Introduction
The term cancer refers to a group of diseases during
the course of which an increase in the number of cells with altered
genotype and phenotype is observed. The characteristic feature of
cancer cells is primarily their uncontrolled and unrestrained
proliferation, as well as their autonomy, which underlies the
carcinogenesis process. Due to the many changes on a molecular
basis (mutation among others), an increase in cancer aggressiveness
is a very complex and long-lasting process that usually proceeds
asymptomatically. A changed sensitivity to a number of growth
factors (cytokines, chemokines and others) is responsible for the
unlimited number of cell divisions, the apoptosis impairment, the
uncontrolled proliferation and the masking and defence against the
host's immune system. One of the final stages in cancer progression
is the formation of cardiovascular and lymphatic vessels (angio-
and lymphangiogenesis, respectively), thanks to which the tumor
grows in volume and acquires the ability to metastasize.
Due to the complexity of the cancer progression
process and, in the case of many types or cancer, the lack of
effective therapy regimens, new molecular markers which could be
potential targets for anti-cancer therapy are increasingly sought
after. The proteins of the SRY-related HMG-box (SOX) family seem to
be a promising element in cancer therapy in this aspect (1). The family of SOX proteins was isolated
and selected in 1990 on the basis of the presence of the high
mobility group box (HMG) conservative protein domain, initially
characteristic for the sex-determining factor found on the Y
chromosome [SRY (sex determining region Y)] (1). Typical for SOX family proteins, the HMG
domain (which consists of 79 amino acids) is characterized by a 50%
homology to that occurring in the SRY gene. The SOX proteins, which
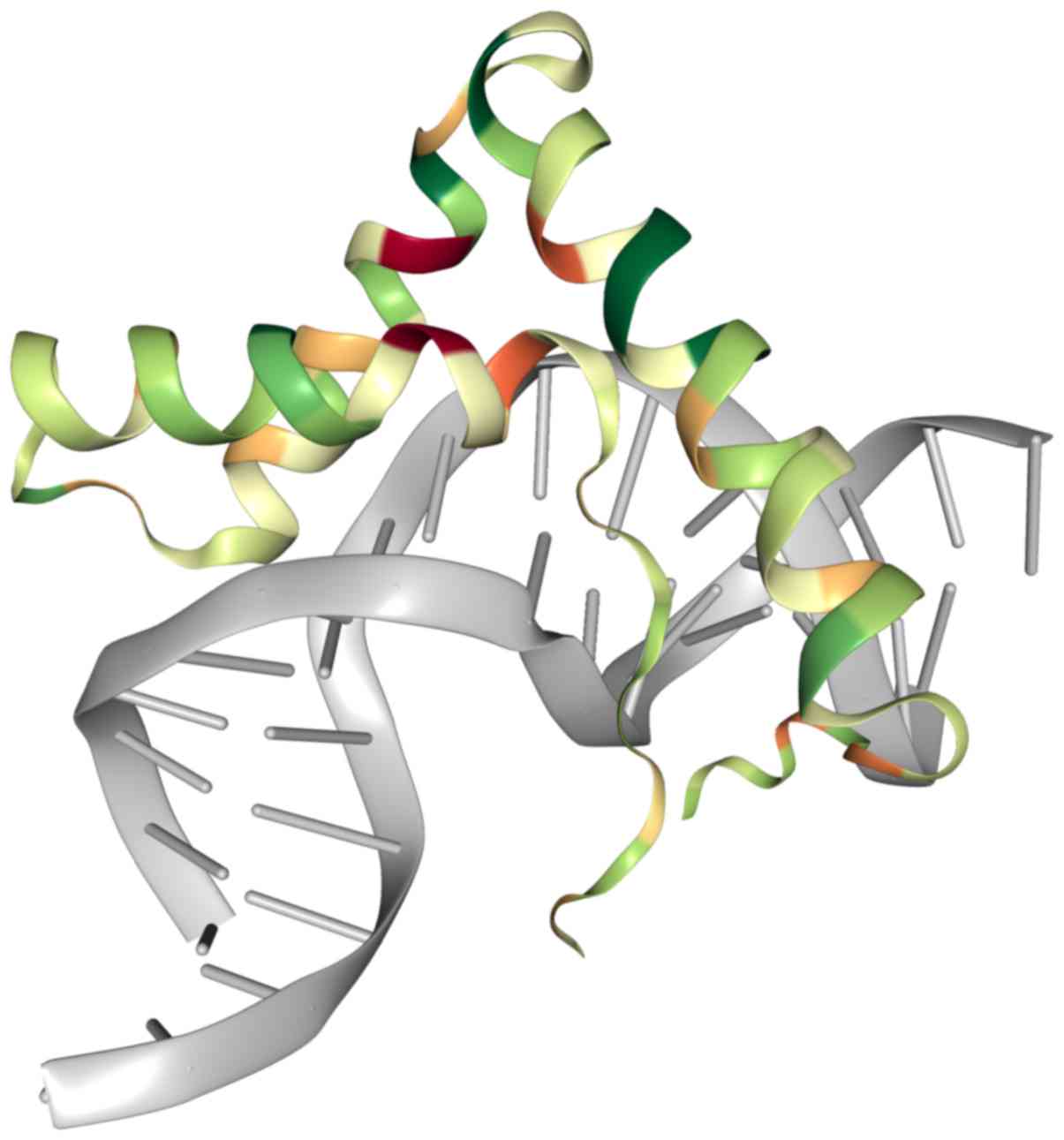
are transcription factors, bind to the 5′-(A/T)(A/T)CAA(A/T)G-3′
sequence precisely through the HMG domain, thus allowing other
transcription factors to bind (Fig.
1) (2,3). The SOX family is comprised of
approximately 20 proteins, divided into 8 main groups (from A to H)
due to their homology and the functions they perform in the body
(4,5).
The F group includes the SOX7, SOX17 and SOX18 proteins, which are
involved in the same pathways as the vascular endothelial growth
factor (VEGF) (6–9).
The SOX18 protein is one of the most important
transcription factors involved in the development of blood and
lymphatic vessels during the embryogenesis process. Likewise,
studies in humans have shown that mutations in the SOX18
gene may result in serious developmental disorders. SOX18 mutation
is responsible for the emergence of the
hypotrichosislymphedematelangiectasia (HLT) syndrome (10,11). This
disease is characterized by the simultaneous appearance of
abnormalities in hair development, the presence of lymphedema and
the widening of small blood vessels. Recent research has also shown
its participation in the pathogenesis of many cancers (12–15). Since
different levels of this protein have been observed in different
types of tumors, it seems that it can perform various functions in
cancer cells (16).
SOX protein family
The SRY protein, coded by the SRY
gene-located in the Y chromosome and conditioning sex (17), belongs to the group A of SOX proteins.
Group B, which is expressed primarily in the central nervous
system, is divided into two subgroups: B1, which consists mainly of
activators (SOX1, SOX2 and SOX3), and B2, which contains mainly
suppressors of the transcription process (SOX4 and SOX21).
Representatives of group C are the proteins SOX11 and SOX12, whose
function has not yet been fully investigated and described.
However, an increase in the expression level of the genes
SOX4 and SOX21 in non-small cell lung carcinomas has
been demonstrated (18,19). The proteins SOX5, SOX6 and SOX13,
which belong to group D, are involved in the processes of
spermatogenesis and chondrogenesis (5,20). SOX8,
SOX9 and SOX10, representatives of group E, are involved in the
development of the nervous system and the testicular nuclei
(21–23). Group F includes the previously
mentioned SOX7, SOX17 and SOX18 proteins. The SOX15 protein is the
only representative of group G, and it is involved in the
development of the testis, the placenta and the muscular system
during the process of embryogenesis (24). The last group of transcription factors
from the SOX family is the group H, a representative of which is
the SOX30 protein, involved mainly in the process of
spermatogenesis (25). Additionally,
it has recently been showed that SOX30 has a suppressor function in
adenocarcinoma (26).
Group F of SOX proteins
The representatives of the group F of SOX proteins
are the proteins SOX7, SOX17 and SOX18, jointly involved in the
development of the cardiovascular and lymphatic systems during the
embryogenesis process (27,28). Since the main characteristic of these
proteins is their high homology, they are able to take over each
other's functions, as demonstrated in a mouse model where the
inhibition of the SOX18 gene expression resulted in an
increased expression of the SOX7 and SOX17 gene
(29).
It has also been proven that, during the embryonic
development, SOX18 interacts with the myocyte enhancer factor-2C
(MEF2C) transcription factor in the vascular endothelial cells. It
has been demonstrated in vivo in a mouse model that the
simultaneous silencing of the SOX18 and MEF2C genes
results in serious cardiovascular malformations, which clearly
confirms the key role of these transcription factors in the initial
stages of the development of blood vessels (30).
An important characteristic of the SOX7, SOX17 and
SOX18 proteins is the presence of the amino acid sequence
DXXEFD/EQYL, responsible for the interaction with the β-catenin in
the WNT/β-catenin pathway (30,31). This
pathway is involved in several key physiological processes at every
stage of the development of the body, and its disorders lead to the
initiation of the carcinogenesis process. Sinner et al have
shown that SOX17 acts as an antagonist to the WNT/β-catenin pathway
by being able to bind simultaneously with the β-catenin and the
t-cell factor (TCF) protein. In vitro studies performed on
the SW480 colorectal cancer line model demonstrated that the
overexpression of the SOX17 gene inhibits the activity of
the β-catenin/TCF complex, thereby reducing the level of
proliferation of the cells of this cancer (32–34).
In turn, in the case of the SOX7 protein, it has
been shown that the downregulation of the expression of its gene
resulted in the inhibition of the WNT/β-catenin pathway in
endometrial cancer cells, which correlated positively with a higher
malignancy degree of the tumor (33–35).
The role of the proteins of the SOX family,
including proteins from the SOX F group, in the development of
cancer has been the focus of many research teams around the world
for many years. It is worth noting that, despite the high homology
of the SOX7, SOX17 and SOX18 proteins, they can act both as
stimulators and inhibitors in the carcinogenesis process (17).
SOX18 protein
In humans, the gene encoding the SOX18 protein is
found in the long arm of chromosome 20 in the locus 20q13.3, and it
consists of 8,901 base pairs. The product of the SOX18 gene is a
protein made of 384 amino acids with a mass of 41 kDa (8). In the embryonic development, this
protein is responsible for the activation of the Prospero Homeobox
1 (Prox-1) promoter and, thus, it is involved in the formation of
new blood and lymphatic vessels (28). The initiation of the expression of the
SOX18 gene and, therefore, of PROX1, in the early
stages of embryogenesis, results in changes in the phenotype of
endothelial cells, which causes their migration and fusion (which
leads to the formation of a network of lymphatic vessels, the
protoplasts of the mature lymphatic system) (28). Among other discoveries, it has been
found that in mouse embryos with a silenced SOX18 gene
expression, the lymphangiogenesis process is inhibited (28).
In addition to the involvement of the SOX18 protein
in the formation of the cardiovascular system in the embryonic
development, its presence has also been demonstrated in many cells
and tissues of the mature human body, for example in the heart, the
lungs, the skeletal muscles, the stomach and the intestines
(8,36). Moreover, an increase in the level of
this protein has been observed in pathological states such as
coronary artery atherosclerosis and would healing (37,38).
Additionally, it has been demonstrated that the
SOX18 transcription factor is able to form not only functional
homodimeric complexes, but also heterodimeric ones with MEF2C.
Furthermore, SOX7 is also capable of creating heterodimers with the
Recombining Binding Protein Suppressor of Hairless (RBPJ) protein,
and SOX17 with Octamer-binding transcription factor 4 (OCT4)
(39). The formation of these
complexes allows the transcription factors of the SOX family to
bind to distant DNA regions, thereby activating or repressing a
larger gene pool.
The observed variable levels of SOX18 in different
kinds of cancer (stomach, colorectal, ovarian, endometrium or liver
cancer) indicate the role of this transcription factor in tumor
progression, metastasis and the development of blood and lymphatic
vessels within the tumor.
Breast cancer
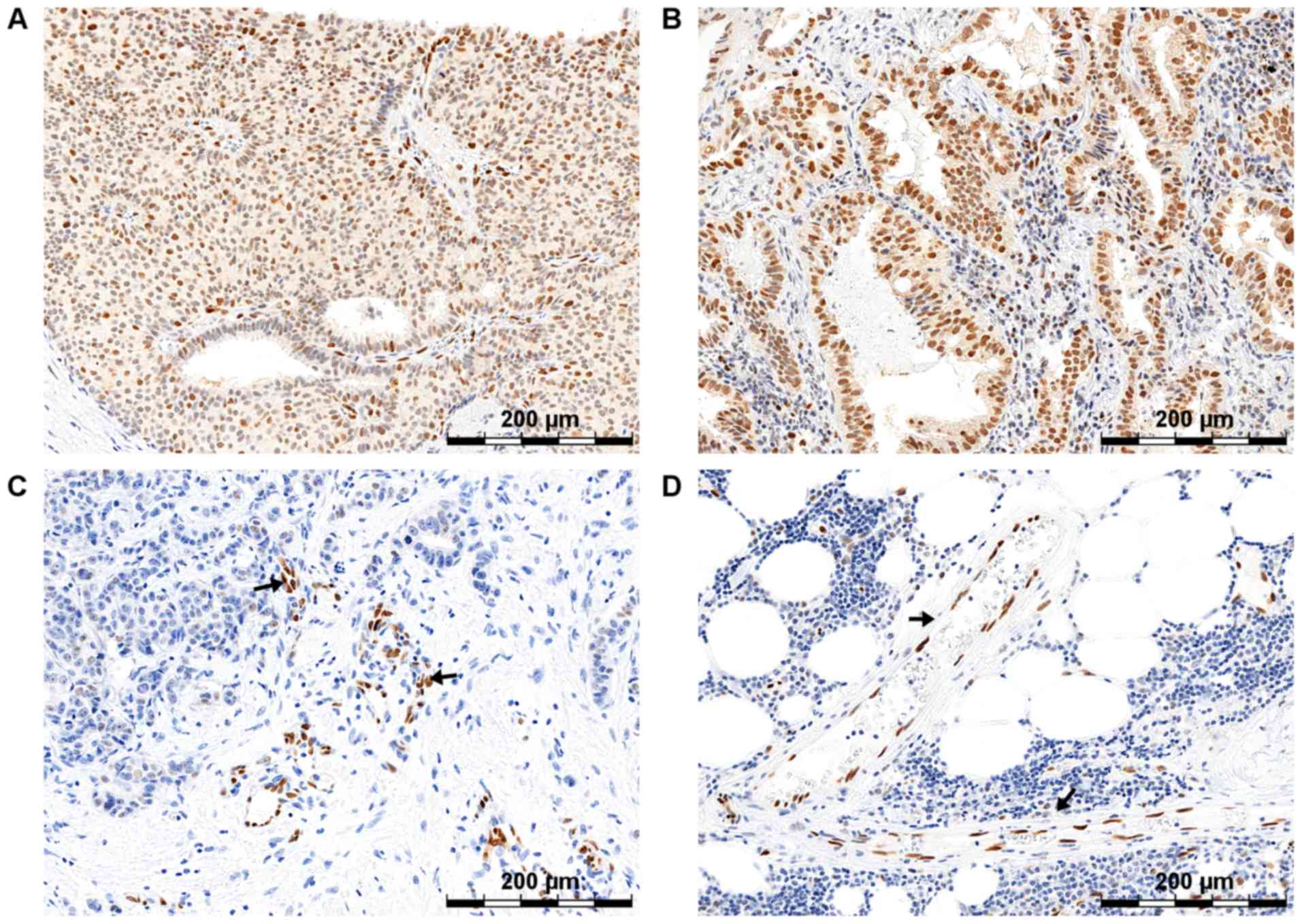
The results of the tests by Pula et al,
conducted using the immunohistochemical (IHC) technique on clinical
material from 122 cases of Invasive Ductal Breast Carcinoma (IDC),
indicated the existence of a significant positive correlation
between an increase in the SOX18 protein level and an increase in
the tumor malignancy grade (Fig. 2)
(14). The expression of the SOX18
factor in the nuclei of the vascular endothelial cells of the tumor
was confirmed, suggesting the participation of SOX18 in the
angiogenesis process. Interestingly, the results of the in
vitro tests on breast cancer human cell lines with increasing
aggressiveness (MCF-7, BT-474, SK-BR-3, MDA-MB-231 i
MDA-MB-231/BO2), on normal human dermal fibroblasts (MHDF) and on
the normal human breast cell line HME1-hTERT confirmed different
levels of the SOX18 protein in the breast cancer cell lines and a
lack of SOX18 expression in the NHDF and the HME1-hTERT control
lines. By using the laser microdissection technique, the authors
clearly demonstrated the existence of an increased expression of
the SOX18 gene and the SOX7 and SOX17 genes in the
vascular endothelial cells, thus confirming the role of these
proteins in the angiogenesis process. Additionally, the level of
SOX18 mRNA correlated with the mRNA level of the gene
VEGFD, which indicates the role of this protein in the
lymphangiogenesis process. Moreover, it was noted that the breast
cancer cell lines not expressing the estrogen receptor (ER-), the
progesterone receptor (PR-) and the human epidermal growth factor
receptor type 2 (HER2-), i.e., the triple negative, were
characterized by a negligible expression of SOX18 (14). This may indicate the involvement of
this transcription factor in the Wnt/β-catenin pathway, responsible
for IDC progression, since similar observations made by other
research teams had already indicated the contribution of the SOX18
protein to the regulation of TCF transcription factors (33).
In vitro studies by Young et al showed
that the inhibition of the expression of the SOX18 transcription
factor in the MCF-7 line results in a decrease in the capacity of
these cells to migrate because of the destabilization of the actin
cytoskeleton (40). As a consequence,
it was claimed that SOX18 inhibition caused a significant reduction
in the size of induced tumors, as well as a decrease in blood
vessel density within the tumor. These observations confirmed the
participation of the SOX18 protein in the angiogenesis process.
Since the number of blood vessels in the tumor closely correlates
with the risk of metastases and shorter survival times, silencing
the expression of the SOX18 gene (which leads to the
reduction of the blood vessel network within the tumor) may
contribute to sensitizing cancer cells to chemotherapeutic agents
by reducing the supply of nutrients supplied along with blood
through the blood vessels. Additionally, an increase in the level
of the SOX18 expression in the MCF-7 line resulted in an increased
proliferation and the creation of capillaries from the Human
Umbilical Vein Endothelial Cells (HUVEC), ultimately confirming the
contribution of the SOX18 protein to the progression of invasive
breast cancer (40).
Ovarian and cervical cancer
Due to the fact that SOX18 is expressed not only by
vascular endothelial cells but also by tumor cells themselves,
Petrovic et al decided to investigate the role of this
transcription factor in cervix cancers (41). So far, the involvement of the SOX18
protein has been confirmed in the Hedgehog pathway in non-cancerous
human cells, primarily during embryonic development. Since the
Hedgehog pathway remains mainly inactive in the mature cells of the
body and its activation is observed in tumors (including cervix
cancer), the modulation of the inhibitors of this pathway may be
important in future targeted therapy. Petrovic et al
demonstrated that the SOX18 protein is regulated by the GL1 and GL2
(glioma-associated oncogene-1, −2) transcription factors, which are
modulators of the Hedgehog pathway (39). It was proved that the inhibitors GL1
and GL2, involved in the regulation of the aforementioned pathway,
are also responsible for the reduction of the expression level of
the SOX18 gene. Moreover, the in vitro tests on the
human cervix cancer cell lines HeLa, SiHa and CaSki showed that a
SOX18 overexpression did not increase the proliferative potential,
but significantly increased the invasiveness and migration of the
cells of this cancer (41). This
could suggest a contribution of the SOX18 protein to the promotion
of tumor progression in cervix cancers.
Other studies carried out on cervix cancer
postoperative material were designed to demonstrate the potential
of the SOX18 protein as a predictive marker in the chemotherapy of
this type of cancer. Pula et al showed that high levels of
SOX18 in ovarian cancer cell nuclei significantly correlated with a
shorter disease-specific survival (DSS) and a shorter disease-free
survival (DFS) (15). Additionally,
the in vitro tests on the human ovarian cancer cell lines
SK-OV-3 and OVCAR3 and the ovarian normal surface epithelial cell
line IOSE 364 indicated the possibility of the participation of the
SOX18 protein in the mechanism of tumor cell resistance to
chemotherapeutics, since the SK-OV-3 and OVCAR3 cells,
characterized by a high drug resistance (including cisplatin),
showed high levels of the SOX18 protein.
Stomach and liver cancer
Research by Jia et al (42) was carried out on material from 196
cases of stomach cancer and the 6 human stomach cancer cell lines
SGC7901, MGC803, BGC823, AGS, N87 and MKN45. In these experiments,
the authors determined the level of methylation of the naked
cuticle homolog (NKD) -1 and -2 gene
promoters, SOX18 direct inhibitors, on the aforementioned material.
It turned out that the methylation of the NKD1 gene is
influenced, among others, by the ability of the cancer cells to
migrate and create metastases (42).
In turn, the active NKD2 protein effectively inhibits cell
proliferation and colony formation, and it also stops the cell
cycle at the G2/M control point. Consecutively, the reactivation of
the NKD2 gene expression sensitized the stomach cancer cells
to docetaxel. The analysis of the expression level of over 200
genes by using the expression microarray technique indicated the
existence of a molecular mechanism of inhibition of the expression
of the SOX18 transcription factor via the NKD2 protein.
SOX18 expression significantly correlated positively with
the methylation profile of the NKD2 gene promoter. It was
also shown that SOX18 directly affects the levels of the
metalloproteinases MMP-2, MMP-7 and MMP-9 (42). Due to the fact that the methylation of
the NKD2 gene promoter entails a reduced expression of the
SOX18 factor (which ultimately results in a decrease in the
metalloproteinase level), SOX18 was found to play a key role in the
progression and metastasis of stomach cancer cells.
In tests on material from 75 cases of primary liver
cancer and the hepatocellular carcinoma (HCC) human cell lines
HepG2, BEL-7404, SMC-7721, MHCC-97L and MHCC-97H, Wang et al
(43) tried to determine the role of
the SOX18 transcription factor in the pathogenesis of this cancer.
A significantly higher level of SOX18 in cases of liver cancer was
observed compared to cases of normal liver tissue from the area
surrounding the tumor, both at the mRNA and protein levels. This
high level of SOX18 correlated with a shorter DFS period. The in
vitro tests carried out in order to inhibit the SOX18
expression clearly showed that a decrease in the level of the SOX18
protein resulted in a cell cycle arrest at the G1 control point and
the initiation of apoptosis in liver cancer cells (43). SOX18-siRNA significantly reduced the
ability of the HCC cells to migrate and invade. The authors were
also able to show the contribution of the SOX18 protein in tumor
progression through direct interaction with the proteins Ras
homolog family member A (RhoA), insulin-like growth factor 1
receptor (IGF1R), platelet derived growth factor Subunit B (PDGFB),
C-C Motif Chemokine Ligand (CCL) 2, CCL3 and CCL5, which are
responsible, among others, for focal adhesion and signalling in the
chemokine signalling pathway (43).
Non-small cell lung cancer
Lung malignant tumors arise as a result of the
accumulation of damage to the genetic material of the cells, which
ultimately leads to the process of tumor transformation (44).
Numerous studies have shown that in NSCLCs mutations
appear most commonly in genes belonging to pathways which are
related to, among others, the activation of the epidermal growth
factor receptor protein (EGFR), i.e., RAS/RAF/MEK/ERK and PI3K,
AKT/mTOR (45). The result of this is
an enhanced proliferation of cancer cells and their ability to form
metastasis, as well as the intensification of the angiogenesis
processes within the tumor. EGFR membrane expression (from the
tyrosine kinase family) is relatively frequent in NSCLC cells, in
contrast to small-cell lung cancer and unchanged tissue. EGFR
overexpression is observed in 40–80% of NSCLC cases, including
24–89% of SQCs and 23–46% of ACs (46).
There are epigenetic mechanisms that are responsible
for the gene and protein expression profile specific for a
determined tissue and regulate it with precision. The basic
epigenetic mechanisms include DNA methylation, post-translational
modifications of proteins and miRNA or siRNA participation
(47). These mechanisms often seem to
have a different course in cancer cells than in normal human cells
(48). It has been recently proven
that in cases of non-small cell lung cancer (NSCLC) there is a high
heterogeneity of the methylation process (49,50). For
example, non-malignant lung tissue (NMLT) is characterized by a
high level of methylation of the SOX18 gene promoter. In turn, the
closer to the tumor area, the lower this level is. Therefore, it
should be assumed that also in the tumor itself, depending on the
area being studied, the level of methylation varies and can affect
the neoplastic process (51).
Azhikina et al (51), in lung
cancer post-operative material and cases of NMLT, determined the
methylation profiles of four gene promoters: Tissue inhibitor of
metalloproteinases 4 (TIMP4), GATA binding protein 4
(GATA4), multiple epidermal growth factor-like domains
protein 7 (EGFL7) and SOX18, whose role in the
neoplastic process was confirmed and documented. They demonstrated
the existence of the heterogeneity of the methylation level of the
SOX18 gene promoter both in NSCLC and NMLT cases, and
pointed out the possibility of using the methylation profile of the
SOX18 gene promoter in NSCLC diagnosis in the future
(51).
The results so far seem to confirm the complexity of
the epigenetic mechanisms in NSCLC (9,16). Based
on the results obtained from our research, different SOX18
expressions were observed in cases of NSCLC both at the mRNA and
protein levels (Fig. 2) (9,16).
Interestingly, the level of mRNA did not in any way reflect the
level of protein, determined with the use of the western blot
technique (16). Later experiments
revealed the participation of two miRNA molecules in this
phenomenon: miR-7a and miR-24-3p (9).
The results of the analysis of the level of these molecules in
cases of lung adenocarcinoma (AC) and squamous-cell lung carcinoma
(SQC) and their corresponding cases of NMLT point to a mechanism
carried out by the cancer cells through which they reduce the
primary high level of the miRNA molecules found in normal lung
tissue (9,52). Therefore, the demonstration of the
role of specific miRNAs and the role of the methylation of the
SOX18 gene promoter in the above-described mechanism may be
used in the future in diagnostics, prognostic assessment and,
possibly, in NSCLC targeted therapy as well.
The prospect of using miRNA interacting with the
SOX18 gene in the treatment of non-small cell lung
carcinomas seems to be promising, since there are already many
similar miRNA molecules with proven pro- or anti-cancer functions
(53,54). Variable miRNA molecule levels can be
detected in the patient's blood, which further increases the
availability and universality of their use in lung cancer
diagnosis. Besides, miRNA examination can provide information not
only about the type, but also about the stage of the cancer.
Skin lymphomas
Jankowska et al (55) attempted to analyse the level of
expression of the SOX18 transcription factor in 80 cases of mycosis
fungoides (MF). A positive correlation was found between the
increase in the SOX18 level in MF cases and the progression of the
neoplastic disease. In contrast to previous publications in which
SOX18 expression was observed both in tumor cells and in the nuclei
of vascular endothelial cells, the results in these studies
indicated the presence of SOX18 expression only in the vascular
endothelial cells (54). Hence, the
authors postulate that in this type of cancer the role of SOX18 is
limited to the process of angiogenesis itself, which is key in
cancer progression, allowing the tumor to nourish, grow and
metastasize. This observation is supported by the fact that a
statistically higher level of SOX18 was observed in the blood
vessels in the cases of MF compared to neoplasms like lichen
planus and eczema. In addition, the high levels of SOX18
correlated positively with the degree of histological
differentiation of the MF, which confirms the role of this
transcription factor in the process of tumor progression (55). The differences between the mRNA and
the SOX18 protein levels in MF cases may indicate, as it happens in
other types of cancers, the complexity of the modulation process of
the transcript of this gene through the participation of, for
example, epigenetic factors. The observation of the existence of a
positive correlation between the SOX18 expression and the
expression of known proliferation markers such as antigen
identified by monoclonal antibody Ki-67 (Ki-67),
Minichromosome Maintenance Complex (MCM)-3, −7, suggests
that the SOX18 protein could be used in the future as a potential
marker of tumor aggressiveness.
Osteosarcoma
An increased expression of the SOX18 transcription
factor was observed in osteosarcoma cells, a fact that may be of
great importance in the future in the diagnosis and therapy of this
type of malignant tumor (56). In the
in vitro and in vivo tests proposed by Wu et
al, the SOX18 expression was effectively silenced in the human
osteosarcoma cell lines U2OS and MG63, which resulted in a decrease
in the cancer cells' ability to form metastases and inhibited tumor
growth, thus confirming the role of SOX18 in tumor cell
proliferation. Moreover, it was noted that the inhibition of the
SOX18 expression results in a cell cycle arrest in the S phase, and
thus results in tumor cell apoptosis (56). Furthermore, there was a reduction of
the osteosarcoma cell adhesion and migration capacity. The
mechanism that underlies the observed phenomena is still unclear,
although the authors hypothesize that SOX18 could be involved in
the pathway of the activation of the proteins platelet-derived
growth factor A (PDGFA), PDGFB, RhoA and transforming growth
factor-β1 (TGF-β1), which are responsible for cancer cell
progression and invasion. As previously described, a similar
mechanism was also observed in liver cancer cells (43). The results of the studies presented
above seem to attribute a suppressor role to the SOX18
transcription factor in osteosarcoma cells.
Conclusions
It has been found that the SOX18 protein may be
expressed differently in some types of human malignant cancers,
although the mechanisms underlying the biological basis of this
process are still not fully understood. The SOX proteins are
involved in the same pathways as the VEGFs, which indicates their
participation in the development of the cardiovascular system and
in lymphangiogenesis (28). The
proteins from this group exhibit a fairly high homology, and
therefore they are able to take over each other's functions in
specific situations. It has been demonstrated, in the absence of
the SOX18 protein, its function, which is the development of the
lymphatic vessels in mice, its fulfilled by the SOX7 and SOX17
proteins (57).
An in-depth study of the epigenetic mechanisms
conditioning these phenomena may allow a more comprehensive
understanding of the molecular basis of cancer development, which
in turn may contribute to the implementation of new targeted
therapy based ultimately on proteins from the SOX family (a
representative of which is SOX18). Great efforts have been recently
made in order to fully understand the multifactorial and multistage
process of carcinogenesis. Since epigenetic changes are also at the
basis of the cancer process, the discovery of new markers of
susceptibility to cancer based, among others, on miRNA-based
regulation can enable the introduction into clinical practice of
new diagnostic and predictive tests, and perhaps also of molecular
therapies.
Considering the role of the proteins of the SOX
family in the progression of many types of human malignant tumors
and their role in the proliferation and migration processes and in
cell apoptosis, it is reasonable to test, identify and understand
the mechanisms in which the SOX transcription factors (including
the SOX18 protein) play a key role.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
MO was involved in the preparation of the
manuscript, gathering of the necessary publications and figure
preparation. PD and MPO contributed to revising the manuscript and
language.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gubbay J, Collignon J, Koopman P, Capel B,
Economou A, Münsterberg A, Vivian N, Goodfellow P and Lovell-Badge
R: A gene mapping to the sex-determining region of the mouse Y
chromosome is a member of a novel family of embryonically expressed
genes. Nature. 346:245–250. 1990. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Harley VR, Lovell-Badge R and Goodfellow
PN: Definition of a consensus DNA binding site for SRY. Nucleic
Acids Res. 22:1500–1501. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lovell-Badge R: The early history of the
Sox genes. Int J Biochem Cell Biol. 42:378–380. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wegner M: From head to toes: The multiple
facets of Sox proteins. Nucleic Acids Res. 27:1409–1420. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bowles J, Schepers G and Koopman P:
Phylogeny of the SOX family of developmental transcription factors
based on sequence and structural indicators. Dev Biol. 227:239–255.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Taniguchi K, Hiraoka Y, Ogawa M, Sakai Y,
Kido S and Aiso S: Isolation and characterization of a mouse
SRY-related cDNA, mSox7. Biochim Biophys Acta. 1445:225–231. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kanai Y, Kanai-Azuma M, Noce T, Saido TC,
Shiroishi T, Hayashi Y and Yazaki K: Identification of two Sox17
messenger RNA isoforms, with and without the high mobility group
box region, and their differential expression in mouse
spermatogenesis. J Cell Biol. 133:667–681. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dunn TL, Mynett-Johnson L, Wright EM,
Hosking BM, Koopman PA and Muscat GE: Sequence and expression of
Sox-18 encoding a new HMG-box transcription factor. Gene.
161:223–225. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Olbromski M, Grzegrzolka J,
Jankowska-Konsur A, Witkiewicz W, Podhorska-Okolow M and Dziegiel
P: MicroRNAs modulate the expression of the SOX18 transcript in
lung squamous cell carcinoma. Oncol Rep. 36:2884–2892. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Irrthum A, Devriendt K, Chitayat D,
Matthijs G, Glade C, Steijlen PM, Fryns JP, van Steensel MA and
Vikkula M: Mutations in the transcription factor gene SOX18
underlie recessive and dominant forms of
hypotrichosis-lymphedema-telangiectasia. Am J Hum Genet.
72:1470–1478. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cermenati S, Moleri S, Cimbro S, Corti P,
Del Giacco L, Amodeo R, Dejana E, Koopman P, Cotelli F and Beltrame
M: Sox18 and Sox7 play redundant roles in vascular development.
Blood. 111:2657–2666. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Castillo SD and Sanchez-Cespedes M: The
SOX family of genes in cancer development: Biological relevance and
opportunities for therapy. Expert Opin Ther Targets. 16:903–919.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pula B, Werynska B, Olbromski M,
Muszczynska-Bernhard B, Chabowski M, Janczak D, Zabel M,
Podhorska-Okolow M and Dziegiel P: Expression of Nogo isoforms and
Nogo-B receptor (NgBR) in non-small cell lung carcinomas.
Anticancer Res. 34:4059–4068. 2014.PubMed/NCBI
|
|
14
|
Pula B, Olbromski M, Wojnar A,
Gomulkiewicz A, Witkiewicz W, Ugorski M, Dziegiel P and
Podhorska-Okolow M: Impact of SOX18 expression in cancer cells and
vessels on the outcome of invasive ductal breast carcinoma. Cell
Oncol (Dordr). 36:469–483. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pula B, Kobierzycki C, Solinski D,
Olbromski M, Nowak-Markwitz E, Spaczynski M, Kedzia W, Zabel M and
Dziegiel P: SOX18 expression predicts response to platinum-based
chemotherapy in ovarian cancer. Anticancer Res. 34:4029–4037.
2014.PubMed/NCBI
|
|
16
|
Jethon A, Pula B, Olbromski M, Werynska B,
Muszczynska-Bernhard B, Witkiewicz W, Dziegiel P and
Podhorska-Okolow M: Prognostic significance of SOX18 expression in
non-small cell lung cancer. Int J Oncol. 46:123–132. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu Y, Li Y, Jun Wei JW and Liu X: The
role of Sox genes in lung morphogenesis and cancer. Int J Mol Sci.
13:15767–15783. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sekido R: A transcriptional activator of
mammalian testis determination. Int J Biochem Cell Biol.
42:417–420. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Medina PP, Castillo SD, Blanco S,
Sanz-Garcia M, Largo C, Alvarez S, Yokota J, Gonzalez-Neira A,
Benitez J, Clevers HC, et al: The SRY-HMG box gene, SOX4, is a
target of gene amplification at chromosome 6p in lung cancer. Hum
Mol Genet. 18:1343–1352. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Penzo-Méndez AI: Critical roles for SoxC
transcription factors in development and cancer. Int J Biochem Cell
Biol. 42:425–428. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lefebvre V: The SoxD transcription
factors-Sox5, Sox6, and Sox13-are key cell fate modulators. Int J
Biochem Cell Biol. 42:429–432. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Haldin CE and LaBonne C: SoxE factors as
multifunctional neural crest regulatory factors. Int J Biochem Cell
Biol. 42:441–444. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Stolt CC and Wegner M: SoxE function in
vertebrate nervous system development. Int J Biochem Cell Biol.
42:437–440. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Barrionuevo F and Scherer G: SOX E genes:
SOX9 and SOX8 in mammalian testis development. Int J Biochem Cell
Biol. 42:433–436. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ito M: Function and molecular evolution of
mammalian Sox15, a singleton in the SoxG group of transcription
factors. Int J Biochem Cell Biol. 42:449–452. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Han F, Wang Z, Wu F, Liu Z, Huang B and
Wang D: Characterization, phylogeny, alternative splicing and
expression of Sox30 gene. BMC Mol Biol. 11:982010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Han F, Liu W, Jiang X, Shi X, Yin L, Ao L,
Cui Z, Li Y, Huang C, Cao J and Liu J: SOX30, a novel epigenetic
silenced tumor suppressor, promotes tumor cell apoptosis by
transcriptional activating p53 in lung cancer. Oncogene.
34:4391–4402. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
François M, Caprini A, Hosking B, Orsenigo
F, Wilhelm D, Browne C, Paavonen K, Karnezis T, Shayan R, Downes M,
et al: Sox18 induces development of the lymphatic vasculature in
mice. Nature. 456:643–647. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sakamoto Y, Hara K, Kanai-Azuma M, Matsui
T, Miura Y, Tsunekawa N, Kurohmaru M, Saijoh Y, Koopman P and Kanai
Y: Redundant roles of Sox17 and Sox18 in early cardiovascular
development of mouse embryos. Biochem Biophys Res Commun.
360:539–544. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bernard P and Harley VR: Acquisition of
SOX transcription factor specificity through protein-protein
interaction, modulation of Wnt signalling and post-translational
modification. Int J Biochem Cell Biol. 42:400–410. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hosking BM, Wang SC, Chen SL, Penning S,
Koopman P and Muscat GE: SOX18 directly interacts with MEF2C in
endothelial cells. Biochem Biophys Res Commun. 287:493–500. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sinner D, Rankin S, Lee M and Zorn AM:
Sox17 and beta-catenin cooperate to regulate the transcription of
endodermal genes. Development. 131:3069–3080. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kormish JD, Sinner D and Zorn AM:
Interactions between SOX factors and Wnt/beta-catenin signaling in
development and disease. Dev Dyn. 239:56–68. 2010.PubMed/NCBI
|
|
34
|
Sinner D, Kordich JJ, Spence JR, Opoka R,
Rankin S, Lin SC, Jonatan D, Zorn AM and Wells JM: Sox17 and Sox4
differentially regulate beta-catenin/T-cell factor activity and
proliferation of colon carcinoma cells. Mol Cell Biol.
27:7802–7815. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chan DW, Mak CS, Leung TH, Chan KK and
Ngan HY: Down-regulation of Sox7 is associated with aberrant
activation of Wnt/b-catenin signaling in endometrial cancer.
Oncotarget. 3:1546–1556. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Saitoh T and Katoh M: Expression of human
SOX18 in normal tissues and tumors. Int J Mol Med. 10:339–344.
2002.PubMed/NCBI
|
|
37
|
García-Ramírez M, Martínez-González J,
Juan-Babot JO, Rodríguez C and Badimon L: Transcription factor
SOX18 is expressed in human coronary atherosclerotic lesions and
regulates DNA synthesis and vascular cell growth. Arterioscler
Thromb Vasc Biol. 25:2398–2403. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Darby IA, Bisucci T, Raghoenath S, Olsson
J, Muscat GE and Koopman P: Sox18 is transiently expressed during
angiogenesis in granulation tissue of skin wounds with an identical
expression pattern to Flk-1 mRNA. Lab Invest. 81:937–943. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim I and Koh GY: Taking aim at Sox18.
Elife. 6(pii): e242382017.PubMed/NCBI
|
|
40
|
Young N, Hahn CN, Poh A, Dong C, Wilhelm
D, Olsson J, Muscat GE, Parsons P, Gamble JR and Koopman P: Effect
of disrupted SOX18 transcription factor function on tumor growth,
vascularization, and endothelial development. J Natl Cancer Inst.
98:1060–1067. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Petrovic I, Milivojevic M, Popovic J,
Schwirtlich M, Rankovic B and Stevanovic M: SOX18 is a novel target
gene of hedgehog signaling in cervical carcinoma cell lines. PLoS
One. 10:e01435912015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jia Y, Cao B, Yang Y, Linghu E, Zhan Q, Lu
Y, Yu Y, Herman JG and Guo M: Silencing NKD2 by promoter region
hypermethylation promotes gastric cancer invasion and metastasis by
up-regulating SOX18 in human gastric cancer. Oncotarget.
6:33470–33485. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang G, Wei Z, Jia H, Zhao W, Yang G and
Zhao H: Knockdown of SOX18 inhibits the proliferation, migration
and invasion of hepatocellular carcinoma cells. Oncol Rep.
34:1121–1128. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ma L, Liu L, Zhang T and Shan L: Detection
and evaluation of EGFR mutation status in serum of patients with
advanced non-small cell lung cancer treated with EGFR-TKIs.
Zhongguo Fei Ai Za Zhi. 16:303–307. 2013.(In Chinese). PubMed/NCBI
|
|
46
|
Meert AP, Martin B, Delmotte P, Berghmans
T, Lafitte JJ, Mascaux C, Paesmans M, Steels E, Verdebout JM and
Sculier JP: The role of EGF-R expression on patient survival in
lung cancer: A systematic review with meta-analysis. Eur Respir J.
20:975–981. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Holzner B, Kemmler G, Greil R, Kopp M,
Zeimet A, Raderer M, Hejna M, Zöchbauer S, Krajnik G, Huber H, et
al: The impact of hemoglobin levels on fatigue and quality of life
in cancer patients. Ann Oncol. 13:965–973. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Guo SC, Yan F, Xu J, Bao Y, Zhu J, Wang X,
Wu J, Li Y, Pu W, Liu Y, et al: Identification and validation of
the methylation biomarkers of non-small cell lung cancer (NSCLC).
Clin Epigenetics. 7:32015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Huang T, Chen X, Hong Q, Deng Z, Ma H, Xin
Y, Fang Y, Ye H, Wang R, Zhang C, et al: Meta-analyses of gene
methylation and smoking behavior in non-small cell lung cancer
patients. Sci Rep. 5:88972015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Azhikina T, Kozlova A, Skvortsov T and
Sverdlov E: Heterogeneity and degree of TIMP4, GATA4, SOX18, and
EGFL7 gene promoter methylation in non-small cell lung cancer and
surrounding tissues. Cancer Genet. 204:492–500. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Olbromski M, Rzechonek A, Grzegrzolka J,
Glatzel-Plucinska N, Chachaj A, Werynska B, Podhorska-Okolow M and
Dziegiel P: Influence of miR-7a and miR-24-3p on the SOX18
transcript in lung adenocarcinoma. Oncol Rep. 39:201–208.
2018.PubMed/NCBI
|
|
53
|
Zhang C, Ge S, Hu C, Yang N and Zhang J:
MiRNA-218, a new regulator of HMGB1, suppresses cell migration and
invasion in non-small cell lung cancer. Acta Biochim Biophys Sin
(Shanghai). 45:1055–1061. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Xie C, Han Y, Liu Y, Han L and Liu J:
miRNA-124 down-regulates SOX8 expression and suppresses cell
proliferation in non-small cell lung cancer. Int J Clin Exp Pathol.
7:7518–7526. 2014.PubMed/NCBI
|
|
55
|
Jankowska-Konsur A, Kobierzycki C, Reich
A, Piotrowska A, Gomulkiewicz A, Olbromski M, Podhorska-Okołów M,
Dzięgiel P and Szepietowski JC: Expression of SOX18 in mycosis
fungoides. Acta Derm Venereol. 97:17–23. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wu Z, Liu J, Wang J and Zhang F: SOX18
knockdown suppresses the proliferation and metastasis, and induces
the apoptosis of osteosarcoma cells. Mol Med Rep. 13:497–504. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bernard P and Harley VR: Acquisition of
SOX transcription factor specificity through protein-protein
interaction, modulation of Wnt signalling and post-translational
modification. Int J Biochem Cell Biol. 42:400–410. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Image from the RCSB PDB of 4Y60: Structure
and decoy-mediated inhibition of the SOX18/Prox1-DNA interaction.
Klaus M., Prokoph N., Girbig M., Wang X., Huang Y.H., Srivastaya
Y., Hou L., Narasimhan K., Kolatkar P.R., Francois M., Jauch R.
(2016) Nucleic Acids Res. 44:3922–3935
|