Introduction
Acute lymphoblastic leukemia (ALL) is an aggressive
type of blood cancer affecting children and adults, with peak
prevalence between 2 and 5 years of age (1). T-cell ALL (T-ALL) is an aggressive
hematological cancer that is caused by the malignant transformation
of thymocyte progenitors (2). T-ALL
accounts for 10–15% of pediatric and 25% of adult ALL cases
(2,3).
The age of the patient at diagnosis, leukocyte count, ethnicity,
gender and immunophenotype are clinical prognostic parameters that
classify ALL patients into different risk groups (4). TALL is classified into a high-risk group
in ALL (4). The prognosis of T-ALL
has improved with the development of high-dose multi-agent
chemotherapy, with a cure rate of ~85% in children and ~50% in
adults (5). However, the treatment is
often accompanied by severe acute toxicities and side effects, such
as primary resistance, early relapse and secondary tumors (2,3). The
identification of new agents for T-ALL patients is urgently
required.
Nightshade, a Chinese herbal medicine, has been used
to treat sores, injuries, swelling and fractures (6). Solanine, the main extract of Nightshade,
is a trisaccharide glycoalkaloid (7).
Solanine has been demonstrated to inhibit the production of
cytokine and nitric oxide in stimulated Jurkat cells LPS-stimulated
Raw macrophages (8).
High concentrations of Solanine result in
cytotoxicity-inducing damage of the plasma membrane, which causes
disorder of metabolism, including reduced NAD(P)H productivity and
the loss or inactivation of NAD(P)H:menadione reductase (9). Solanine was demonstrated to have a
proliferation-inhibiting and apoptosis-promoting effect on multiple
cancer cells, including prostate cancer, pancreatic carcinoma and
melanoma cancer cells (6–9). Studies have also shown that Solanine
suppresses proliferation and metastasis, and promotes apoptosis, in
pancreatic cancer cells (9,10). Solanine induces apoptosis of HepG2
cells by facilitating the opening of the PT channels in the
mitochondria and suppressing the expression of Bcl-2 (11,12).
Additionally, Solanine has been reported to inhibit human melanoma
cells and human prostate cancer cell invasion at non-toxic doses
(6,7).
However, to the best of our knowledge, the efficacy and the
associated molecular mechanisms of Solanine promoting apoptosis in
Jurkat cells have not been explored. In the present study, the
effects of Solanine on the inhibition of proliferation and
induction of apoptosis in Jurkat cells and the underlying molecular
mechanism were investigated. Additionally, the effect of Solanine
on the chemosensitivity of Jurkat cells to Adriamycin was assessed.
The findings indicated the potential of Solanine to improve the
therapeutic outcome of T-ALL.
Materials and methods
Chemicals and reagents
Adriamycin (Melone Pharmaceutical Co. Ltd., Dalian,
China) was dissolved to a concentration of 2 g/l in dH2O
and divided into 25 aliquots (1.5 ml). Solanine (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) was dissolved in dimethyl sulfoxide
(HyClone; GE Healthcare Life Sciences, Logan, UT, USA) to generate
a stock solution (100 µg/ml), and diluted to each designated
concentration in RPMI-1640 (HyClone; GE Healthcare Life Sciences).
The rabbit polyclonal antibodies against B-cell lymphoma-2 (Bcl-2)
and Bcl-2-associated X protein (Bax) were obtained from Beijing
ZhongShan Golden Bridge Technology Co., Ltd. (Beijing, China). The
rabbit polyclonal antibody against GADPH was obtained from Good
here Biotechnology Co., Ltd., Hangzhou, China.
Cell lines and cell culture
The human T-ALL Jurkat cells were obtained from Key
Laboratory of Tumour Molecular Biology of Binzhou Medical
University (Binzhou, China). The cells were cultured in RPMI-1640
medium supplemented with 10% fetal bovine serum (HyClone; GE
Healthcare Life Sciences) at 37°C in a humidified atmosphere
containing 5% CO2.
Cell proliferation assay
Cell Counting Kit 8 (CCK-8; Dojindo Molecular
Technologies, Inc., Shanghai, China) was used to determine the cell
viability in the presence of Solanine (0, 2, 4, 8 or 16 µg/ml),
with/without 0.15 µg/l Adriamycin (4), incubated for 24 h at 37at
24atedci2 in air. Briefly, cells were seeded into
96-well plates at a density of 1×104 cells/well, and,
subsequent to treatment with Solamine (0, 2 4, 6, 8 or 16 µg/ml)
with/without 0.15 µg/l Adriamycin, 10 µl CCK-8 solution was added
to each well and incubated for 4 h at 37°C in a humidified
incubator with 5% CO2 in air. Cells in control group
were supplemented with the equivalent quantity of DMSO The
absorbance was then measured at a wavelength of 490 nm using a
fluorescence spectrofluorometer (F-7000; Hitachi Ltd., Tokyo,
Japan). A blank well containing only medium and drugs was used as a
control.
Flow cytometry analysis
The Jurkat cells (3×105 cells/ml) were
seeded into 6 well plates and incubated with various concentrations
of Solanine (0, 4 and 16 µg/ml) for 24 h at 37°C in a humidified
atmosphere containing 5% CO2 in air. Cells were
collected and washed twice with PBS. Cells were then suspended in
binding buffer (KeyGen Biotech Co., Ltd., Nanjing, China) and
double-stained with annexin V fluorescein isothiocyanate
(FITC)/propidium iodide (PI; KeyGen Biotech Co., Ltd.) for 15 min
in the dark at room temperature. The cell-associated mean
fluorescence intensity (MFI) was detected by flow cytometer using a
FACSCalibur (Beckman Coulter, Brea, CA, USA) to analyzed the
apoptotic cells.
Determination of gene expression by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR)
Total RNA was isolated from Solanine-treated cells
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), according to the manufacturer's protocol.
Reverse transcription was performed to generate first strand cDNA
(Takara Biotechnology Co., Ltd., Dalian, China) using 2 µg of total
RNA. The reverse transcription reaction was implemented with
PrimeScript™ RT reagent kit with gDNA Eraser (Takara Bio, Shiga,
Japan). The primers (Table I) used in
this experiment were designed using Primer 5 version 5.6.0 software
(PREMIER Biosoft Co., Ltd., CA, USA) and synthesized by Sangon
Biotech Co., Ltd., Shanghai, China.
 | Table I.Primers used in reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers used in reverse
transcription-quantitative polymerase chain reaction.
| Gene | Primer sequence,
5′-3′ | Product length,
bp |
|---|
| Bax |
| 117 |
|
Forward |
CCGAGAGGTCTTTTTCCGAG |
|
|
Reverse |
GTGCACAGGGCCTTGAGC |
|
| Bcl-2 |
| 152 |
|
Forward |
GGATTGTGGCCTTCTTTGAG |
|
|
Reverse |
TACCCAGCCTCCGTTATCCT |
|
| GAPDH |
| 121 |
|
Forward |
TGACTTCAACAGCGACACCCA |
|
|
Reverse |
CACCCTGTTGCTGTAGCCAAA |
|
qPCR was performed on an ABI PRISM 7500 qPCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.) by using
SYBRGreen reaction kit (Takara Bio, Inc., Otsu, Japan). The PCR
reaction system consisted of SYBR Green reagent, forward and
reverse primers, template cDNA and nuclease-free distilled water.
The PCR conditions were 95°C for 30 sec, followed by 45 cycles of
95°C for 5 sec and 60°C for 30 sec. GADPH was used as an internal
control. qPCR for each gene of each cDNA sample was assayed in
triplicate. The results were calculated using the 2−ΔΔCq
method (4). The following equations
were used: ΔCq=Cq (target gene)-Cq (GADPH); ΔΔCq=ΔCq
(Solanine-treated cells)-ΔCq (untreated control).
Western blot analysis
The Jurkat cells were incubated with Solanine (0, 4
or 16 µg/ml) for 24 h at 37°C. The cells were then harvested. Lysis
buffer (100 µl; Beyotime Institute of Biotechnology, Shanghai,
China) was added and the protein concentration of the lysate was
determined using a bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology). The lysed samples containing 50 µg
total protein were separated on 10–12% SDS-PAGE (Beyotime Institute
of Biotechnology), with a constant voltage of 80 V for 0.5 h, and
then 100 V for another 1.5 h. The resolved proteins were
electrophoretically transferred to polyvinylidene difluoride
membranes (Merck KGaA) and blocked with 5% skimmed milk for 2 h at
room temperature. Subsequently, the membranes were incubated
overnight at 4°C with specific antibodies. The primary antibodies
used were rabbit polyclonal antibodies against Bcl-2 (dilution,
1:500), Bax (dilution, 1:500) and GAPDH (dilution, 1:1,000). The
following day, the membranes were incubated in horseradish
peroxidase-labeled goat anti-rabbit immunoglobulin G (dilution,
1:5,000; catalog no., ZB-5301; Beijing ZhongShan Golden Bridge
Technology Co., Ltd.) for 2 h at room temperature. Finally, images
were captured using a FluorChem FC2 gel imaging system
(ProteinSimple; Bio-Techne, Minneapolis, MN, USA). The intensity of
each band was normalized by GADPH for their respective lanes.
Data analysis
Statistical analyses were performed using SPSS 13.0
software (SPSS, Inc., Chicago, IL, USA). Data are expressed as the
mean ± standard deviation. Differences between ≥3 groups were
evaluated by one-way analysis of variance followed by
Student-Newman-Keuls test. Independent two sample t-tests were used
to compare the differences between 2 groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Solanine decreased the viability of
Jurkat cells
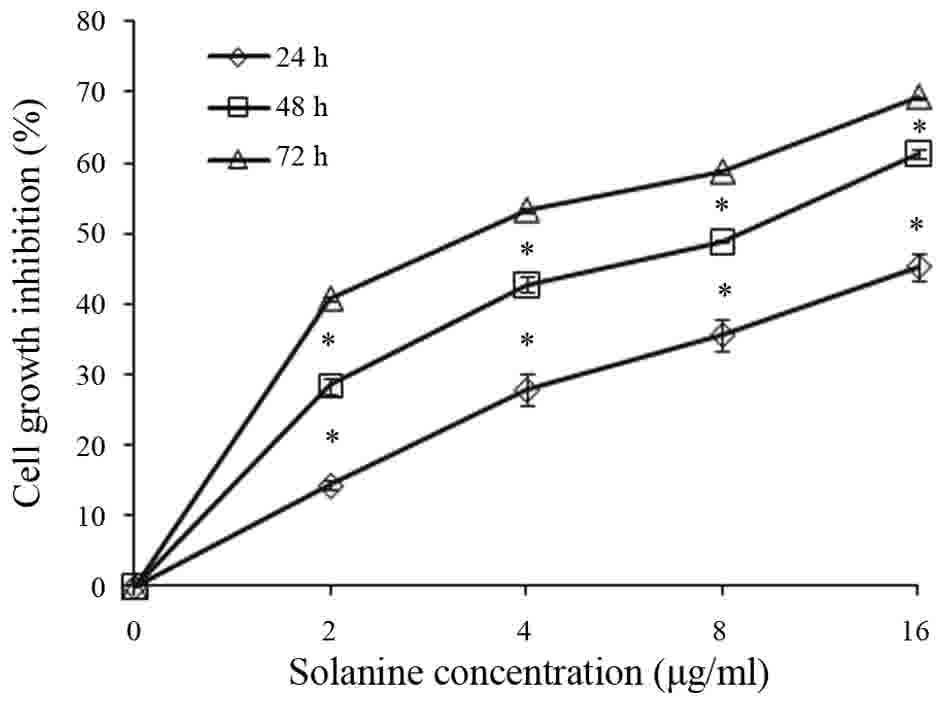
The ability of Solanine to inhibit Jurkat cell
proliferation was assessed. Jurkat cells were treated with various
concentrations of Solanine (0, 2, 4, 8 or 16 µg/ml) for 24, 48 and
72 h. Cell viability was determined by CCK-8 assay. The results
demonstrated that Jurkat cells were sensitive to Solanine, and
Solanine inhibited Jurkat cell proliferation in a dose- and
time-dependent manner (Fig. 1;
P<0.05).
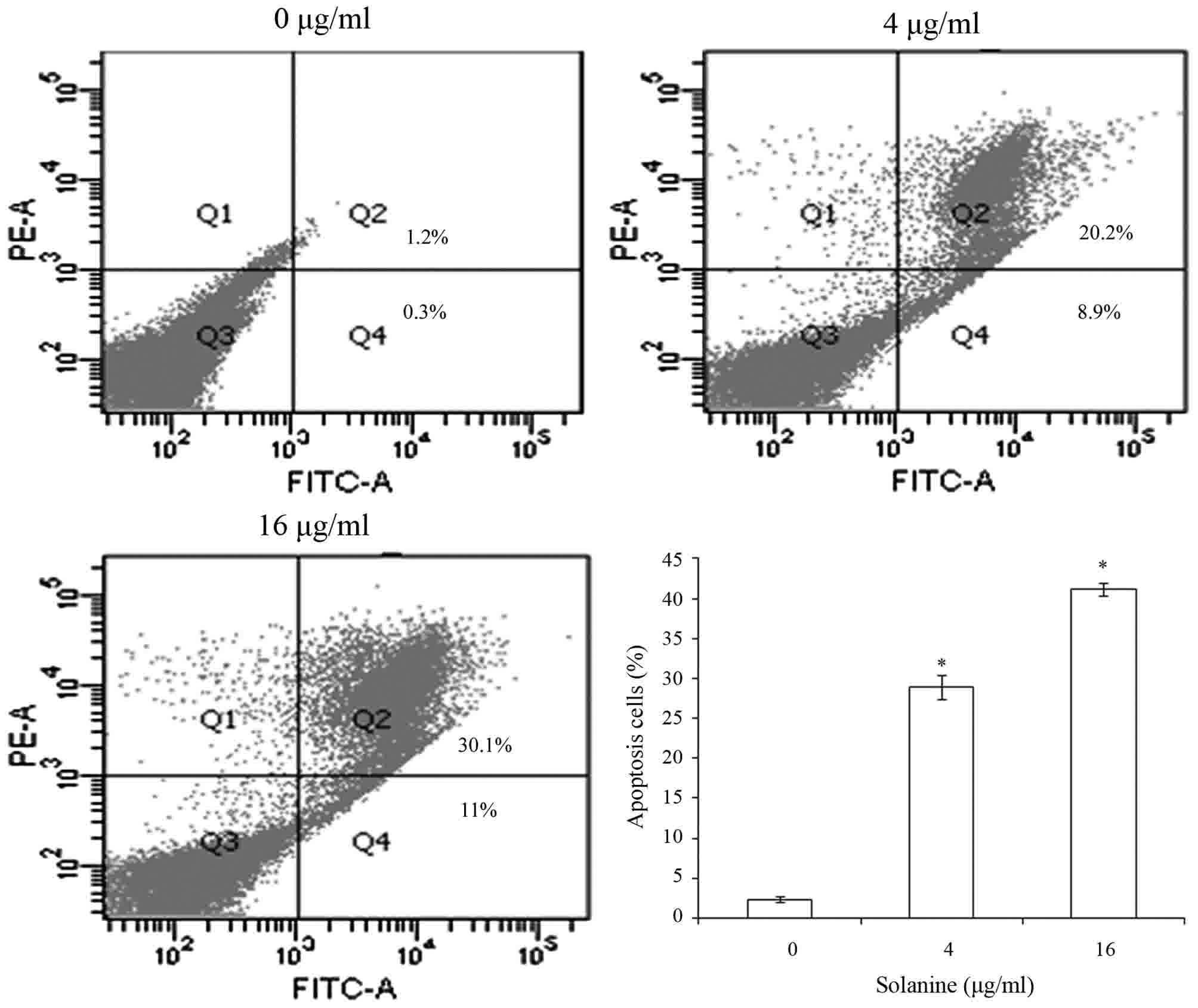
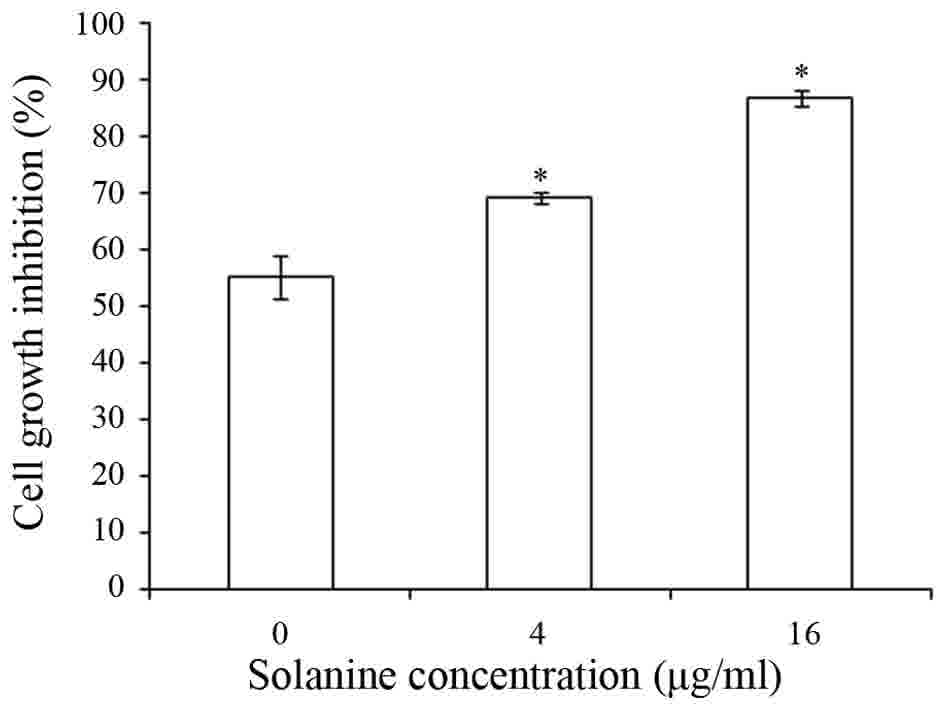
Solanine induced Jurkat cell
apoptosis
Subsequent to treatment of Jurkat cells with various
concentrations of Solanine (0, 4 and 16 µg/ml) for 24 h, the
apoptotic rate of Jurkat cells was detected by flow cytometry to
confirm that Solanine induces Jurkat cell apoptosis. The results
revealed that Solanine promotes Jurkat cell apoptosis in a
dose-dependent manner (Fig. 2;
P<0.05).
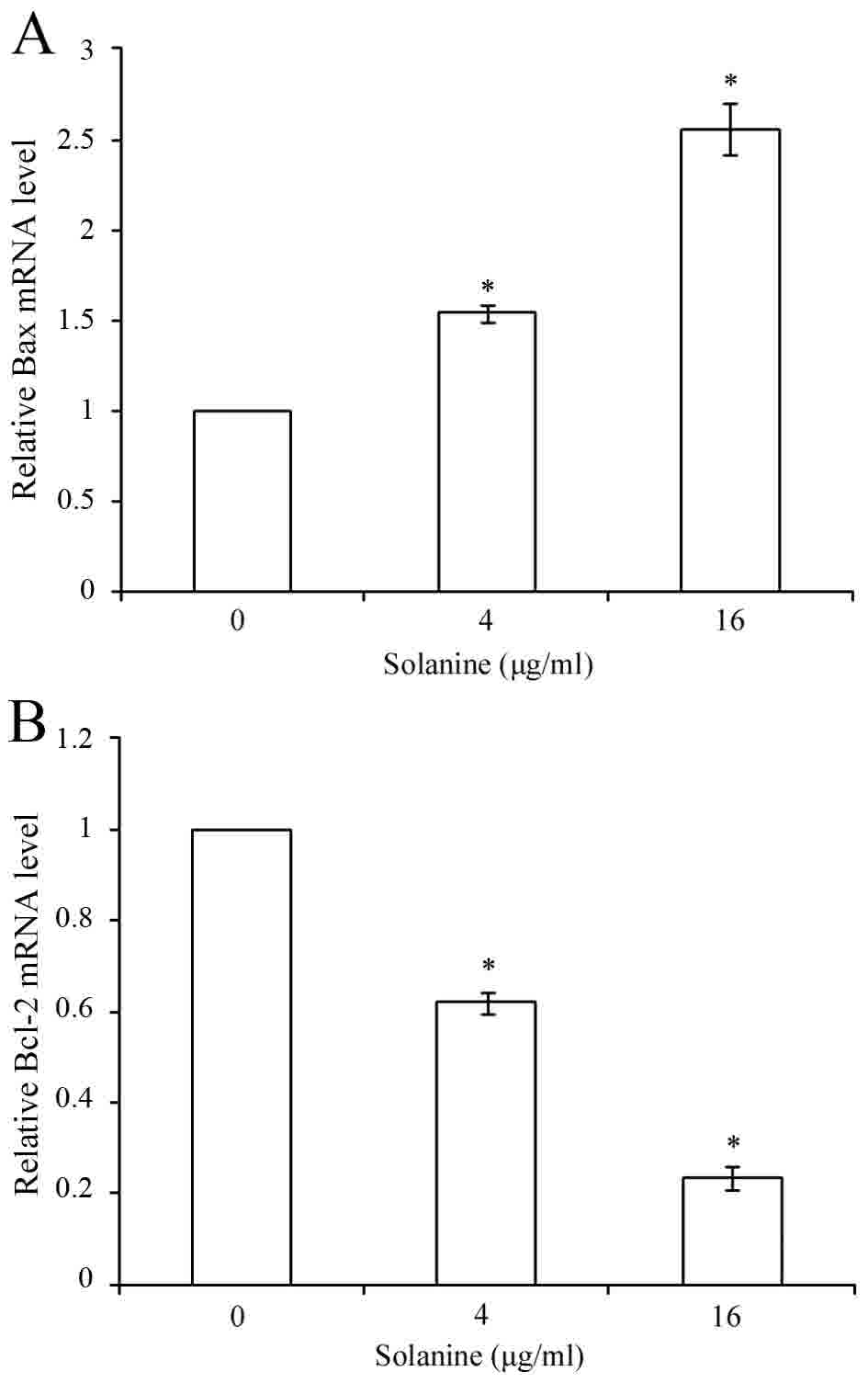
Effect of Solanine on mRNA levels of
Bcl-2 and Bax
RT-qPCR analysis was performed to assess whether
Solanine modulates the expression of Bcl-2 and Bax genes. The
results showed that Bcl-2 mRNA level decreased and Bax mRNA level
increased subsequent to treatment with various concentrations of
Solanine (Fig. 3; P<0.05).
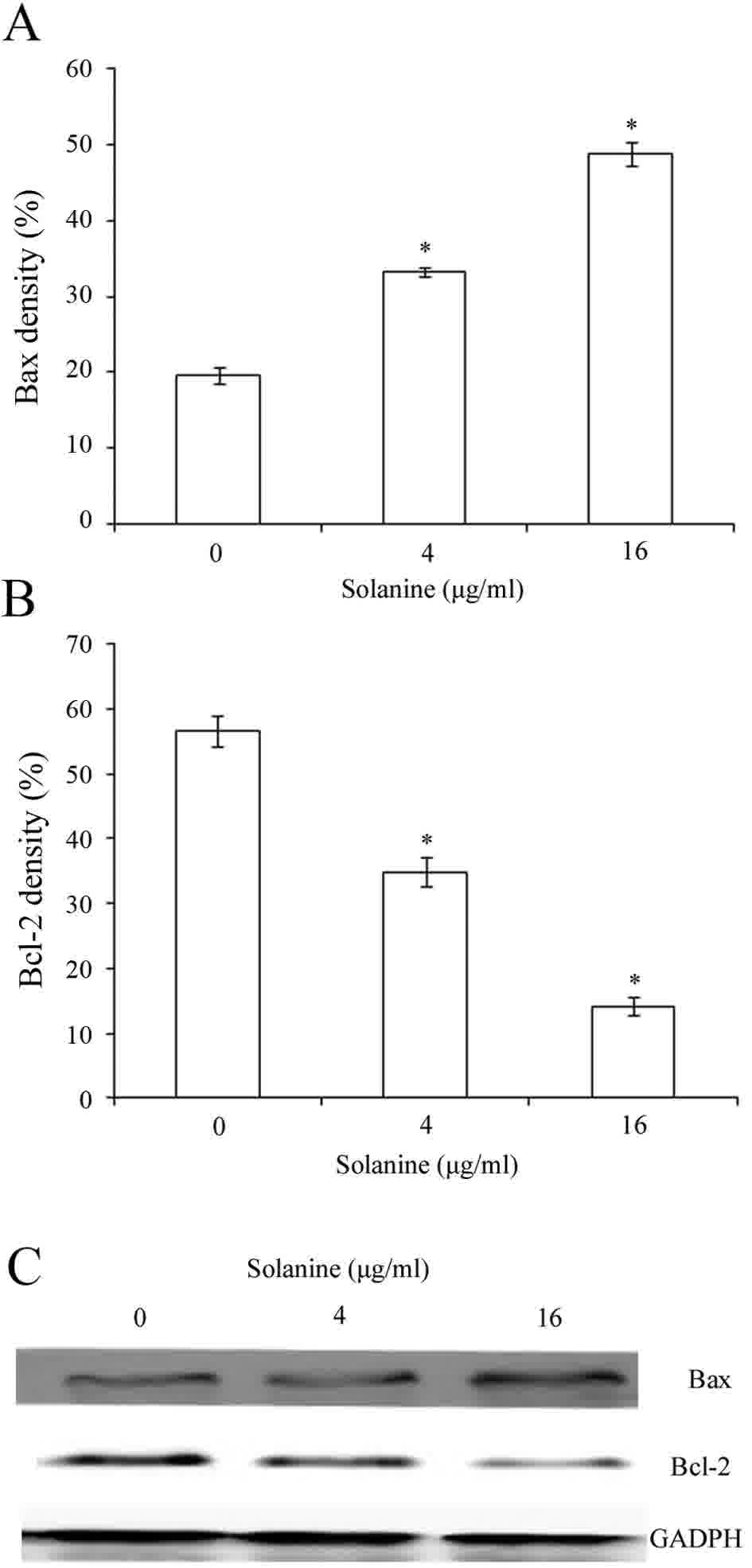
Effect of Solanine on the expression
of Bcl-2 and Bax protein in Jurkat cells
Bcl-2 and Bax are involved in cell apoptosis. The
expression of Bcl-2 and Bax was measured to explore the molecular
mechanism underlying Solanine-induced apoptosis. The western blot
assay showed that the expression of Bcl-2 decreased significantly
while the expression of Bax increased subsequent to Jurkat cell
incubation with various concentrations of Solanine for 24 h
(Fig. 4; P<0.05).
Solanine enhanced chemosensitivity of
Jurkat cells to Adriamycin
Based on the aforementioned findings, the present
study explored whether Solanine could increase the chemosensitivity
of Jurkat cells to Adriamycin. The cells were incubated with
various concentrations of Solanine (0, 4 and 16 µg/ml) in the
presence of Adriamycin for 24 h, and inhibition of cell
proliferation was measured by CCK-8 assay. As shown in Fig. 5, Solanine significantly increased the
Adriamycin-induced inhibitory rate of Jurkat cell proliferation,
which indicated that Solanine enhanced sensitivity to Adriamycin
compared to the controls [Cells treated with Adriamycin (0.15 µg/l)
in the absence of Solanine].
Discussion
Glycoalkaloids are secondary plant metabolites that
contain nitrogen and are found in Solanaceous plants and possess
anticarcinogenic activity (13).
Solanine is a steroidal glycoalkaloid (9). Studies have shown that Solanine has
antitumor potency (6–9), and inhibited the proliferation of U937
cells (14). In addition, data has
shown that Solanine has antitumor activity in other types of
cancer. Solanine has been shown to induce apoptosis of mice breast
cancer cells by inducing Bax expression and decreasing Bcl-2
expression (14). Solanine also
inhibited the activity of matrix metalloproteinase (MMP)-2 and
MMP-9 by suppressing the phosphoinositide 3-kinase/Akt and c-Jun
N-terminal kinase signaling pathways at non-toxic doses, causing
inhibition of migration and invasion in melanoma cells (7). Additionally, Solanine suppressed
pancreatic cancer cell migration and invasion by inhibiting MMP-2
and MMP-9 expression (9,10). Solanine downregulated the Bcl-2/Bax
ratio and processed the capase-3 zymogen into an active form,
thereby promoting pancreatic cancer cell apoptosis (10). However, to the best of our knowledge,
the effect of Solanine on T-ALL cells remains unknown. The present
study aimed to investigate the cellular functions of Solanine to
elucidate the mechanism by which it contributes to promote
apoptosis and inhibit proliferation in T-ALL Jurkat cells in
vitro.
The CCK-8 assay showed that Solanine significantly
inhibited Jurkat cell proliferation in a time- and dose-dependent
manner (Fig. 1). Therefore, the
results of the present study demonstrated that Solanine treatment
inhibited T-ALL Jurkat cell proliferation in vitro.
Apoptosis, also termed programmed cell death, is
characterized by morphological changes, including cell shrinkage,
chromatin condensation and membrane blebbing without disruption of
the plasma membrane (15). Apoptosis
plays an important role in homeostasis (16). Therefore, apoptosis plays a crucial
role in cancer treatment (17). In
the present study, Solanine induced apoptosis of Jurkat cells,
which was demonstrated by flow cytometry (Fig. 2).
Apoptosis is triggered by two pathways; one is the
death receptor-mediated extrinsic pathway, and the other is the
mitochondrial-dependent intrinsic pathway (17,18). Bcl-2
family members are key components of the mitochondrial-dependent
intrinsic apoptosis pathway (19).
Bcl-2 family members are classified into three subgroups: The
pro-apoptotic proteins, including Bax and Bak; the anti-apoptotic
proteins, including Bcl-2, myeloid cell leukemia-1 and Bcl-extra
large; and the BH3-only proteins, including BH3 interacting domain
death agonist, p53 upregulated modulator of apoptosis and Noxa
(20). Bax and Bcl-2 are the most
characterized apoptosis regulators in mitochondrial-associated
apoptosis (21). Bax was the first
identified pro-apoptotic protein member of the Bcl-2 protein
family, which is able to promote apoptosis (21). In the presence of apoptotic stimuli,
Bax translocates to the mitochondria, promoting the release of
cytochrome c into the cytosol, leading ultimately to apoptotic cell
death (22). Bcl-2, a major
anti-apoptotic protein of the Bcl-2 family inhibits cells apoptosis
by protecting mitochondrial membrane integrity and blocking the
release of cytochrome c (22). The
Bax/Bcl-2 ratio determines whether a cell will survive or undergo
apoptosis (18). It has been reported
that a reduced level of Bax and increased level of Bcl-2 affect the
relapse rate of patients with ALL (21). In the present study, RT-qPCR revealed
that Solanine inhibited Bcl-2 and promoted Bax mRNA expression
(Fig. 3). In addition, the results of
the present study indicated that Solanine increased Bax protein
expression and decreased Bcl-2 protein expression in a
dose-dependent manner, promoting the apoptosis of Jurkat cells
(Fig. 4). Therefore, the results of
the present study confirmed that Solanine regulates the expression
of apoptosis-associated genes and proteins in the T-ALL Jurkat cell
line.
Adriamycin is a highly effective anthracycline that
is widely used in chemotherapy treatment of a wide range of cancer,
including leukemia. In the present study, it was demonstrated that
Solanine significantly enhanced the cytotoxicity of Adriamycin in
T-ALL cells. The results indicated that Solanine may sensitize
T-ALL cells to Adriamycin. The results from the present study
indicated the potential of Solanine as an attractive therapeutic
strategy for T-ALL.
In conclusion, the present results indicated that
Solanine possesses antitumor activity in Jurkat cells.
Additionally, the results showed that the anticancer activity of
Solanine in Jurkat cells may be associated with inhibition of
proliferation and induction of apoptosis by regulating the
expression of apoptosis-associated genes and proteins. Solanine may
significantly increase the chemosensitivity of Jurkat cells to
Adriamycin. Therefore, these findings may provide a novel approach
for the development of T-ALL therapy using Solanine.
Acknowledgements
This study was supported by the Natural Science
Foundation of Shandong Province (grant no. ZR2014HL032), Projects
of Medical and Health Technology Development Program in Shandong
Province (grant no. 2014WS0183) and Shandong Science and Technology
Committee (grant no. 2010GSF10264).
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chopra A, Soni S, Verma D, Kumar D,
Dwivedi R, Vishwanathan A, Vishwakama G, Bakhshi S, Seth R, Gogia
A, et al: Prevalence of common fusion transcripts in acute
lymphoblastic leukemia: A report of 304 cases. Asia Pac J Clin
Oncol. 11:293–298. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Durinck K, Goossens S, Peirs S, Wallaert
A, Van Loocke W, Matthijssens F, Pieters T, Milani G, Lammens T,
Rondou P, et al: Novel biological insights in T-cell acute
lymphoblastic leukemia. Exp Hematol. 43:625–639. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
D'Angelo V, Iannotta A, Ramaglia M,
Lombardi A, Zarone MR, Desiderio V, Affinita MC, Pecoraro G, Di
Martino M, Indolfi P, et al: EZH2 is increased in paediatric T-cell
acute lymphoblastic leukemia and is a suitable molecular target in
combination treatment approaches. J Exp Clin Cancer Res. 34:832015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Akahane K, Sanda T, Mansour MR, Radimerski
T, DeAngelo DJ, Weinstock DM and Look AT: HSP90 inhibition leads to
degradation of the TYK2 kinase and apoptotic cell death in T-cell
acute lymphoblastic leukemia. Leukemia. 30:219–228. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shen KH, Liao AC, Hung JH, Lee WJ, Hu KC,
Lin PT, Liao RF and Chen PS: α-solanine inhibits invasion of human
prostate cancer cell by suppressing epithelial-mesenchymal
transition and MMPs expression. Molecules. 19:11896–11914. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lu MK, Shih YW, Chien Chang TT, Fang LH,
Huang HC and Chen PS: α-solanine inhibits human melanoma cell
migration and invasion by reducing matrix metalloproteinase-2/9
activities. Biol Pharm Bull. 33:1685–1691. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kenny OM, McCarthy CM, Brunton NP, Hossain
MB, Rai DK, Collins SG, Jones PW, Maguire AR and O'Brien NM:
Anti-inflammatory properties of potato glycoalkaloids in stimulated
Jurkat and Raw 264.7 mouse macrophages. Life Sci. 92:775–782. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lv C, Kong H, Dong G, Liu L, Tong K, Sun
H, Chen B, Zhang C and Zhou M: Antitumor efficacy of α-solanine
against pancreatic cancer in vitro and in vivo. PLoS One.
9:e878682014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun H, Lv C, Yang L, Wang Y, Zhang Q, Yu
S, Kong H, Wang M, Xie J, Zhang C and Zhou M: Solanine induces
mitochondria-mediated apoptosis in human pancreatic cancer cells.
Biomed Res Int. 2014:8059262014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gao SY, Wang QJ and Ji YB: Effect of
solanine on the membrane potential of mitochondria in HepG2 cells
and [Ca2+]i in the cells. World J Gastroenterol. 12:3359–3367.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ji YB, Gao SY, Ji CF and Zou X: Induction
of apoptosis in HepG2 cells by solanine and Bcl-2 protein. J
Ethnopharmacol. 115:194–202. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Friedman M: Chemistry and anticarcinogenic
mechanisms of glycoalkaloids produced by eggplants, potatoes, and
tomatoes. J Agric Food Chem. 63:3323–3337. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Friedman M, Lee KR, Kim HJ, Lee IS and
Kozukue N: Anticarcinogenic effects of glycoalkaloids from potatoes
against human cervical, liver, lymphoma, and stomach cancer cells.
J Agric Food Chem. 53:6162–6169. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kwak CH, Lee SH, Lee SK, Ha SH, Suh SJ,
Kwon KM, Chung TW, Ha KT, Chang YC, Lee YC, et al: Induction of
apoptosis and antitumor activity of eel skin mucus, containing
lactose-binding molecules, on human leukemic K562 cells. Mar Drugs.
13:3936–3949. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Scarfò L and Ghia P: Reprogramming cell
death: BCL2 family inhibition in hematological malignancies.
Immunol Lett. 155:36–39. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qi L, Ren K, Fang F, Zhao DH, Yang NJ and
Li Y: Over expression of BCL2 and low expression of caspase 8
related to TRAIL resistance in brain cancer stem cells. Asian Pac J
Cancer Prev. 16:4849–4852. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Christodoulou MI, Kontos CK, Halabalaki M,
Skaltsounis AL and Scorilas A: Nature promises new anticancer
agents: Interplay with the apoptosis-related BCL2 Gene Family.
Anticancer Agents Med Chim. 14:375–399. 2014. View Article : Google Scholar
|
|
19
|
Wang S, Zhou M, Ouyang J, Geng Z and Wang
Z: Tetraarsenictetrasulfide and arsenic trioxide exert synergistic
effects on induction of apoptosis and differentiation in acute
promyelocytic leukemia Cells. PLoS One. 10:e01303432015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Subburaj Y, Cosentino K, Axmann M,
Pedrueza-Villalmanzo E, Hermann E, Bleicken S, Spatz J and
García-Sáez AJ: Bax monomers form dimer units in the membrane that
further self-assemble into multiple oligomeric species
Introduction. Nat Commun. 6:80422015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cingeetham A, Vuree S, Dunna NR, Gorre M,
Nanchari SR, Edathara PM, Meka P, Annamaneni S, Digumarthi R, Sinha
S and Satti V: Influence of BCL2-938C>A and BAX-248G>A
promoter polymorphisms in the development of AML: Case-control
study from South India. Tumour Biol. 36:7967–7976. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Stamati L, Avgeris M, Kosmidis H, Baka M,
Anastasiou T, Piatopoulou D, Scorilas A and Gourgiotis D:
Overexpression of BCL2 and BAX following BFM induction therapy
predicts ch-ALL patients' poor response to treatment and short-term
relapse. J Cancer Res Clin Oncol. 141:2023–2036. 2015. View Article : Google Scholar : PubMed/NCBI
|