LH: Hepatitis B
virus X protein accelerates the development of hepatoma. Cancer
Biol Med. 11:182–190. 2014.PubMed/NCBI
|
|
4
|
Wang Y, Cui F, Lv Y, Li C, Xu X, Deng C,
Wang D, Sun Y, Hu G, Lang Z, et al: HBsAg and HBx knocked into the
p21 locus causes hepatocellular carcinoma in mice. Hepatology.
39:318–324. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Arzumanyan A, Reis HM and Feitelson MA:
Pathogenic mechanisms in HBV- and HCV-associated hepatocellular
carcinoma. Nat Rev Cancer. 13:123–135. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fyffe C and Falasca M:
3-Phosphoinositide-dependent protein kinase-1 as an emerging target
in the management of breast cancer. Cancer Manag Res. 5:271–280.
2013.PubMed/NCBI
|
|
7
|
Cho JY and Park J: Contribution of natural
inhibitors to the understanding of the PI3K/PDK1/PKB pathway in the
insulin-mediated intracellular signaling cascade. Int J Mol Sci.
9:2217–2230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mora A, Komander D, van Aalten DM and
Alessi DR: PDK1, the master regulator of AGC kinase signal
transduction. Semin Cell Dev Biol. 15:161–170. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Castel P, Ellis H, Bago R, Toska E, Razavi
P, Carmona FJ, Kannan S, Verma CS, Dickler M, Chandarlapaty S, et
al: PDK1-SGK1 signaling sustains AKT-independent mTORC1 activation
and confers resistance to PI3Kα inhibition. Cancer Cell.
30:229–242. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fujiwara S, Kawano Y, Yuki H, Okuno Y,
Nosaka K, Mitsuya H and Hata H: PDK1 inhibition is a novel
therapeutic target in multiple myeloma. Br J Cancer. 108:170–178.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wada M, Horinaka M, Yasuda S, Masuzawa M,
Sakai T and Katoh N: PDK1 is a potential therapeutic target against
angiosarcoma cells. J Dermatol Sci. 78:44–50. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Du J, Yang M, Chen S, Li D, Chang Z and
Dong Z: PDK1 promotes tumor growth and metastasis in a spontaneous
breast cancer model. Oncogene. 35:3314–3323. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang J, Liu F, Ao P, Li X, Zheng H, Wu D,
Zhang N, She J, Yuan J and Wu X: Correlation of PDK1 expression
with clinicopathologic features and prognosis of hepatocellular
carcinoma. Onco Targets Ther. 9:5597–5602. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Verissimo F and Jordan P: WNK kinases, a
novel protein kinase subfamily in multi-cellular organisms.
Oncogene. 20:5562–5569. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang CL, Cha SK, Wang HR, Xie J and Cobb
MH: WNKs: Protein kinases with a unique kinase domain. Exp Mol Med.
39:565–573. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Vitari AC, Deak M, Collins BJ, Morrice N,
Prescott AR, Phelan A, Humphreys S and Alessi DR: WNK1, the kinase
mutated in an inherited high-blood-pressure syndrome, is a novel
PKB (protein kinase B)/Akt substrate. Biochem J. 378:257–268. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun X, Gao L, Yu RK and Zeng G:
Down-regulation of WNK1 protein kinase in neural progenitor cells
suppresses cell proliferation and migration. J Neurochem.
99:1114–1121. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shyamasundar S, Lim JP and Bay BH: miR-93
inhibits the invasive potential of triple-negative breast cancer
cells in vitro via protein kinase WNK1. Int J Oncol. 49:2629–2636.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Costa V, Esposito R, Ziviello C, Sepe R,
Bim LV, Cacciola NA, Decaussin-Petrucci M, Pallante P, Fusco A and
Ciccodicola A: New somatic mutations and WNK1-B4GALNT3 gene fusion
in papillary thyroid carcinoma. Oncotarget. 6:11242–11251. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xie T, D'Ario G, Lamb JR, Martin E, Wang
K, Tejpar S, Delorenzi M, Bosman FT, Roth AD, Yan P, et al: A
comprehensive characterization of genome-wide copy number
aberrations in colorectal cancer reveals novel oncogenes and
patterns of alterations. PLoS One. 7:e420012012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Moniz S and Jordan P: Emerging roles for
WNK kinases in cancer. Cell Mol Life Sci. 67:1265–1276. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kew MC: Epidemiology of chronic hepatitis
B virus infection, hepatocellular carcinoma, and hepatitis B
virus-induced hepatocellular carcinoma. Pathol Biol (Paris).
58:273–277. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang X, Zhang H and Ye L: Effects of
hepatitis B virus X protein on the development of liver cancer. J
La Clin Med. 147:58–66. 2006.
|
|
25
|
Khaar E, Mukherji A and Kumar V: Akt
augments the oncogenic potential of the HBx protein of hepatitis B
virus by phosphorylation. FEBS J. 279:1220–1230. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hsieh A, Kim HS, Lim SO, Yu DY and Jung G:
Hepatitis B viral X protein interacts with tumor suppressor
adenomatous polyposis coli to activate Wnt/β-catenin signaling.
Cancer Lett. 300:162–172. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cha MY, Kim CM, Park YM and Ryu WS:
Hepatitis B virus X protein is essential for the activation of
Wnt/beta-catenin signaling in hepatoma cells. Hepatology.
39:1683–1693. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Waris G, Huh KW and Siddiqui A:
Mitochondrially associated hepatitis B virus X protein
constitutively activates transcription factors STAT-3 and NF-kappa
B via oxidative stress. Mol Cell Biol. 21:7721–7730. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Teng J, Wang X, Xu Z and Tang N:
HBx-dependent activation of Twist mediates STAT3 control of
epithelium-mesenchymal transition of liver cells. J Cell Biochem.
114:1097–1104. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Thorpe LM, Yuzugullu H and Zhao JJ: PI3K
in cancer: Divergent roles of isoforms, modes of activation and
therapeutic targeting. Nat Rev Cancer. 15:7–24. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fruman DA and Rommel C: PI3K and cancer:
Lessons, challenges and opportunities. Nat Rev Drug Discov.
13:140–156. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Eser S, Reiff N, Messer M, Seidler B,
Gottschalk K, Dobler M, Hieber M, Arbeiter A, Klein S, Kong B, et
al: Selective requirement of PI3K/PDK1 signaling for Kras
oncogene-driven pancreatic cell plasticity and cancer. Cancer cell.
23:406–420. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Miller TW, Rexer BN, Garrett JT and
Arteaga CL: Mutations in the phosphatidylinositol 3-kinase pathway:
Role in tumor progression and therapeutic implications in breast
cancer. Breast Cancer Res. 13:2242011. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Raimondi C and Falasca M: Targeting PDK1
in cancer. Curr Med Chem. 18:2763–2769. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jin YM, Yun C, Park C, Wang HJ and Cho H:
Expression of hepatitis B virus X protein is closely correlated
with the high periportal inflammatory activity of liver diseases. J
Viral Hepat. 8:322–330. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Scortegagna M, Lau E, Zhang T, Feng Y,
Sereduk C, Yin H, De SK, Meeth K, Platt JT, Langdon CG, et al: PDK1
and SGK3 contribute to the growth of BRAF-mutant melanomas and are
potential therapeutic targets. Cancer Res. 75:1399–1412. 2015.
View Article : Google Scholar : PubMed/NCBI
|