Introduction
Globally, ~200,000 women succumb to mortality from
cervical cancer every year and the age of onset of this disease has
decreased over time (1), indicating
that it presents a notable threat to the health of women. At
present, the treatment methods for cervical cancer include surgical
treatment, chemotherapy, radiotherapy, biological therapy and
traditional Chinese medicine. In clinical practice, surgical
treatment with radiotherapy or chemotherapy is the gold-standard.
However, this treatment option has a number of shortcomings,
including damage to the body and toxicity. Therefore, it is of
interest to identify a novel treatment method for cervical
cancer.
Allyl isothiocyanate (AITC) is widely distributed in
cruciferous plants and their by-products, including mustard and
horseradish (2,3). A number of studies have considered AITC
to be associated with tumors. For instance, AITC may arrest cancer
cells in mitosis, which in turn leads to apoptosis via B-cell
lymphoma 2 (Bcl-2) protein phosphorylation (4–6). AITC may
affect the proliferation of various tumor cells by inducing
apoptosis and cell cycle arrest, or by inhibiting their invasion
and metastasis, including in breast (7), bladder (8–10), lung
(11), colon (12,13), liver
(14,15) and prostate cancer cells (16). On the basis of the inhibitory effect
AITC has on the proliferation of bladder cancer cells,
population-based survey results have suggested that the intake of
cruciferous plants may improve the survival rate of patients with
bladder cancer (17), which may
provide further evidence for the anticancer effect of AITC.
However, few studies have reported on whether AITC has anticancer
effects on cervical cancer. Therefore, in the present study,
cervical cancer HeLa cells were treated with varying concentrations
of AITC in order to investigate the effect of AITC on the viability
and apoptosis of HeLa cells. In addition, the gene expression of
Bcl-2 and Bcl-2-associated X protein (Bax), which are associated
with apoptosis, was detected and the potential mechanisms
underlying the effects of AITC on the apoptosis of HeLa cells was
preliminarily discussed.
Materials and methods
Culture of HeLa cells
The cervical cancer HeLa cell line was sourced from
the American Type Culture Collection (Manassas, VA, USA) and was
cultured in Dulbecco's modified Eagle medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) containing 10% fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc.), prior to being
incubated with 5% CO2 at 37°C. Cells were passaged every
2–3 days, during which the culture medium in the culture dish was
removed and discarded, and the cells were rinsed with
phosphate-buffered saline (PBS) twice, prior to being digested with
trypsin for 1 min at 37°C. Subsequently, the adherent cells were
pipetted into single cell suspensions in PRMI-1640 medium
(Invitrogen; Thermo Fisher Scientific, Inc.) and centrifuged at
1,000 × g for 5 min at 37°C. The supernatant was discarded. The
cells were resuspended with 1 ml culture medium and then 200 µl was
removed for subculture (subculture ratio 1:5).
Cell viability using a Cell Counting
kit-8 (CCK8) assay
The HeLa cells, which had been digested and counted
during the logarithmic growth phase, were seeded onto 96-well
plates at a density of 1×104 cells/well. After 12 h of
incubation, the cells were treated with varying concentrations (0,
5, 15 and 45 µM) of AITC (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany). The treatment duration was set as 24, 48 and 72 h at a
temperature of 37°C. A total of 5 replicate wells were used for
each concentration and a blank control group was treated with an
equivalent volume of PBS (7).
Following treatment, the cells were rinsed using PBS three times
and 100 µl CCK8 reagent (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan) was added to each well (the ratio of CCK8 reagent
to medium was 1:10). The cells were incubated at 37°C in the dark
for 2 h. Subsequently, the absorbance values were determined using
an enzyme microplate reader at a wavelength of 450 nm and the cell
viability rate was calculated according to following expression:
Cell viability rate=(Atreatment group-Ablank
control group)/(Acontrol group-Ablank control
group), where A denotes the absorbance value.
Apoptosis detection by flow
cytometry
The HeLa cell suspension was added to a 6-well plate
at a density of 1×105 cells/well. After 12 h of
incubation, the cells were treated with varying concentrations of
AITC (0, 5, 15 and 45 µM) for 48 h. A total of 3 parallel samples
were used for each concentration. Following completion of the
treatment, the cells were digested with trypsin for 1 min at 37°C,
washed twice with PBS, centrifuged at 1,000 × g for 5 min at 37°C
and resuspended in PBS. Subsequently, 1×105 cells were
counted and centrifuged again at 1,000 × g for 5 min at 37°C. The
supernatant was discarded and 195 µl Annexin V-fluorescein
isothiocyanate (FITC), as part of a cell apoptosis detection kit
(Beyotime Institute of Biotechnology, Haimen, China) was added for
resuspension and agitated gently to mix following the addition of 5
µl Annexin V-FITC. The mixture was gently agitated to mix again
following the addition of 10 µl propidium iodide staining solution
(part of the aforementioned cell apoptosis detection kit). Next,
the mixture was incubated at room temperature in the dark for 20
min, prior to being placed in an ice bath at 0°C for 20 min. The
apoptosis rate was subsequently detected using a Gallios™ flow
cytometer (Version no. A75199AA; BD Biosciences, Franklin Lakes,
NJ, USA) and analyzed using FCS Express 3.0 (DeNovo software,
Glendale, CA, USA) and a cell apoptosis detection kit (Beyotime
Institute of Biotechnology, Haimen, China).
Detection of Bax and Bcl-2 mRNA
levels
The concentrations of AITC were set to 0, 5, 15 and
45 µM. After 48 h of treatment at 4°C, total RNA was extracted from
the HeLa cells using the TRIzol RNA extraction reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). The purity and content of RNA were
detected using a nucleic acid and protein analyzer. At the same
time, the integrity of RNA was identified by 1% agarose gel
electrophoresis (Abcam, Cambridge, MA, USA). RNA (1 µg) was used
for cDNA synthesis through reverse transcription using a reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) kit
(Takara Biotechnology Co., Ltd., Dalian, China), according to the
manufacturer's protocols. The primers of all genes were as follows:
Bax forward, TCCTCATCGCCATGCTCAT and reverse,
CCTTGGTCTGGAAGCAGAAGA; Bcl-2 forward, GATGACCGAGTACCTGAACC and
reverse, CAGGAGAAATCGAACAAAGGC; and β-actin forward,
TGCTGTGTTCCCATCTATCG and reverse, TTGGTGACAATACCGTGTTCA. The
2−∆∆Cq method was used for quantification (18), and an RT-qPCR reaction system was
established according to the following conditions: 5 µl 2 X
SYBR-Green mixture (Qiagen GmbH, Hilden, Germany), 0.5 µl cDNA, 0.5
µl primer and 4 µl purified H2O. Reaction conditions
were as follows: Following pre-denaturation at 95°C for 10 min
(denaturation at 95°C for 15 sec, annealing and extension at 60°C
for 60 sec), 40 PCR cycles were performed on a ViiA7 quantitative
fluorescence PCR machine (ABI Corporation, Lee's Summit, MO, USA)
at 95°C for 5 sec, 55°C for 30 sec and 72°C for 20 sec. A total of
3 parallel samples were set for each experiment, and β-actin was
used as the internal control gene.
Detection of Bax and Bcl-2 protein
levels using western blot analysis
The concentrations of AITC were set to 0, 5, 15 and
45 µM. Following 48 h of treatment at 37°C, the cells were added to
cell lysis solution (155 mM ammonium chloride, 10 mM sodium
bicarbonate and 0.5 mM EDTA; Miltenyi Biotec, Inc., Auburn, CA,
USA) for 2 h at 37°C, and placed in a homogenizer for
homogenization. Total cell protein was extracted using a total
protein extraction kit (BestBio Co., Shanghai, China), according to
the manufacturer's protocol, and was centrifuged at 1,000 × g for 5
min at 37°C, prior to the supernatant being obtained. The amount of
protein was determined using the Coomassie brilliant blue protein
assay kit (Shanghai Majorbio Pharmaceutical Technology Co., Ltd.,
Shanghai, China), according to the manufacturer's protocol. Next,
the Laemmli sample buffer, containing 60 mM Tris-Cl (pH 6.8), 2%
SDS, 10% glycerol, 5% β-mercaptoethanol and 0.01% bromophenol blue
(Sigma-Aldrich; Merck KGaA), was added and boiled at 100°C.
SDS-PAGE gel (6–12%; Sigma-Aldrich; Merck KGaA) was prepared as
follows: The protein sample (50 µg) was loaded for electrophoretic
separation for 3 h and then placed into the electric transducers,
in which the transfer buffer was added to the transmembrane for 1.5
h and the target protein was transferred onto nitrocellulose (NC)
membranes. Non-specific binding was blocked using PBS with Tween
(PBST; Sigma-Aldrich; Merck KGaA) containing 5% skimmed milk
powder, and agitated at room temperature for 2 h. Bcl-2 (dilution,
1:100; cat. no. MS-123-A1; Biotium, Inc., Freemont, CA, USA), Bax
(dilution, 1:100; cat. no. Rb-1486-R1; Biotium, Inc.) and GAPDH
antibodies (dilution, 1:100; cat. no. MS-168-P1; Biotium, Inc.)
were utilized. The membrane was washed 3 times with PBST and the
horseradish peroxidase-conjugated rabbit anti-mouse IgG antibody
(dilution, 1:1,000; cat. no. A-21422; Sigma-Aldrich; Merck KGaA)
was added and incubated in a 4°C refrigerator overnight. The
membrane was washed using PBST for 30 min, and the secondary
horseradish peroxidase-conjugated rabbit anti-mouse IgG antibody
(dilution, 1:1,000; cat. no. A-2418, Sigma-Aldrich; Merck KGaA),
prepared with PBST (containing 2.5% skimmed milk powder), was added
and was incubated in a shaker at 58°C for 60 min. The NC membranes
were washed with PBST 3 times and was then evenly coated with
reagents from an enhanced chemiluminescence detection kit
(Sigma-Aldrich; Merck KGaA) in a darkroom for 5 min. Following
washing, the film was placed in a fixing solution (40% methanol and
7% acetic acid) for 5 min at 37°C, and hung to dry following
flushing. The gel imaging system was sourced from ABI Corporation.
The optical densities of the protein bands of Bax, Bcl-2 and GADPH
were analyzed using the GIS-2020D gel image analysis system
(Sigma-Aldrich; Merck KGaA), while the expression intensities of
Bax and Bcl-2 proteins were expressed as the ratio of the optical
density of the Bax protein band to the optical density of the
β-actin protein band and the ratio of the optical density of the
Bcl-2 protein band to the optical density of the β-actin protein
band by using SPSS 19.0 software (IBM Corp., Armonk, NY, USA).
Statistical methods
Data were statistically analyzed using SPSS 19.0
software (IBM Corp.). Normally distributed data are presented as
the mean ± standard deviation. The normally distributed data in
different groups were compared using one-way analysis of variance
and the data of two groups were compared using Fisher's least
significant difference test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effect of AITC on the viability of
Hela cells
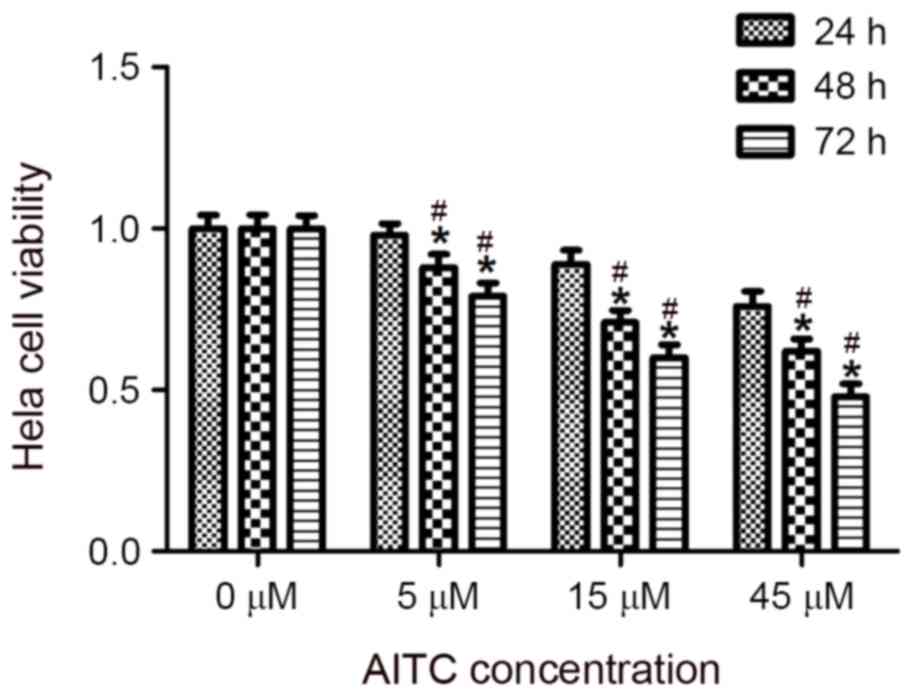
HeLa cells were treated with varying concentrations
of AITC for different durations, and a CCK8 assay was conducted to
determine the effect of AITC on the viability of HeLa cells, the
results of which are presented in Table
I and Fig. 1. The results
revealed that AITC inhibited the viability of HeLa cells, an effect
that was most significant when the cells were treated with 45 µM
AITC for 72 h (P=0.024).
 | Table I.Effect of AITC on the viability of
HeLa cells. |
Table I.
Effect of AITC on the viability of
HeLa cells.
|
| Treatment duration,
h |
|---|
|
|
|
|---|
| Treatment
concentration, µM | 24 | 48 | 72 |
|---|
| 0 | 1.00±0.042 | 1.00±0.043 | 1.00±0.041 |
| 5 | 0.98±0.036 |
0.88±0.040a,b |
0.79±0.042a,b |
| 15 |
0.89±0.043a |
0.71±0.037a,b |
0.60±0.041a,b |
| 45 |
0.76±0.045a |
0.62±0.038a,b |
0.48±0.039a,b |
Effect of AITC on the apoptosis of
HeLa cells
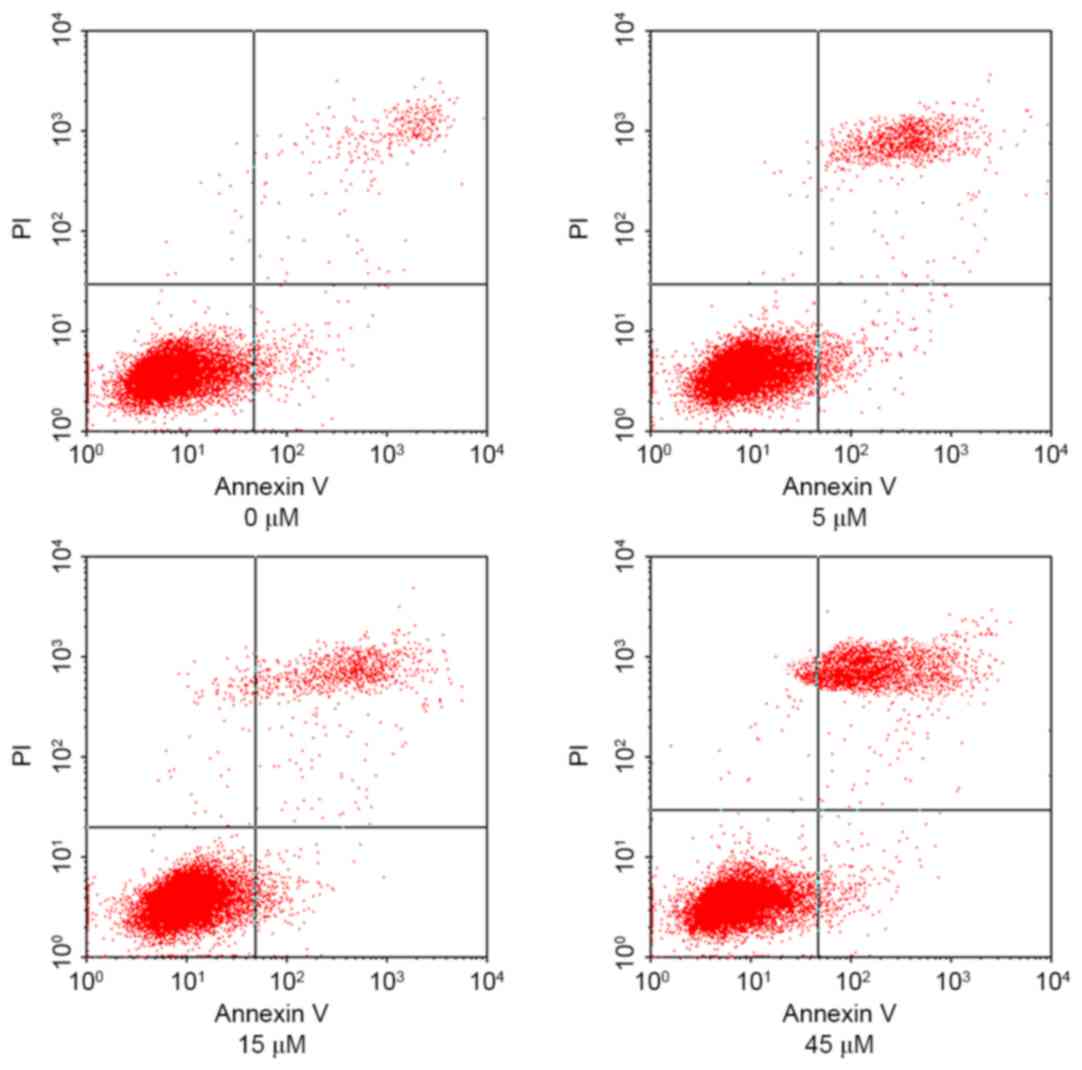
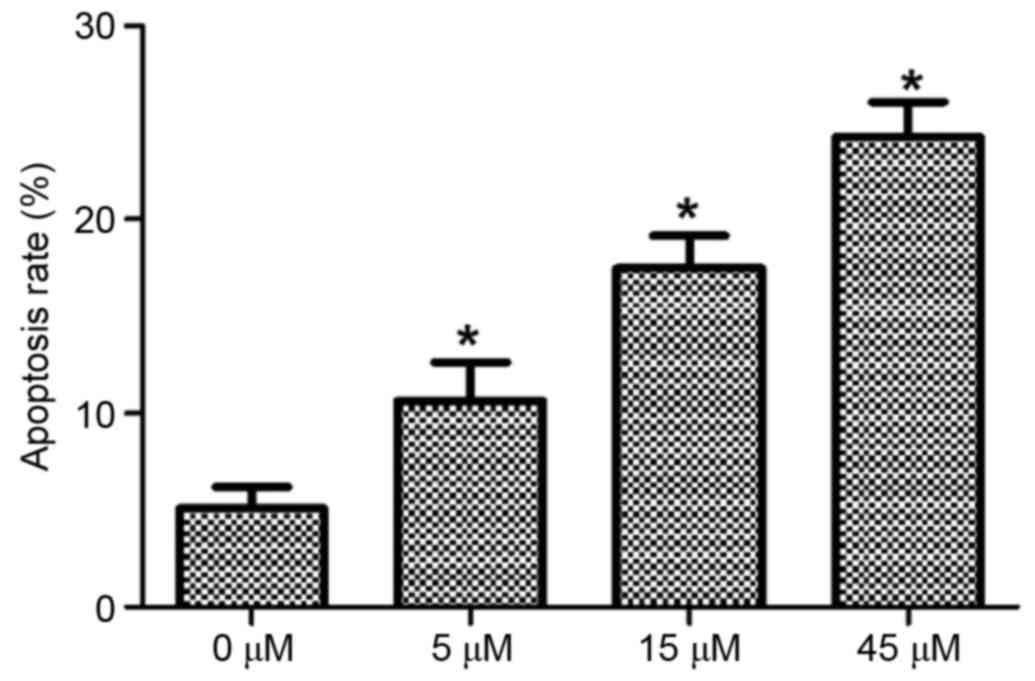
HeLa cells were treated with 0, 5, 15 and 45 µM AITC
for varying durations and the apoptosis rates of HeLa cells in
different groups were detected using flow cytometry, the results of
which are presented in Fig. 2 (in
which the right upper quadrant presents the results for late
apoptotic cells and the right lower quadrant presents the results
for early apoptotic cells) and Fig.
3. As determined by flow cytometry, the apoptosis rate in the
control group was 5.08±1.12%, which was significantly lower
compared with 10.65±1.98, 17.49±1.68 and 24.26±1.83% in the 5, 15
and 45 µM AITC treatment groups, respectively. The apoptosis rates
in the treatment groups were significantly higher than the
apoptosis rate in the control group at all drug concentrations
(P<0.05).
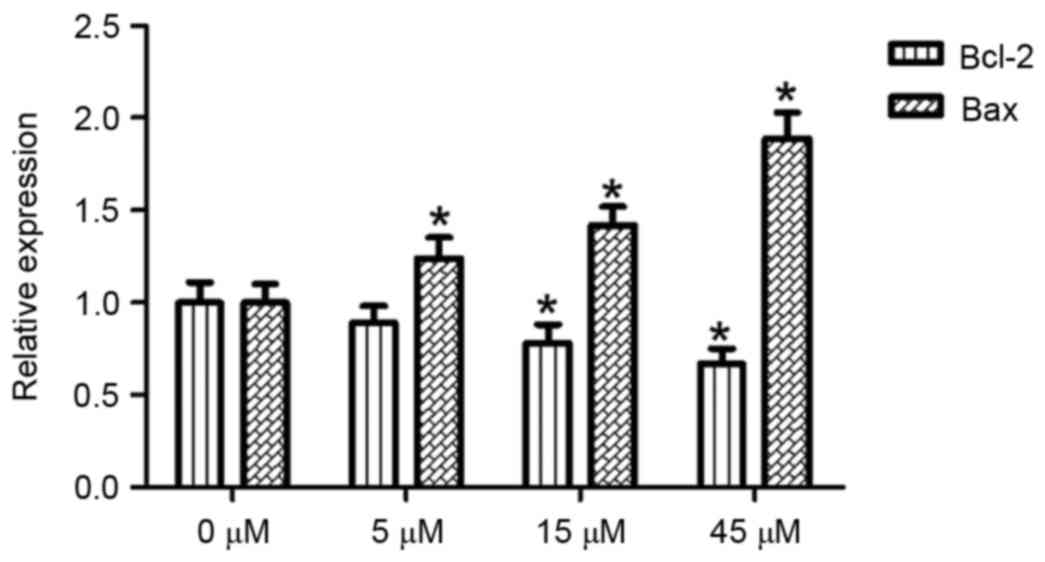
Effects of AITC on Bax and Bcl-2 mRNA
expression in HeLa cells
With the 0 µM AITC treatment group serving as a
control and β-actin acting as an internal reference gene, Bax and
Bcl-2 mRNA expression levels in HeLa cells were detected using a
semi quantitative method following 48 h of AITC treatment, the
results of which are presented in Fig.
4. Compared with the control group, the Bcl-2 mRNA expression
level was significantly decreased in the 15 and 45 µM treatment
groups (P<0.05). The differences between the treatment groups
(at all concentrations) and the control group were statistically
significant (P<0.05).
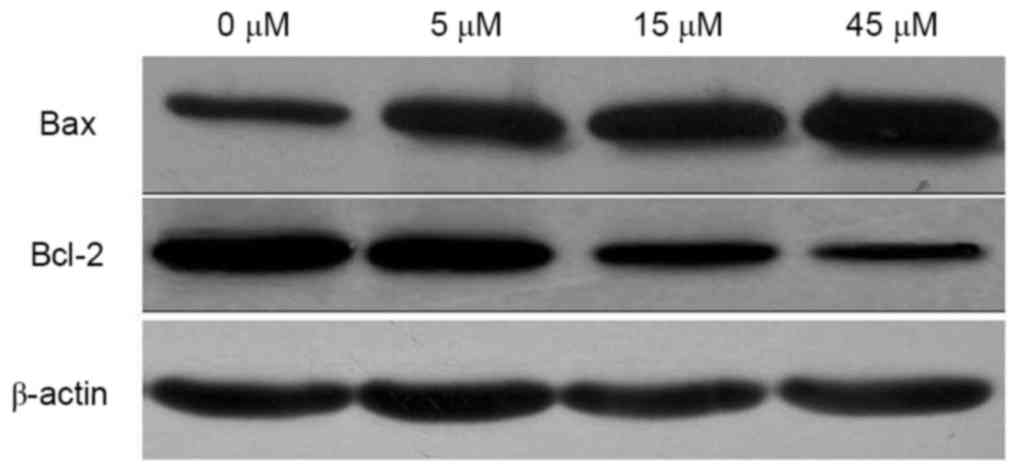
Effects of AITC on Bax and Bcl-2
protein expression in HeLa cells
Following treatment with 0, 5, 15 and 45 µM AITC for
48 h, Bax and Bcl-2 protein expression levels in HeLa cells were
detected. As illustrated in Fig. 5
and Table II, compared with the
control group, the expression level of Bax protein was
significantly upregulated, whereas the expression level of Bcl-2
protein was significantly downregulated following AITC treatment
(all P<0.001).
| Table II.Effects of AITC treatment on Bax and
Bcl-2 protein expression levels. |
Table II.
Effects of AITC treatment on Bax and
Bcl-2 protein expression levels.
|
| Concentration,
µM |
|---|
|
|
|
|---|
| Protein | 0 | 5 | 15 | 45 | P-value |
|---|
| Bax | 0.58±0.07 |
0.71±0.06a |
0.83±0.04a |
1.02±0.08a | <0.001 |
| Bcl-2 | 0.98±0.09 | 0.89±0.08 |
0.65±0.07a |
0.50±0.05a | <0.001 |
| Bcl-2/Bax | 1.69±0.14 |
1.25±0.13a |
0.78±0.11a |
0.49±0.12a | <0.001 |
Discussion
At present, the drugs used for the treatment of
cervical cancer are often highly toxic and induce severe side
effects, which may cause damage to patient tissues during the
treatment of cancer and may reduce the chance of survival (2,3).
Therefore, the identification of a safe, effective and low toxicity
anticancer drug has attracted interest from researchers, and the
development of these drugs from natural sources has become an
important strategy in the development of antitumor drugs. As a type
of plant chemical that is prevalent in the natural diet of humans,
AITC may affect the proliferation of a variety of tumor cells
(19,20), and has attracted extensive attention
from researchers. The anticancer effect of AITC has been verified
by studies in vivo and in vitro (21,22), but
the function of AITC in the treatment of cervical cancer has not
been reported. Therefore, the present study confirmed the effect of
AITC on cervical cancer cells through cell line experiments and
preliminarily explored the underlying mechanisms of this.
The inhibitory effect on cell viability was most
significant when the cells were treated with 45 µM AITC for 72 h
(P=0.024). At present, the effect of AITC on HeLa cells has not
previously been reported. However, a similar study was reported by
Hasegawa et al (23), which
revealed that isothiocyanate, including phenyl-ethyl isothiocyanate
and benzyl isothiocyanate, may inhibit HeLa cell viability through
cell cycle arrest. AITC is a type of isothiocyanate, which means
that the conclusion of the present study is consistent with that of
Hasegawa et al (23). In
addition, when HeLa cells were treated with AITC for 48 h, the
apoptosis rate of cells exhibited a dose-dependent increase,
suggesting that AITC may inhibit HeLa cell viability by inducing
cell apoptosis. However, the mechanism of AITC inhibiting cell
viability is complex. For example, AITC may inhibit HeLa cell
viability through cell cycle arrest and the induction of cell
apoptosis.
Bax and Bcl-2 have attracted attention due to their
regulation of cell apoptosis, and their function in promoting or
inhibiting cell apoptosis may be realized through their protein
expression (24). Bax and Bcl-2 are a
pair of positive-negative apoptosis regulating genes. In addition
to promoting cell apoptosis, Bax may also reduce the inhibitory
effect of Bcl-2 on cell apoptosis by forming a dimer with Bcl-2
(25). Therefore, the occurrence of
cell apoptosis is associated with an imbalance of Bcl-2/Bax
expression. The present study revealed that AITC may induce the
apoptosis of HeLa cells. By detecting Bcl-2 and Bax expression
levels, it was demonstrated that, with an increase in the
concentration of AITC, the expression levels of Bax mRNA and
protein increased whilst Bcl-2 mRNA and protein expression levels
decreased, resulting in a decrease in the ratio of Bcl-2/Bax
proteins. The results of the present study suggested that the
apoptosis of HeLa cells induced by AITC may be associated with an
imbalance in Bcl-2/Bax expression. Sávio et al (8) also revealed that the apoptosis induced
by AITC was associated with a Bcl-2/Bax expression imbalance.
The present study demonstrated that AITC may inhibit
HeLa cell viability through the induction of cell apoptosis and
that the imbalance of Bcl-2/Bax expression may be the mechanism
accounting for AITC-induced cell apoptosis, which may provide
evidence for further research for novel anticancer drugs for
cervical cancer. However, the results of the present study remain
far from being used for clinical applications and require further
verification in future studies.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xu K and Thornalley PJ: Studies on the
mechanism of the inhibition of human leukaemia cell growth by
dietary isothiocyanates and their cysteine adducts in vitro.
Biochem Pharmacol. 60:221–231. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bhattacharya A, Li Y, Wade KL, Paonessa
JD, Fahey JW and Zhang Y: Allyl isothiocyanate-rich mustard seed
powder inhibits bladder cancer growth and muscle invasion.
Carcinogenesis. 31:2105–2110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Geng F, Tang L, Li Y, Yang L, Choi KS,
Kazim AL and Zhang Y: Allyl isothiocyanate arrests cancer cells in
mitosis, and mitotic arrest in turn leads to apoptosis via Bcl-2
protein phosphorylation. J Biol Chem. 286:32259–32267. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xiao D, Srivastava SK, Lew KL, Zeng Y,
Hershberger P, Johnson CS, Trump DL and Singh SV: Allyl
isothiocyanate, a constituent of cruciferous vegetables, inhibits
proliferation of human prostate cancer cells by causing G2/M arrest
and inducing apoptosis. Carcinogenesis. 24:891–897. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu CL, Huang AC, Yang JS, Liao CL, Lu HF,
Chou ST, Ma CY, Hsia TC, Ko YC and Chung JG: Benzyl isothiocyanate
(BITC) and phenethyl isothiocyanate (PEITC)-mediated generation of
reactive oxygen species causes cell cycle arrest and induces
apoptosis via activation of caspase-3, mitochondria dysfunction and
nitric oxide (NO) in human osteogenic sarcoma U-2 OS cells. J
Orthop Res. 29:1199–1209. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bo P, Lien JC, Chen YY, Yu FS, Lu HF, Yu
CS, Chou YC, Yu CC and Chung JG: Allyl isothiocyanate induces cell
toxicity by multiple pathways in human breast cancer cells. Am J
Chin Med. 44:415–437. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sávio AL, da Silva GN and Salvadori DM:
Inhibition of bladder cancer cell proliferation by allyl
isothiocyanate (mustard essential oil). Mutat Res. 771:29–35. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bhattacharya A, Li Y, Geng F, Munday R and
Zhang Y: The principal urinary metabolite of allyl isothiocyanate,
N-acetyl-S-(N-allylthiocarbamoyl) cysteine, inhibits the growth and
muscle invasion of bladder cancer. Carcinogenesis. 33:394–398.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Savio AL, da Silva GN, de Camargo EA and
Salvadori DM: Cell cycle kinetics, apoptosis rates, DNA damage and
TP53 gene expression in bladder cancer cells treated with allyl
isothiocyanate (mustard essential oil). Mutat Res. 762:40–46. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tripathi K, Hussein UK, Anupalli R,
Barnett R, Bachaboina L, Scalici J, Rocconi RP, Owen LB, Piazza GA
and Palle K: Allyl isothiocyanate induces replication-associated
DNA damage response in NSCLC cells and sensitizes to ionizing
radiation. Oncotarget. 6:5237–5252. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lau WS, Chen T and Wong YS: Allyl
isothiocyanate induces G2/M arrest in human colorectal
adenocarcinoma SW620 cells through down-regulation of Cdc25B and
Cdc25C. Mol Med Rep. 3:1023–1030. 2010.PubMed/NCBI
|
|
13
|
Lai KC, Lu CC, Tang YJ, Chiang JH, Kuo DH,
Chen FA, Chen IL and Yang JS: Allyl isothiocyanate inhibits cell
metastasis through suppression of the MAPK pathways in epidermal
growth factor-stimulated HT29 human colorectal adenocarcinoma
cells. Oncol Rep. 31:189–196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hwang ES and Kim GH: Allyl isothiocyanate
influences cell adhesion, migration and metalloproteinase gene
expression in SK-Hep1 cells. Exp Biol Med (Maywood). 234:105–111.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Garcia A, Haza AI, Arranz N, Rafter J and
Morales P: Protective effects of isothiocyanates alone or in
combination with vitamin C towards N-nitrosodibutylamine or
N-nitrosopiperidine-induced oxidative DNA damage in the single-cell
gel electrophoresis (SCGE)/HepG2 assay. J ApplToxicol. 28:196–204.
2008.
|
|
16
|
Xu C, Shen G, Yuan X, Kim JH,
Gopalkrishnan A, Keum YS, Nair S and Kong AN: ERK and JNK signaling
pathways are involved in the regulation of activator protein 1 and
cell death elicited by three isothiocyanates in human prostate
cancer PC-3 cells. Carcinogenesis. 27:437–445. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tang L, Zirpoli GR, Guru K, Moysich KB,
Zhang Y, Ambrosone CB and McCann SE: Intake of cruciferous
vegetables modifies bladder cancer survival. Cancer Epidemiol
Biomarkers Prev. 19:1806–1811. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Louhivuori LM, Bart G, Larsson KP,
Louhivuori V, Näsman J, Nordström T, Koivisto AP and Akerman KE:
Differentiation dependent expression of TRPA1 and TRPM8 channels in
IMR-32 human neuroblastoma cells. J Cell Physiol. 221:67–74. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen NG, Chen KT, Lu CC, Lan YH, Lai CH,
Chung YT, Yang JS and Lin YC: Allyl isothiocyanate triggers G2/M
phase arrest and apoptosis in human brain malignant glioma GBM 8401
cells through a mitochondria-dependent pathway. Oncol Rep.
24:449–455. 2010.PubMed/NCBI
|
|
21
|
Zhu Y, Zhuang JX, Wang Q, Zhang HY and
Yang P: Inhibitory effect of benzyl isothiocyanate on proliferation
in vitro of human glioma cells. Asian Pac J Cancer Prev.
14:2607–2610. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gupta P, Kim B, Kim SH and Srivastava SK:
Molecular targets of isothiocyanates in cancer: Recent advances.
Mol Nutr Food Res. 58:1685–1707. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hasegawa T, Nishino H and Iwashima A:
Isothiocyanates inhibit cell cycle progression of HeLa cells at
G2/M phase. Anticancer Drugs. 4:273–279. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cory S and Adams JM: The Bcl2 family:
Regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Oltvai ZN, Milliman CL and Korsmeyer SJ:
Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that
accelerates programmed cell death. Cell. 74:609–619. 1993.
View Article : Google Scholar : PubMed/NCBI
|