Introduction
PI3K/AKT signaling pathway plays an important role
in the development of numerous types of tumors as it is involved in
cell growth, proliferation and apoptosis (1–3). There are
several pathways that PI3K/AKT regulates its downstream targets.
PI3K/AKT indirectly inhibits mTOR to cause cell cycle arrest and
cell apoptosis (4). AKT activates
MDM2 indirectly to decrease p53 level and activity, and promote p53
translation and protein stability (5–7).
Metformin is a biguanide used as an oral
anti-diabetic drug, and the Food and Drug Administration (FDA)
recently has approved the first clinical study to investigate its
protective effects against numerous comorbidities. It has received
attention as a potential therapeutic agent for cancer treatment
(8,9).
Trametinib and paclitaxel are well-known chemotherapeutic drugs,
which are frequently used to treat breast and lung cancers, and
both have synergistic effects when used in combination with
metformin (4,10). Metformin exerts anti-proliferative
activity on glioblastoma cells by inhibiting AKT signaling
(11). Metformin induces p53 activity
by regulating AMPK/mTOR signaling pathway (10,12) and
activates AMPK to phosphorylate and inactivate MDMX, finally
results in stabilization and activation of p53 in cancer cells
(13). As a major p53 repressor, MDMX
overexpresses in many human cancers, which inhibits DNA repair
signals and promotes transformation to contribute to tumorigenesis.
When MDMX is inhibited, the activity of p53 is enhanced and p53
degradation is blocked (14).
Flavonoids are safe and easily obtainable by
extraction from natural plants. Many of them are part of human
daily diet (15,16). Flavonoids inhibit PI3K/AKT pathway to
induce apoptosis (17,18). Tangeretin inhibits growth of HL-60
cells and induces apoptosis without causing serious side-effect on
immune cells (19). Luteolin has low
incidence of side effects in therapeutic potential (20). Pre-treatment with cardamonin
significantly attenuates nephrotoxic effects, oxidative stress,
inflammation and decreases caspase3 expression and Bax/Bcl-2 ratio
induced by cisplatin (21). Flavone
(2-phenyl-4H-1-benzopyran-4-one) is the core structure of
flavonoids, which induces apoptosis potent in human colon carcinoma
cells (22,23). Flavone inhibits PI3K/AKT and increases
FoxO3a activation to induce apoptosis in breast cancer cells
(24).
The PI3K/AKT pathway that metformin mediated with or
without flavonoids remains to be fully determined in breast cancer
cells. Inhibition of PI3K/AKT pathway may gain importance as
potential mono- and combination therapies in the clinical trials.
Therefore, our study was designed to investigate whether flavone
enhances cell apoptosis induced by metformin and whether the
combination shares a common intracellular signal pathway that
regulates MDMX/p53 via PI3K/AKT in breast cancer cells.
Materials and methods
Reagents and antibodies
Metformin (MET) and flavone (FLA) were purchased
from Sigma-Aldrich (St. Louis, MO, USA). Antibodies against PI3K
(cat. no. 4249S), p-AKT(ser-473) (cat. no. 4060S) and caspase3
(cat. no. 8665P) were from Cell Signaling Technology (Danvers, MA,
USA); MDMX (cat. no. sc-14738) from Santa Cruz Biotechnology
(Dallas, TX, USA); p53 (cat. no. BS1272) and Bax (cat. no. BS1030)
from Bioworld Technology, Inc. (St. Louis Park, MN, USA). AKT1
(cat. no. BF0570), Bcl-2 (cat. no. AF0769) and cleaved caspase3
(cat. no. AF7022) were from Affinity Biosciences (Cincinnati, OH,
USA); β-actin (cat. no. 60008-1-lg) was from ProteinTech Group,
Inc. (Chicago, IL, USA). PI3K inhibitor (LY294002) was obtained
from Beyotime Institute of Biotechnology (Haimen, China).
Cell culture
MCF-10A human breast epithelial cells, MDA-MB-231
and MCF-7 human breast cancer cells were from American Type Culture
Collection (Manassas, VA, USA). Cells were cultured in Dulbecco's
modified Eagle's medium supplemented with 10% fetal calf serum, and
maintained at 37°C in humidified atmosphere with 5%
CO2.
MTT assay
MTT assay is based on the conversion of MTT
(Amresco, LLC, Solon, OH, USA) to formazan crystals by
mitochondrial dehydrogenases. Briefly, MDA-MB-231 cells were plated
in 96-well plates at a density of 1×104 cells per well
for 16 h. The cells were respectively treated with flavone and
metformin under various concentrations (100, 150, 200, 250 µM) and
(5, 10, 15, 20, 25 mM) for 24 h to select a suitable inhibitory
concentration. Similarly, the cells were treated with 10 mM
metformin, 10 mM metformin:100 µM flavone, 10 mM metformin:200 µM
flavone; and 20 mM metformin, 20 mM metformin:100 µM flavone, 20 mM
metformin:200 µM flavone for 24 h. Then MTT solution (5 mg/ml) was
added (20 µl/well) to the precipitate and incubated at 37°C for 4
h. The solution was aspirated, and the formazan crystals were
dissolved with 150 µl DMSO (Amresco, LLC). The absorbance was
measured at 492 nm by an enzyme-linked immunosorbent assay reader
(Awareness Technology, Inc., Palm City, FL, USA).
Hoechst/propidium iodide (PI)
staining
MCF-10A, MDA-MB-231 and MCF-7 cells were exposed to
20 mM metformin, 20 mM metformin:100 µM flavone for 24 h and
stained with Hoechst 33342 and PI (Beyotime Institute of
Biotechnology) for 20–30 min. The nuclei of apoptotic cells were
observed visually by fluorescence microscope. The cells were
differentiated with normal live cells (exhibited light blue),
apoptotic cells (exhibited bright blue) and necrotics cells
(exhibited red).
Western blot analysis
The cells were treated with 20 mM metformin, 100 µm
flavone and 20 mM metformin:100 µM flavone for 24 h. Protein
concentration was determined by the BCA Assay (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). An equal amount of protein
from each sample was loaded per lane, separated by SDS-PAGE and
transferred onto a polyvinylidene difluoride membrane. The membrane
was blocked in 5% milk for 90 min at room temperature, incubated
with primary antibodies overnight at 4°C, then washed 3 times and
incubated with appropriate HRP-conjugated secondary antibodies.
After washing, protein expression was detected with the ECL system
(Beyotime Institute of Biotechnology).
Statistical analysis
Each experiment was analysed in three biological
replicates. Data were expressed as mean ± standard deviation.
Multiple comparisons between the groups have been performed using
S-N-K method after ANOVA. P-values of <0.05 were considered
statistically significant.
Results
Combined metformin and flavone
inhibits cell viability, and induces apoptosis
MDA-MB-231 cells were treated by 0, 5, 10, 15, 20
and 25 mM metformin for 24 h, the inhibition rates on cell
viability were 0, 7.64±5.90, 13.51±0.35, 13.10±0.39, 20.93±1.038
and 24.20±0.012%, the inhibitory effects on cell viability
increased with the extension of treatment concentrations, and cells
were sensitive to 10/20 mM metformin treatment. The inhibition
rates were 0, 3.83±7.30, 9.15±8.14, 18.97±2.94 and 19.67±3.78%,
respectively when treated by 0, 100, 150, 200, and 250 µM flavone
for same time. Cell viability was inhibited by 150, 200 and 250 µM
flavone and was not significantly effected by 100 µM flavone
(P<0.05 and P<0.01; Fig. 1A).
The concentrations of 10 and 20 mM metformin were chosen for the
combination treatment. When 100/200 µM flavone was supplemented to
the cells which were treated with 10 mM metformin for 24 h, the
inhibition rates increased to 24.3±1.5 and 24.9±1.5% respectively
compared to an inhibition rate of 15.7±0.2% with 10 mM metformin
only. Similarly, when 100/200 µM flavone was added to the cells
treated with 20 mM metformin, the inhibition rates became higher
(34.0±1.9 and 36.8±1.4%, respectively) than 20 mM metformin only
(24.6±2.5%). The results demonstrated that combination treatment of
metformin and flavone increased significantly inhibition rates to
the breast cancer cells (P<0.05 and P<0.01; Fig. 1B). The inhibition rates were higher
when the cells treated by 20 mM metformin combined with 100/200 µM
flavone than that by 10 mM metformin with same concentration
flavone. Meanwhile, the inhibition rates had no change when the
cells co-treated by 10/20 mM metformin with 100/200 µM flavone
respectively. The combination of 20 mM metformin:100 µM flavone was
used for the following experiments.
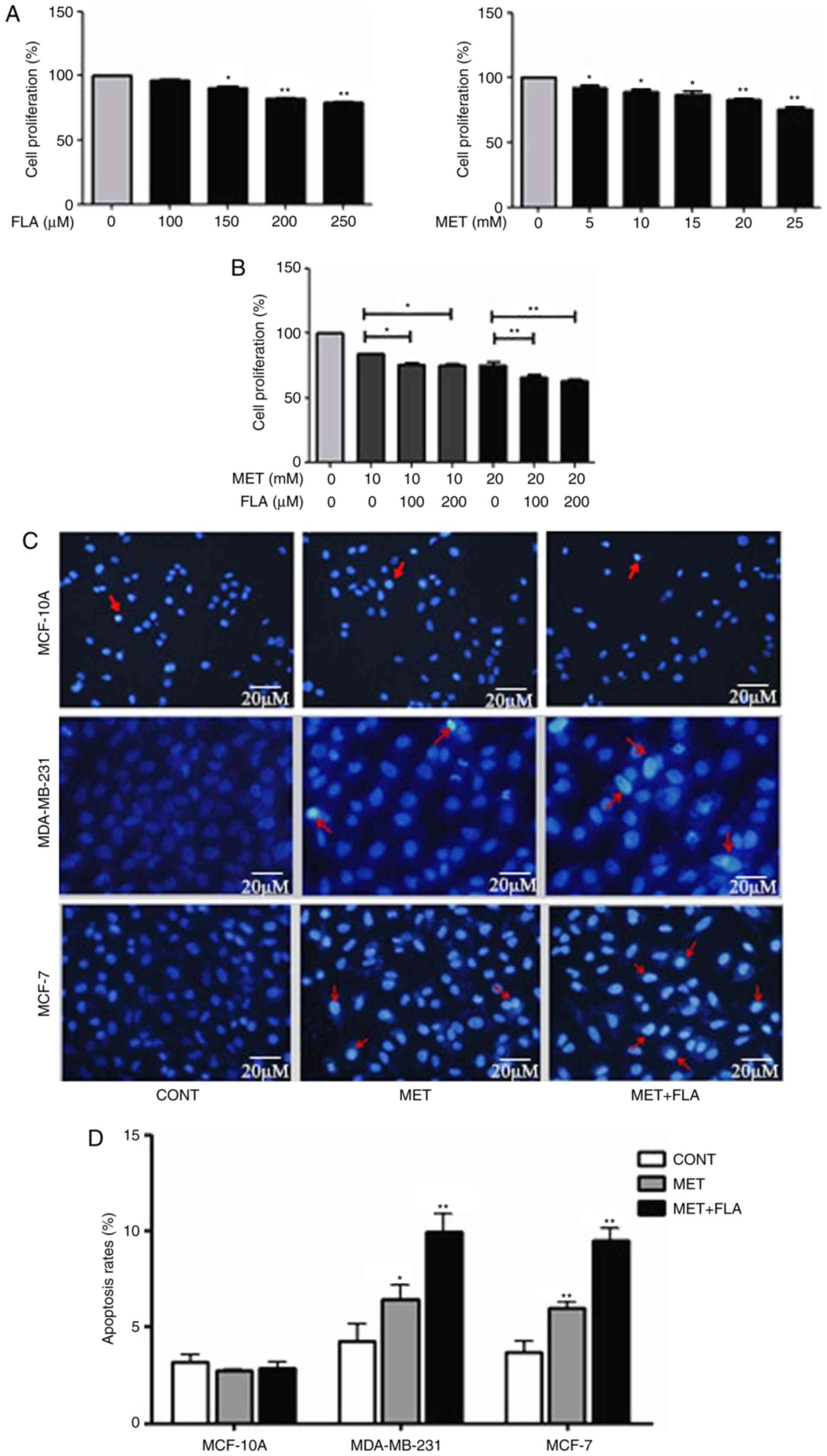 | Figure 1.The combination of metformin and
flavone inhibited cell viability and induced apoptosis in breast
cancer cells with a concentration dependent manner. (A) MDA-MB-231
cell viability was measured after treatment with FLA (0, 100, 150,
200, 250 µM) (*P<0.05, 150 µM FLA vs. control; **P<0.01, 200
µM FLA vs. control, 250 µM FLA vs. control) and MET under various
concentrations (0, 5, 10, 15, 20, 25 mM) (*P<0.05, 5 mM MET vs.
control, 10 mM MET vs. control, 15 mM MET vs. control; **P<0.01,
20 mM MET vs. control, 25 mM MET vs. control) for 24 h. (B)
MDA-MB-231 cells were treated by 10 mM MET, 10 mM MET:100 µM FLA,
10 mM MET:200 µM FLA and treated by 20 mM MET, 20 mM MET:100 µM
FLA, 20 mM MET:200 µM FLA for 24 h (*P<0.05, 10 mM MET:100 µM
FLA vs. 10 mM MET, 10 mM MET:200 µM FLA vs. 10 mM MET; **P<0.01,
20 mM MET:100 µM FLA vs. 20 mM MET, 20 mM MET:200 µM FLA vs. 20 mM
MET). (C) Representative photomicrographs of MCF-10A, MDA-MB-231
cells stained with Hoechst 33342 and propidium iodide fluorescent
dye after incubated in the treatment with 20 mM MET, 20 mM MET:100
µM FLA for 24 h. The arrows show the apoptotic cells distinguished
by condensed or fragmented nuclei. (D) The quantitative results of
apoptosis rates with different treatment (*P<0.05, 20 mM MET vs.
control; **P<0.01, 20 mM MET:100 µM FLA vs. control) and MCF-7
(**P<0.01, 20 mM MET vs. control, 20 mM MET:100 µM FLA vs.
control). FLA, flavone; MET, metformin. |
To investigate effects of metformin and flavone on
cell apoptosis, Hoechst and PI nuclear staining assay were
performed. The apoptosis rates were 2.71±0.81 and 2.86±0.79% in
MCF-10A, 6.43±0.78 and 9.96±0.94% in MDA-MB-231 and 5.89±1.02 and
9.37±0.86% in MCF-7 after metformin or combination treatment
compared to 3.16±0.95, 4.23±0.90 and 3.46±0.80% of control groups,
respectively (P<0.05 and P<0.01; Fig. 1C and D). Our results revealed that
MCF-10A cells were less sensitive to metformin or
combination-induced apoptosis than MDA-MB-231 or MCF-7 cells
respectively. Metformin and co-treatment increased breast cancer
cell apoptosis but barely affected on MCF-10A cells, and the higher
apoptosis rate appeared by co-treatment than metformin treatment in
MDA-MB-231 and MCF-7 cells.
Combined metformin and flavone
synergistically inhibits the PI3K/AKT pathway
In MDA-MB-231 and MCF-7 cells, the results of
western blotting showed that metformin, flavone and co-treatment
had no effect on AKT1 expression, but metformin decreased PI3K and
p-AKT(ser-473) protein levels compared with control group. The
combination of metformin and flavone significantly decreased
expressions of PI3K and p-AKT compared to metformin or flavone
single-treatment (P<0.05 and P<0.01; Fig. 2). These results showed that induction
of cell apoptosis may involve modulation of the PI3K/AKT signaling
via the combination treatment.
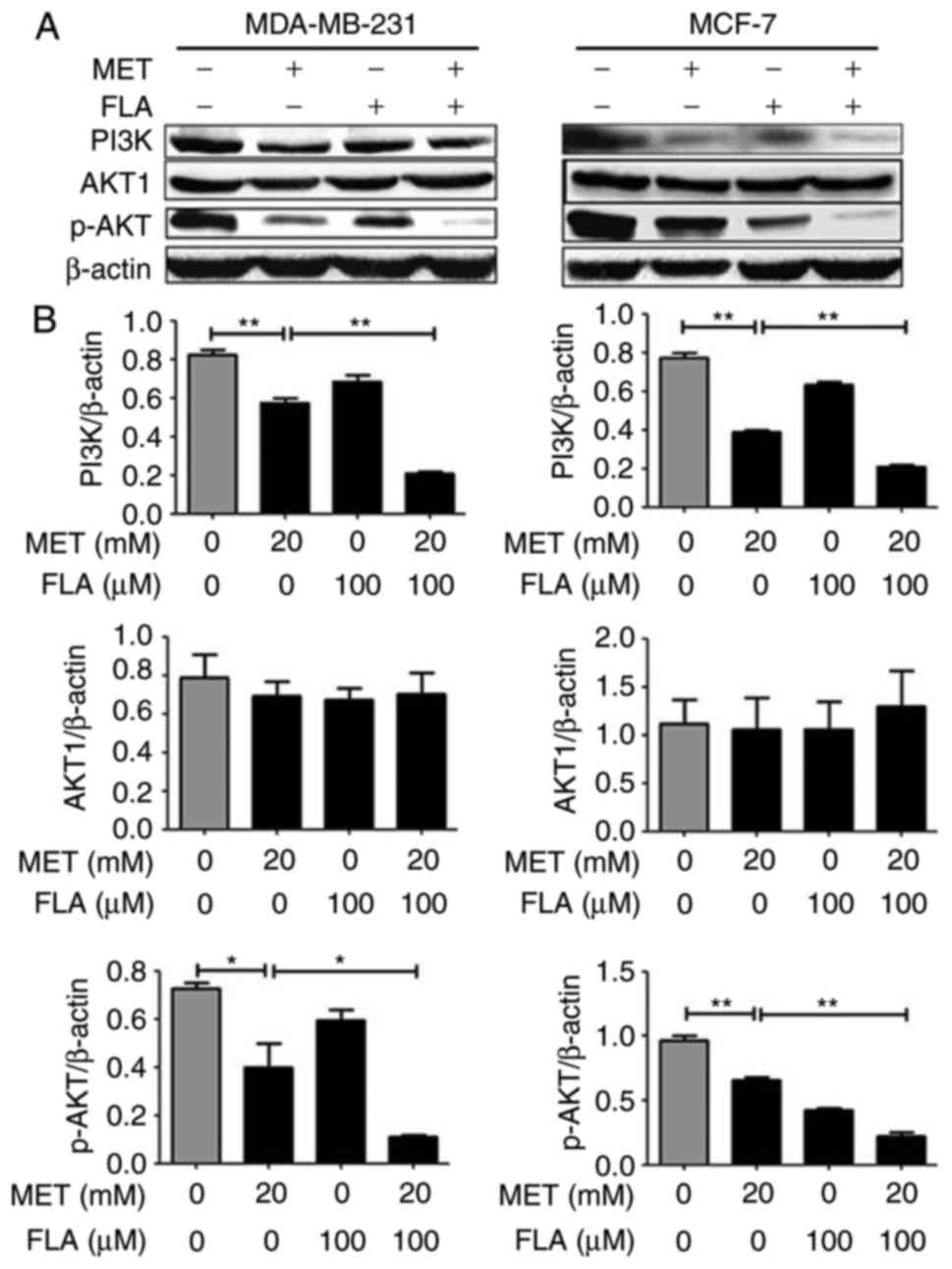 | Figure 2.The combination of metformin and
flavone synergistically inhibited the PI3K/AKT pathway in breast
cancer cells. (A) Protein levels for PI3K, AKT and p-AKT(ser-473)
were detected with western blotting when MDA-MB-231 cells (left
column) were treated with 0, 20 mM MET, 100 µM FLA and the
combination of 20 mM MET:100 µM FLA for 24 h, and MCF-7 cells
(right column) were the same treatment to test PI3K, AKT and
p-AKT(ser-473) expressions, (B) and their densitometry results over
β-actin at least three separate experiments. **P<0.01, 20 mM MET
vs. control, 20 mM MET:100 µM FLA vs. 20 mM MET; *P<0.05, 20 mM
MET vs. control, 20 mM MET:100 µM FLA vs. 20 mM MET. PI3K,
phosphatidylinositol 3-kinase; AKT, serine/threonine kinase; FLA,
flavone; MET, metformin. |
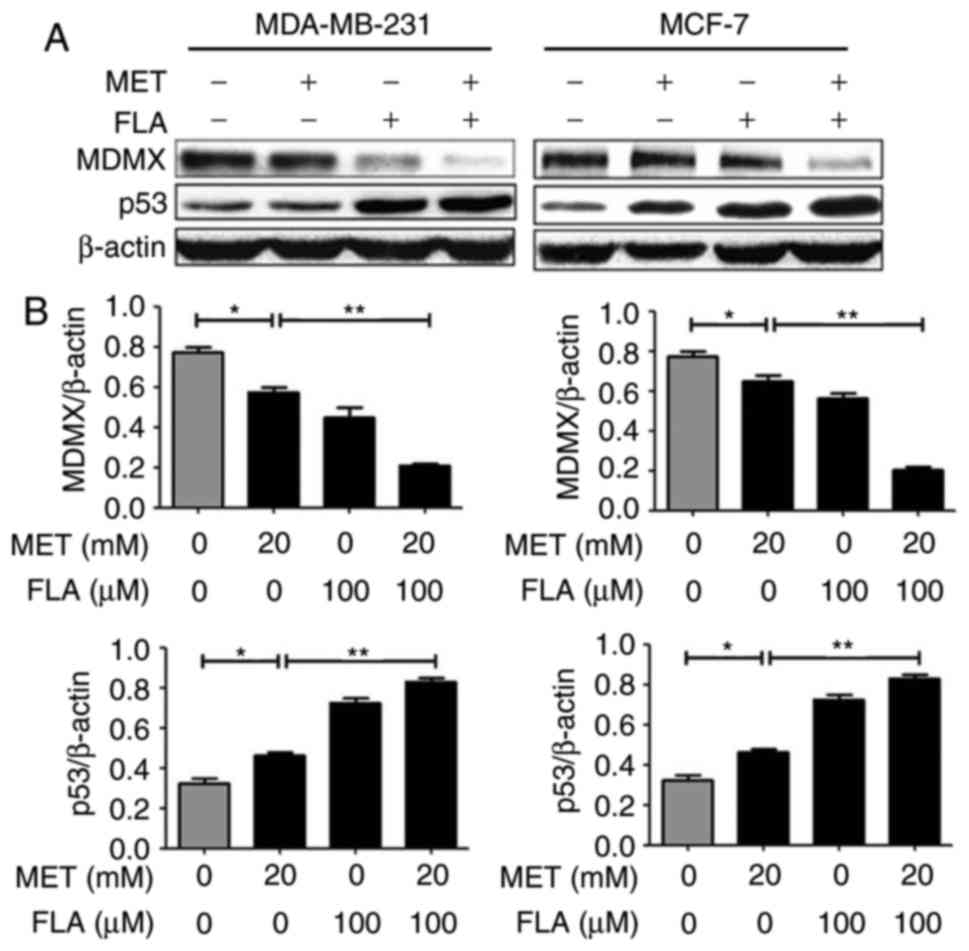
PI3K/AKT pathway is associated with
MDMX/p53 regulation for treatment with the combination of metformin
and flavone
When two cell lines were treated with metformin or
flavone alone, MDMX expression decreased and p53 increased. When
flavone was supplemented together into the cells, the effects of
metformin were potentiated (P<0.05 and P<0.01; Fig. 3). These results suggested that the
combination induced apoptosis by reducing MDMX and up-regulating
p53 level.
To further examine the relationship of p53 with
PI3K/AKT pathway after the combination treatment, PI3K inhibitor
LY294002 was used to examine p53 expression. An increase in p53
protein level appeared when the cells were treated with combination
or LY294002 compared to control group. p53 levels increased
significantly when LY294002 was added to co-treatment compared to
combination group or LY294002 in MDA-MB-231 and MCF-7 cells
(P<0.05 and P<0.01; Fig.
4).
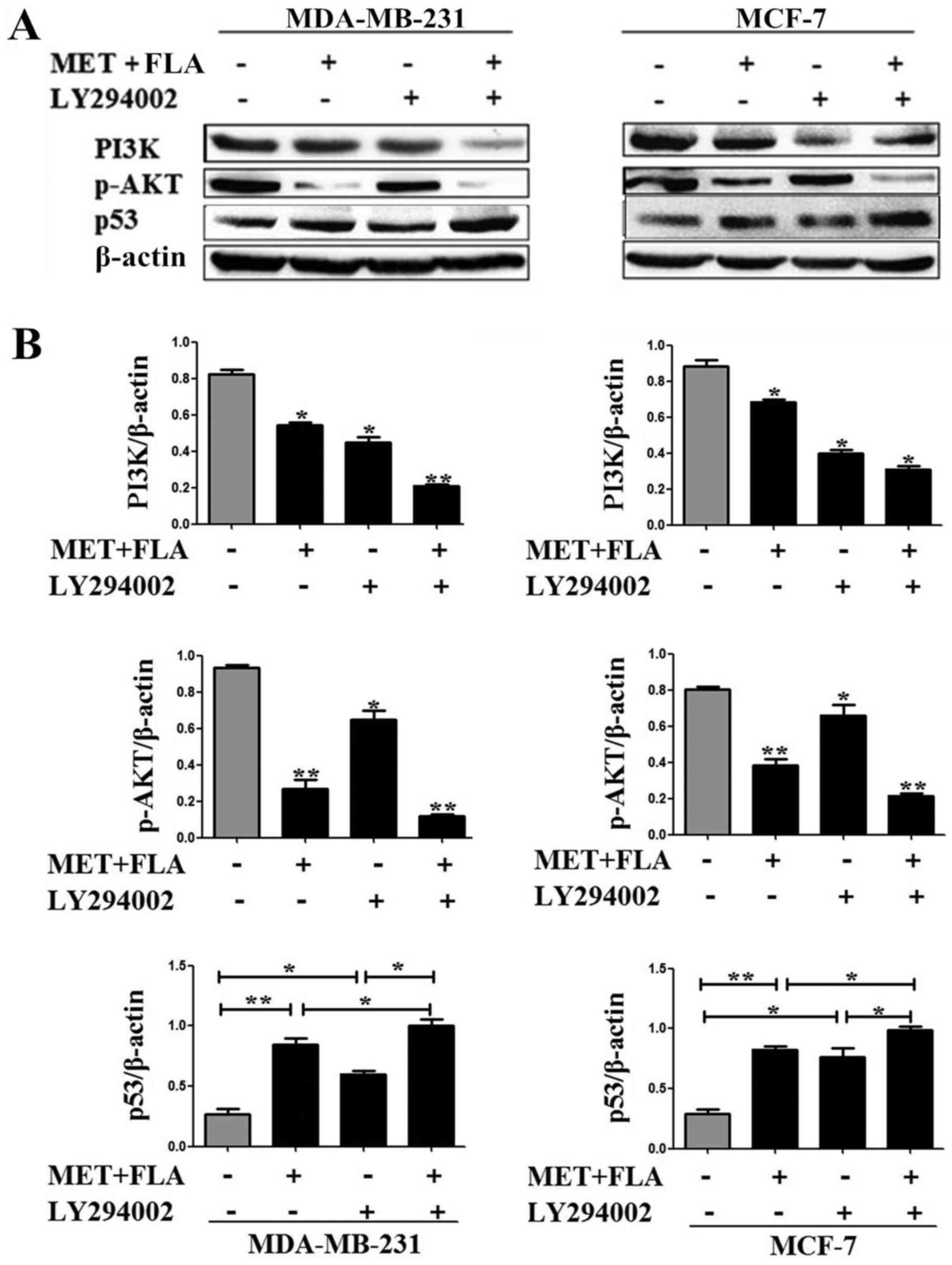 | Figure 4.PI3K/AKT pathway was associated with
MDMX/p53 regulation treated with the combination of metformin and
flavone. (A) Expressions of PI3K (*P<0.05, combination vs.
control, LY294002 vs. control; **P<0.01, the combination and
LY294002 vs. control), p-AKT(ser-473) (*P<0.05, LY294002 vs.
control; **P<0.01 combination vs. control, the combination and
LY294002 vs. control) and p53 (*P<0.05, LY294002 vs. control,
the combination and LY294002 vs. combination, the combination and
LY294002 vs. LY294002; **P<0.01, combination vs. control) with
0, the combination, LY294002, the combination and LY294002
treatment in MDA-MB-231 cells. MCF-7 cells were the same treatment
to detect PI3K (*P<0.05, combination vs. control, Ly294002 vs.
control, the combination and LY294002 vs. control), p-AKT(ser-473)
(*P<0.05, LY294002 vs. control; **P<0.01, combination vs.
control, the combination and LY294002 vs. control) and p53
(*P<0.05, LY294002 vs. control, the combination and LY294002 vs.
combination, the combination and LY294002 vs. LY294002;
**P<0.01, combination vs. control) expression. (B) Bar graphs
showed their relative levels to β-actin. PI3K, phosphatidylinositol
3-kinase; AKT, serine/threonine kinase; MDMX, murine double minute
X; FLA, flavone; MET, metformin. |
Combined metformin and flavone
regulates the expression of apoptosis-associated proteins
Bcl-2 and Bax are p53-dependent target genes. Our
results showed that metformin, flavone and co-treatment decreased
Bcl-2, and increased Bax as well as cleaved caspase3 expressions,
but it had no effect on caspase3 level. Combination group showed
higher expression of cleaved caspase3 than metformin or flavone
group. These results showed that metformin, flavone and
co-treatment induced apoptosis via apoptotic protein Bcl-2, Bax and
caspase pathway in breast cancer cells (P<0.05 and P<0.01;
Fig. 5).
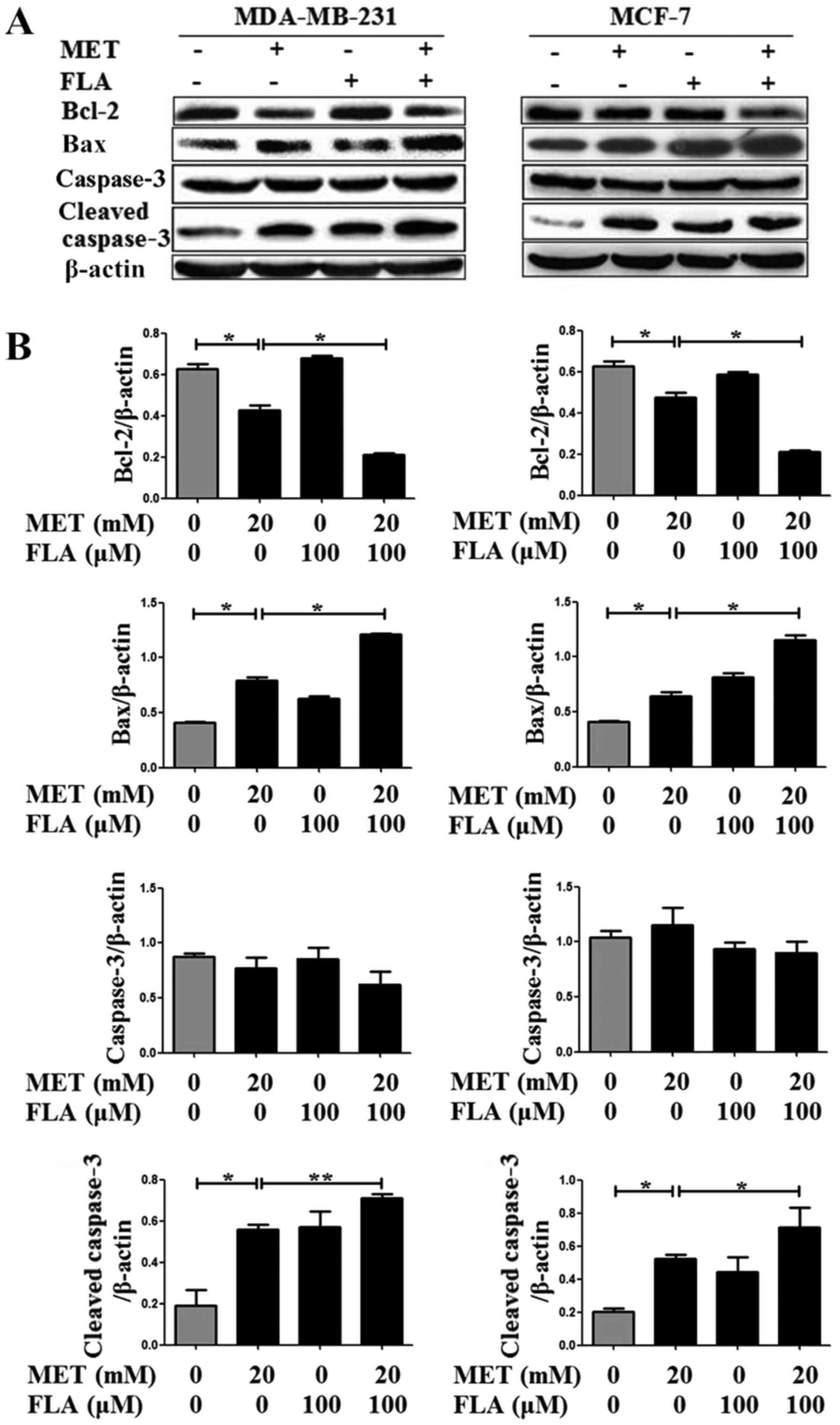 | Figure 5.The combination of metformin and
flavone regulated downstream targets of p53. (A) Protein levels of
Bcl-2, Bax were detected with western blotting when MDA-MB-231 and
MCF-7 cells were treated with 0, 20 mM MET, 100 µM FLA and the
combination of 20 mM MET:100 µM FLA for 24 h (*P<0.05, 20 mM MET
vs. control, 20 mM MET:100 µM FLA vs. 20 mM MET), caspase3 were
detected no significance and cleaved caspase3 were detected when
MDA-MB-231 (*P<0.05, 20 mM MET vs. control; **P<0.01, 20 mM
MET:100 µM FLA vs. 20 mM MET) and MCF-7 cells (*P<0.05, 20 mM
MET vs. control, 20 mM MET:100 µM FLA vs. 20 mM MET) were the same
treatment, (B) and their densitometry results over β-actin in at
least three separate experiments. FLA, flavone; MET, metformin. |
Discussion
Metformin is an oral anti-diabetic drug and has a
well-known safety profile. It interferes with PI3K/AKT pathway to
induce apoptosis in many cancer cells (11,25,26). AKT
can activate eIF4E by inhibiting mTOR to enhance cyclin D1,
resulting in cell cycle arrest in G1 phase (27,28). As a
novel AMPK activator, metformin increases nuclear accumulation and
protein stability of FoxO3a by inhibiting AKT-MDM2 signaling
pathway to reduce invasive and metastatic capacity of aggressive
cancer cells (29). Therefore,
PI3K/AKT pathway plays a significant role in anticancer
effects.
Flavone inhibits PI3K/AKT pathway to involve in the
progression and apoptosis of cancer cells (18,30,31). Our
previous results have presented that 100 µM flavone reduced
viability slightly in breast cancer cells (32). We herein showed that metformin
monotherapy had low inhibition rates on cell viability and
apoptosis (Fig. 1A and C). The
addition of flavone to metformin led to enhance anti-cancer effects
of metformin (Fig. 1B and C).
Apoptosis rates are 27 and 37% for the cells treated with flavone
compared to 13 and 25% for control group respectively in MDA-MB-231
and MCF-7 cells (24). Apoptosis is
increased about 2.4- to 3.1-fold induced by flavonoid for 36 h in
LNCaP cells (33). Our previous study
showed that apoptotic rate was 15.15±0.20% when treated by 200 µM
flavone compared to 4.90±1.23% of control group in breast cancer
cells (32). Apoptosis rates in our
results increased 1.5- to 3-fold. It is consistent with others'
results. Similarly, metformin decreased PI3K and p-AKT(ser-473)
protein expressions compared with control group. When the cancer
cells were treated with the combination, PI3K and p-AKT(ser-473)
protein levels became lower than those in metformin treatment
(Fig. 2). Therefore, the combination
treatment significantly decreased expressions of PI3K and
p-AKT(ser-473). MCF-7 cells represent an ERa positive model and
MDA-MB-231 ERa negative. Both cells are PTEN inactive mutation
(34). In addition, mTORC2 can
regulate AKT phosphorylation at S473 and induce cellular signal
pathway in human wild-type PTEN or PTEN-null cancer cell lines
(35). Therefore, the constitutive
AKT activation after PI3K inhibitor treatment might due to
deregulated p-AKT level by mTORC2 inhibition with co-treatment
other than dependence on PI3K only.
MDMX binds to p53 and is involved in inhibiting p53
transactivation activity, which represses function of p53 and
destabilizes the protein in breast cancer cells (14,36). Our
data indicated that metformin reduced MDMX level and activated p53
to regulate apoptosis-related proteins. It is in accordance with a
previous result that metformin phosphorylates and inactivates MDMX
to activate p53 in HCT116 colon cancer cells (13). The combination of metformin and
flavone markedly inhibited MDMX and activated p53 compared to
metformin treatment only. Flavone supplement enhanced the
regulation of metformin on MDMX and p53 (Fig. 3). When MDMX was inhibited, p53
expression was enhanced, which showed that the combination
increased p53 level through inhibiting MDMX. Some reports suggest
that PI3K/AKT/p53 was activated by mammalian reovirus to inhibit
virus replication/infection (37). A
p53 target gene, Sesn2, is independent of p53 but requires the
PI3K/AKT pathway to protect cells against oxidative and genotoxic
stresses (38). Pacilitaxel can
promote apoptosis and growth inhibition of human nasopharyngeal
cancer cell line CNE2 by suppressing PI3K, p-AKT and increasing p53
level (39). However, the molecular
mechanism of PI3K/AKT pathway regulates MDMX/p53 is presently
unclear when the cancer cells were treated with combination of
metformin and flavone. In our study, p53 expression significantly
increased after co-treatment and LY294002 treatment (Fig. 4), indicating that p53 was the
downstream of PI3K and the combination increased expression of p53
by inhibiting PI3K/AKT pathway. Although the mechanism was not
fully understood, it brings unexpected insight into
PI3K/AKT-mediated regulation of MDMX/p53 on apoptosis by
co-treatment in breast cancer cells.
p53 directly activates apoptosis-related Bax to
induce release of cytochrome c from mitochondria, and then
activates caspase (40). In addition,
p53 reduces Bcl-2 transcription (41). Flavones induce caspase-dependent
apoptosis in A549 cells (42) and
primary human neutrophils (43). Our
present study showed that metformin, flavone and combination
induced apoptosis by decreasing Bcl-2, increasing Bax and cleaved
caspase3 protein level.
Taken together, it is the first time for the
combination of metformin and flavone to inhibit cell viability and
to induce apoptosis more effectively than metformin and flavone
alone in breast cancer cells. The combination potentiated apoptosis
by decreasing MDMX and increasing p53 expression via PI3K/AKT
signaling pathway. Further studies should be performed to fully
define the action of the combination and underlying mechanism in
vivo, and this combination will be a promising approach for
clinical cancer therapy.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 31672377), Major Key
Technology of Science and Technology in Shandong Province
(2015ZDJS04003), the Key Program of Shandong Provincial Natural
Science Foundation of China (ZR2013CZ002), Science and Technology
Program of Jinan (201202033).
References
|
1
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-kinase-AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu Y, Zhang Y, Jia K, Dong Y and Ma W:
Metformin inhibits the proliferation of A431 cells by modulating
the PI3K/Akt signaling pathway. Exp Ther Med. 9:1401–1406. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pappalardo F, Russo G, Candido S, Pennisi
M, Cavalieri S, Motta S, McCubrey JA, Nicoletti F and Libra M:
Computational modeling of PI3K/AKT and MAPK signaling pathways in
melanoma cancer. PLoS One. 11:e01521042016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vujic I, Sanlorenzo M, Posch C,
Esteve-Puig R, Yen AJ, Kwong A, Tsumura A, Murphy R, Rappersberger
K and Ortiz-Urda S: Metformin and trametinib have synergistic
effects on cell viability and tumor growth in NRAS mutant cancer.
Oncotarget. 6:969–978. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou BP, Liao Y, Xia W, Zou Y, Spohn B and
Hung MC: HER-2/neu induces p53 ubiquitination via Akt-mediated MDM2
phosphorylation. Nat Cell Biol. 3:973–982. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ashcroft M, Ludwig RL, Woods DB, Copeland
TD, Weber HO, MacRae EJ and Vousden KH: Phosphorylation of HDM2 by
Akt. Oncogene. 21:1955–1962. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dong C, Zhao B, Long F, Liu Y, Liu Z, Li
S, Yang X, Sun D, Wang H, Liu Q, et al: Nogo-B receptor promotes
the chemoresistance of human hepatocellular carcinoma via the
ubiquitination of p53 protein. Oncotarget. 7:8850–8865. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Decensi A, Puntoni M, Goodwin P, Cazzaniga
M, Gennari A, Bonanni B and Gandini S: Metformin and cancer risk in
diabetic patients: A systematic review and meta-analysis. Cancer
Prev Res (Phila). 3:1451–1461. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Franciosi M, Lucisano G, Lapice E,
Strippoli GF, Pellegrini F and Nicolucci A: Metformin therapy and
risk of cancer in patients with type 2 diabetes: Systematic review.
PLoS One. 8:e715832013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rocha GZ, Dias MM, Ropelle ER,
Osório-Costa F, Rossato FA, Vercesi AE, Saad MJ and Carvalheira JB:
Metformin amplifies chemotherapy-induced AMPK activation and
antitumoral growth. Clin Cancer Res. 17:3993–4005. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Würth R, Pattarozzi A, Gatti M, Bajetto A,
Corsaro A, Parodi A, Sirito R, Massollo M, Marini C, Zona G, et al:
Metformin selectively affects human glioblastoma tumor-initiating
cell viability: A role for metformin-induced inhibition of Akt.
Cell Cycle. 12:145–156. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Song CW, Lee H, Dings RP, Williams B,
Powers J, Santos TD, Choi BH and Park HJ: Metformin kills and
radiosensitizes cancer cells and preferentially kills cancer stem
cells. Sci Rep. 2:3622012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He G, Zhang YW, Lee JH, Zeng SX, Wang YV,
Luo Z, Dong XC, Viollet B, Wahl GM and Lu H: AMP-activated protein
kinase induces p53 by phosphorylating MDMX and inhibiting its
activity. Mol Cell Biol. 34:148–157. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang H, Ma X, Ren S, Buolamwini JK and Yan
C: A small-molecule inhibitor of MDMX activates p53 and induces
apoptosis. Mol Cancer Ther. 10:69–79. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ravishankar D, Rajora AK, Greco F and
Osborn HM: Flavonoids as prospective compounds for anti-cancer
therapy. Int J Biochem Cell Biol. 45:2821–2831. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sak K: Cytotoxicity of dietary flavonoids
on different human cancer types. Pharmacogn Rev. 8:122–146. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim SJ, Kim HJ, Kim HR, Lee SH, Cho SD,
Choi CS, Nam JS and Jung JY: Antitumor actions of baicalein and
wogonin in HT-29 human colorectal cancer cells. Mol Med Rep.
6:1443–1449. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Koosha S, Alshawsh MA, Looi CY, Seyedan A
and Mohamed Z: An association map on the effect of flavonoids on
the signaling pathways in colorectal cancer. Int J Med Sci.
13:374–385. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hirano T, Abe K, Gotoh M and Oka K: Citrus
flavone tangeretin inhibits leukaemic HL-60 cell growth partially
through induction of apoptosis with less cytotoxicity on normal
lymphocytes. Br J Cancer. 72:1380–1388. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Iakovleva I, Begum A, Pokrzywa M,
Walfridsson M, Sauer-Eriksson AE and Olofsson A: The flavonoid
luteolin, but not luteolin-7-O-glucoside, prevents a transthyretin
mediated toxic response. PLoS One. 10:e01282222015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
El-Naga RN: Pre-treatment with cardamonin
protects against cisplatin-induced nephrotoxicity in rats: Impact
on NOX-1, inflammation and apoptosis. Toxicol Appl Pharmacol.
274:87–95. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wenzel U, Kuntz S, Brendel MD and Daniel
H: Dietary flavone is a potent apoptosis inducer in human colon
carcinoma cells. Cancer Res. 60:3823–3831. 2000.PubMed/NCBI
|
|
23
|
Herzog A, Kindermann B, Döring F, Daniel H
and Wenzel U: Pleiotropic molecular effects of the pro-apoptotic
dietary constituent flavone in human colon cancer cells identified
by protein and mRNA expression profiling. Proteomics. 4:2455–2464.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin CH, Chang CY, Lee KR, Lin HJ, Chen TH
and Wan L: Flavones inhibit breast cancer proliferation through the
Akt/FOXO3a signaling pathway. BMC cancer. 15:9582015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Quinn BJ, Kitagawa H, Memmott RM, Gills JJ
and Dennis PA: Repositioning metformin for cancer prevention and
treatment. Trends Endocrinol Metab. 24:469–480. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cerezo M, Tomic T, Ballotti R and Rocchi
S: Is it time to test biguanide metformin in the treatment of
melanoma? Pigment Cell Melanoma Res. 28:8–20. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Manning BD and Cantley LC: AKT/PKB
signaling: Navigating downstream. Cell. 129:1261–1274. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mamane Y, Petroulakis E, Rong L, Yoshida
K, Ler LW and Sonenberg N: eIF4E-from translation to
transformation. Oncogene. 23:3172–3179. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chou CC, Lee KH, Lai IL, Wang D, Mo X,
Kulp SK, Shapiro CL and Chen CS: AMPK reverses the mesenchymal
phenotype of cancer cells by targeting the Akt-MDM2-Foxo3a
signaling axis. Cancer Res. 74:4783–4795. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Franzen CA, Amargo E, Todorović V, Desai
BV, Huda S, Mirzoeva S, Chiu K, Grzybowski BA, Chew TL, Green KJ
and Pelling JC: The chemopreventive bioflavonoid apigenin inhibits
prostate cancer cell motility through the focal adhesion kinase/Src
signaling mechanism. Cancer Prev Res (Phila). 2:830–841. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shukla S, Bhaskaran N, Babcook MA, Fu P,
Maclennan GT and Gupta S: Apigenin inhibits prostate cancer
progression in TRAMP mice via targeting PI3K/Akt/FoxO pathway.
Carcinogenesis. 35:452–460. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhu W, Yang B, Fu H, Ma L, Liu T, Chai R,
Zheng Z, Zhang Q and Li G: Flavone inhibits nitric oxide synthase
(NOS) activity, nitric oxide production and protein S-nitrosylation
in breast cancer cells. Biochem Biophys Res Commun. 458:590–595.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Seo YJ, Kim BS, Chun SY, Park YK, Kang KS
and Kwon TG: Apoptotic effects of genistein, biochanin-A and
apigenin on LNCaP and PC-3 cells by p21 through transcriptional
inhibition of polo-like kinase-1. J Korean Med Sci. 26:1489–1494.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dey N, De P and Leyland-Jones B:
PI3K-AKT-mTOR inhibitors in breast cancers: From tumor cell
signaling to clinical trials. Pharmacol Ther. 175:91–106. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sarbassov DD, Guertin DA, Ali SM and
Sabatini DM: Phosphorylation and regulation of Akt/PKB by the
rictor-mTOR complex. Science. 307:1098–1101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ling X, Xu C, Fan C, Zhong K, Li F and
Wang X: FL118 induces p53-dependent senescence in colorectal cancer
cells by promoting degradation of MdmX. Cancer Res. 74:7487–7497.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang X, Wu H, Liu C, Tian J and Qu L:
PI3K/Akt/p53 pathway inhibits reovirus infection. Infect Genet
Evol. 34:415–422. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ben-Sahra I, Dirat B, Laurent K, Puissant
A, Auberger P, Budanov A, Tanti JF and Bost F: Sestrin2 integrates
Akt and mTOR signaling to protect cells against energetic
stress-induced death. Cell Death Differ. 20:611–619. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li T: Pacilitaxel induces human
nasopharyngeal carcinoma cell line CNE2 apoptosis and growth
inhibition by suppressing PI3K/AKT/p53 signaling pathway. Lin Chung
Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 29:2147–2150. 2015.(In
Chinese). PubMed/NCBI
|
|
40
|
Chipuk JE, Kuwana T, Bouchier-Hayes L,
Droin NM, Newmeyer DD, Schule M and Green DR: Direct activation of
Bax by p53 mediates mitochondrial membrane permeabilization and
apoptosis. Science. 303:1010–1014. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Miyashita T, Krajewski S, Krajewska M,
Wang HG, Lin HK, Liebermann DA, Hoffman B and Reed JC: Tumor
suppressor p53 is a regulator of bcl-2 and bax gene expression in
vitro and in vivo. Oncogene. 9:1799–1805. 1994.PubMed/NCBI
|
|
42
|
Zhong LR, Chen X and Wei KM: Radix
tetrastigma hemsleyani flavone induces apoptosis in human lung
carcinoma a549 cells by modulating the MAPK pathway. Asian Pac J
Cancer Prev. 14:5983–5987. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lucas CD, Allen KC, Dorward DA, Hoodless
LJ, Melrose LA, Marwick JA, Tucker CS, Haslett C, Duffin R and
Rossi AG: Flavones induce neutrophil apoptosis by down-regulation
of Mcl-1 via a proteasomal-dependent pathway. FASEB J.
27:1084–1094. 2013. View Article : Google Scholar : PubMed/NCBI
|