Introduction
Pancreatic cancer (PC) is one of the leading causes
of cancer-associated mortality in developed and developing
countries and one of the most lethal malignant neoplasms in the
world (1–4). Based upon the estimates of GLOBOCAN
2012, >331,000 mortalities per year were caused by PC,
accounting for 4% of all mortalities, and PC was considered to be
the seventh leading cause of cancer-associated mortality in males
and females (1). With regards to
treatment, surgery is the only option with the potential to cure
PC; however, as a result of the concealed pathogenesis, rapid
progress and high metastatic rate, a limited number patients with
PC (15–20%) are candidates for radical surgery at the time of
disease diagnosis (5). Less than 30%
of the patients who undergo surgery and adjuvant chemotherapy
survive >5 years following treatment (5). Life expectancy is decreased by up to 98%
in patients with PC compared with healthy patients (6).
The causes of PC remain unknown, but certain risk
factors have been reported, including cigarette smoking, a family
history of PC, excessive alcohol consumption, diabetes mellitus,
obesity, dietary factors and a lack of physical exercise (7–10). The
present study focused on the Jumonji AT-rich interactive domain 1B
(JARID1B) gene, which is a member of JmjC domain-containing protein
family. JARID1B specifically removes the methyl residues from tri-,
di- and mono-methylated lysine 4 on histone H3 (H3K4) that are
associated with gene activation (11,12).
JARID1B acts as a transcriptional repressor due to its intrinsic
histone demethylase activity. Trimethylation at H3K4 is an
important histone modification associated with the activation of
transcribed genes, including PTEN (13), and JARID1B specifically demethylates
H3K4me3 (tri-methylated histone H3 at lysine 4) to a
transcriptionally inactive state that will repress the activation
of target genes (14).
Expression of JARID1B has been reported be elevated
in a number of different types of cancer, including breast cancer,
bladder cancer, lung cancer, colorectal cancer, prostate cancer and
malignant melanoma and is required for the proliferation of cancer
cells and tumor growth (11,15–22). It
has been reported that the depletion of JARID1B inhibited the
proliferation of breast cancer cells and restrained tumor growth in
xenografts (23) and a syngeneic
mouse mammary tumor model (24).
Similar results were obtained in lung, bladder and colorectal
tumors (19–21). To the best of our knowledge, the
present study was the first to reveal that the expression of
JARID1B was elevated in PC, and that this was responsible for the
inhibition of cell proliferation and tumor growth. Furthermore, it
was also previously revealed that JARID1B is associated with the
inactivation of phosphatase and tensin homolog (PTEN) in
hepatocellular carcinoma (13) which
was verified in the present study. The results of the present study
reveal a novel function of JARID1B in PC and may provide
perspective in order to develop novel therapeutic interventions for
PC.
Materials and methods
Clinical specimens and cell
culture
The present study was conducted with the approval of
the Institutional Ethical Review Board of the Second Affiliated
Hospital of Soochow University (Suzhou, Jiangsu, China). Between
June 2014 and April 2015, a total of 42 paired (25 male and 17
female, from 37 to 62 years old, mean 50.6 years old) PC specimens
and adjacent tissue samples, frozen in liquid nitrogen, were
obtained from the pathology laboratory at the Second Affiliated
Hospital of Soochow University. None of the patients had received
any antitumor treatments prior to biopsy. PC cell lines H6c7, MIA
PaCa-2, AsPC-1, BxPC-3, Su86.86, UACC462 were obtained from the
American Type Culture Collection (Manassas, VA, USA) and cultured
in adaptive culture medium (Sigma-Aldrich; Merck KgaA, Darmstadt,
Germany) according to ATCC and cultured at 37°C in 5%
CO2.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from freshly-frozen samples
or cells using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Total RNA was reverse
transcribed at 42°C for 60 min using a First Strand cDNA Synthesis
kit (Invitrogen; Thermo Fisher Scientific, Inc.). qPCR reactions
were conducted using Platinum SYBR Green qPCR SuperMix-UDG reagents
(Invitrogen; Thermo Fisher Scientific, Inc.) on the PRISM 7900HT
system (Applied Biosystems; Thermo Fisher Scientific, Inc.)
followed the following primers: JARID1B forward primer
5′-AGAGGCTGAATGAGCTGGAG-3′ and reverse primer
5′-TGGCAATTTTGGTCCATTTT-3′; GAPDH forward primer
5′-ATCACTGCCACCCAGAAGAC-3′ and reverse primer
5′-ATGAGGTCCACCACCCTGTT-3′. The thermocycling conditions were as
follows: 95°C for 10 min, followed by 40 cycles of 95°C for 10 sec,
50°C for 30 sec and 72°C for 20 sec. All reactions were performed
in triplicate and reactions without reverse transcriptase were used
as negative controls. GAPDH was used as an endogenous control and
the 2−ΔΔCq method was used to calculate the relative
expression levels (25).
Western blot analysis
Western-blot assay was used to analyze the
expressions of candidate proteins in indicated cell lines. Cells
were cultured in 100 mm dishes and radioimmunoprecipitation assay
buffer (Beyotime Institute of Biotechnology, Haimen, China)
containing protein inhibitor cocktail (cat. no. P9599;
Sigma-Aldrich; Merck KGaA) was input at 70–90% density. Whole
protein was extracted by centrifugation (13,000 × g) for 20 min at
4°C. And the concentration of proteins was measured by NanoDrop
(NanoDrop2000; Thermo Fisher Scientific, Inc.). Samples containing
30–50 ug of protein were separated by 10% SDS-PAGE. Proteins were
transferred to polyvinylidene fluoride membranes (Merck KGaA) which
was blocked in 5% bovine serum albumin (Beijing Seasky Bio
Technology, Co., Ltd., Beijing, China) or 5% skim milk at room
temperature for 1 h. Then the membrane was probed overnight at 4°C
in 5% bovine serum albumin (Beijing Seasky Bio Technology, Co.,
Ltd.) with primary antibodies (Cell Signaling Technology, Inc.,
Danvers, MA, USA) against Actin (1:2,000; cat. no. CST 3700S),
anti-JARID1B (1:1,000; cat. no. CST 3273T), anti-PTEN (1:1,000;
cat. no. CST 9188T), anti-protein kinase B (Akt) (1:1,000; cat. no.
CST 2920S), anti-p-Akt (1:1,000; cat. no. CST 4060T), anti-P13K
(1:1,000; cat. no. CST 4249T), anti-p-P13K (1:1,000; cat. no. CST
4228T), anti-p53 (1:1,000; cat. no. CST 2527T), anti-p21 (1:1,000;
cat. no. CST 2947T), anti-p27 (1:1,000; cat. no. CST 3686T),
anti-H3K4me3 (1:1,000; cat. no. CST 9751T), anti-K3K9me3 (1:1,000;
cat. no. CST 4658T), anti-H3K27me3 (1:1,000; cat. no. CST 9728T)
followed by washing in Tris-buffered saline with Tween-20 (TBST;
0.02M Tris PH 7.6, 0.8% NaCl, 01% Tween-20) and incubated in TBST
containing HRP secondary antibodies (1:10,000; cat. no. ZB2301;
ZSGB-BIO) for 1 h at room temperature. Following washing in TBST
again, the Chemiluminescent HRP substrate (cat. no. P90720; Merck
KGaA) was added, according to the manufacturer's protocol, and the
fluorescence was assessed using FluorChem E (version 4.1.3;
Proteinsimple, FE0444). All antibodies were purchased from Cell
Signaling Technology.
Establishment of cell lines
Human JARID1B shRNA was cloned into a pSuper-puro
vector (GENEWIZ, South Plainfield, NJ, USA) while JARID1B mRNA was
cloned into a pBabe-puro vector (GENEWIZ). A total of 10 µg plasmid
was transfected into Phoenix packaging cells by X-tremeGENE HP
(Roche Applied Science, Penzberg, Germany) and retrovirus
supernatants containing pSuper and pSuper-sh.JARID1B, pBabe and
pBabe-JARID1B were collected and filtered. AsPC-1 was incubated in
retrovirus supernatants contaning pBabe and pBabe-JARID1B, but
UACC-462 was incubated in retrovirus supernatants containing pSuper
and pSuper-shJARID1B 1 or 2. A total of 4 µg/ml polybrene
(Sigma-Aldrich; Merck KGaA) was added to accelerate the
transfection. Then AsPC-1-pBabe-JARID1B, UACC-462-pSuper-shJARID1B
1 or 2 and control cells were subsequently selected by puromycin (2
µg/ml) as previously described (26).
Proliferation assay
An MTT assay was used to detect the proliferative
rate of AsPC-1-pBabe-JARID1B, UACC-462-pSuper-shJARID1B 1 or 2 and
control cell lines. A total of 1,000 cells were plated into a
96-well plate and were cultured at 37°C. MTT was added at 70%
density and the 96-well plate was incubated for 4 h, followed by
the addition of 150 µl dimethyl sulfoxide in order to dissolve the
purple formazan. The optical density values at 492 nm were measured
using Multiskan 3 at 12, 24, 48 and 72 h.
In vivo tumor growth model
A total of 20 Male BALB/c nude mice (weight ~15 g)
aged 4–6 weeks were purchased from the Hunan Slac Jingda Laboratory
Animal Company, Ltd. (Changsha, China) and provided with sterile
food and water in the specific pathogen free (SPF) laboratory
animal room (22°C, 40% humidity, noise ≤60 dB, clean pass box). All
animals were used in accordance with institutional guidelines and
the current experiments were approved by the Animal Care and Use
Ethics Committe. For the tumor growth assay, AsPC-1-pBabe-JARID1B,
UACC-462-pSuper-shJARID1B 2 and control cells were resuspended in
phosphate-buffered saline and 4×106 cells (100 µl) were
subcutaneously injected into the axilla of the nude mice. Six weeks
later, the mice were sacrificed using 1 mg/kg chloral hydrate, and
the tumors were dissected and weighed. Animal handling and research
protocols were approved by the Animal Care and Use Ethics
Committee.
Chromatin immunoprecipitation
(ChIP)-qPCR
A ChIP kit was purchased from EMD Millipore
(Billerica, MA, USA) and ChIP experiments were performed as
previously described (27).
Immunoprecipitated DNA was analyzed on the ABI PRISM 7900HT
sequence detection system. The PTEN promoter primers of −1,235 to
−1,072 bp (forward primer, 5′-CGCCCAGCTCCTTTTCCC-3′; reverse
primer, 5′-CTGCCGCCGATTCTTAC-3′) and 260 to 512 bp (forward primer,
5′-GCTCGCACCCAGAGCTAC-3′; reverse primer, 5′-GGAGAGAACTGAGC-3′)
used for detection of promoters after ChIP are available upon
request.
Statistical analysis
Statistical analysis data are presented as the mean
± standard deviation Associations between JAEID1B and PTEN, Akt,
P13K, p53, p21 and p27 expression in PC tissues was assessed using
Spearman's rank correlation. Comparisons between different groups
were undertaken using one-way analysis of variance with Tukey's
post hoc test. P<0.05 was considered to indicate a statistically
significant difference. Statistical analysis was performed using
SPSS 11.0 software for Windows (SPSS, Inc., Chicago, IL, USA).
Results
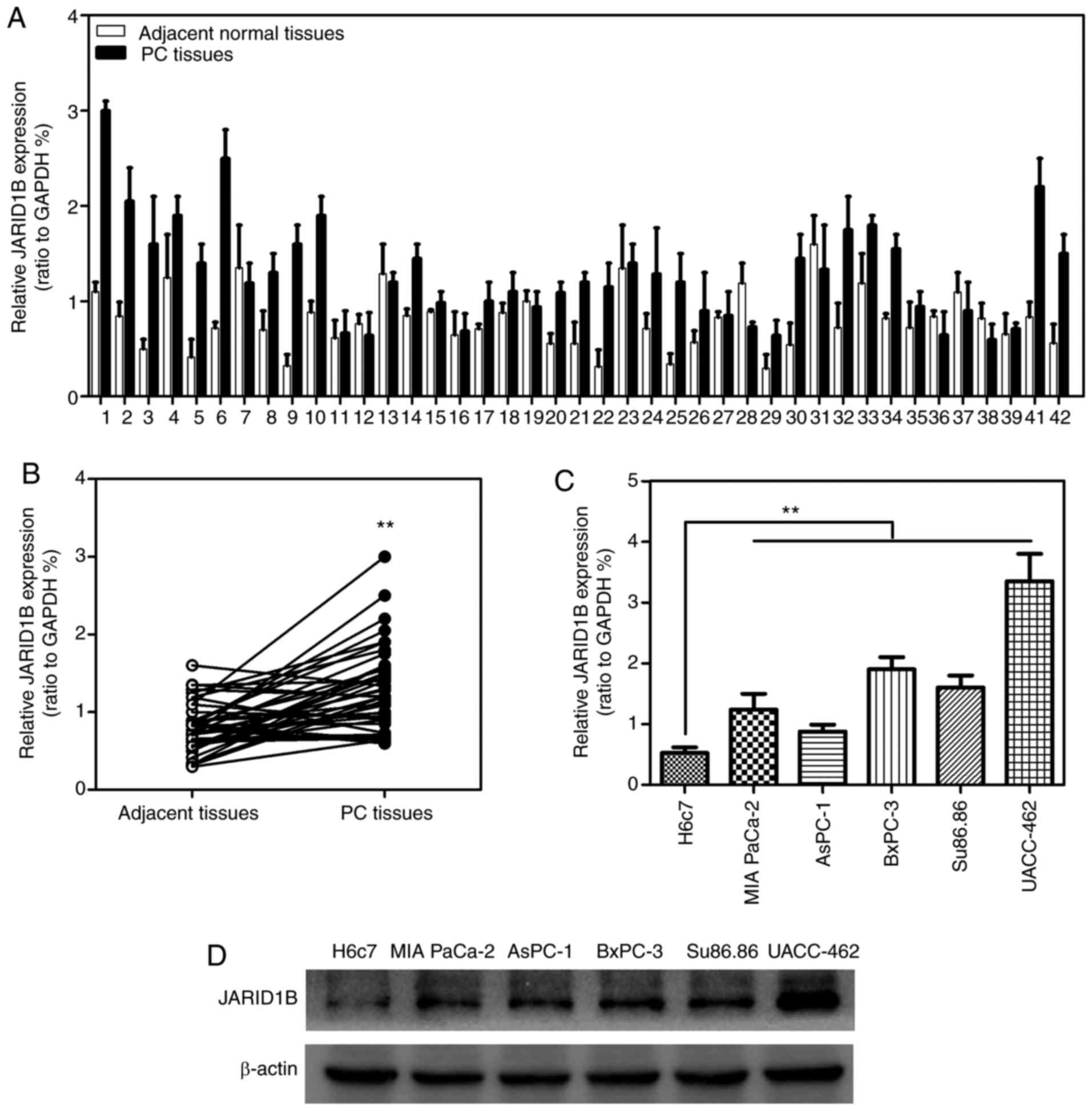
JARID1B is highly expressed in PC
In order to investigate whether JARID1B was
associated with the progression of PC, the expression levels of
JARID1B in 42 patient PC samples RT-qPCR. As demonstrated in
Fig. 1A and B, JARID1B was
significantly overexpressed in 42 PC tissues compared with the
normal adjacent tissues. To further confirm the elevated expression
of JARID1B in PC, the expression levels of JARID1B in distinct PC
cell lines were determined. The results revealed that JARID1B was
significantly overexpressed in PC cell lines including MIA PaCa-2,
AsPC-1, BxPC-3, Su86.86 and UACC-462 compared with expression in
the control cell line H6c7 (Fig. 1C).
Furthermore, the expression levels of JARID1B in these cell lines
described above were verified using western blot analysis (Fig. 1D). The results revealed that JARID1B
was overexpressed in PC.
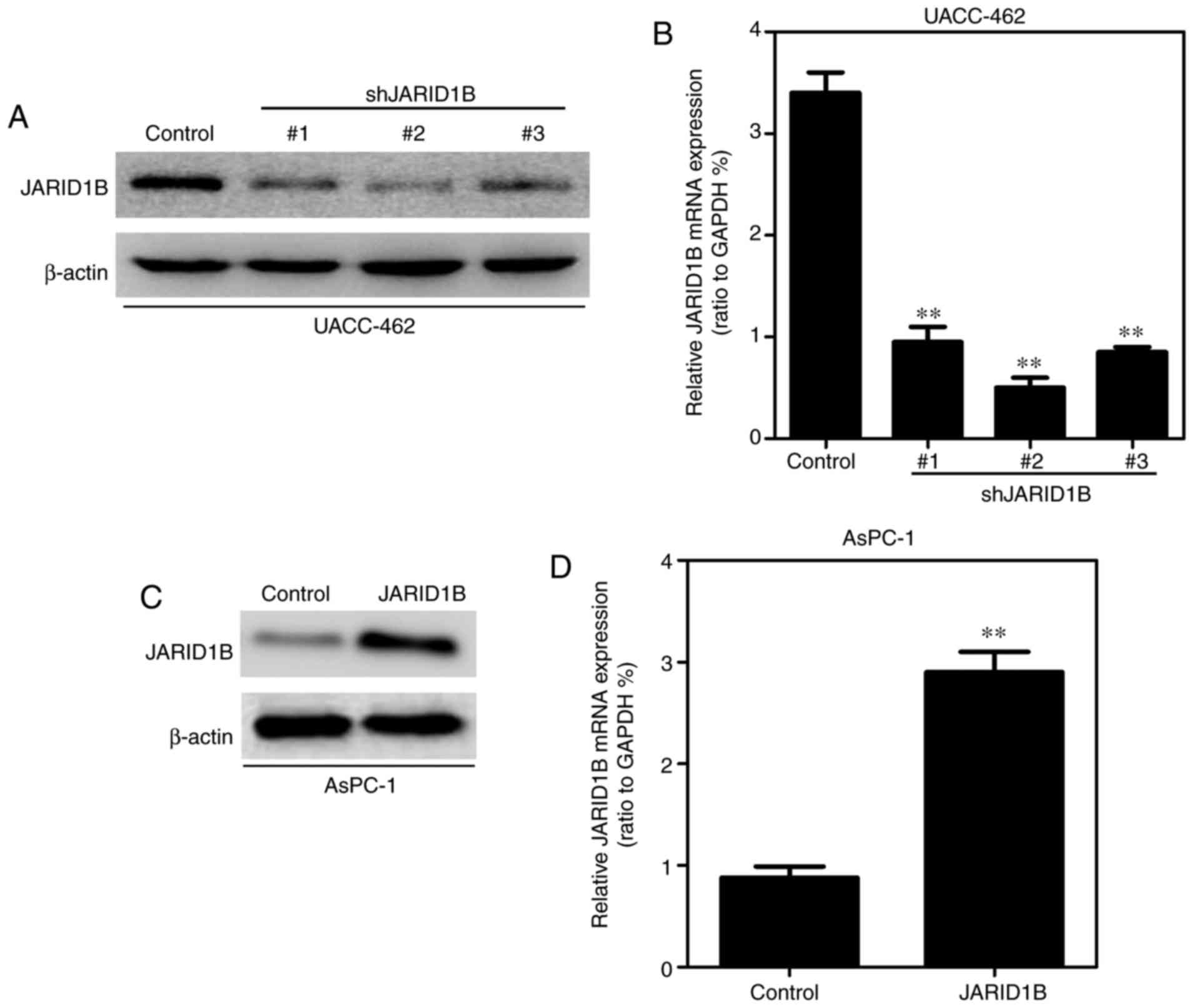
JARID1B promotes the proliferative
capacity of PC cells in vitro and in vivo
In order to further identify the role of JARID1B in
PC cells, the pancreatic cell lines exhibiting overexpression or
silencing of JARID1B were established using a lentiviral vector and
the expression levels of JARID1B were examined by western blot
analysis and RT-qPCR. As demonstrated in Fig. 2A, the expression levels of JARID1B in
UACC-462 cells with three different shRNAs were examined by western
blot analysis and JARID1B shRNA2 was the most effective. Similar
results were observed in RT-qPCR (Fig.
2B). Overexpression of JARID1B in the AsPC-1 cell line was
observed in western blot analysis (Fig.
2C) and RT-qPCR (Fig. 2D).
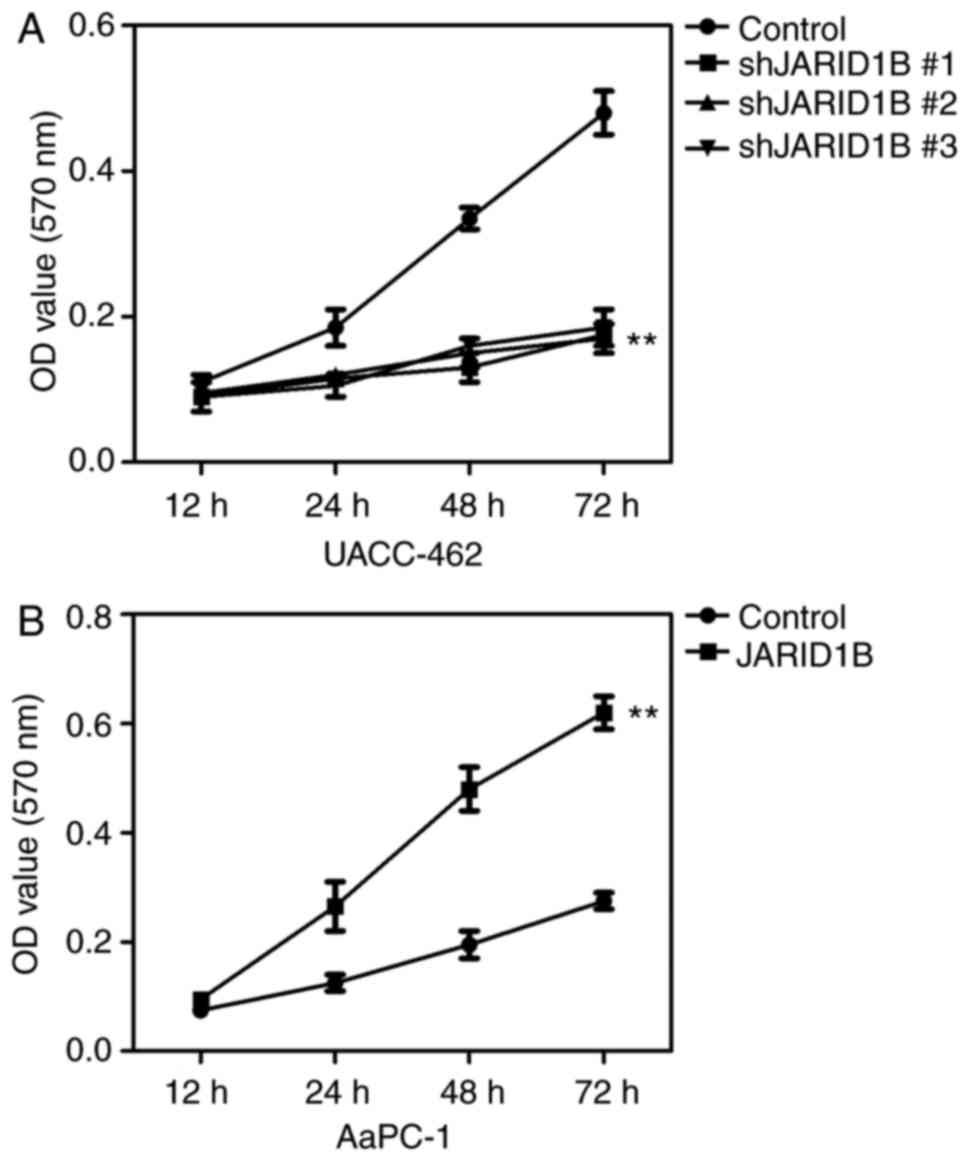
Based on the established cell lines, the effect of
JARID1B silencing on the proliferation of PC cancer cells was
subsequently determined using an MTT assay. The results revealed
that silencing of JARID1B significantly inhibited the proliferation
of UACC-462 cells (Fig. 3A), while
overexpression of JARID1B promoted the proliferation of AsPC-1
cells (Fig. 3B).
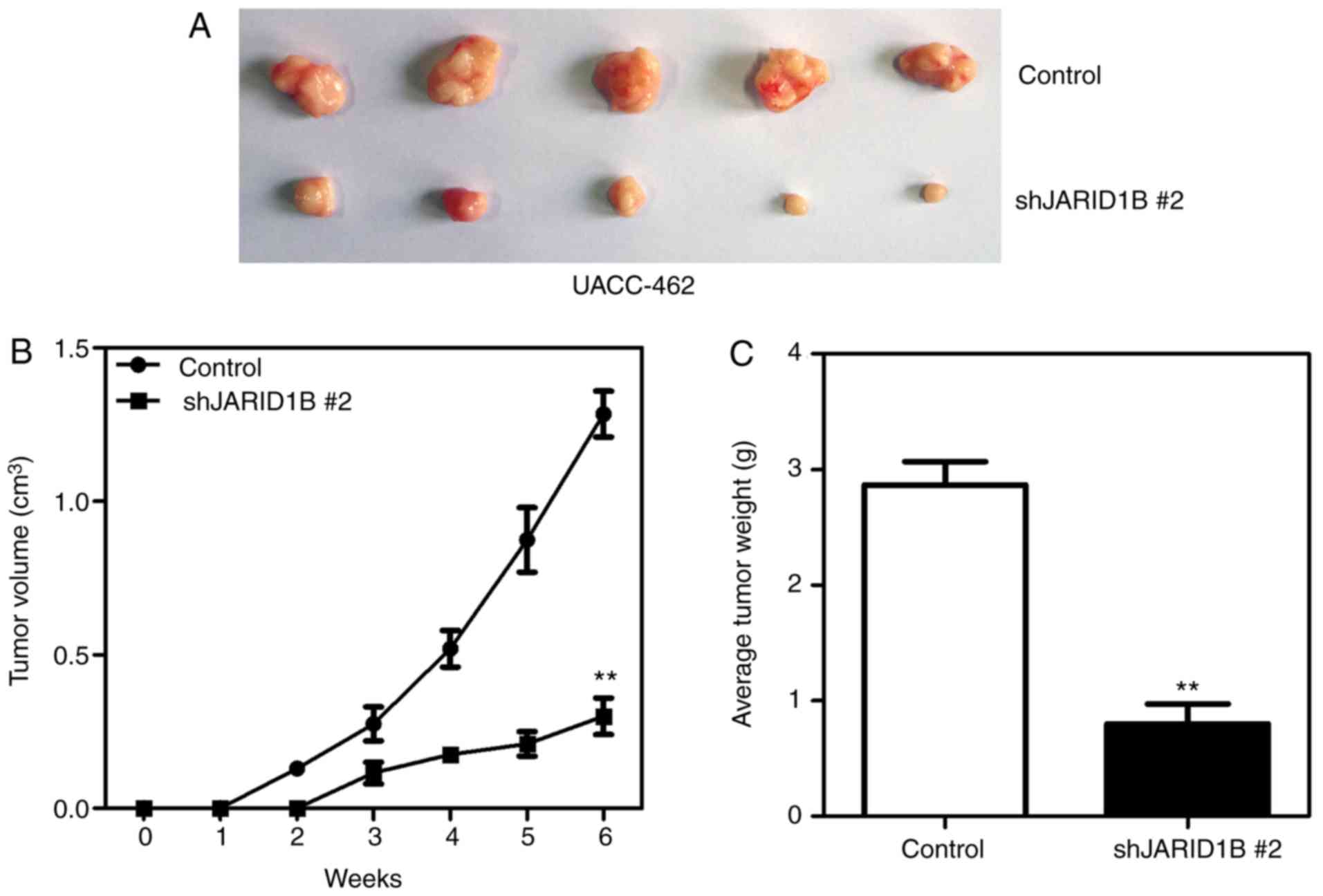
In order to further confirm the effect of JARID1B on
the proliferation of PC cells, xenografts were used to reveal
whether JARID1B affected the proliferation rate of pancreatic
tumors. AsPC-1-pBabe-JARID1B, UACC-462-pSuper-shJARID1B 2 and
control cells were injected subcutaneously into the axilla of nude
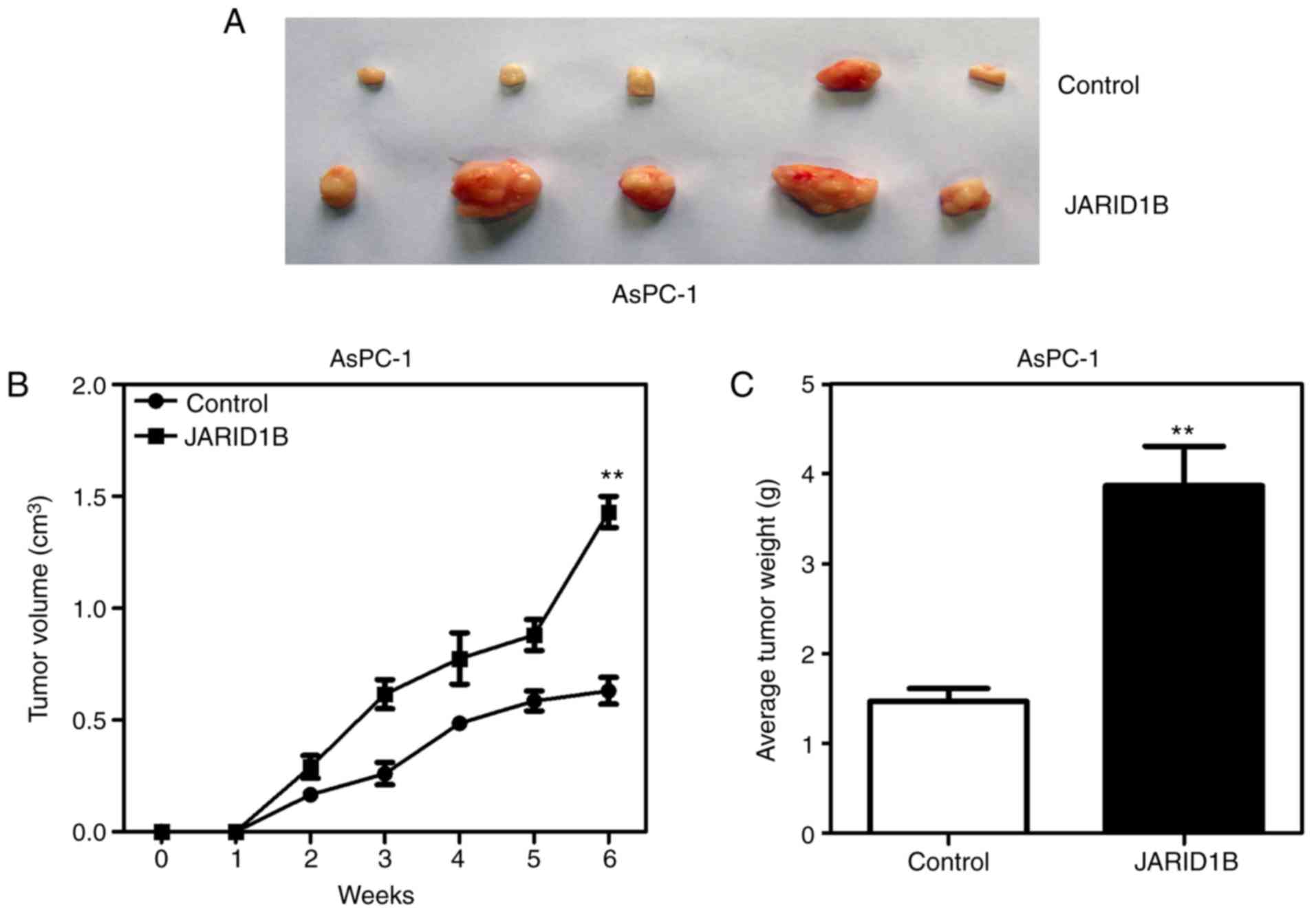
mice. As demonstrated in Fig. 4,
silencing of JARID1B significantly inhibited the proliferation of
pancreatic tumors formed in nude mice (Fig. 4A), which were smaller in size
(Fig. 4B) and lighter in weight
(Fig. 4C). However, overexpression of
JARID1B significantly promoted the proliferation of pancreatic
tumors formed by the injection of AsPC-1 cells (Fig. 5A), in terms of tumor size (Fig. 5B) and weight (Fig. 5C). The aforementioned results
demonstrated that JARID1B may promote the proliferation of PC cells
in vitro and tumorigenesis in vivo.
JARID1B regulates PTEN expression
through the demethylation of H3K4me3
It has been demonstrated that JARID1B may affect the
demethylation of H3K4me3 and that the activation of PTEN is
regulated by H3K4me3 (13). In order
to determine whether JARID1B regulates the tumorigenesis of PC
through the PTEN/Akt signaling pathway, the expression levels of
proteins participating in the PTEN/Akt signaling pathway were
examined. As demonstrated in Fig. 6,
silencing of JARID1B significantly elevated the expression of PTEN
in UACC-462 cells (Fig. 6A), while
overexpression of JARID1B inhibited the expression of PTEN in
AsPC-1 cells (Fig. 6B). It has been
reported that PTEN, as well as p53, p27 and p21, is able to
regulate the phosphoinositide 3-kinase (PI3K)/Akt pathway in a
number of types of tumor cell (28,29).
Therefore, whether or not JARID1B-mediated inhibition of PTEN may
affect the expression of the aforementioned proteins was
subsequently investigated. Notably, silencing of JARID1B followed
by ectopic expression of PTEN significantly inhibited the level of
phosphorylated PI3K and Akt but increased the level of p53, p27 and
p21 (Fig. 6A), while overexpression
of JARID1B induced the opposite effect (Fig. 6B). In further research, silencing of
JARID1B significantly increased the mRNA level of PTEN and
p53 (Fig. 6C), while
overexpression of JARID1B decreased the mRNA level of PTEN
and p53 (Fig. 6D). Western
blot analysis and RT-qPCR revealed that JARID1B did not affect the
level of P13K and Akt (Fig. 6A-D).
These data revealed that JARID1B may inhibit the transcription of
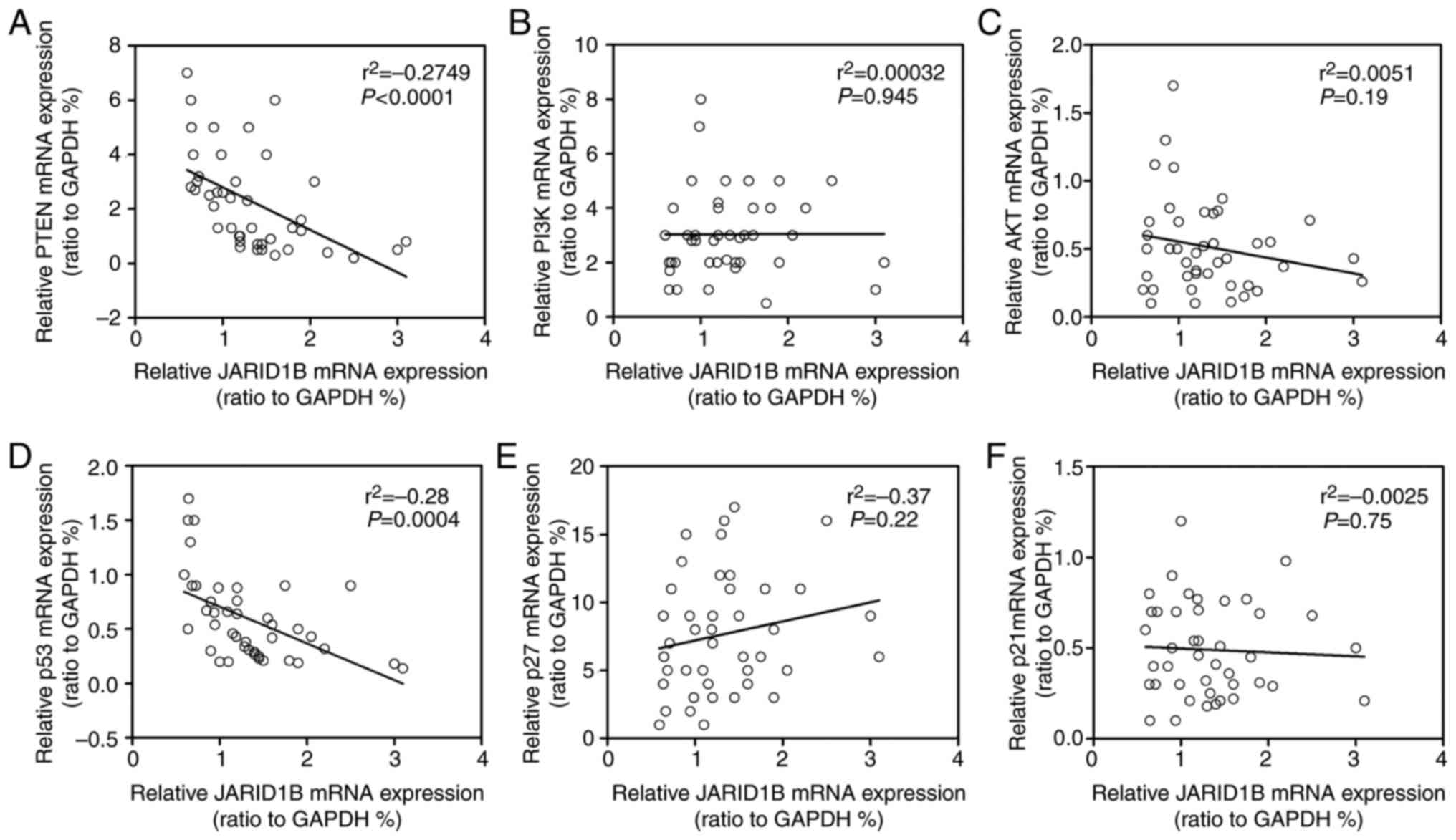
PTEN and p53. In order to further confirm the
correlation between JARID1B and PTEN or p53,
Spearman's rank correlation analysis was performed. As demonstrated
in Fig. 7, the mRNA level of
JARID1B was inversely correlated with the mRNA levels of
PTEN (Fig. 7A) and p53
(Fig. 7D), while no association was
observed between JARID1B and P13K, Akt, p27 or p21 (Fig. 7B, C, E and F). Based upon these
aforementioned data, JARID1B inhibited the expression of PTEN and
p53 at the RNA level.
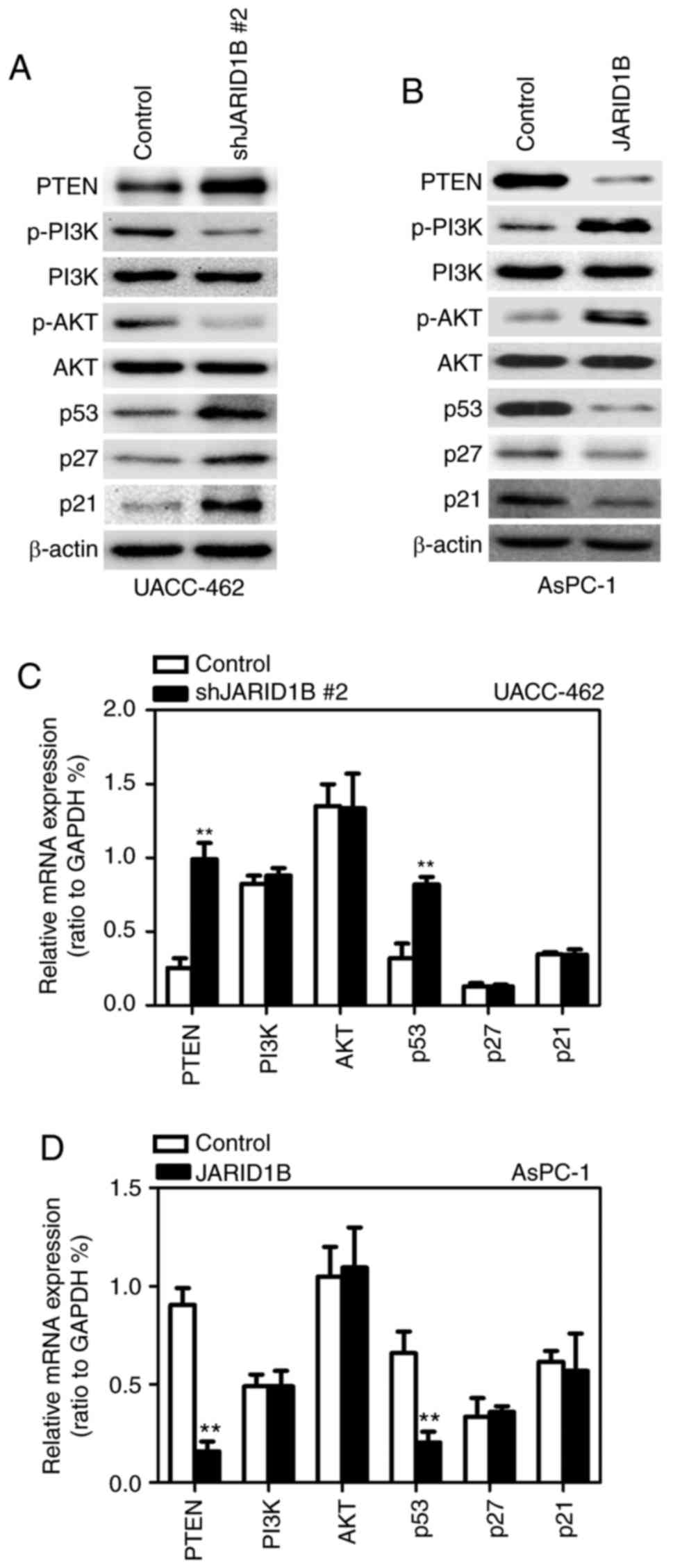 | Figure 6.JARID1B activates the PTEN signaling
pathway. (A) Western blot analysis of the proteins involved in the
PTEN signaling pathway, and p53, p27 and p21 in UACC-462 cells with
silencing of JARID1B. (B) Western blot analysis of the proteins
involved in the PTEN signaling pathway, and p53, p27 and p21 in
AsPC-1 cells overexpressing JARID1B. (C) RT-qPCR analysis of the
proteins involved in the PTEN signaling pathway, and p53, p27 and
p21 in UACC-462 cells with silencing of JARID1B. (D) RT-qPCR
analysis of the proteins involved in the PTEN signaling pathway,
and p53, p27 and p21 in AsPC-1 cells overexpressing JARID1B.
**P<0.01 compared with control. Error bars represent the
standard deviation. JARID1B, Jumonji AT-rich interactive domain 1B;
PTEN, phosphatase and tensin homolog; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction. |
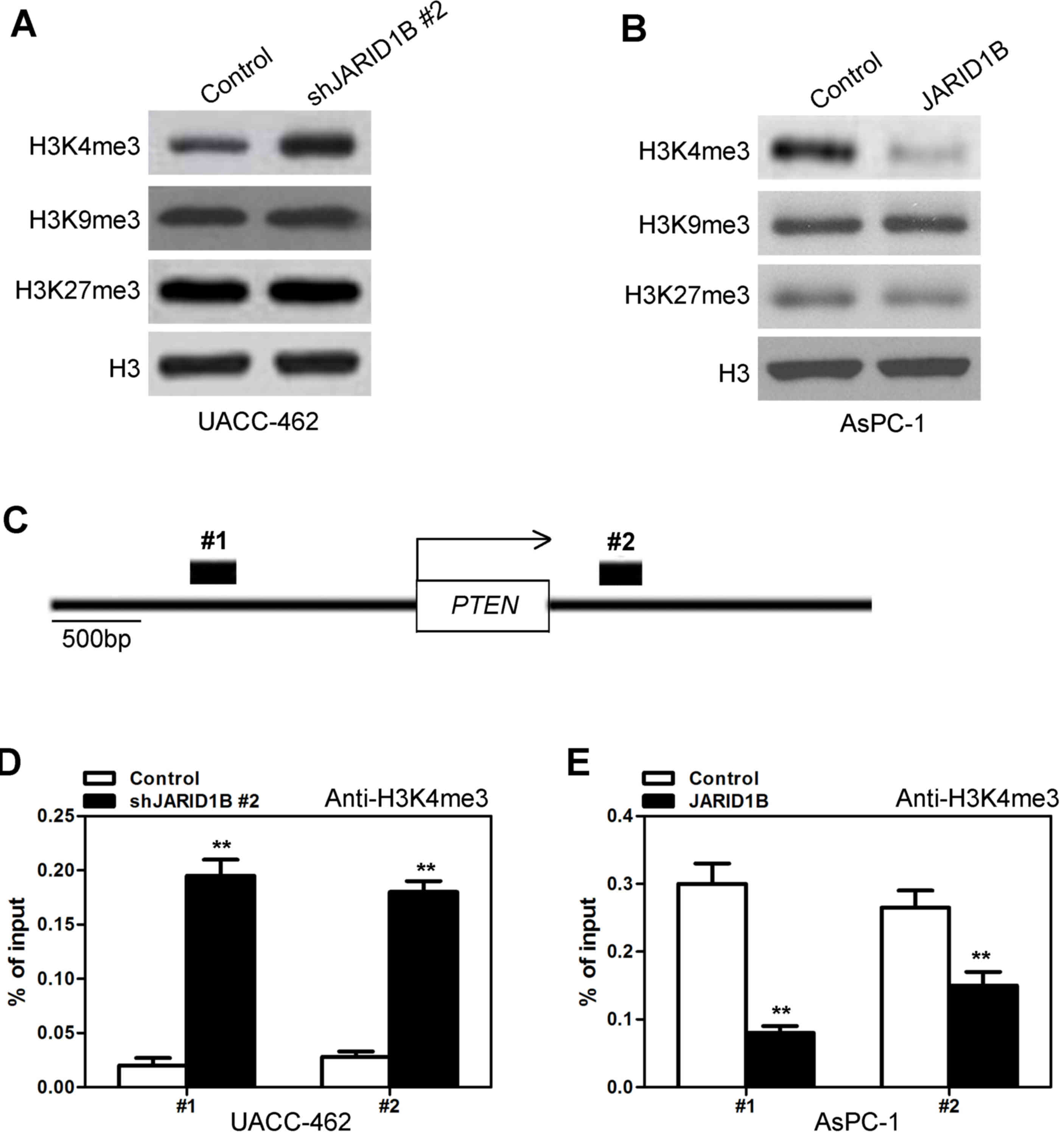
Subsequently, the manner in which JARID1B inhibited
the expression of PTEN was investigated. It has previously been
demonstrated that JARID1B regulates the methylation of H3K4
(11). To begin with, the expression
levels of H3K4me3 with silencing or overexpression of JARID1B were
investigated. Silencing of JARID1B significantly increased the
level of H3K4me3 (Fig. 8A), while
overexpression of JARID1B decreased the level of H3K4me3 (Fig. 8B). However, silencing and
overexpression of JARID1B did not affect the levels of H3K9me3 or
H3K27me3 (Fig. 8A and B). Due to the
fact that H3K4me3 is associated with the active transcription of a
number of genes, including PTEN, whether or not JARID1B
expression was associated with the H3K4me3 modification at the
promoter of PTEN in PC cells was investigated. Quantitative
chromatin immunoprecipitation (qChIP) assays were performed in
AsPC-1-pBabe-JARID1B, UACC-462-pSuper-shJARID1B and their
corresponding control cells. Silencing of JARID1B was revealed to
be associated with increased H3K4me3 levels at the region −1,235 to
−1,072 bp (Fig. 8C1) and 260 to 512
bp (Fig. 8C2) in the PTEN gene
regions (Fig. 8D). Furthermore, less
occupancy of those PTEN gene regions by H3K4me3 was detected
in AsPC-1 cells with overexpression of JARID1B (Fig. 8E). Taken together, these results
indicated that JARID1B induced inactive transcription of
PTEN due to decreased levels of H3K4me3 in the PTEN
gene.
Discussion
PC has been reported to be the most lethal malignant
neoplasm, with a difficult diagnosis and a high rate of relapse.
The results of the present study demonstrated that JARID1B was
aberrantly expressed in patients with PC and that elevated JARID1B
significantly promoted the cell and tumor proliferation of PC,
while silencing of JARID1B had the opposite effect. With regards to
the mechanism of JARID1B-promoted cell and tumor proliferation, it
was revealed that PTEN serves an essential role in these processes.
JARID1B may regulate the methylation of H3K4 in order to affect the
activation of PTEN, which ultimately promotes the progression of
PC. Therefore, JARID1B may be a novel target for the diagnosis of
PC.
In recent years, a number of novel insights into the
pathogenesis of PC have been reported. Extracellular
signal-regulated kinase promoted the degradation of FBXW7 through
phosphorylation, which resulted in the inhibition of the tumor
suppressor function of FBXW7 and eventually promoted the occurrence
of PC (30). Ataxia telangiectasia
group D complementing gene (gene-ATDC), which serves an essential
role from the early development of invasive cancer to metastatic
cancer, was also reported to be associated with 90% of the
proliferation of PC (31). Patients
with high gene-ATDC levels often experience very early metastasis,
resulting in a <30% survival rate in patients with early PC
(32). Another study reported that
>95% of the patients with PC exhibited KRAS mutations (32), while a different study revealed that
miRNA-21 serves a key role in the drug resistance of PC (33). These data may aid doctors in better
understanding the pathogenesis of PC. The present study revealed
the novel role of JARID1B in the proliferation of PC, which may
further improve the pathogenesis of PC and may serve as a novel
therapeutic target for PC.
Overexpression of JARID1B occurs in a wide variety
of cancer types, but the function of JARID1B is not fully
understood (15–22). It was recently observed that the
demethylation of H3K4 regulated by JARID1B contributed to the
silencing of retinoblastoma target genes in senescent cells,
presumably through closing the chromatin in which the silencing of
retinoblastoma trigger genes was involved (34). In esophageal cancer, JARID1B promoted
cell proliferation and tumor growth following an effect of JARID1B
on the activation of PTEN (35). The
association between JARID1B and PTEN has also been demonstrated in
previous studies (13,36,37). The
present study also revealed that JARID1B promoted the cell
proliferation and tumor growth of PC by regulating the activation
of the PTEN gene through demethylation of H3K4me3, which
further confirmed the regulatory association between JARID1B and
PTEN.
In recent years, good diagnostic markers, drug
targets and therapeutic strategies remain insufficient for the
successful treatment of PC. The results of the present study
identified the role and mechanism of JARID1B in the cell
proliferation and tumor growth of PC, which may contribute toward
the diagnosis and therapeutic strategy of PC patients.
Acknowledgements
Not applicable.
Funding
No funding received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' Contributions
PD was responsible for the conception and design of
the present study. XS, GC and PD developed the methodology. XS, GC,
LX, WW and ZC undertook the acquisition of data. XS, GC, LX, WW and
ZC were responsible for the analysis and interpretation of data. XS
and GC wrote and undertook any manuscript revisions and PD was
responsible for study supervision and any administrative,
technical, or material support.
Ethics approval and consent to
participate
The present study was conducted with the approval of
the Institutional Ethical Review Board of the Second Affiliated
Hospital of Soochow University (Suzhou, Jiangsu, China).
Consent to publish
The study participants provided their consent for
the publication of this data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hidalgo M, Cascinu S, Kleeff J, Labianca
R, Löhr JM, Neoptolemos J, Real FX, Van Laethem JL and Heinemann V:
Addressing the challenges of pancreatic cancer: Future directions
for improving outcomes. Pancreatology. 15:8–18. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vincent A, Herman J, Schulick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Duell EJ, Lucenteforte E, Olson SH, Bracci
PM, Li D, Risch HA, Silverman DT, Ji BT, Gallinger S, Holly EA, et
al: Pancreatitis and pancreatic cancer risk: A pooled analysis in
the international pancreatic cancer Case-control consortium
(PanC4). Ann Oncol. 23:2964–2970. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tumas J, Kvederaviciute K, Petrulionis M,
Kurlinkus B, Rimkus A, Sakalauskaite G, Cicenas J and Sileikis A:
Metabolomics in pancreatic cancer biomarkers research. Med Oncol.
33:1332016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Carrato A, Falcone A, Ducreux M, Valle JW,
Parnaby A, Djazouli K, Alnwick-Allu K, Hutchings A, Palaska C and
Parthenaki I: A systematic review of the burden of pancreatic
cancer in europe: Real-world impact on survival, quality of life
and costs. J Gastrointest Cancer. 46:201–211. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peto R: The fraction of cancer
attributable to lifestyle and environmental factors in the UK in
2010. Br J Cancer. 105 Suppl 2:S12011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ezzati M, Henley SJ, Lopez AD and Thun MJ:
Role of smoking in global and regional cancer epidemiology: Current
patterns and data needs. Int J Cancer. 116:963–971. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Willett WC: Diet and cancer. Oncologist.
5:393–404. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xiang Y, Zhu Z, Han G, Ye X, Xu B, Peng Z,
Ma Y, Yu Y, Lin H, Chen AP and Chen CD: JARID1B is a histone H3
lysine 4 demethylase up-regulated in prostate cancer. Proc Natl
Acad Sci USA. 104:19226–19231. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Scibetta AG, Santangelo S, Coleman J, Hall
D, Chaplin T, Copier J, Catchpole S, Burchell J and
Taylor-Papadimitriou J: Functional analysis of the transcription
repressor PLU-1/JARID1B. Mol Cell Biol. 27:7220–7235. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tang B, Qi G, Tang F, Yuan S, Wang Z,
Liang X, Li B, Yu S, Liu J, Huang Q, et al: JARID1B promotes
metastasis and epithelial-mesenchymal transition via PTEN/AKT
signaling in hepatocellular carcinoma cells. Oncotarget.
6:12723–12739. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Benevolenskaya EV: Histone H3K4
demethylases are essential in development and differentiation.
Biochem Cell Biol. 85:435–443. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Barrett A, Madsen B, Copier J, Lu PJ,
Cooper L, Scibetta AG, Burchell J and Taylor-Papadimitriou J: PLU-1
nuclear protein, which is upregulated in breast cancer, shows
restricted expression in normal human adult tissues: A new
cancer/testis antigen? Int J Cancer. 101:581–588. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lu PJ, Sundquist K, Baeckstrom D, Poulsom
R, Hanby A, Meier-Ewert S, Jones T, Mitchell M, Pitha-Rowe P,
Freemont P and Taylor-Papadimitriou J: A novel gene (PLU-1)
containing highly conserved putative DNA/chromatin binding motifs
is specifically up-regulated in breast cancer. J Biol Chem.
274:15633–15645. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cancer Genome Atlas Network: Comprehensive
molecular portraits of human breast tumours. Nature. 490:61–70.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hayami S, Yoshimatsu M, Veerakumarasivam
A, Unoki M, Iwai Y, Tsunoda T, Field HI, Kelly JD, Neal DE, Yamaue
H, et al: Overexpression of the JmjC histone demethylase KDM5B in
human carcinogenesis: Involvement in the proliferation of cancer
cells through the E2F/RB pathway. Mol Cancer. 9:592010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li X, Su Y, Pan J, Zhou Z, Song B, Xiong E
and Chen Z: Connexin 26 is down-regulated by KDM5B in the
progression of bladder cancer. Int J Mol Sci. 14:7866–7879. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shen X, Zhuang Z, Zhang Y, Chen Z, Shen L,
Pu W, Chen L and Xu Z: JARID1B modulates lung cancer cell
proliferation and invasion by regulating p53 expression. Tumour
Biol. 36:7133–7142. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ohta K, Haraguchi N, Kano Y, Kagawa Y,
Konno M, Nishikawa S, Hamabe A, Hasegawa S, Ogawa H, Fukusumi T, et
al: Depletion of JARID1B induces cellular senescence in human
colorectal cancer. Int J Oncol. 42:1212–1218. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roesch A, Vultur A, Bogeski I, Wang H,
Zimmermann KM, Speicher D, Körbel C, Laschke MW, Gimotty PA,
Philipp SE, et al: Overcoming intrinsic multidrug resistance in
melanoma by blocking the mitochondrial respiratory chain of
slow-cycling JARID1B(high) cells. Cancer Cell. 23:811–825. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Catchpole S, Spencer-Dene B, Hall D,
Santangelo S, Rosewell I, Guenatri M, Beatson R, Scibetta AG,
Burchell JM and Taylor-Papadimitriou J: PLU-1/JARID1B/KDM5B is
required for embryonic survival and contributes to cell
proliferation in the mammary gland and in ER+ breast cancer cells.
Int J Oncol. 38:1267–1277. 2011.PubMed/NCBI
|
|
24
|
Yamane K, Tateishi K, Klose RJ, Fang J,
Fabrizio LA, Erdjument-Bromage H, Taylor-Papadimitriou J, Tempst P
and Zhang Y: PLU-1 is an H3K4 demethylase involved in
transcriptional repression and breast cancer cell proliferation.
Mol Cell. 25:801–812. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wen M, Kwon Y, Wang Y, Mao JH and Wei G:
Elevated expression of UBE2T exhibits oncogenic properties in human
prostate cancer. Oncotarget. 6:25226–25239. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Y, Wen M, Kwon Y, Xu Y, Liu Y, Zhang
P, He X, Wang Q, Huang Y, Jen KY, et al: CUL4A induces
epithelial-mesenchymal transition and promotes cancer metastasis by
regulating ZEB1 expression. Cancer Res. 74:520–531. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang H, Quah SY, Dong JM, Manser E, Tang
JP and Zeng Q: PRL-3 down-regulates PTEN expression and signals
through PI3K to promote epithelial-mesenchymal transition. Cancer
Res. 67:2922–2926. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim SR, Lee KS, Park SJ, Min KH, Lee KY,
Choe YH, Lee YR, Kim JS, Hong SJ and Lee YC: PTEN down-regulates
IL-17 expression in a murine model of toluene diisocyanate-induced
airway disease. J Immunol. 179:6820–6829. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ji S, Qin Y, Shi S, Liu X, Hu H, Zhou H,
Gao J, Zhang B, Xu W, Liu J, et al: ERK kinase phosphorylates and
destabilizes the tumor suppressor FBW7 in pancreatic cancer. Cell
Res. 25:561–573. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang L, Yang H, Abel EV, Ney GM, Palmbos
PL, Bednar F, Zhang Y, Leflein J, Waghray M, Owens S, et al: ATDC
induces an invasive switch in KRAS-induced pancreatic
tumorigenesis. Genes Dev. 29:171–183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liou GY, Döppler H, Necela B, Edenfield B,
Zhang L, Dawson DW and Storz P: Mutant KRAS-induced expression of
ICAM-1 in pancreatic acinar cells causes attraction of macrophages
to expedite the formation of precancerous lesions. Cancer Discov.
5:52–63. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ma Y, Wu Q, Li X, Gu X, Xu J and Yang J:
Pancreatic cancer: From bench to bedside. Ann Transl Med.
4:4582016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chicas A, Kapoor A, Wang X, Aksoy O,
Evertts AG, Zhang MQ, Garcia BA, Bernstein E and Lowe SW: H3K4
demethylation by Jarid1a and Jarid1b contributes to
retinoblastoma-mediated gene silencing during cellular senescence.
Proc Natl Acad Sci USA. 109:8971–8976. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kano Y, Konno M, Ohta K, Haraguchi N,
Nishikawa S, Kagawa Y, Hamabe A, Hasegawa S, Ogawa H, Fukusumi T,
et al: Jumonji/Arid1b (Jarid1b) protein modulates human esophageal
cancer cell growth. Mol Clin Oncol. 1:753–757. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lu W, Liu S, Li B, Xie Y, Adhiambo C, Yang
Q, Ballard BR, Nakayama KI, Matusik RJ and Chen Z: SKP2
inactivation suppresses prostate tumorigenesis by mediating JARID1B
ubiquitination. Oncotarget. 6:771–788. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yuan P, Ito K, Perez-Lorenzo R, Del Guzzo
C, Lee JH, Shen CH, Bosenberg MW, McMahon M, Cantley LC and Zheng
B: Phenformin enhances the therapeutic benefit of BRAF(V600E)
inhibition in melanoma. Proc Natl Acad Sci USA. 110:18226–18231.
2013. View Article : Google Scholar : PubMed/NCBI
|