Introduction
Gastric cancer (GC), often diagnosed at advanced
stage, is the second-leading cause of cancer-associated mortality
worldwide, although its incidence has been substantially declining
for the past decades (1,2). GC frequently invades the surrounding
tissues to spread cancer cells, leading to high mortality rates for
patients suffering from GC (3);
understanding the molecular mechanisms of GC invasion and
metastasis, including the alterations to metastasis-associated
genes, oncogenes, and tumor suppressor genes, may inform potential
treatment avenues for patients with GC (4–6).
Investigating the molecular mechanisms underlying GC invasion and
metastasis, and identifying novel biomarkers involved in GC
invasion, has been a focus for cancer research (7–9), but has
proven difficult thus far.
Matrix metalloproteinases (MMPs) mediate the
degradation of the extracellular matrix to affect tumor cell
adhesion and migration. MMPs are upregulated in numerous types of
cancer, and their expression is closely associated with the
occurrence, invasion and prognosis of cancer (10,11);
previous studies have demonstrated that the increased expression of
MMPs is associated with the enhanced invasiveness of GC cells, and
is associated with poor prognosis in patients with GC (12–14).
MMP-11 has been revealed to have a crucial role in the
proliferation and invasion of GC, and its expression is associated
with the expression of insulin-like growth factor-1 (IGF-1),
indicating the presence of a possible association between MMP-11
and IGF-1 in the development and progression of GC (15,16). IGF-1
may stimulate a range of biological processes, including cellular
proliferation, differentiation, migration and survival, by binding
to the IGF-1 receptor (IGF-1R) (17–19). The
altered expression of IGF-1 has been reported in certain tumor
types, including in liver and breast cancer, and GC (20–22);
however, the specific mechanisms associated with IGF-1
dysregulation have not yet been fully characterized. A recent study
demonstrated that IGF-1R knockdown not only suppressed the growth
of GC cells via G1 cell cycle arrest and apoptosis, but also
inhibited cancer cell invasion (23).
Therefore, in the present study, the expression of IGF-1 and MMP-11
in GC tissues was analyzed, and the specific mechanism underlying
GC proliferation and invasion associated with IGF-1 was
investigated.
Materials and methods
Reagents
Recombinant human IGF-1 (25, 50, 100 ng/ml), NT157
(5, 10 µM) and piceatannol (10, 20 µM) were all obtained from
Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). Different doses of
IGF-1 were employed Antibodies against phospho (p)-STAT3 (sc-8059),
p-JAK1 (sc-16773), p-JAK2 (sc-21870), p-JAK3 (sc-16567), p-IGFR
(sc-81499), β-actin (sc-47778), horseradish peroxidase (HRP)
coupled goat anti-mouse IgG (sc-2031) and HRP coupled rabbit
anti-goat IgG (sc-2768) secondary antibodies were all purchased
from Santa Cruz Biotechnology, Inc.
Clinical specimens
From September 2008 to November 2011, Specimens of
GC tissue (male 7, female 3, 58±13.5, n=10), para-carcinoma tissue
(male 7, female 3, 58±13.5, n=10), normal gastric tissue (male 7,
female 3, 58±13.5, n=10), and gastric ulcer tissue (male 8, female
2, 55.3±15.6, n=10) were collected from the Department of
Gastrointestinal Surgery in Renji Hospital (Shanghai, China). The
present study was approved by the Institutional Research Ethics
Committee of Renji Hospital. Written informed consent was obtained
from all participants for the use of tissue samples. None of the
patients recruited to the present study had received other
anticancer treatments prior to surgery. Sections from each specimen
were independently examined by two pathologists, and histological
typing was performed using Lauren's classification (24). Tumor, node and metastasis
classification of malignant tumors was assigned in accordance to
the International Union Against Cancer (25).
Cells lines and culture
The human GC SGC-7901 cell line was obtained from
the American Type Culture Collection (Manassas, VA, USA). Cells
were maintained in RPMI-1640 medium (Invitrogen; Thermo Fisher
Scientific, Inc., USA) supplemented with 10% fetal bovine serum
(FBS, Sigma-Aldrich; Merck KGaA) and 100 U/ml of
penicillin/streptomycin (Sigma-Aldrich; Merck KGaA) at 37°C in a
humidified atmosphere with 5% CO2. SGC-7901 cells at
logarithmic phase were cultured with different doses of IGF-1,
NT157 or piceatannol for different time (20, 40, and 60 min). The
Stat3 siRNA or Scramble siRNA was transfected into SGC-7901 cells
using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc.). Stat3 siRNA: sense:
5′-CACCGCAACAGATTGCCTGCATTGGTTCTGCAGGCAATCTGTTGCTTTTTTG-3′;
antisense:
5′-GATCCAAAAAAGCAACAGATTGCCTGCATTGGTCTCTTGAACCAATGCAGGCAATCTGTTGC-3′;
Scramble siRNA: sense:
5′-CACCGTTCTCCGAACGTGTCACGTCAAGAGATTACGTGACACGTTCGGAGAATTTTTTG-3′;
antisense:
5′-GATCCAAAAAATTCTCCGAACGTGTCACGTAATCTCTTGACGTGACACGTTCGGAGAAC-3′.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
RT-qPCR was performed as reported previously
(26), with modifications. Briefly,
total RNA was extracted from cells using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) and refined using an
RNeasy Mini kit (Qiagen, Inc., Valencia, CA, USA) according to the
manufacturers' protocols. Samples (1 µg RNA) were
reverse-transcribed using a first-strand cDNA synthesis kit
(Invitrogen; Thermo Fisher Scientific, Inc.). The synthesized cDNA
was used for qPCR with the Chromo 4 instrument and
SsoFast™ EvaGreen Supermix, and then analyzed with
Opticon Monitor Analysis software version 2.0 (all Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Primers used for
amplification were as follows: IGF-1 sense:
5′-CAACAAGCCCACAGGGTATGGC-3′; antisense:
5′-ACAGGTAACTCGTGCAGAGCAAAGC-3′; MMP-1 sense:
5′-ACATCGTGTTGCGGCTCATGA-3′; antisense:
5′-TTTGGGGTTTGTGGGCCGATGG-3′; GAPDH sense:
5′-GTGGACATCCGCAAAGAC-3′; antisense: 5′-AAAGGGTGTAACGCAACTAA-3′.
The PCR cycle included an initial denaturation at 95°C for 10 min
followed by 40 cycles of 95°C for 15 sec, and 60°C for 1 min.
Specificity was determined by electrophoretic analysis of the
reaction products. GAPDH was used as an internal standard. Data
were analyzed using the 2−ΔΔCT method as described
elsewhere (27).
Western blotting
Western blotting was performed as previously
described (28), with modifications.
SGC-7901 cells were harvested in radioimmunoprecipitation assay
lysis buffer (Bioteke Corporation, Beijing, China) and 35 µg
protein per lane was separated by 10% SDS-PAGE mini-gel and
transferred to a polyvinylidene fluoride membrane (EMD Millipore,
Billerica, MA, USA) for 60 min at 100 V. Following incubation in
blocking buffer (Tris-buffered saline containing 5% non-fat dry
milk, 150 mM NaCl, 50 nM Tris, 0.05% Tween-20, pH 7.5) for 1 h at
room temperature, the membrane was hybridized in blocking buffer
with the aforementioned primary antibodies against p-STAT3 (1:200),
p-IGF-1R (1:500), p-JAK1 (1:200), p-JAK2 (1:500), p-JAK3 (1:200),
and β-actin (1:500) overnight at 4°C, then incubated with HRP
coupled goat anti-mouse IgG (1:3,000) or rabbit anti-goat IgG
(1:5,000) secondary antibodies followed by detection with an
enhanced chemiluminescence reagent (GE Healthcare, Chicago, IL,
USA). The band densities were analyzed with ImageJ software
(version 1.47, National Institutes of Health, Bethesda, MD,
USA).
Cell proliferation and invasion
assays
Cell proliferation and invasion analyses were
performed as previously described (29), with alterations. The proliferation
ability of SGC-7901 cells was assessed by an MTT spectrophotometric
dye (Sigma-Aldrich; Merck KGaA) assay. SGC-7901 cells were seeded
in 24-well plates at a density of 8×103 cells per well.
The proliferation rate was measured at 0, 24, 48, 72, 96, 120 and
144 h after seeding. Cells were incubated for 4 h in 20 µl MTT at
37°C, and the supernatant was removed. MTT was dissolved by adding
150 µl/well dimethylsulfoxide. The absorbance was determined at 450
nm using a microplate reader.
For cell invasion analysis, transwell chambers
(24-well; pore size, 8 µm; BD Biosciences, San Jose, CA, USA) were
coated with Matrigel (BD Biosciences) prior to adding the cells
(5×105 cells/ml) and incubated at 37°C for 24 h,
allowing the gel to solidify. RPMI 1640 with 10% FBS (400 µl) was
added to the lower chamber to act as the chemotactic agent.
Invasive cells on the lower side were fixed with cold methanol
(−20°C) for 10 min and then air-dried. Cells were stained with 0.1%
crystal violet (dissolved in methanol) for 30 min at room
temperature, and then counted using a brightfield microscope at a
magnification of ×200.
Statistical analysis
All experiments were performed at least three times.
Statistical analyses were performed using GraphPad Prism 6 software
(GraphPad Software Inc., La Jolla, CA, USA). A one-way analysis of
variance was performed to determine the differences between
multiple groups followed by the Tukey's test. Data were presented
as the mean ± standard error of the mean. P<0.05 was considered
to indicate a statistically significant difference.
Results
Expression of MMP-11 and IGF-1 is
significantly upregulated in GC tissues
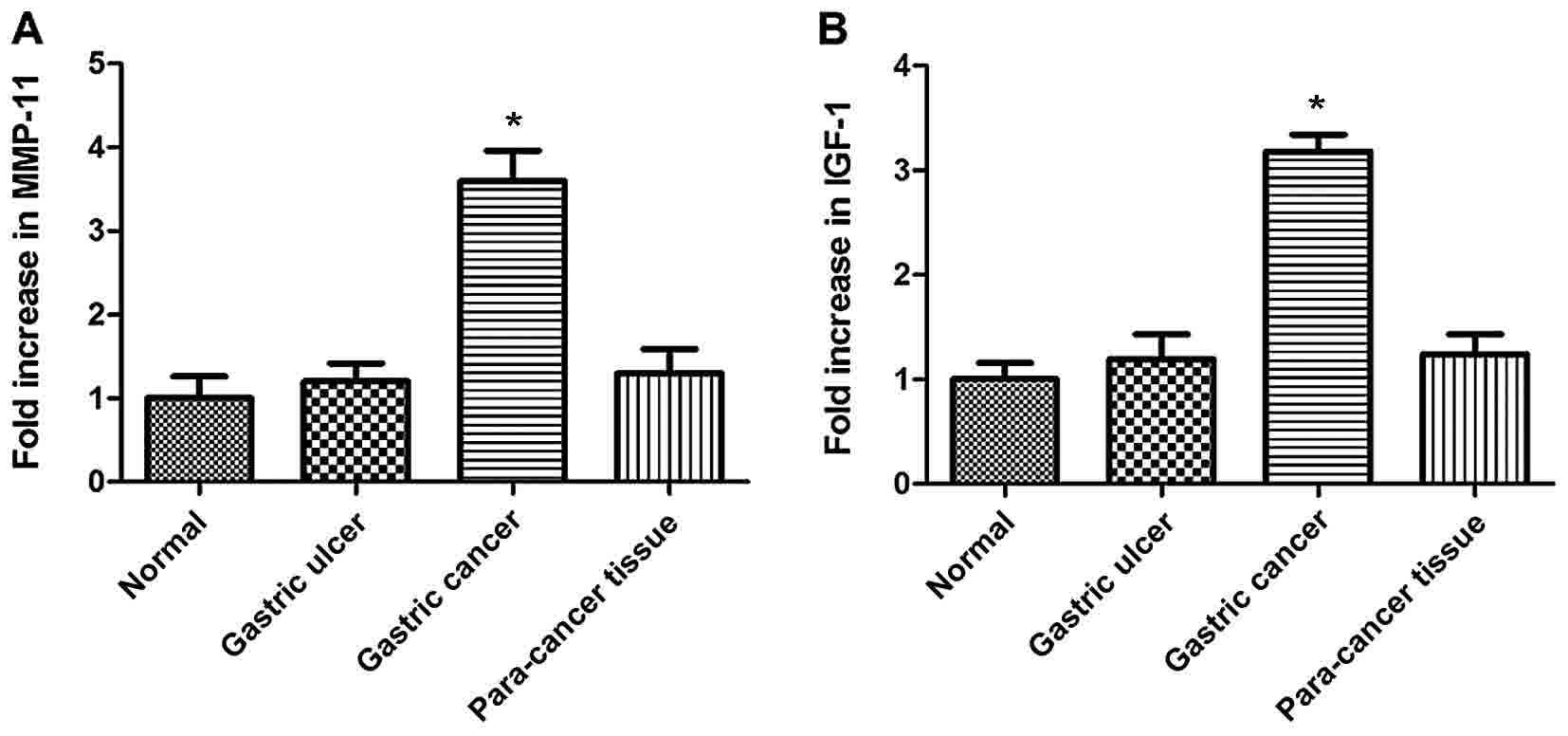
The expression of MMP-11 and IGF-1 was determined in
GC, normal gastric, gastric ulcer and para-carcinoma tissue samples
using RT-qPCR. The expression of MMP-11 did not differ
significantly between normal, gastric ulcer and para-carcinoma
tissue samples, whereas it was significantly higher in GC tissues
(Fig. 1A; P<0.05). Fig. 1B illustrates the differences in IGF-1
expression between the four groups. Similarly, no significant
alterations to IGF-1 expression were observed between normal
gastric, gastric ulcer and para-carcinoma tissue samples, whereas
GC tissues exhibited significantly higher expression of IGF-1
(P<0.05).
IGF-1 elevates the phosphorylation of
STAT3, JAK1, and IGF-1R in SGC-7901 cells
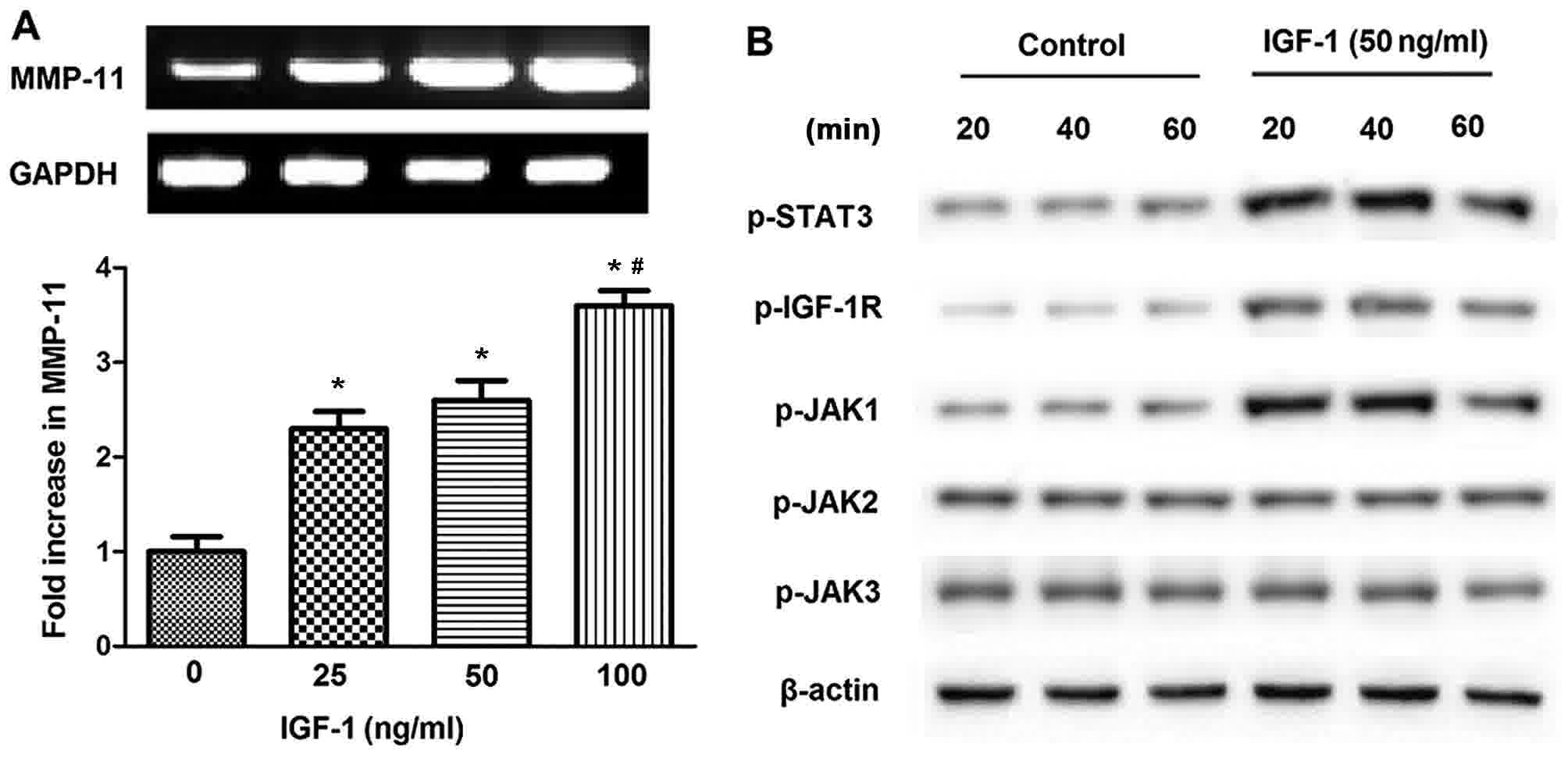
Next, the association between IGF-1 and MMP-11
expression in SGC-7901 cells was examined. IGF-1 treatment
significantly promoted MMP-11 expression in SGC-7901 cells in
parallel with increases in the dose of IGF-1 (P<0.05; Fig. 2A). Given as the JAK/STAT pathway may
be activated in response to a variety of cytokines and growth
factors, including epidermal growth factor, interleukin-6 and
platelet-derived growth factor (30),
western blotting was performed to investigate the time-dependent
action of IGF-1 on the phosphorylation of JAK family kinases and
STAT3 in SGC-7901 cells. The addition of 50 ng/ml IGF-1 to SGC-7901
cells enhanced the phosphorylation of STAT3, JAK1 and IGF-1R at 20,
40 and 60 min, although this was observed to a lesser extent at 60
min (Fig. 2B). By contrast, the
phosphorylation of JAK2 and 3 was not affected following IGF-1
stimulation. These data indicated that STAT3 and JAK1, but not JAK2
and JAK3, were associated with the effect of IGF-1 signaling in
SGC-7901 cells.
NT157 reduces the phosphorylation of
STAT3, JAK1, and IGF-1R in SGC-7901 cells
Considering the role of IGF-1R in cell
proliferation, differentiation, migration and survival following
the binding of IGF-1, the IGF-1R inhibitor NT157 was utilized to
examine the effect of IGF-1R on MMP-11 expression, and the
phosphorylation of STAT3, JAK1 and IGF-1R in SGC-7901 cells. As
demonstrated in Fig. 3A, NT157
treatment caused a significant decline in the elevated expression
of MMP-11 induced by IGF-1 in SGC-7901 cells that was inversely
associated with the concentration of NT157. In addition, the
IGF-1-induced phosphorylation of STAT3, JAK1, and IGF-1R was also
inhibited in SGC-7901 cells following NT157 treatment (5 µM;
Fig. 3B). The results in Fig. 3 indicated that the phosphorylation of
STAT3, JAK1 and IGF-1R following IGF-1 treatment was induced by
IGF-1R activity in SGC-7901 cells.
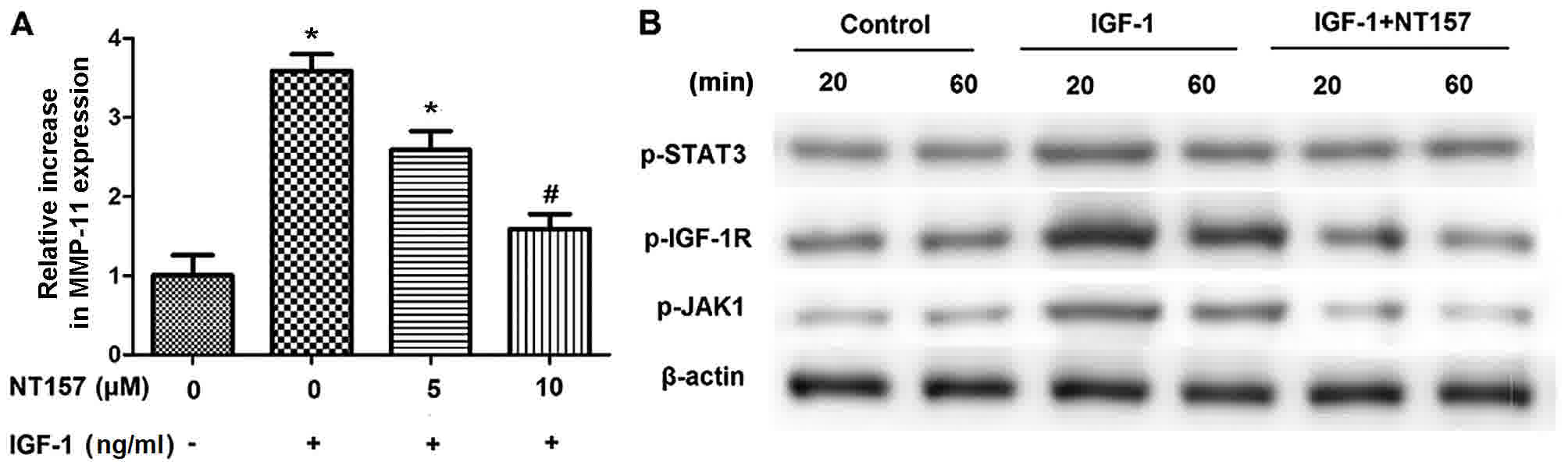 | Figure 3.IGF-1R is implicated in the elevated
expression of MMP-11 as well as the enhanced phosphorylation of
STAT3, JAK1 and IGF-1R induced by IGF-1 in SGC-7901 cells. (A)
NT157, an IGF-1R inhibitor, significantly reduced the extent that
IGF-1 increased the expression of MMP-11 in SGC-7901 cells. Reverse
transcription-quantitative polymerase chain reaction analysis was
performed to detect the change in the expression of MMP-11; GAPDH
was used as the internal control. (B) STAT3, JAK1, and IGF-1R
phosphorylation induced by IGF-1 (50 ng/ml) was also inhibited in
SGC-7901 cells following treatment with 5 µM NT157. Western
blotting was performed to measure the phosphorylation of JAK1,
STAT3, and IGF-1R; β-actin was used as protein loading control.
*P<0.05 vs. no treatment. #P<0.05, vs. IGF-1
alone. IGF-1R, insulin-like growth factor-1 receptor; MMP-11,
matrix metalloproteinase-11; STAT3, signal transducer and activator
of transcription 3; JAK1, Janus kinase 1. |
JAK1/STAT3 pathway is implicated in
the elevated expression of MMP-11 elicited by IGF-1 in SGC-7901
cells
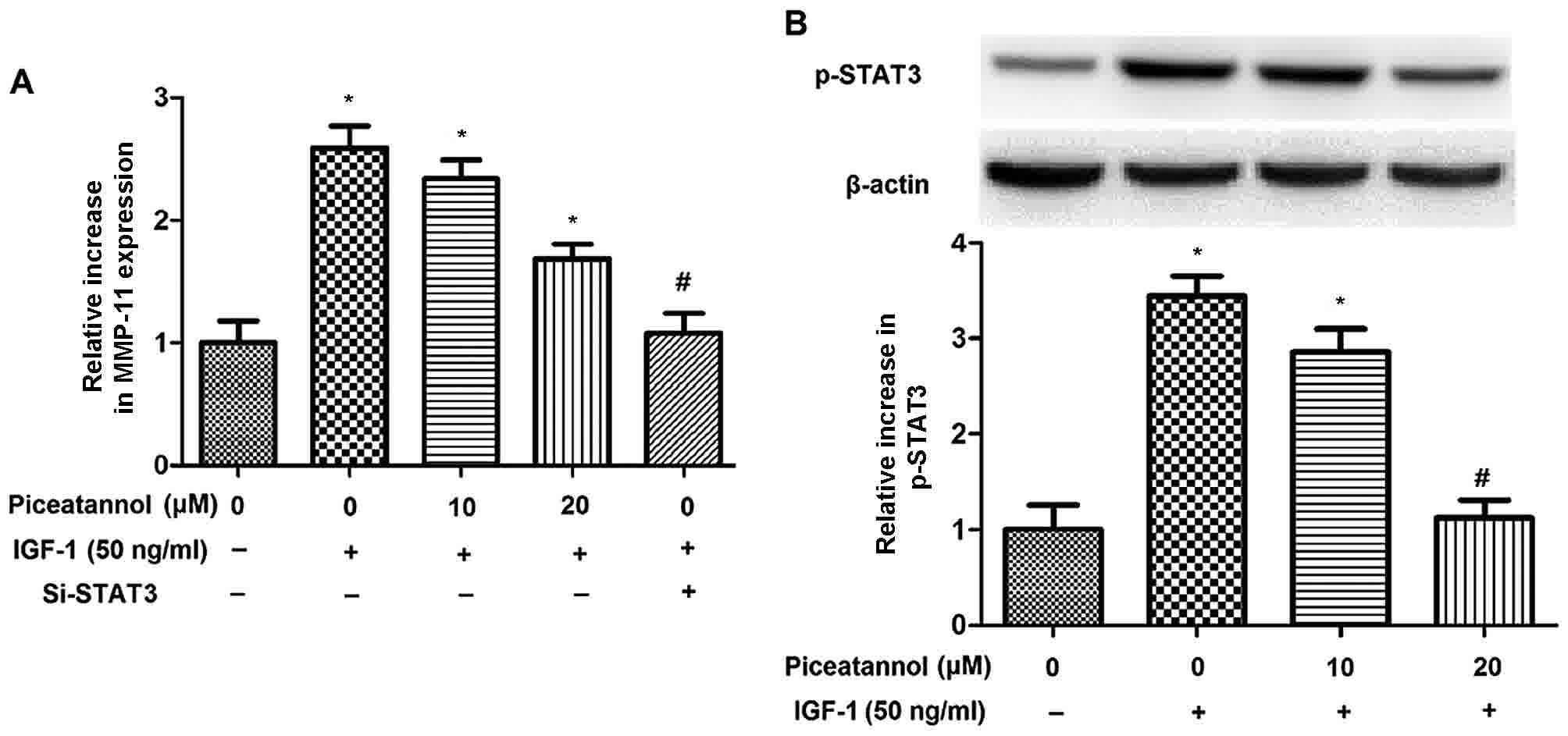
The present study has demonstrated that IGF-1
regulated MMP-11 expression and the phosphorylation of STAT3 and
JAK1 in SGC-7901 cells; however, there is limited data regarding
the association between IGF-1-induced MMP-11, STAT3 and JAK1
activation. The present study therefore examined the effects of
STAT3 and JAK1 on the IGF-1-induced activation of MMP-11 in
SGC-7901 cells using piceatannol (a JAK1 inhibitor) and small
interfering RNA targeted at STAT3 (si-STAT3). As indicated by
Fig. 4A, piceatannol treatment
significantly decreased the elevated expression of MMP-11 induced
by IGF-1 in SGC-7901 cells in a dose-dependent manner; the maximal
decline in MMP-11 expression was observed following STAT3-knockdown
with si-STAT3. Furthermore, the enhanced phosphorylation of STAT3
induced by IGF-1 was reduced in SGC-7901 cells proportional to the
concentration of piceatannol (Fig.
4B). These results demonstrate the association of the
JAK1/STAT3 pathway with the increased expression of MMP-11 induced
by IGF-1 in SGC-7901 cells.
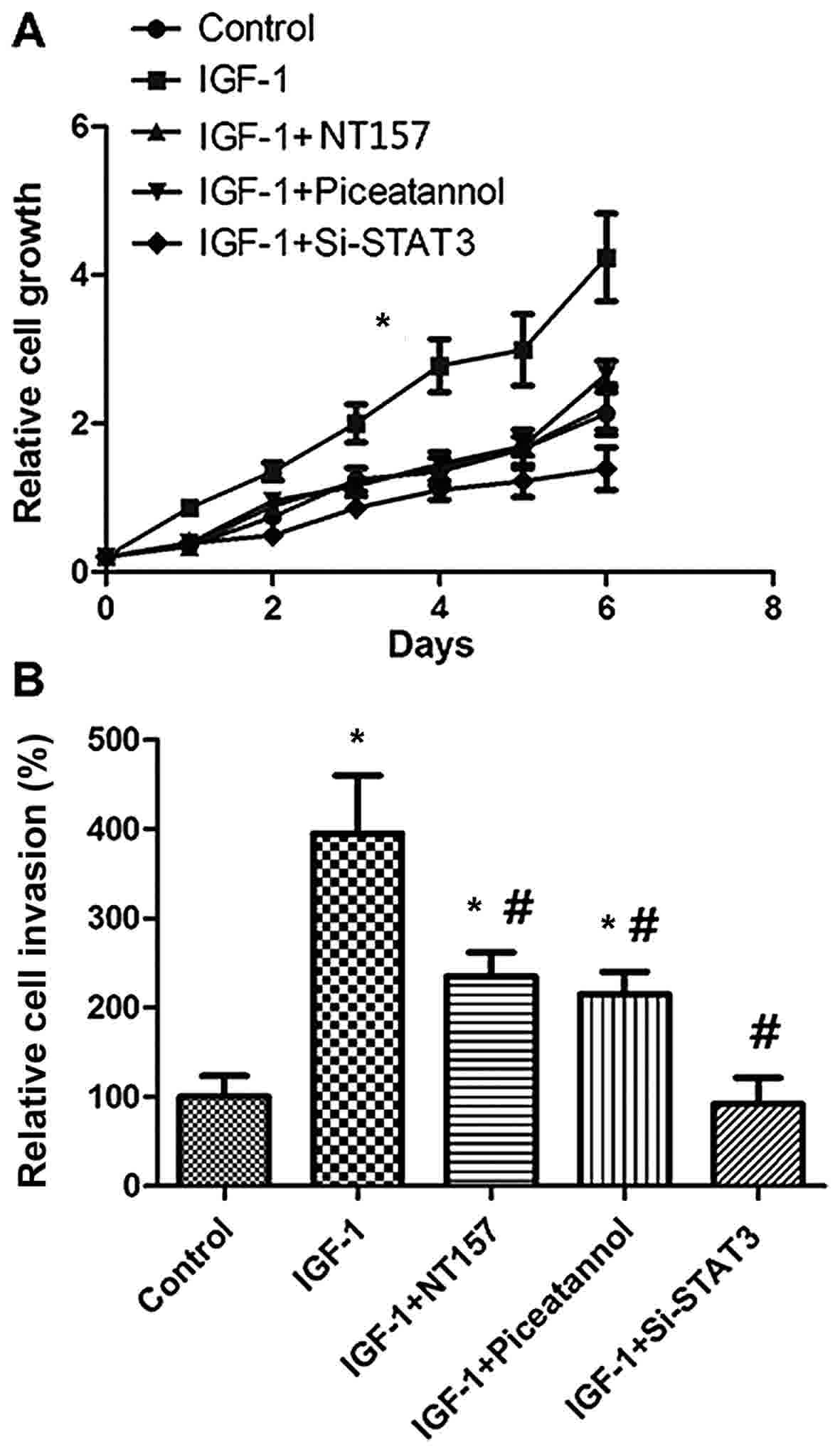
IGF-1 promotes the proliferation and
invasion of SGC-7901 cells via the JAK1/STAT3 pathway
The data of the present study have indicated the
association of the JAK1/STAT3 pathway with the IGF-1-induced
expression of MMP-11 in SGC-7901 cells; the role of MMP-11 in GC
proliferation and invasion has been previously established
(16). Thus, the present study next
evaluated the proliferation and invasion of SGC-7901 cells
following stimulation with IGF-1. IGF-1 treatment induced the
proliferation and invasion of SGC-7901 cells; this effect was less
pronounced following treatment with NT157, piceatannol, or si-STAT3
(Fig. 5A and B). The data of Fig. 5 indicate that the IGF-1-induced
expression of MMP-11 may promote the proliferation and invasion of
SGC-7901 cells through the JAK1/STAT3 pathway.
Discussion
MMP-11 and IGF-1 have been demonstrated to be
associated with the proliferation and invasion of GC (15,16);
however, the specifics of the interaction between these signaling
pathways remain uncharacterized. Although the JAK/STAT pathway
serves a role in the progression of GC, the mechanism by which this
pathway is associated with the IGF-1-induced proliferation and
invasion of GC has not been identified. The present study
demonstrated that the expression of IGF-1 and MMP-11 was
significantly upregulated in GC tissue relative to normal gastric
tissue. IGF-1 induced the expression of MMP-11, as well as the
phosphorylation of STAT3, IGF-1R and JAK1 in SGC-7901 cells.
Treatment with an IGF-1R inhibitor reversed the enhanced
phosphorylation of STAT3, JAK1 and IGF-1R induced by IGF-1 in
SGC-7901 cells. In addition, the JAK1/STAT3 pathway was associated
with the increased expression of MMP-11 induced by IGF-1 in
SGC-7901 cells. Finally, the present study demonstrated that
IGF-1-induced MMP-11 expression may have facilitated the
proliferation and invasion of SGC-7901 cells via JAK1/STAT3
pathway.
Zhao et al (15) reported that the increased expression
of MMP-11 was associated with an elevation in IGF-1 expression in
GC tissues. Kou et al (16)
reported that MMP-11 knockdown repressed the proliferative and
invasive activities of SGC-7901 cells, with a corresponding
decrease in the expression of IGF-1, PCNA and VEGF. The present
study reported the increased expression of IGF-1 and MMP-11 in GC
tissue compared with non-cancerous tissue, and that IGF-1 treatment
induced MMP-11 expression in SGC-7901 cells. The data of the
present study concerning IGF-1 and MMP-11 expression in GC tissues
are consistent with a previous study (15).
The JAK/STAT pathway regulates cell development and
survival. Giorgetti-Peraldi et al (31) identified that insulin promoted the
tyrosine phosphorylation of JAK1 in fibroblasts overexpressing the
insulin receptor, but did not alter the tyrosine phosphorylation
status of JAK2. However, Saad et al (32) demonstrated that insulin stimulated the
tyrosine phosphorylation of JAK2 in the insulin-sensitive tissues
of rats. Subsequently, Gual et al (33) reported that in mouse fibroblast NIH
3T3 cells overexpressing insulin and IGF-1 receptors, treatment
with insulin and IGF-1 resulted in the phosphorylation and
activation of JAK1 and JAK2, with JAK1 interacting directly with
phosphorylated insulin and IGF-1 receptors. Previous studies have
also demonstrated that insulin stimulates the phosphorylation and
activation of STAT1, 3 and 5 (34,35). The
importance of the JAK/STAT3 pathway in the survival of neurons in
response to IGF-1 treatment has also been noted (36). The present study additionally
demonstrated that IGF-1 treatment stimulated the phosphorylation of
JAK1, STAT3 and IGF-1R in SGC-7901 cells, but not JAK2 or JAK3. An
explanation for the inconsistencies between the present study and
Gual et al (33) may be the
different cell types employed. We hypothesize that JAK1 and STAT3
are associated with IGF-1-regulated signaling in SGC-7901
cells.
The present study demonstrated that treatment with
the IGF-1R inhibitor NT157 reversed the elevated expression of
MMP-11 and the phosphorylation of STAT3, JAK1 and IGF-1R as induced
by IGF-1 in SGC-7901 cells. NT157 was previously demonstrated to
affect IGF-1R and STAT3 in the inhibition of colorectal cancer
development (37). Zong et al
(38) demonstrated that STAT3, and
not STAT5, was activated in response to IGF-1 in 293T cells
overexpressing IGF-1R, and that the IGF-1 stimulation of endogenous
IGF-1R promoted the tyrosine phosphorylation of STAT3, JAK1 and 2.
The specific mediation of STAT3 activation by insulin and IGF-1
receptors has previously been demonstrated (39). A recent study identified that the
alterations in cellular behavior induced by IGF-1R and p38
mitogen-activated protein kinase inhibitors were accompanied by
alterations to the level of STAT3 in human dental pulp stem cell
quiescence, proliferation and differentiation, indicating that
STAT3 may be a target for IGF-1R (40).
Quelle et al (41) demonstrated that STATs were direct
targets for activated JAKs. Thus, the present study raised the
question of the physiological role of JAK1 activation by IGF-1.
Shimoda et al (42) reported
that the role of JAK1 in STAT activation and receptor
phosphorylation could be induced by granulocyte colony-stimulating
factor, whereas Sawka-Verhelle et al (43) observed that insulin and IGF-1 caused
STAT5B phosphorylation, and that JAKs were not associated with
insulin-induced STAT5B activation in Cercopithecus aethiops
Cos-7 kidney cells, whereas JAK2 was essential for the activation
of STAT5B by growth hormones. The present study revealed that JAK1
and STAT3 were involved in regulating MMP-11 via IGF-1 in SGC-7901
cells. Previous studies have indicated that cell proliferation,
invasion and metastasis in certain types of tumor require the
involvement of the JAK/STAT pathway (44,45). The
JAK1/STAT3 pathway was implicated in the proliferation and invasion
of SGC-7901 cells, and it can be inferred that the IGF-1-induced
expression of MMP-11 promoted the proliferation and invasion of
SGC-7901 cells through the JAK1/STAT3 pathway.
In summary, the results of the present study
demonstrated that IGF-1 and MMP-11 expression is significantly
upregulated in GC tissues, and that IGF-1 stimulates MMP-11
expression and the phosphorylation of STAT3, IGF-1R and JAK1 in
SGC-7901 cells. Furthermore, the elevated expression of MMP-11, and
the enhanced phosphorylation of STAT3, JAK1 and IGF-1R induced by
IGF-1, are associated with IGF-1R in SGC-7901 cells. Based on these
data, it can be concluded that IGF-1-induced MMP-11 expression
induces the proliferation and invasion of SGC-7901 cells via the
JAK1/STAT3 pathway.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author, on reasonable
request.
Authors' Contributions
CS designed the project and reviewed the manuscript.
WW conducted the experiments and drafted the manuscript. CW
analyzed the data and drafted the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Research Ethics Committee of Renji Hospital. Written informed
consent was obtained from all participants for the use of tissue
samples.
Consent for publication
Study participants provided approval for
publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferro A, Peleteiro B, Malvezzi M, Bosetti
C, Bertuccio P, Levi F, Negri E, La Vecchia C and Lunet N:
Worldwide trends in gastric cancer mortality (1980-2011), with
predictions to 2015 and incidence by subtype. Eur J Cancer.
50:1330–1344. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Bray F, Pisani P and Parkin DM:
Globocan 2000: Cancer Incidence, Mortality and Prevalence
Worldwide. IARC Press; Lyon: 2001
|
|
3
|
Velho S, Fernandes MS, Leite M, Figueiredo
C and Seruca R: Causes and consequences of microsatellite
instability in gastric carcinogenesis. World J Gastroenterol.
20:16433–16442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oue N, Aung PP, Mitani Y, Kuniyasu H,
Nakayama H and Yasui W: Genes involved in invasion and metastasis
of gastric cancer identified by array-based hybridization and
serial analysis of gene expression. Oncology. 69 Suppl 1:S17–S22.
2005. View Article : Google Scholar
|
|
5
|
Li X, Zhang Y, Zhang H, Liu X, Gong T, Li
M, Sun L, Ji G, Shi Y, Han Z, et al: miRNA-223 promotes gastric
cancer invasion and metastasis by targeting tumor suppressor
EPB41L3. Mol Cancer Res. 9:824–833. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ganesan K, Ivanova T, Wu Y, Rajasegaran V,
Wu J, Lee MH, Yu K, Rha SY, Chung HC, Ylstra B, et al: Inhibition
of gastric cancer invasion and metastasis by PLA2G2A, a novel
beta-catenin/TCF target gene. Cancer Res. 68:4277–4286. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dun B, Sharma A, Teng Y, Liu H, Purohit S,
Xu H, Zeng L and She JX: Mycophenolic acid inhibits migration and
invasion of gastric cancer cells via multiple molecular pathways.
PLoS One. 8:e817022013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang L, Guo J, Wang Q, Zhou J, Xu C, Teng
R, Chen Y, Wei Q and Liu ZP: LZTFL1 suppresses gastric cancer cell
migration and invasion through regulating nuclear translocation of
β-catenin. J Cancer Res Clin Oncol. 140:1997–2008. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang ZZ, Shen ZY, Shen YY, Zhao EH, Wang
M, Wang CJ, Cao H and Xu J: HOTAIR long noncoding RNA promotes
gastric cancer metastasis through suppression of poly r(C)-binding
protein (PCBP) 1. Mol Cancer Ther. 14:1162–1170. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Deryugina EI and Quigley JP: Matrix
metalloproteinases and tumor metastasis. Cancer Metastasis Rev.
25:9–34. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu D, Nakano J, Ishikawa S, Yokomise H,
Ueno M, Kadota K, Urushihara M and Huang CL: Overexpression of
matrix metalloproteinase-7 (MMP-7) correlates with tumor
proliferation, and a poor prognosis in non-small cell lung cancer.
Lung Cancer. 58:384–391. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kubben FJ, Sier CF, van Duijn W, Griffioen
G, Hanemaaijer R, van de Velde CJ, van Krieken JH, Lamers CB and
Verspaget HW: Matrix metalloproteinase-2 is a consistent prognostic
factor in gastric cancer. Br J Cancer. 94:1035–1040. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shim KN, Jung SA, Joo YH and Yoo K:
Clinical significance of tissue levels of matrix metalloproteinases
and tissue inhibitors of metalloproteinases in gastric cancer. J
Gastroenterol. 42:120–128. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Łukaszewicz-Zając M, Mroczko B and
Szmitkowski M: Gastric cancer-The role of matrix metalloproteinases
in tumor progression. Clin Chim Acta. 412:1725–1730. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao ZS, Chu YQ, Ye ZY, Wang YY and Tao
HQ: Overexpression of matrix metalloproteinase 11 in human gastric
carcinoma and its clinicopathologic significance. Hum Pathol.
41:686–696. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kou YB, Zhang SY, Zhao BL, Ding R, Liu H
and Li S: Knockdown of MMP11 inhibits proliferation and invasion of
gastric cancer cells. Int J Immunopathol Pharmacol. 26:361–370.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stewart CE and Rotwein P: Growth,
differentiation, and survival: Multiple physiological functions for
insulin-like growth factors. Physiol Rev. 76:1005–1026. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
LeRoith D, Werner H, Beitner-Johnson D and
Roberts CT Jr: Molecular and cellular aspects of the insulin-like
growth factor I receptor. Endocr Rev. 16:143–163. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Delafontaine P, Song YH and Li Y:
Expression, regulation, and function of IGF-1, IGF-1R, and IGF-1
binding proteins in blood vessels. Arterioscler Thromb Vasc Biol.
24:435–444. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Moerkens M, Ramaiahgari S, de
Bont H, Price L, Meerman J and van de Water B: Elevated
insulin-like growth factor 1 receptor signaling induces
antiestrogen resistance through the MAPK/ERK and PI3K/Akt signaling
routes. Breast Cancer Res. 13:R522011. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kang HJ, Yi YW, Kim HJ, Hong YB, Seong YS
and Bae I: BRCA1 negatively regulates IGF-1 expression through an
estrogen-responsive element-like site. Cell Death Dis. 3:e3362012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li S, Lei X, Zhang J, Yang H, Liu J and Xu
C: Insulin-like growth factor 1 promotes growth of gastric cancer
by inhibiting foxo1 nuclear retention. Tumour Biol. 36:4519–4523.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ge J and Chen Z, Huang J, Yuan W, Den Z
and Chen Z: Silencing insulin-like growth factor-1 receptor
expression inhibits gastric cancer cell proliferation and invasion.
Mol Med Rep. 11:633–638. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Seoane A, Bessa X, Balleste B, O'Callaghan
E, Panadès A, Alameda F, Navarro S, Gallén M, Andreu M and Bory F:
Helicobacter pylori and gastric cancer: Relationship with
histological subtype and tumor location. Gastroenterol Hepatol.
28:60–64. 2005.(In Spanish). View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ajani J, D'Amico TA, Hayman JA, Meropol NJ
and Minsky B: National Comprehensive Cancer Network: Gastric
cancer. Clinical practice guidelines in oncology. J Natl Compr Canc
Netw. 1:28–39. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Matsubara J, Yamada Y, Nakajima TE, Kato
K, Hamaguchi T, Shirao K, Shimada Y and Shimoda T: Clinical
significance of insulin-like growth factor type 1 receptor and
epidermal growth factor receptor in patients with advanced gastric
cancer. Oncology. 74:76–83. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim JE, Lee MH, Nam DH, Song HK, Kang YS,
Lee JE, Kim HW, Cha JJ, Hyun YY, Han SY, et al: Celastrol, an NF-κB
inhibitor, improves insulin resistance and attenuates renal injury
in db/db mice. PLoS One. 8:e620682013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang BG, Li JF, Yu BQ, Zhu ZG, Liu BY and
Yan M: microRNA-21 promotes tumor proliferation and invasion in
gastric cancer by targeting PTEN. Oncol Rep. 27:1019–1026. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Vignais ML, Sadowski HB, Watling D, Rogers
NC and Gilman M: Platelet-derived growth factor induces
phosphorylation of multiple JAK family kinases and STAT proteins.
Mol Cell Biol. 16:1759–1769. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Giorgetti-Peraldi S, Peyrade F, Baron V
and Van Obberghen E: Involvement of Janus kinases in the insulin
signaling pathway. Eur J Biochem. 234:656–660. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Saad MJ, Carvalho CR, Thirone AC and
Velloso LA: Insulin induces tyrosine phosphorylation of JAK2 in
insulin-sensitive tissues of the intact rat. J Biol Chem.
271:22100–22104. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gual P, Baron V, Lequoy V and Van
Obberghen E: Interaction of Janus kinases JAK-1 and JAK-2 with the
insulin receptor and the insulin-like growth factor-1 receptor.
Endocrinology. 139:884–893. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen J, Sadowski HB, Kohanski RA and Wang
LH: Stat5 is a physiological substrate of the insulin receptor.
Proc Natl Acad Sci USA. 94:2295–2300. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sawka-Verhelle D, Filloux C,
Tartare-Deckert S, Mothe I and Van Obberghen E: Identification of
Stat 5B as a substrate of the insulin receptor. Eur J Biochem.
250:411–417. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yadav A, Kalita A, Dhillon S and Banerjee
K: JAK/STAT3 pathway is involved in survival of neurons in response
to insulin-like growth factor and negatively regulated by
suppressor of cytokine signaling-3. J Biol Chem. 280:31830–31840.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sanchez-Lopez E, Flashner-Abramson E,
Shalapour S, Zhong Z, Taniguchi K, Levitzki A and Karin M:
Targeting colorectal cancer via its microenvironment by inhibiting
IGF-1 receptor-insulin receptor substrate and STAT3 signaling.
Oncogene. 35:2634–2644. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zong CS, Chan J, Levy DE, Horvath C,
Sadowski HB and Wang LH: Mechanism of STAT3 activation by
insulin-like growth factor I receptor. J Biol Chem.
275:15099–15105. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang W, Zong CS, Hermanto U,
Lopez-Bergami P, Ronai Z and Wang LH: RACK1 recruits STAT3
specifically to insulin and insulin-like growth factor 1 receptors
for activation, which is important for regulating
anchorage-independent growth. Mol Cell Biol. 26:413–424. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Vandomme J, Touil Y, Ostyn P, Olejnik C,
Flamenco P, El Machhour R, Segard P, Masselot B, Bailliez Y,
Formstecher P and Polakowska R: Insulin-like growth factor 1
receptor and p38 mitogen-activated protein kinase signals inversely
regulate signal transducer and activator of transcription 3
activity to control human dental pulp stem cell quiescence,
propagation, and differentiation. Stem Cells Dev. 23:839–851. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Quelle FW, Thierfelder W, Witthuhn BA,
Tang B, Cohen S and Ihle JN: Phosphorylation and activation of the
DNA binding activity of enriched Stat1 by the Janus
protein-tyrosine kinases and the epidermal growth factor receptor.
J Biol Chem. 270:20775–20780. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shimoda K, Feng J, Murakami H, Nagata S,
Watling D, Rogers NC, Stark GR, Kerr IM and Ihle JN: Jak1 plays an
essential role for receptor phosphorylation and Stat activation in
response to granulocyte colony-stimulating factor. Blood.
90:597–604. 1997.PubMed/NCBI
|
|
43
|
Sawka-Verhelle D, Tartare-Deckert S,
Decaux JF, Girard J and Van Obberghen E: Stat 5B, activated by
insulin in a Jak-independent fashion, plays a role in glucokinase
gene transcription. Endocrinology. 141:1977–1988. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Macha MA, Satyanarayana R, Suprit G, Pai
P, Ponnusamy MP, Batra SK and Jain M: Guggulsterone decreases
proliferation and metastatic behavior of pancreatic cancer cells by
modulating JAK/STAT and Src/FAK signaling. Cancer Lett.
341:166–177. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kowshik J, Baba AB, Giri H, Reddy Deepak
G, Dixit M and Nagini S: Astaxanthin inhibits JAK/STAT-3 signaling
to abrogate cell proliferation, invasion and angiogenesis in a
hamster model of oral cancer. PLoS One. 9:e1091142014. View Article : Google Scholar : PubMed/NCBI
|