Introduction
Gastric cancer was the second most common cause of
cancer-associated mortality worldwide in 2014 (1). The long-term survival of patients with
gastric cancer remains unsatisfactory because of increased
incidences of recurrence and chemotherapy resistance (2). A substantial proportion of gastric
cancer cases are either inherently resistant to chemotherapy or
develop resistance during the course of therapy. Therefore, an
improved understanding of the molecular mechanisms involved will
ultimately result in more effective methods of overcoming
chemotherapy resistance and developing novel antineoplastic
treatment strategies.
MicroRNAs (miRNAs/miRs) are genomically encoded,
small, non-coding RNAs (ncRNAs) that negatively regulate gene
expression by controlling either translation or stability of mRNAs
through an RNA interference-like pathway (3). miRNAs comprise 1–3% of the human genome
(4) and regulate 30% of human gene
expression (5). The majority of
miRNAs are located within the introns of either the protein-coding
or non-coding transcriptional units and are expressed with their
host genes coordinately (6). A few
miRNA genes are located in the exons of ncRNAs (6). miRNAs that cluster in the same genome
region are transcribed as polycistronic transcripts (6). Studies have demonstrated that miRNAs are
associated with multiple physiological processes, including aging,
differentiation, hematopoiesis and endocrine functions, and also
function as key regulators in the progression of a number of human
diseases, including heart disease and cancer (7–11).
For the past three decades, changes to the
expression of certain gene has been proposed to be the major
factors of tumorigenesis as well as metastasis (12). The majority of cancer-associated genes
are thought to be protein-coding genes. Previous studies have
demonstrated that there are associations between cancer and miRNA
(13–15). In addition, silencing the expression
of miRNA-processing factors in a transgenic mouse model increases
the susceptibility of patients to cancer (16). Evidence indicates that miRNAs serve
significant functions in almost all aspects of cancer biology,
including cell proliferation, apoptosis, angiogenesis, invasion,
metastatic lesions and drug resistance (17). Recently, multiple dysregulated miRNAs
were found to participate in numerous aspects of gastric cancer
(18). miR-31, miR-106a and miR-21
are reported to possess clinical significance (9). Functions for miR-451, miR-141, miR-34a
and miR-27a have been identified in the progression of gastric
cancer (19–22). miR-15b and miR-16 modulate multidrug
resistance of gastric cancer cells by negatively regulating B-cell
lymphoma 2 (BCL2) expression (23).
Although multiple miRNAs have been shown to function as bona fide
oncogenes or tumor suppressors, the precise functions and the
molecular mechanism underlying their dysregulation of the gastric
cancer progression, as well as the development of chemotherapy
resistance, remain largely unknown (15,24,25). In
addition to individual miRNAs, the functions and regulation of the
genomic miRNA cluster have also not been clearly elucidated in
gastric cancer (26).
The present study analyzed the expression of
microRNAs in gastric cancer tissues using microRNA arrays. In
vitro and in vivo experiments were applied to
investigate their oncogenic functions in gastric cancers.
Materials and methods
Patients and microRNA arrays
Gastric cancer and its corresponding adjacent normal
tissues were obtained from 68 patients who received surgical
resection in the Department of General Surgery, Taipei Veterans
General Hospital (Taipei, Taiwan). A total of 5 patients were
<65 years old, while the remainder were >65 years old.
Overall, 5 patients were female and the remainder were male. Tumor
and adjacent normal tissues were snap-frozen and stored in liquid
nitrogen. The Institutional Review Board of Taipei Veterans General
Hospital approved the use of these tissues. The total RNA from 68
pairs of gastric cancer tissues and their adjacent normal tissues
were extracted by TRIzol® (Life Technologies; Thermo
Fisher Scientific, Inc., Waltham, MA, USA). MicroRNA analyses were
commissioned by the High-throughput Genome Analysis Core Facility
of the VYM Genome Research Center (National Yang-Ming University,
Taipei, Taiwan) on NCode™ Multi-Species miRNA
microarrays (Invitrogen; Thermo Fisher Scientific, Inc.). Only 21
pairs of total RNA passed the quality check of the core facility
for microarray analysis and were subsequently used. Results were
analyzed using the Partek Genomics suite V6.6 (Partek, Inc., St.
Louis, MO, USA) for multi-dimensional scaling, clustering, and heat
map drawing.
Cell lines, cell culture and
transfection
The gastric cancer SC-M1 cell line was obtained from
the American Type Culture Collection (Manassas, VA, USA) and
maintained in RPMI-1640 (Invitrogen; Thermo Fisher Scientific,
Inc.) containing 100 mg/ml penicillin-streptomycin (Invitrogen;
Thermo Fisher Scientific, Inc.) and 10% fetal bovine serum
(Hyclone; GE Healthcare Life Sciences, Logan, UT, USA). A total of
5×105 SC-M1 cells were seeded into 6-well plates for
12-h, with fresh medium replaced to a total volume of 500 µl 1 h
prior to transfection. TransIT TKO reagent (6 µl; Mirus Bio, LLC,
Madison, WI, USA) was mixed with 200 µl serum-free Dulbecco's
modified Eagle's medium (Gibco; Thermo Fisher Scientific, Inc.) for
20 min and then 2 µl microRNA inhibitors (Dharmacon Inc.,
Lafayette, CT, USA) for hsa-miR-23a (IH-300494-05-0010),
hsa-miR-27a (cat. no. IH-300502-05-0020), hsa-miR-24-2
(IH-300497-05-0010) and the control oligo (cat. no.
IN-001-005-01-20) were added for 20 min. The mixture was added into
the well and incubated with the cells at room temperature for 30
min.
Nucleic acid (nuclear) staining/DAPI
staining
Cells pre-treated with 30 nM miRNA inhibitor mix or
scramble RNA for 24 h were washed 3 times in PBS. DAPI stain
solution (300 nM; Thermo Fisher Scientific, Inc.) was added at a
sufficient quantity to cover the cells, then protected from light
and incubated at room temperature for 5 min. The cells were washed
3 times in PBS and visualized by fluorescence microscopy. Normal
nuclear and apoptotic bodies were visualized and distinguished.
Clonogenic assays
For the clonogenic assay, 100 SC-M1 cells that were
pre-treated with 30 or 90 nM of a miR-23a, miR-27a or miR-24
inhibitor, either independently or together at 24 h after
transfection, were seeded in 35-mm dishes. The seeding density was
11 cells/cm2. All experiments were conducted in
triplicate. The dishes were incubated at 37°C in a 5%
CO2 incubator for 15 days and the medium was changed
every 3 days. Colonies were fixed with methanol (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) at room temperature for 10 min,
stained with 1% crystal violet (Sigma-Aldrich; Merck KGaA)
incubated at room temperature for 30 min, and then counted using
ImageQuant TL (version 7.0; GE Healthcare, Chicago IL, USA).
Tumorigenesis in nude mice
All animal experiments were approved by and
performed in accordance with the guidelines of the Institutional
Animal Care and Use Committee of National Yang-Ming University
(Taipei, Taiwan). In total, 3×106 viable SC-M1 cells
pre-treated with either the control oligo or miRNA inhibitor
mixture were washed in PBS and injected subcutaneously into the 2 h
indlimbs of 5-week-old male BALB/c nude (nu/nu) mice to assess
tumor formation (initial bodyweight, 17.6±2.9 g). The mice were
supplied by the National Laboratory Animal Center (Taipei, Taiwan).
Each group contained 5 mice. The housing conditions followed the
guidelines of the Animal Center, National Yang-Ming University
(Taipei, Taiwan) (temperature, 20–22°C; humidity, 50–70%; normal
diet, specific pathogen-free). Primary tumor volume was monitored
with a ruler every 3 days. The tumor size was considered to be the
humane endpoint, with a maximum tumor diameter of 20 mm. The mice
were sacrificed after 27 days. The mice were sacrificed using 30
psi CO2 for <20 sec until cardiac arrest. Tumor
volume was calculated by the formula: Tumor volume=length ×
width2 ×0.5.
microRNA target gene prediction by
TargetScanHuman
TargetScanHuman (http://www.targetscan.org) predicts target genes of
miRNAs by searching for the presence of conserved 6-8mer sites that
match the seed region of each miRNA. It considers matches to human
3′-untranslated regions (UTRs) and their orthologs.
Dual luciferase reporter assay
Luciferase reporter constructs were generated by
cloning a specific miRNA-binding sequence (wild-type/mutants),
3′UTR of SOCS6, into the NotI and XbaI site
located at the psiCHECK2 dual luciferase expression vector (Promega
Corporation, Madison, WI, USA). In total, 1.5×105 SC-M1
cells were seeded in 12-well plates for 16 h and co-transfected
with 100 ng luciferase expression vector and 90 nM miRNA
inhibitors, either independently or jointly, with TransIT TKO
reagent (Mirus Bio, LLC). Subsequent to a 24-h incubation at 37°C,
5% CO2, cell lysates were collected and luciferase
activity (Dual-Luciferase Reporter Assay System; Promega
Corporation) was detected using a microplate reader. Activity was
normalized by comparison with Renilla luciferase.
Western blot analysis
Cell lysates were prepared in RIPA buffer [(150 mM
NaCl, 1% NP-40, 0.5% deoxycholic acid, 0.1% SDS, 50 mM Tris-HCl,
and 5 mM EDTA (pH 7.5)] containing cocktails of protease inhibitors
(Roche, Mannheim, Germany). The relative concentration of protein
in the lysates was determined by BCA protein assay kit (Pierce;
Thermo Fisher Scientific, Inc.). For each lane of 10% SDS-PAGE, 20
µg of protein lysates was separated in gel and subsequently
transferred onto Hybond ECL membranes (Amersham; GE Healthcare,
Chicago, IL, USA). The membranes were gently agitated in the
blocking solution (5% skimmed mik in 1X Tris-buffered saline with
Tween-20 at 4°C overnight) and then were probed with anti-SOCS6
antibody (cat. no. sc-5608; 1:500 dilution; Santa Cruz
Biotechnology, Inc., Callas, TX, USA) and with anti-β-actin
antibody (cat. no. A5316; 1:20,000 dilution; Sigma-Aldrich; Merck
KGaA), as an internal control for protein loading at 4°C 6 h. The
secondary antibodies were anti-mouse IgG and anti-rabbit IgG (cat.
nos. NA931 and NA934; 1:10,000 dilution, Amersham; GE Healthcare,
Chicago, IL, USA). The western blot analysis was detected with
SuperSignal enhanced chemiluminescence reagents (Pierce; Thermo
Fisher Scientific, Inc.) and film autoradiography.
Statistical analysis
Based on SPSS v20 (IBM Corp, Armonk, NY, USA)
differences between groups were analyzed using three-way analysis
of variance followed by Tukey's post-hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Kaplan-Meier survival analysis was also conducted, with P-values
calculated by log-rank test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression profile of miRNA in gastric
cancer
To identify dysregulated miRNAs that may participate
in the tumorigenesis of gastric cancer, miRNA microarray analyses
were performed on 21 pairs of gastric cancer tissues and adjacent
non-neoplastic stomach tissues. Using a three-way analysis of
variance, dysregulated miRNAs were defined as having ≥1.5 folds of
change in their expression levels. The P-value was set at 0.01. A
total of 19 miRNAs were significantly differentially expressed
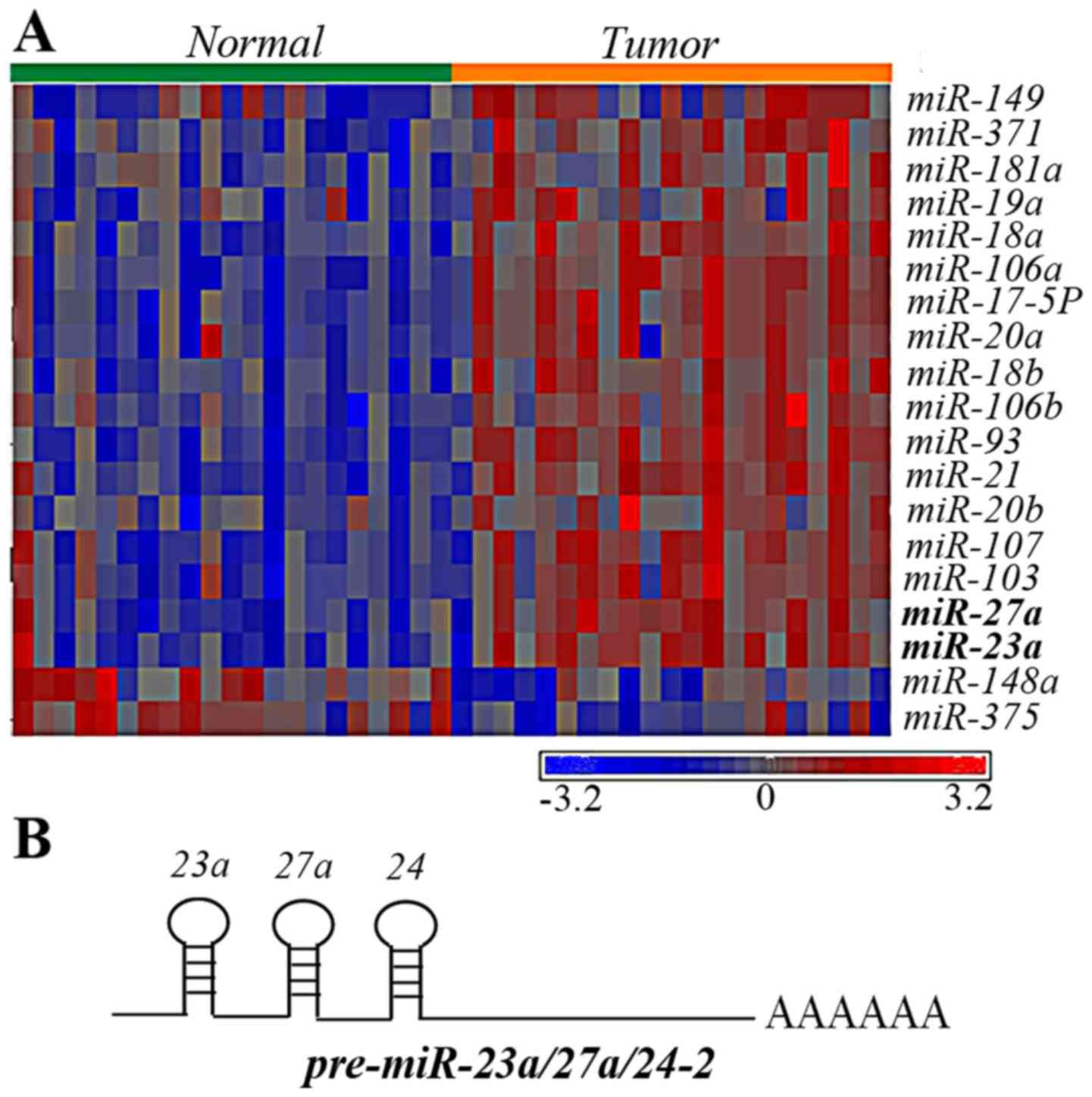
compared with the paired tissues (Fig.
1A). Of the upregulated miRNAs, miR-23a and miR27a, attracted
attention, since they belong to the miR-23a/27a/24-2 cluster
(Fig. 1B). In addition, even though
miR-24 barely reached a significant P-value (P=0.0319), it was
upregulated in 15 out of 21 patients. Therefore, in the present
study it was hypothesized that the miR-23a/27a/24-2 cluster was
upregulated in gastric cancer and possessed oncogenic
activities.
Clonogenic, apoptosis and
proliferation assays for miR inhibitor treatment
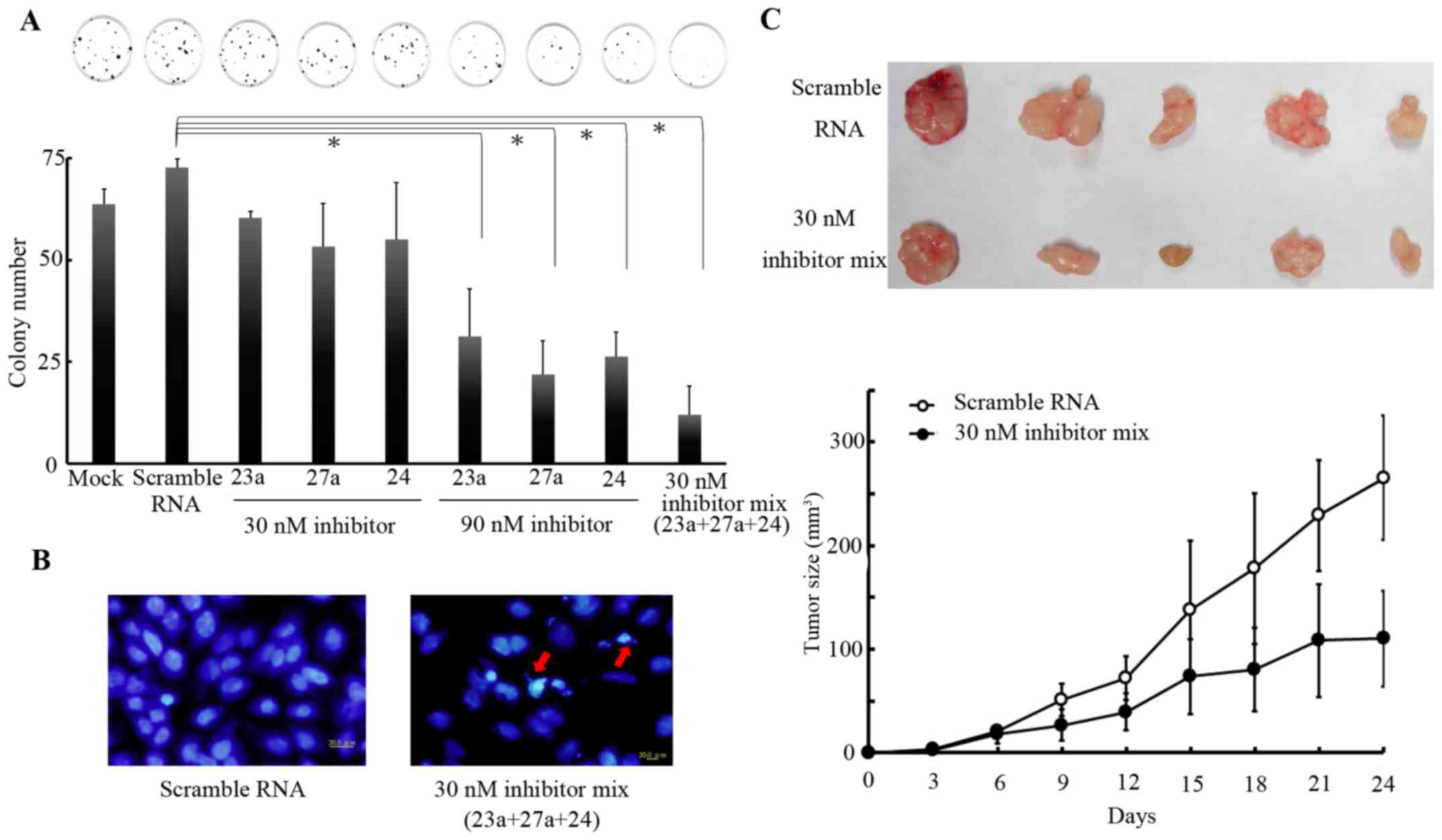
To confirm the aforementioned hypothesis, SC-M1
cells were selected, owing to their increased expression levels of
miR-23a/27a/24-2 cluster. Clonogenic assays were performed in SC-M1
cells that were treated with 30 or 90 nM of a miR-23a, miR-27a or
miR-24 inhibitor, either independently or together at 24 h after
transfection. Results revealed that treatments with either or both
miRNA inhibitors repressed cell proliferation (Fig. 2A). The miR23a/27a/24-2 cluster
inhibitors caused repression of the proliferation of gastric cancer
cells in a synergetic manner. In addition, the treatment with the
decreased dose, 30nM, of the 3 miRNA inhibitors induced the
formation of apoptotic body (Fig.
2B). These results indicated that the miR-23a/27a/24-2 cluster
promoted the proliferation of gastric cancer cells. To validate
these observations in vivo, SC-M1 cells pre-treated with
either the control oligo or miRNA inhibitor mixture were injected
into limbs of nude mice. Tumor size was measured once every 3 days
for 24 days. Tumor growth in the group treated with miRNA inhibitor
mixture was significantly decreased compared with that in the
control group following an incubation period of 21 days (P<0.05;
Fig. 2C). The tumor masses isolated
from mice in the group treated with the 3 miRNA inhibitors were
smaller compared with those in the control group (Fig. 2C). Taken together, these results
indicated that miR-23a/27a/24-2 cluster may possess oncogenic
activities.
Search for the downstream target genes
of miR-23a, miR-27a and miR-24
The downstream targets of miRNAs are critical in
determining the roles of miRNA. Since expression consistency and
growth repression by the mRNA inhibitor mixture was presented in a
synergistic manner, it was presumed that miR-23a, miR-27a, and
miR-24 may act in combination to perform their functions by
targeting either a common downstream gene or various genes in the
same signaling pathway. TargetScanHuman was used to identify the
putative downstream target genes of the miR-23a/27a/24-2 cluster
that may control either cell growth or cell death. Accordingly, the
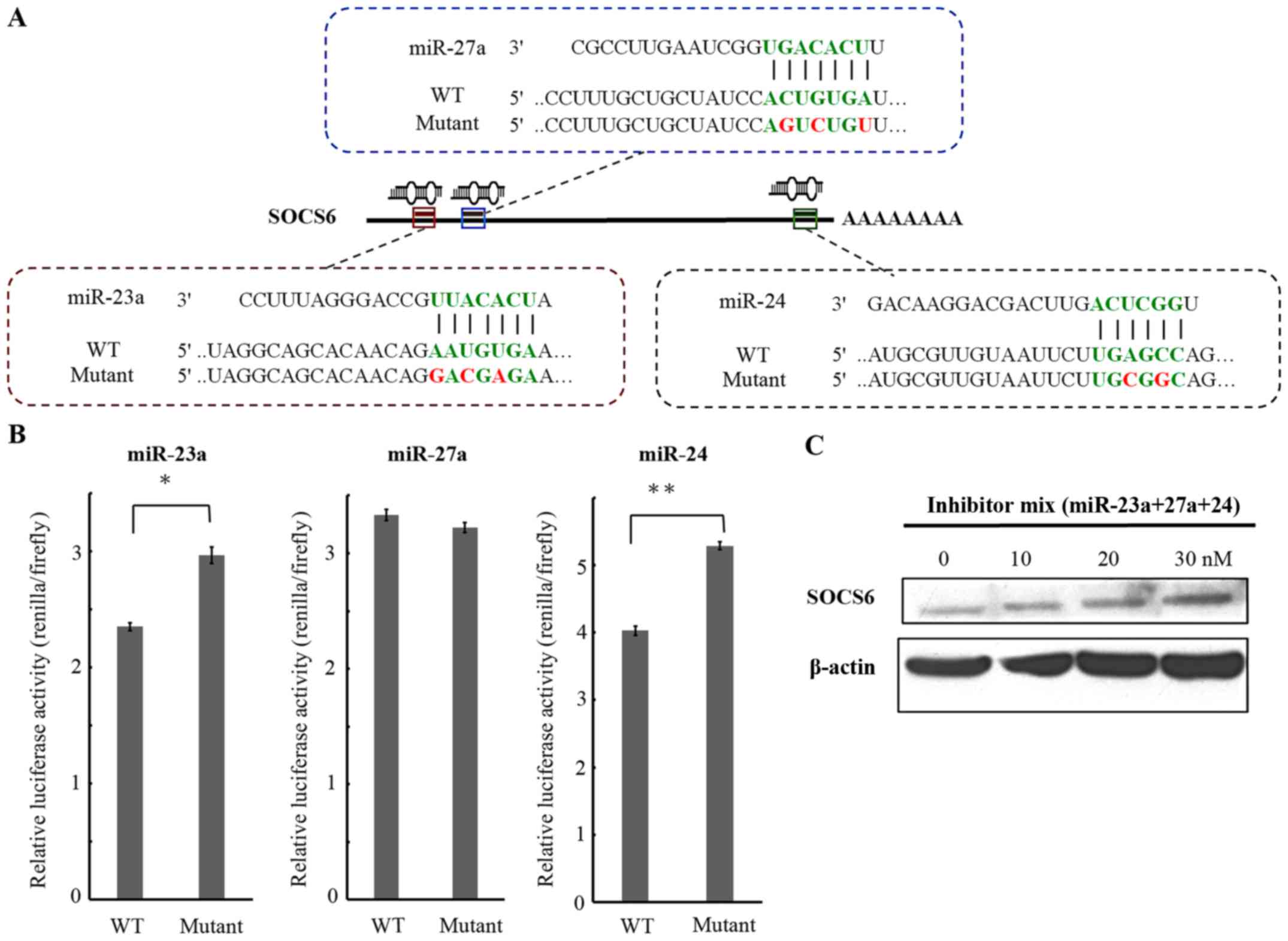
suppressor of cytokine singaling-6 (SOCS6), which belongs to the
SOCS family of negative regulators of the cytokine signaling
pathway (27), was predicted to be a
common potential target for miR-23a, miR-27a and miR-24. There were
3 putative binding sites, located at 240–282, 298–319, and
3099–3120 nt after the stop codon of the SOCS6 mRNA for miR-23a,
miR-27a and miR-24, respectively (Fig.
3A). To demonstrate that the miR-23a/27a/24-2 cluster is able
to target directly to the 3′-UTR of the SOCS6 mRNA, a luciferase
reporter assay was conducted. To perform this assay, 3 reporter
plasmids containing dual luciferase genes followed by DNA fragments
containing the putative target sites for miR-23a, miR-27a, and
miR-24 were constructed. There were also three mutant constructs
with a 2/3-nucleotide alteration within the seed regions of binding
sites generated (Fig. 3A). SC-M1
cells were transfected with the reporter plasmid and three microRNA
inhibitors, either independently or jointly and incubated.
Luciferase activity was measured at 24 h after transfection. In the
three mutant type constructs, when treated with the miRNA
inhibitors individually, the relative luciferase activity increased
31% in the miR-23 mutant and 25% in the miR-24a mutant, compared
with the wild-type-treated group (Fig.
3B). Analysis of the protein expression of SOCS6 revealed that
treatment with the miR-23a, miR27a and miR24 inhibitors were able
to restore the SOCS6 protein expression level in a dose-dependent
manner (Fig. 3C).
Analysis of the expression of SOCS6 in
gastric cancer patients
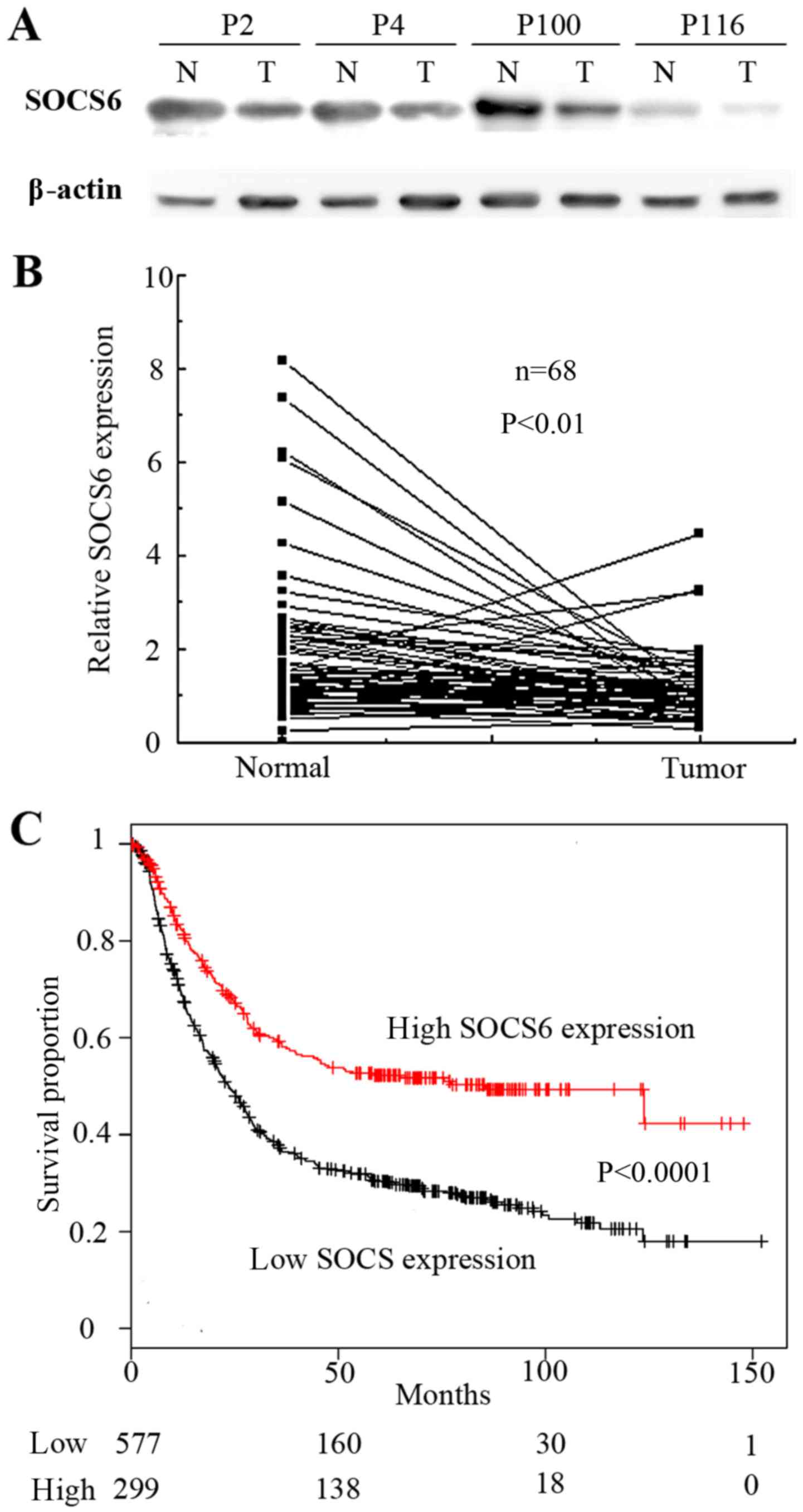
To confirm that SOCS6 was downregulated by the
miR-23a/27a/24-2 cluster in gastric cancer, 68 pairs of clinical
tissue samples were examined by western blot analysis (Fig. 4A). Quantification of western blot
analysis revealed that the relative expression of SOCS6 in the
tumor tissue was significantly lower than that in the normal tissue
(P<0.01; Fig. 4B). To validate
these observations, data from a microarray of 876 gastric cancer
tissues obtained from the Kaplan-Meier plotter (http://kmplot.com/analysis/) (28). Kaplan-Meier analyses showed
downregulation of SOCS6 was significantly associated with poor
patient survival rates (P<0.00001; Fig. 4C). Taken together, these results
indicate that SOCS6 is a downstream target gene of the
miR-23a/27a/24-2 cluster in tumorigenesis of gastric cancer.
Discussion
The present study revealed that the miR-23a/27a/24-2
cluster was highly expressed in gastric cancer. In vivo and
in vitro experiments indicated that miR-23a, miR-27a and
miR-24 promoted tumor formation by enhancing cell growth. Target
site prediction and luciferase reporter assays indicated that SOCS6
was regulated by miR-23a and miR-24. Finally, western blotting and
survival analysis revealed that SOCS6 was downregulated in gastric
cancers and positively associated with patient survival rates.
The miR-23a/27a/24-2 cluster is associated with
multiple diseases (29–33). Even though the 3 miRNAs in this
cluster are derived from a single primary transcript, their
expression patterns vary depending on different biological
conditions (34). For example,
studies on colorectal cancer demonstrated that miR23a and miR27a
were downregulated, but that miR-24 was upregulated in tumor
samples (35). Such complex
expression patterns may be due to post-transcriptional regulation
during precursor processing; however, this mechanism requires
further investigation (36). In the
analysis of miRNA expression profiles in gastric cancer tissues,
the miR-23a/27a/24-2 cluster was considered to be an oncogene owing
to its increased fold-change of upregulation compared with that in
adjacent normal regions. Although the degree of alteration to this
upregulated expression of miR-23a, miR-27a and miR-24 exhibited
variance, this may be due to individual differences between the
miRNAs (data not shown). However, the trend of miR-23a, miR-27a and
miR-24 upregulation was consistent.
Functionally, the miR-23a/27a/24-2 cluster has been
proposed to control the cell cycle, cell proliferation, cell death
and cell differentiation (24–26,28).
In gastric cancer, miR-23a has been reported to promote cell
proliferation by targeting interferon regulator factor 1 and
interleukin-6 receptor (37,38). miR-27a also inhibits the expression of
B-cell translocation gene 2 (BTG2) and prohibitin to facilitate
cell proliferation in gastric cancer cell lines (39,40).
BCL2L11 was targeted by miR-24 to regulate cell proliferation and
apoptosis (41). Yuan et al
(42) demonstrated that miRNAs of a
miRNA cluster may work in combination to accomplish their function.
In this study, miR-23a, miR-27a, and miR-24 were consistently
upregulated in gastric cancer and may function cooperatively to
serve the same function. The results of the present study, which
indicated that the mixture of 3 microRNA inhibitors repressed cell
proliferation in a synergistic manner, strongly supports this
possibility.
SOCS6, located at chromosome 18q22, belongs to a
member of the SOCS family of E3 ubiquitin ligases, a number of
which have been implicated in the negative regulation of cytokine
receptor signaling (27). Unlike
other members of the SOCS family, SOCS6 neither binds to
intermediate components of cytokine signaling pathways nor
represses cytokine receptor signaling (43). SOCS6 is mainly associated with the
negative regulation of receptor signaling by increasing the
degradation, mediated by ubiquitination, of receptors or substrate
proteins and induces apoptosis by targeting mitochondrial proteins
(27). In gastric cancer cells, SOCS6
was reported to inhibit cell proliferation and colony formation
ability, with SOCS6 expression inactivated by either loss of
heterozygosity or epigenetic modification (44). These results also indicated that SOCS6
is commonly downregulated in patients with gastric cancer and is
associated with poor survival rates. However, these results cannot
fully explain those lower-expressed SOCS6 cases, the SOCS6 promoter
regions of which were not hypermethylated. By contrast, the present
study revealed that the expression of SOCS6 was negatively
regulated by miR-23a and miR-24 in gastric cancer, providing an
additional novel mechanism of SOCS6 regulation.
Taken together, the data presented in the present
study indicated the miR-23a/27a/24-2 cluster functions as oncogenic
miRNAs in gastric cancer. Downregulation of SOCS6 expression by
miR-23a and miR-24 may cause the activation of those cell
proliferation signaling pathways and the suppression of the
apoptosis signals, resulting in poorer survival of patients with
gastric cancer. The combined inhibition of miR-23a, miR-27a and
miR-24 may represent an efficient gastric cancer therapy.
Acknowledgements
The authors acknowledge the technical services
provided by High-throughput Genome & Big Data Analysis Core
Facility of VYM Genome Research Center (National Yang-Ming
University). The National Core Facility Program supports the Core
Facility for Biotechnology (NCFPB), Ministry of Science and
Technology.
Funding
The present study was supported by the Yen Tjing
Ling Medical Foundation (grant nos. CI-100-16 and CI-103-14), the
Center of Excellence for Cancer Research at Taipei Veterans General
Hospital (grant nos. DOH101-TD-C-111-007, DOH102-TD-C-111-007 and
MOHW103-TD-B-111-02), the Ministry of Education, Aim for the Top
University Plan (grant nos. 101AC-T406, 103AC-T607 and 104AC-T506)
and the Ministry of Science and Technology, Taiwan (grant no.
MOST105-2319-B-010-001).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
KH designed the study, performed experiments and
bioinformatics analysis, analyzed data, and wrote the manuscript.
YTC performed experiments and analyzed the data. CFC conducted
bioinformatics analysis and assisted interpretation of results.
Y-ST performed experiments and analyzed data. TTH performed animal
experiments and analyzed data. YCL performed experiments and
analyzed data. TSY provided technical and material support and
assisted with analysis and interpretation of results. KHY provided
clinical samples. HCL provided technical support and assisted with
analysis and interpretation of results. MTH assisted with analysis
and interpretation of results. CWC provided clinical samples and
assisted with analysis and interpretation of results. CWW provided
clinical samples. CHL assisted with analysis and interpretation of
results. YHP designed the study, assisted with analysis and
interpretation of results, and wrote the manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Taipei Veterans General Hospital and written informed consent was
obtained from all patients. This study was carried out in strict
accordance with the recommendations in the Guide for the Care and
Use of Laboratory Animals of National Yang Ming University.
Consent for publication
The study was approved by the Ethics Committee of
Taipei Veterans General Hospital and written informed consent was
obtained from all patients.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ang TL and Fock KM: Clinical epidemiology
of gastric cancer. Singapore Med J. 55:621–628. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wu CW, Lo SS, Shen KH, Hsieh MC, Chen JH,
Chiang JH, Lin HJ, Li AF and Lui WY: Incidence and factors
associated with recurrence patterns after intended curative surgery
for gastric cancer. World J Surg. 27:153–158. 2003.PubMed/NCBI
|
|
3
|
Kim VN: MicroRNA biogenesis: Coordinated
cropping and dicing. Nat Rev Mol Cell Biol. 6:376–385. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bentwich I, Avniel A, Karov Y, Aharonov R,
Gilad S, Barad O, Barzilai A, Einat P, Einav U, Meiri E, et al:
Identification of hundreds of conserved and nonconserved human
microRNAs. Nat Genet. 37:766–770. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liang H and Li WH: Lowly expressed human
microRNA genes evolve rapidly. Mol Biol Evol. 26:1195–1198. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rodriguez A, Griffiths-Jones S, Ashurst JL
and Bradley A: Identification of mammalian microRNA host genes and
transcription units. Genome Res. 14:1902–1910. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee YS and Dutta A: MicroRNAs in cancer.
Annu Rev Pathol. 4:199–227. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhu K, Liu D, Lai H, Li J and Wang C:
Developing miRNA therapeutics for cardiac repair in ischemic heart
disease. J Thorac Dis. 8:E918–E927. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liang W, Gao S, Liang L, Huang X, Hu N, Lu
X and Zhao Y: miRNA-34b is directly involved in the aging of
macrophages. Aging Clin Exp Res. 29:599–607. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shi C, Huang F, Gu X, Zhang M, Wen J, Wang
X, You L, Cui X, Ji C and Guo X: Adipogenic miRNA and
meta-signature miRNAs involved in human adipocyte differentiation
and obesity. Oncotarget. 7:40830–40845. 2016.PubMed/NCBI
|
|
11
|
Petriv OI, Hansen CL, Humphries RK and
Kuchenbauer F: Probing the complexity of miRNA expression across
hematopoiesis. Cell Cycle. 10:2–3. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hunter T: Cooperation between oncogenes.
Cell. 64:249–270. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He L, Thomson JM, Hemann MT,
Hernando-Monge E, Mu D, Goodson S, Powers S, Cordon-Cardo C, Lowe
SW, Hannon GJ and Hammond SM: A microRNA polycistron as a potential
human oncogene. Nature. 435:828–833. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu Q, Yang X, Duan W, Li C, Luo Y and Lu
S: miRNA-346 promotes proliferation, migration and invasion in
liver cancer. Oncol Lett. 14:3255–3260. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Y, Wu YY, Jiang JN, Liu XS, Ji FJ
and Fang XD: MiRNA-3978 regulates peritoneal gastric cancer
metastasis by targeting legumain. Oncotarget. 7:83223–83230.
2016.PubMed/NCBI
|
|
16
|
Kumar MS, Lu J, Mercer KL, Golub TR and
Jacks T: Impaired microRNA processing enhances cellular
transformation and tumorigenesis. Nat Genet. 39:673–677. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Garzon R, Marcucci G and Croce CM:
Targeting microRNAs in cancer: Rationale, strategies and
challenges. Nat Rev Drug Discov. 9:775–789. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Song JH and Meltzer SJ: MicroRNAs in
pathogenesis, diagnosis, and treatment of gastroesophageal cancers.
Gastroenterology. 143:35–47, e2. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Su Z, Zhao J, Rong Z, Geng W and Wang Z:
MiR-451, a potential prognostic biomarker and tumor suppressor for
gastric cancer. Int J Clin Exp Pathol. 8:9154–9160. 2015.PubMed/NCBI
|
|
20
|
Du Y, Xu Y, Ding L, Yao H, Yu H, Zhou T
and Si J: Down-regulation of miR-141 in gastric cancer and its
involvement in cell growth. J Gastroenterol. 44:556–561. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao X, Yang L and Hu J: Down-regulation
of miR-27a might inhibit proliferation and drug resistance of
gastric cancer cells. J Exp Clin Cancer Res. 30:552011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cao W, Fan R, Wang L, Cheng S, Li H, Jiang
J, Geng M, Jin Y and Wu Y: Expression and regulatory function of
miRNA-34a in targeting survivin in gastric cancer cells. Tumour
Biol. 34:963–971. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xia L, Zhang D, Du R, Pan Y, Zhao L, Sun
S, Hong L, Liu J and Fan D: miR-15b and miR-16 modulate multidrug
resistance by targeting BCL2 in human gastric cancer cells. Int J
Cancer. 123:372–379. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang L, Liang H, Wang Y, Gao S, Yin K, Liu
Z, Zheng X, Lv Y, Wang L, Zhang CY, et al: MiRNA-203 suppresses
tumor cell proliferation, migration and invasion by targeting Slug
in gastric cancer. Protein Cell. 7:383–387. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang R, Li F, Wang W, Wang X, Li S and
Liu J: The effect of antisense inhibitor of miRNA 106b~25 on the
proliferation, invasion, migration, and apoptosis of gastric cancer
cell. Tumour Biol. 37:10507–10515. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kabir NN, Sun J, Ronnstrand L and Kazi JU:
SOCS6 is a selective suppressor of receptor tyrosine kinase
signaling. Tumour Biol. 35:10581–10589. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Szász AM, Lánczky A, Nagy A, Förster S,
Hark K, Green JE, Boussioutas A, Busuttil R, Szabó A and Győrffy B:
Cross-validation of survival associated biomarkers in gastric
cancer using transcriptomic data of 1,065 patients. Oncotarget.
7:49322–49333. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Abu-Elneel K, Liu T, Gazzaniga FS,
Nishimura Y, Wall DP, Geschwind DH, Lao K and Kosik KS:
Heterogeneous dysregulation of microRNAs across the autism
spectrum. Neurogenetic. 9:153–161. 2008. View Article : Google Scholar
|
|
30
|
Sayed D, Hong C, Chen IY, Lypowy J and
Abdellatif M: MicroRNAs play an essential role in the development
of cardiac hypertrophy. Circ Res. 100:416–424. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
van Rooij E, Sutherland LB, Liu N,
Williams AH, McAnally J, Gerard RD, Richardson JA and Olson EN: A
signature pattern of stress-responsive microRNAs that can evoke
cardiac hypertrophy and heart failure. Proc Natl Acad Sci USA.
103:18255–18260. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Murakami Y, Yasuda T, Saigo K, Urashima T,
Toyoda H, Okanoue T and Shimotohno K: Comprehensive analysis of
microRNA expression patterns in hepatocellular carcinoma and
non-tumorous tissues. Oncogene. 25:2537–2545. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Perkins DO, Jeffries CD, Jarskog LF,
Thomson JM, Woods K, Newman MA, Parker JS, Jin J and Hammond SM:
microRNA expression in the prefrontal cortex of individuals with
schizophrenia and schizoaffective disorder. Genome Biol. 8:R272007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chhabra R, Dubey R and Saini N:
Cooperative and individualistic functions of the microRNAs in the
miR-23a~27a~24-2 cluster and its implication in human diseases. Mol
Cancer. 9:2322010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xi Y, Shalgi R, Fodstad O, Pilpel Y and Ju
J: Differentially regulated micro-RNAs and actively translated
messenger RNA transcripts by tumor suppressor p53 in colon cancer.
Clin Cancer Res. 12:2014–2024. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ma S, Liu M, Xu Z, Li Y, Guo H, Ge Y, Liu
Y, Zheng D and Shi J: A double feedback loop mediated by
microRNA-23a/27a/24-2 regulates M1 versus M2 macrophage
polarization and thus regulates cancer progression. Oncotarget.
7:13502–13519. 2016.PubMed/NCBI
|
|
37
|
Liu X, Ru J, Zhang J, Zhu LH, Liu M, Li X
and Tang H: miR-23a targets interferon regulatory factor 1 and
modulates cellular proliferation and paclitaxel-induced apoptosis
in gastric adenocarcinoma cells. PLoS One. 8:e647072013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhu LH, Liu T, Tang H, Tian RQ, Su C, Liu
M and Li X: MicroRNA-23a promotes the growth of gastric
adenocarcinoma cell line MGC803 and downregulates interleukin-6
receptor. FEBS J. 277:3726–3734. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhou L, Liang X, Zhang L, Yang L, Nagao N,
Wu H, Liu C, Lin S, Cai G and Liu J: MiR-27a-3p functions as an
oncogene in gastric cancer by targeting BTG2. Oncotarget.
7:51943–51954. 2016.PubMed/NCBI
|
|
40
|
Liu T, Tang H, Lang Y, Liu M and Li X:
MicroRNA-27a functions as an oncogene in gastric adenocarcinoma by
targeting prohibitin. Cancer Lett. 273:233–242. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang H, Duan J, Qu Y, Deng T, Liu R,
Zhang L, Bai M, Li J, Ning T, Ge S, et al: Onco-miR-24 regulates
cell growth and apoptosis by targeting BCL2L11 in gastric cancer.
Protein Cell. 7:141–151. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yuan X, Liu C, Yang P, He S, Liao Q, Kang
S and Zhao Y: Clustered microRNAs' coordination in regulating
protein-protein interaction network. BMC Syst Biol. 3:652009.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Masuhara M, Sakamoto H, Matsumoto A,
Suzuki R, Yasukawa H, Mitsui K, Wakioka T, Tanimura S, Sasaki A,
Misawa H, et al: Cloning and characterization of novel CIS family
genes. Biochem Biophys Res Commun. 239:439–446. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lai RH, Hsiao YW, Wang MJ, Lin HY, Wu CW,
Chi CW, Li AF, Jou YS and Chen JY: SOCS6, down-regulated in gastric
cancer, inhibits cell proliferation and colony formation. Cancer
Lett. 288:75–85. 2010. View Article : Google Scholar : PubMed/NCBI
|