Introduction
Acute myeloid leukemia (AML), which is a clonal
disorder of the myeloid line of blood cells, progresses rapidly and
is associated with poor clinical prognosis (1). AML most commonly affects older adults,
with a median age at diagnosis of 67 years, and 1/3 AML patients
are >75 years old (1). The ‘3+7′
regimen (3 days of anthracycline and 7 days of cytarabine
treatment) has remained unchanged for >30 years, and has a
60–80% complete response (CR) rate in adult patients with AML
<60 years old, and a 40–60% CR rate in patients >60 years
old. The overall curative rate of conventional chemotherapy is only
30–40% in patients <60 years old, and <10% in elderly
patients or those with adverse karyotypes (2,3). Patients
who experience relapsed or refractory AML, are associated with poor
outcomes (3).
The low-dose cytarabine, aclarubicin and
granulocyte-colony stimulating factor (G-CSF) (CAG) priming regimen
was first developed by Yamada et al (4) for the treatment of relapsed AML in 1995.
CAG chemotherapy has since been investigated as an effective
treatment for patients with relapsed or refractory AML and advanced
myelodysplastic syndrome (MDS) in Japan and China (5–9). The
rationale for this regimen is that G-CSF potentiates the
anti-leukemic effect of cytosine arabinoside (AraC) (10–12) by
recruiting quiescent G0 leukemic cells into the cell cycle
(11), thus improving the CR rate of
patients with refractory and relapsed leukemia.
Hematopoietic stem cells (HSCs) exist within the
niche of the bone marrow microenvironment (BMM) which regulates
stem cell survival, proliferation, differentiation and apoptosis
(13). The stromal-derived factor-1α
(SDF-1α)/C-X-C chemokine receptor type 4 (CXCR4) axis is the key
factor associated with HSC chemotaxis (14–16),
homing (17,18) and survival/anti-apoptosis (19,20). It
has also been demonstrated to dynamically mediate HSC-trafficking
in the bone marrow (18). Inhibition
of the SDF-1α/CXCR4 axis using the CXCR4 antagonist, AMD3100, has
been indicated to induce leukemia cells to enter the peripheral
circulation and induce higher sensitivity to chemotherapeutic drugs
(21). G-CSF, an effective stem
cell-mobilizing agent, induces SDF-1α secretion from bone marrow
stromal cells into the blood, thus recruiting CXCR4+
cells, including HSCs, into the peripheral circulation (22). It has been established that G-CSF can
promote the expression of transcriptional repressor growth factor
independence-1 (Gfi-1), which binds to DNA sequences upstream of
the CXCR4 gene and reduces CXCR4 expression in myeloid cells
(23).
It has been demonstrated that the BMM serves a
crucial role in leukemia development and progression, and that it
becomes immunosuppressive. Several types of immunosuppressive
cells, including regulatory T cells (Tregs) (24), myeloid-derived suppressor cells
(MDSCs) (25) and tumor-associated
macrophages (TAMs) (26), contribute
to the immunologically permissive microenvironment and aid in tumor
immune evasion. Tregs and MDSCs mediate their suppressive effects
and promote tumor progression by inhibiting T-cell priming
(24,25). Furthermore, they provide a favorable
microenvironment in which cancer cells can proliferate, expand and
evade host immunosurveillance (24,25). As
previously reported, SDF-1α is involved in the chemotaxis and
adhesion of Tregs and HSCs (27,28). G-CSF
can mobilize Tregs from the bone marrow into the peripheral blood
by reducing bone marrow-derived SDF-1α expression (27). G-CSF has been identified as a major
factor in the differentiation of granulocytic MDSCs (gMDSCs), and
tumor-derived granulocyte-macrophage colony-stimulating factor
(GM-CSF) has been demonstrated to serve a key role in monocytic
MDSC (mMDSC) production (29–31).
The present study examined the alteration of the
immunosuppressive BMM following administration of G-CSF in
combination with low-dose chemotherapy. An established syngeneic
leukemia mouse model using the murine AML WEHI-3 cell line
(32,33) was used to determine whether the BMM
would be altered in vivo following treatment with the CAG
priming regimen.
Materials and methods
Cells and reagents
The murine AML cell line, WEHI-3, was maintained in
the laboratory of the Department of Clinical Hematology, Second
Affiliated Hospital, Medical School of Xi'an Jiao Tong University
(Xi'an, China) in DMEM medium supplemented with 10% fetal bovine
serum (Biological Industries, Israel) and 1%
penicillin-streptomycin (100 U/ml penicillin and 100 mg/ml
streptomycin; cat no. 15140-122; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). The following antibodies were used for
flow cytometry (FCM): Fluorescein isothiocyanate (FITC) anti-mouse
cluster of differentiation (CD)4 (cat no. 11-0042), allophycocyanin
(APC) anti-mouse CD25 (cat no. 17-0251), phycoerythrin (PE)
anti-mouse/rat Forheac box P3 (Foxp3) (cat no. 12-5773) and PE
anti-mouse CD184 (CXCR4; cat no. 12-9991), and all were purchased
from eBioscience (Thermo Fisher Scientific, Inc.). FITC anti-mouse
CD11b (cat no. 557396), PE anti-mouse lymphocyte antigen 6G (Ly-6G)
and Ly-6C (cat no. 561084) were purchased from BD Biosciences (San
Jose, CA, USA). The mouse SDF-1α (cat. no. MCX120), transforming
growth factor β (TGF-β; cat. no. MB100B) and interleukin 10 (IL-10;
cat. no. M1000B) ELISA kits were purchased from R&D Systems,
Inc. (Minneapolis, MN, USA), and the arginase 1 (Arg-1; cat. no.
JL13668) ELISA kit was purchased from and Shanghai Jiang Lai
Biotechnology Co., Ltd. (Shanghai, China; http://www.laibio.com/).
Leukemia mouse model and chemotherapy
regimens
All animal experiments were reviewed and approved by
the Ethics Committee of the Medical College, Xi'an Jiao Tong
University (Xi'an, China). Male BALB/c wild-type mice (6–8 weeks
old, weighed 21±2 g) were purchased from the Laboratory Animal
Center, Medical College of Xi'an Jiao Tong University. The mice
were maintained under specific pathogen-free conditions, in a 12
/12 h light/dark cycle at 21±2°C, with access to food and water
ad libitum. A total of 21 mice were randomly divided into 3
groups of 7. For the syngeneic acute myelomonocytic leukemia model
(AML-M4), BALB/c mice were injected intravenously into the tail
vein on day 0 with 1×106 WEHI-3 cells in 100 µl PBS as
previously described (32–34). On day 15, all 7 mice in the AML
(control) group were humanely sacrificed and the spleen, liver,
femur and blood were collected and stored for further experiments.
For the daunorubicin and cytarabine (DNR-AraC; DA) group, 7 mice
were treated with 10 mg/kg DNR intravenously on days 15–17 and 50
mg/kg AraC intraperitoneally on days 15–21. Mice in DA group were
sacrificed on day 22 and the spleen, liver, femur and blood were
collected for further experiments. For the CAG group, 7 mice were
treated with 6.25 mg/kg AraC via hypodermic injection (IH) on days
15–18, 3 mg/kg aclarubicin (ACR) intravenously on days 15–28 and
0.2 mg/kg G-CSF via IH on days 15–28. All mice in CAG group were
sacrificed on day 29 and the spleen, liver, femur and blood were
collected for further experiments.
ELISA
SDF-1α, IL-10 and Arg-1 levels in the blood plasma
were determined using the aforementioned commercially available
mouse ELISA kits, according the manufacturers' protocols.
Absorbance was measured at 450 nm on a microplate reader (Thermo
Fisher Scientific, Inc.).
FCM analysis
Single-cell suspensions of spleen cells and bone
marrow mononuclear cells (BMMCs) were stained with FITC-conjugated
anti-mouse CD4 (dilution, 1:100) and APC-conjugated anti-mouse CD25
(dilution, 1:100) for 30 min on ice. The cells were washed with
flow cytometry staining buffer (eBioscience; Thermo Fisher
Scientific, Inc.), resuspended in 1 ml fixation/permeabilization
working solution (eBioscience; Thermo Fisher Scientific, Inc.) and
incubated at 4°C, in the dark, overnight. The cells were washed
twice with 1× permeabilization buffer (eBioscience; Thermo Fisher
Scientific, Inc.) and incubated for 30 min at 4°C in 1×
permeabilization buffer containing PE-conjugated anti-mouse Foxp3
(dilution, 1:40) antibody. Forward vs. side scatter was plotted for
lymphocytes, followed by gating for CD4+ T cells. These
cells were then analyzed for the expression of CD25 and FoxP3. For
the detection of MDSCs and CXCR4, single-cell suspensions were
stained with CD11b (dilution, 1:200), Gr-1 (Ly-6G and Ly-6C;
dilution, 1:200) or CXCR4 (dilution, 1:20) antibodies. FCM was
performed using a FACS flow cytometer (BD Biosciences, Franklin
Lakes, NJ, USA) and the data was analyzed using CellQuest Pro
(version 6.0; BD Biosciences).
Pathological examinations
The femur, liver and spleen tissues were harvested
and immediately fixed in 10% neutral-buffered formalin for 24 h.
Liver and spleen tissues were embedded in paraffin and the femurs
were decalcified in 10% EDTA for ~30 days at room temperature prior
to being embedded in paraffin. A total of 54-µm-thick sections were
cut from each paraffin block. The sections were stained at room
25°C using hematoxylin (H) for 6 min and eosin (E) for 1 min (cat
no. C0105; Beyotime Institute of Biotechnology, Shanghai, China),
according to standard histochemical procedures. Images were
acquired using a light microscope (DP71; Olympus Corporation,
Tokyo, Japan).
Statistical analysis
All data are expressed as the mean ± standard
deviation. Differences between treatment groups were determined
using analysis of variance followed by Fisher's Least Significant
Difference post-hoc test. P<0.05 was considered to indicate a
statistically significant difference. All analyses were performed
using SPSS 18.0 software (SPSS Inc., Chicago, IL, USA).
Results
CAG treatment regimen decreases the
number of Tregs and MDSCs in bone marrow
To determine whether the CAG and DA treatment
regimens affect AML immunosuppressive cells in the BMM, FCM was
performed to investigate the percentage of
CD4+CD25+FoxP3+ Tregs and
CD11b+Gr-1+ MDSCs in BMMCs. It was revealed
that the levels of Tregs and MDSCs in the DA and CAG groups were
significantly lower than in the AML group (P<0.01; Fig. 1A). Furthermore, compared with mice in
the DA group, mice in CAG group exhibited significantly decreased
levels of Tregs and MDSCs in BMMCs (P<0.05; Fig. 1A).
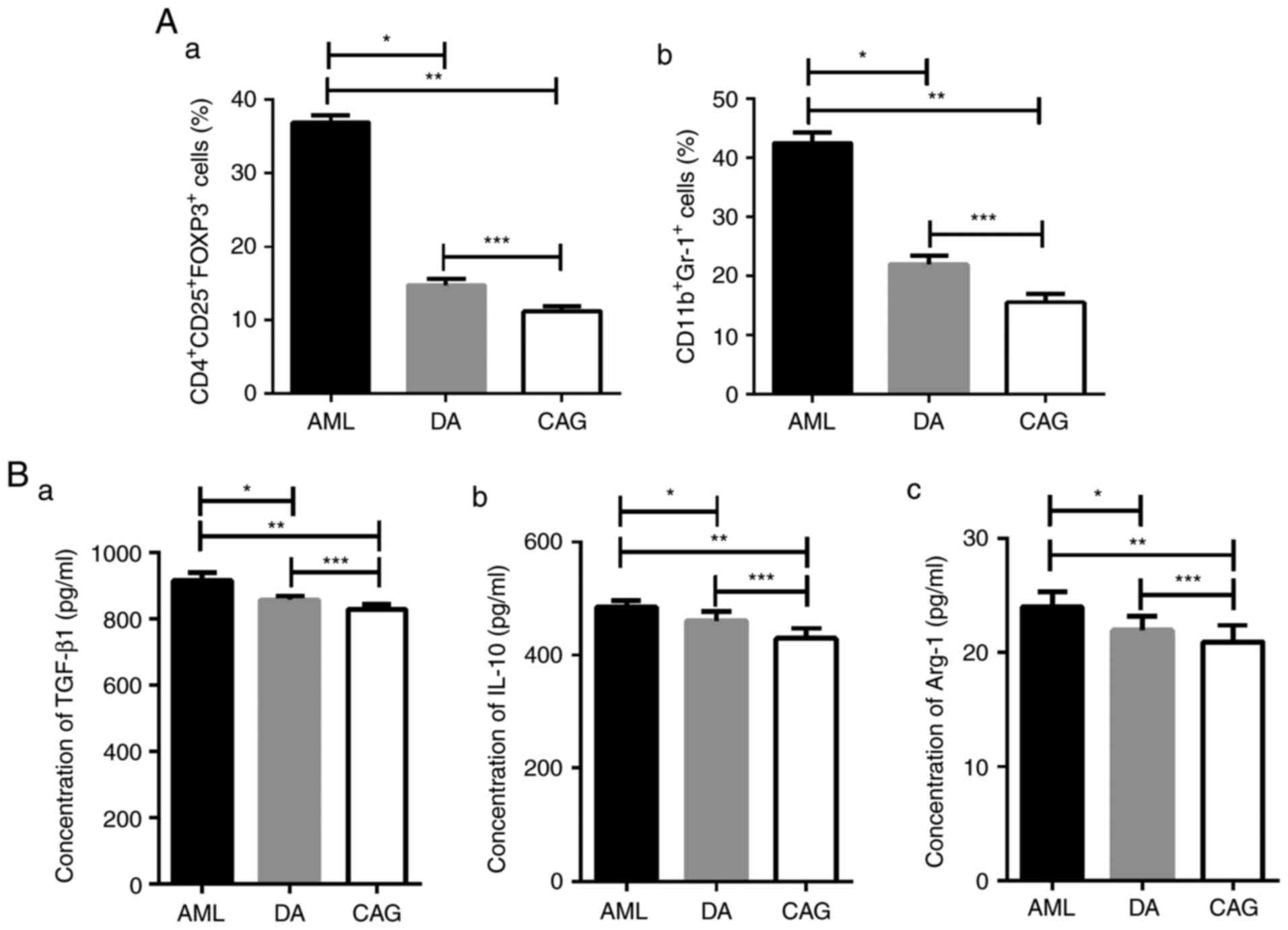 | Figure 1.Tregs and MDSC numbers and cytokines
levels in the bone marrow are reduced with the CAG regimen. (A) (a)
Percentage of CD4+CD25+Foxp3+
Tregs in each group, analyzed by FCM. *P<0.01, **P<0.01,
***P<0.05; (b) Percentage of CD11b+Gr-1+
MDSCs in each group, analyzed by FCM. *P<0.01, **P<0.01,
***P<0.05. (B) (a) Concentration of TGF-β1 in the bone marrow
supernatant, analyzed by ELISA. *P<0.01, **P<0.01,
***P<0.01; (b) Concentration of IL-10 in bone marrow
supernatant, analyzed by ELISA. *P<0.01, **P<0.01,
***P<0.01; (c) Concentration of Arg-1 in bone marrow
supernatant, analyzed by ELISA. *P<0.01, **P<0.01,
***P<0.05. Tregs, regulatory T cells; MDSCs, myeloid-derived
suppressor cells; CAG, cytarabine + aclarubicin +
granulocyte-colony stimulating factor; CD, cluster of
differentiation; Foxp3, forkhead box protein 3; FCM, flow
cytometry; TGF-β1, transforming growth factor-β1; IL-10,
interleukin-10; Arg-1, arginase-1; AML, acute myeloid leukemia; DA,
daunorubicin + cytarabine. |
CAG treatment regimen decreases Tregs
and MDSCs associated cytokine levels in the BMM
TGF-β and IL-10 are anti-inflammatory and
immunosuppressive cytokines secreted by Tregs (35,36), while
Arg-1, which suppresses T-lymphocyte function, is expressed by
MDSCs (37,38). The ELISA results demonstrated that the
concentrations of TGF-β, IL-10 and Arg-1 in the bone marrow
supernatant were significantly decreased in the DA (P<0.01) and
CAG (P<0.01) groups compared with the AML group (Fig. 1B). The CAG group exhibited lower
TGF-β1 (P<0.01), IL-10 (P<0.01) and Arg-1 (P<0.05)
concentrations compared with the DA group (Fig. 1B). These results suggest that the CAG
treatment regimen may rescue immune suppression in AML bone
marrow.
Compared with the DA treatment
regimen, the CAG treatment regimen increases Treg and MDSC numbers
in the spleen
The percentages of Tregs and MDSCs in mice spleen
single cell suspensions were tested by FCM. As shown in Fig. 2, mice in the DA and CAG groups both
exhibited lower percentages of Tregs and MDSCs in spleen than AML
group (P<0.01). However, there were significantly higher levels
of Tregs (P<0.01) and MDSCs (P<0.05) in the CAG group than
the DA group.
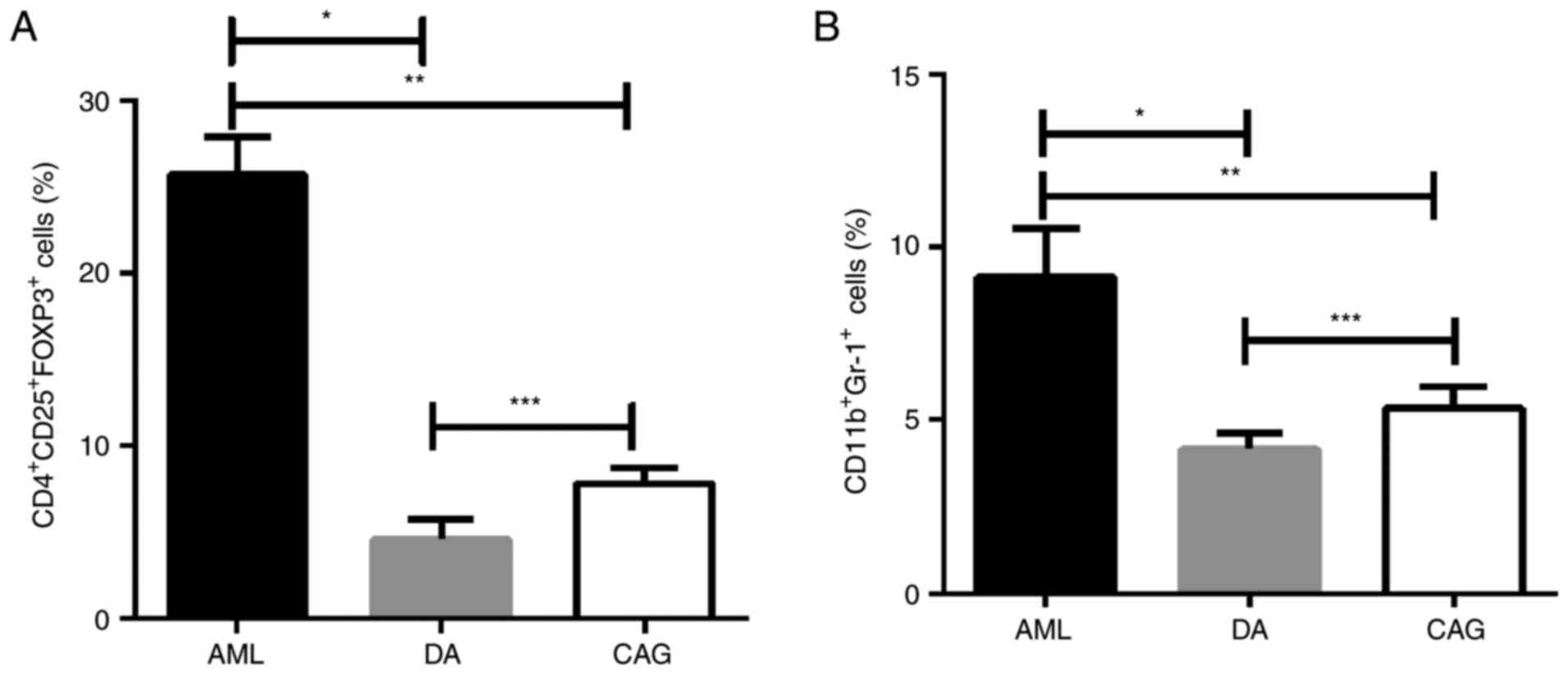 | Figure 2.CAG regimen increases Tregs and MDSC
numbers in the spleen compared with the DA regimen. (A) Percentage
of CD4+CD25+FoxP3+ Tregs in each
group, analyzed by FCM. *P<0.01, **P<0.01, ***P<0.01. (B)
Percentage of CD11b+Gr-1+ MDSCs in each
group, analyzed by FCM. *P<0.01, **P<0.01, ***P<0.05. CAG,
cytarabine + aclarubicin + granulocyte-colony stimulating factor;
Tregs, regulatory T cells; MDSCs, myeloid-derived suppressor cells;
DA, daunorubicin + cytarabine; CD, cluster of differentiation;
Foxp3, forkhead box protein 3; FCM, flow cytometry; AML, acute
myeloid leukemia. |
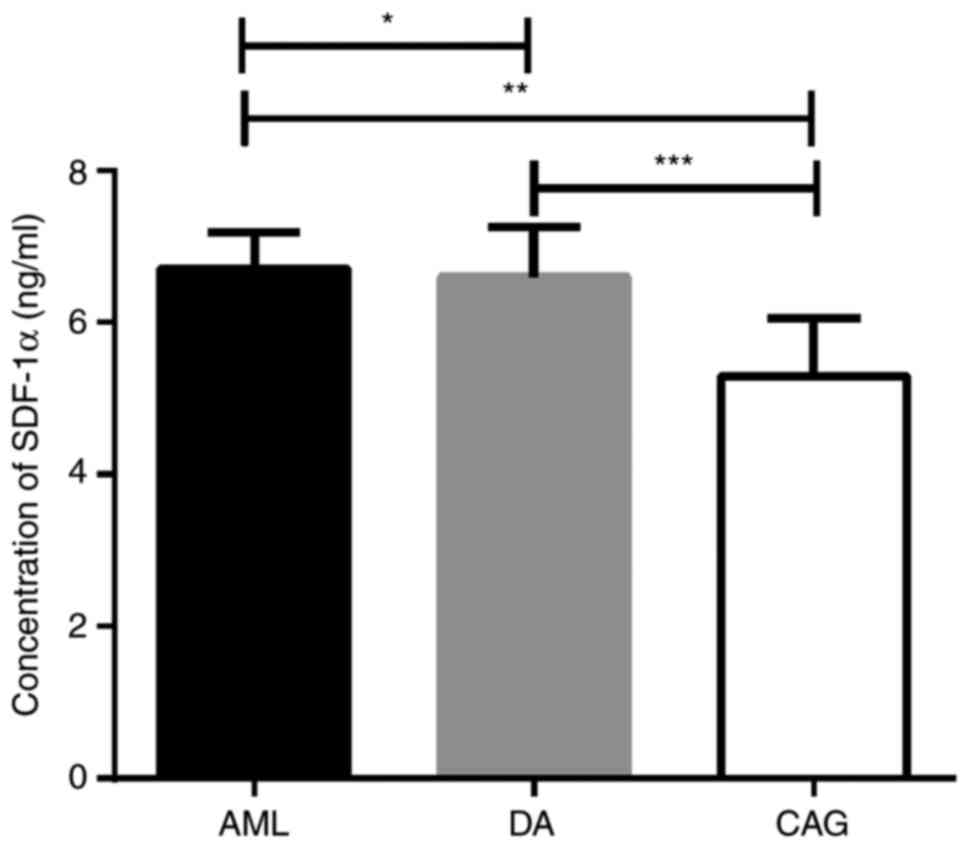
CAG treatment regimen downregulates
SDF-1α levels in the bone marrow
The level of SDF-1α in bone marrow supernatant was
detected by ELISA, and the results revealed that the concentration
of SDF-1α in the CAG group was significantly lower than the AML and
DA groups (P<0.01). However, there was no significant difference
(in the concentration of bone marrow SDF-1α between AML and DA
groups (P>0.05; Fig. 3).
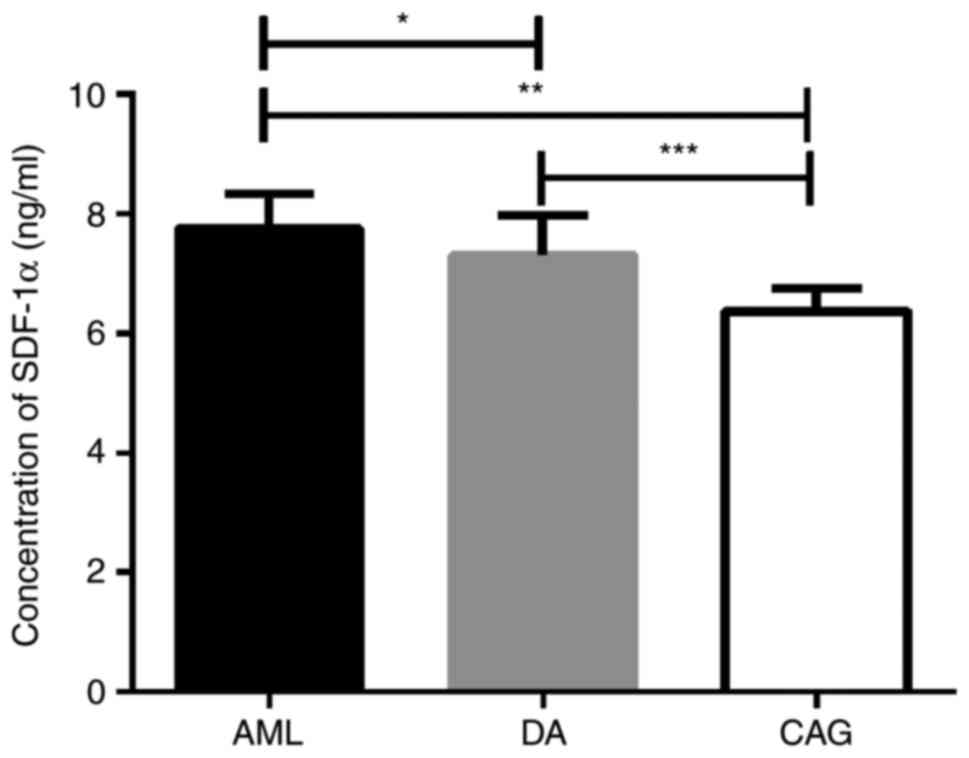
CAG treatment regimen reduces SDF-1α
levels in the peripheral blood
ELISA results demonstrated that the CAG group
exhibited the lowest levels of SDF-1α in the peripheral blood among
the 3 groups of mice (Fig. 4). In
consistence with the concentration of SDF-1α in the bone marrow,
there was no significant difference in the concentration of SDF-1α
in the peripheral blood between the AML and DA groups
(P>0.05).
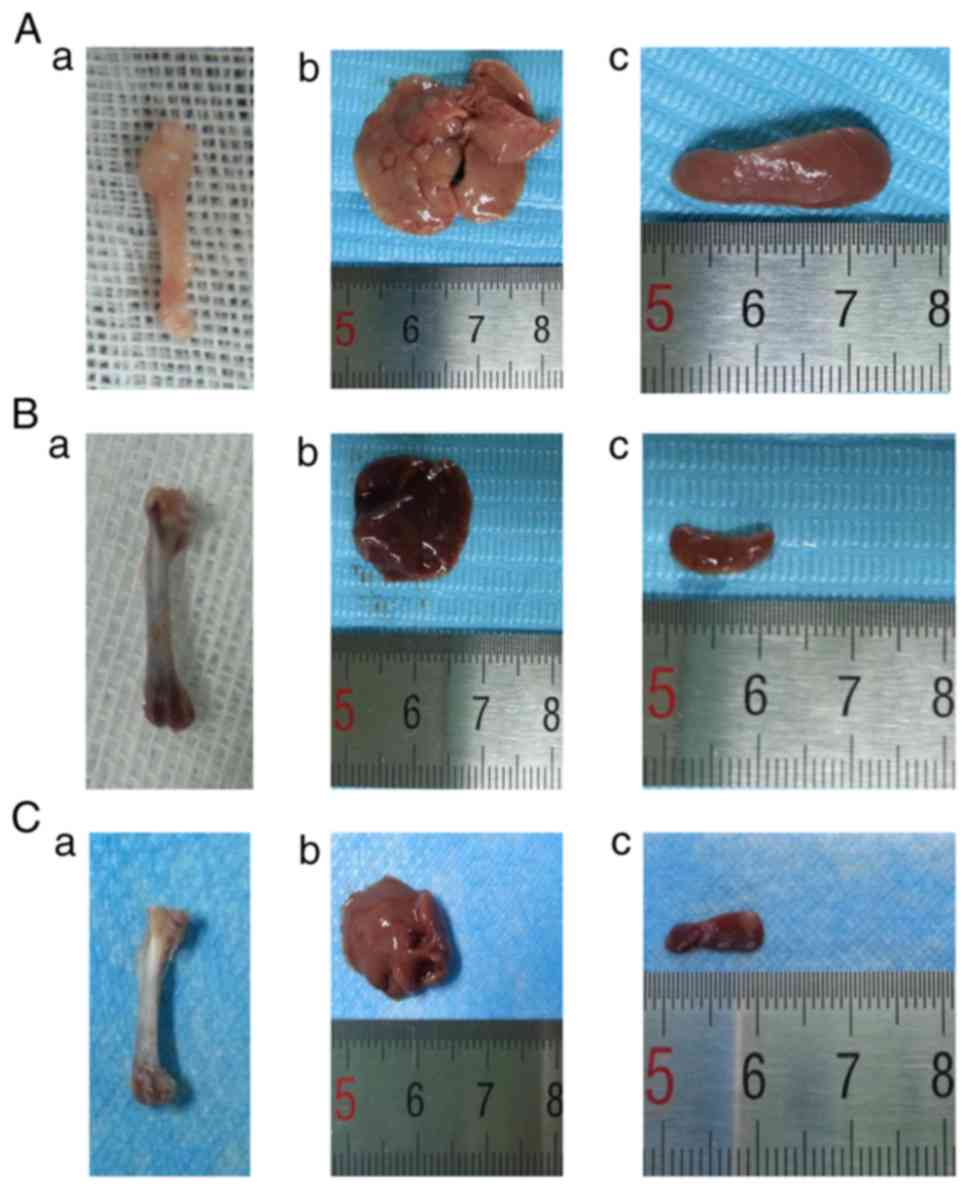
Histopathology of the femur, liver and
spleen tissues
Macroscopically, the femurs of AML mice were pale
(Fig. 5Aa) and the liver and spleen
were swollen (Fig. 5Ab and c)
compared with the DA and CAG mice. The DA (Fig. 5Ba) and CAG (Fig. 5Ca) treatment regimens reddened the
femurs and decreased the size of the liver and spleen (Fig. 5Bb and c; Fig. 5Cb and c). Leukemic cells in the bone
marrow cavity of mice from the AML group were visible under the
light microscope (Fig. 6Aa). By
contrast, the femur sections from mice of the DA and CAG groups
exhibited a recovered hematopoietic system (Fig. 6Ba and Ca). In AML mice, leukemic cells
infiltrated the liver and spleen (Fig.
6Ab and c). However, in the DA group, fewer leukemic cells
infiltrated the liver and spleen and the pathological sections
exhibited liver fibrosis and spleen trabecula fibrosis (Fig. 6Bb and c). In the CAG group, leukemic
cells infiltrated the hepatic portal and central veins (Fig. 6Cb) indicated by the arrows. In the
spleens, the boundary of red pulp widened and the white pulp was
clear (Fig. 6Cc).
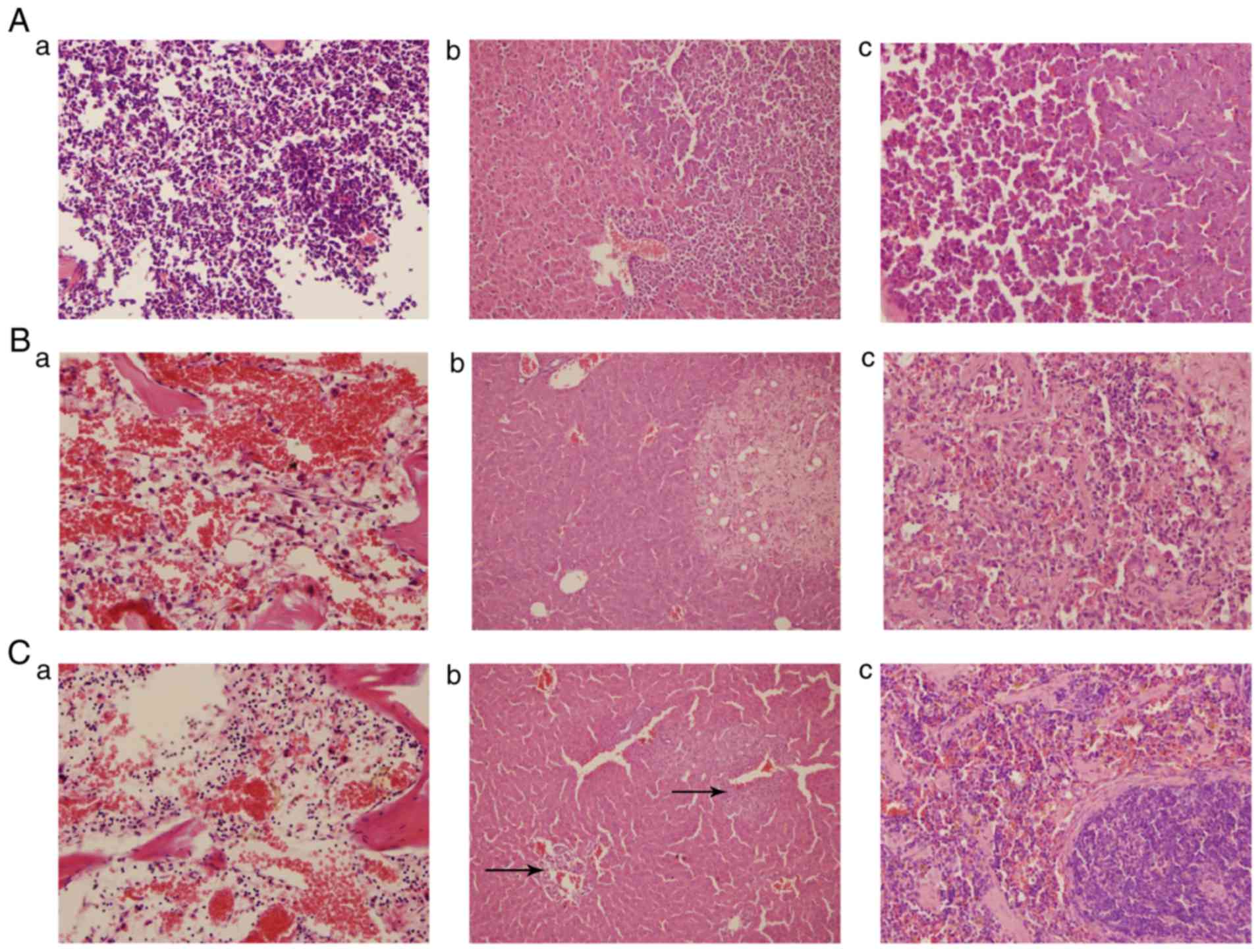 | Figure 6.H&E staining of the femur, liver
and spleen. (A) AML group: (a) femur, magnification, ×400; (b)
liver, magnification, ×200; (c) spleen, magnification, ×400. (B) DA
group: (a) femur, magnification, ×400; (b) liver, magnification,
×200; (c) spleen, magnification, ×400. (C) CAG group: (a) femur,
magnification, ×400; (b) liver, arrows indicate the infiltration of
leukemic cells into the hepatic portal and central veins;
magnification, ×200; (c) spleen, magnification, ×400. H&E,
hematoxylin and eosin; AML, acute myeloid leukemia; DA,
daunorubicin + cytarabine; CAG, cytarabine + aclarubicin +
granulocyte-colony stimulating factor. |
Discussion
G-CSF has been widely used in the treatment of
refractory and relapsed AML together with chemotherapeutic drugs,
and has improved the CR rate (13).
Although it is well-established that G-CSF has an important
influence on the BMM (22), there is
little research regarding the effect of G-CSF priming chemotherapy
on the BMM. G-CSF can promote leukemia cells to be released from
the bone marrow into the peripheral circulation by blocking the
interaction of SDF-1α and CXCR4, thus enhancing the anti-leukemia
effect (13). G-CSF also mobilizes
immunosuppressive Tregs from the bone marrow into the peripheral
blood (27). We hypothesize that
there is an effect of the G-CSF priming regimen on the BMM in
AML.
An AML-M4 BALB/c mouse model was established to
study the BMM in vivo in mice receiving different
chemotherapy regimens. Changes in the SDF-1α/CXCR4 axis, Tregs,
MDSCs and associated cytokines were investigated. In accordance
with previous studies, it was demonstrated that the CAG regimen had
a positive effect on the immunosuppressive microenvironment in
AML.
In the AML-M4 BALB/c mouse model, the number of
Tregs and MDSCs in the bone marrow decreased following CAG
chemotherapy. The concentrations of immunosuppressive cytokines
(TGF-β1, IL-10 and Arg-1), secreted by Tregs and MDSCs, were lower
in the CAG group compared with the DA group. The number of Tregs
and MDSCs in the spleen, the largest immune organ, was high. This
suggests that G-CSF may mobilize Tregs and MDSCs from the bone
marrow to the periphery, and that the G-CSF priming regimen may
relieve AML-associated BMM immune suppression by reducing the
number of MDSCs and Tregs in the bone marrow. This may help to
explain how G-CSF administration reduces the severity of acute
graft-vs.-host disease (GVHD) (27).
The BMM provides a protective niche for HSCs
(28) and it is the primary site of
minimal residual disease (MRD) following chemotherapy (39). By hijacking the HSC niche (40), leukemia stem cells (LSCs) in the bone
marrow niche serve roles in leukemia initiation, maintenance and
recurrence (41). G-CSF priming
chemotherapy treatment significantly increased LSC apoptosis and
increases responsiveness to chemotherapy in vivo by
modifying the functional behavior of LSCs (42). On the other hand, G-CSF inhibits the
interaction of SDF-1α and CXCR4, thus reducing LSC migration to the
BMM, which involves CXCR4 (43). In
the present study, it was also demonstrated that mice in the CAG
group expressed the lowest level of SDF-1α in the bone marrow and
peripheral blood, which indicates that the SDF-1α/CXCR4 axis may
have been downregulated in the CAG treatment regimen.
However, the CAG treatment regimen is limited in
that there were more residual leukemic cells subsequent to
treatment in the livers of the CAG mice than the DA mice. This may
result from the low dose of the chemotherapeutic agents, AraC and
ACR. However, the low doses used in the CAG regimen result in less
severe clinical side-effects than common induction chemotherapy,
such as the DA regimen. Due to its milder toxicity and a short
duration of G-CSF neutropenia treatment, the CAG regimen is
suitable for elderly patients (5,44,45).
The present study suggests that the CAG leukemia
treatment regimen has a positive effect on the immunosuppressive
microenvironment in AML and relieves AML-associated BMM immune
suppression by decreasing the number of Tregs and MDSCs in the bone
marrow. The results indicate that downregulation of the
SDF-1α/CXCR4 axis in the bone marrow and peripheral blood is caused
by CAG chemotherapy. The lower dose of chemotherapeutic agents in
the CAG regimen leads to milder side-effects but more residual
leukemic cells. Overall, the CAG regimen relieves immune
suppression in the leukemic BMM and is suitable for elderly
patients.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81270597).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JC performed the murine model and chemotherapy
regimens, performed the majority of the experiments and was a major
contributor in writing the manuscript. NY performed FCM, analyzed
the FCM data and helped in writing the manuscript. HL and HY
sacrificed all the mice and harvested the organs and pretreated
them. JW performed pathological examinations and acquired data of
these experiments. YY performed ELISA and acquired data of this
experiment. WZ designed the study, revised the manuscript and gave
final approval of the version to be published. All authors read and
approved the final manuscript
Ethics approval and consent to
participate
All animal experiments were reviewed and approved by
the Ethics Committee of the Medical College, Xi'an Jiaotong
University (Xi'an, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Podoltsev NA, Stahl M, Zeidan AM and Gore
SD: Selecting initial treatment of acute myeloid leukaemia in older
adults. Blood Rev. 31:43–62. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Döhner H, Weisdorf DJ and Bloomfield CD:
Acute myeloid leukemia. N Engl J Med. 373:1136–1152. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Döhner H, Estey EH, Amadori S, Appelbaum
FR, Büchner T, Burnett AK, Dombret H, Fenaux P, Grimwade D, Larson
RA, et al: Diagnosis and management of acute myeloid leukemia in
adults: Recommendations from an international expert panel, on
behalf of the European LeukemiaNet. Blood. 115:453–474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yamada K, Furusawa S, Saito K, Waga K,
Koike T, Arimura H, Aoyagi A, Yamato H, Sakuma H, Tsunogake S, et
al: Concurrent use of granulocyte colony-stimulating factor with
low-dose cytosine arabinoside and aclarubicin for previously
treated acute myelogenous leukemia: A pilot study. Leukemia.
9:10–14. 1995.PubMed/NCBI
|
|
5
|
Minakata D, Fujiwara S, Ito S, Mashima K,
Umino K, Nakano H, Kawasaki Y, Sugimoto M, Yamasaki R, Yamamoto C,
et al: A low-dose cytarabine, aclarubicin and granulocyte
colony-stimulating factor priming regimen versus a daunorubicin
plus cytarabine regimen as induction therapy for older patients
with acute myeloid leukemia: A propensity score analysis. Leuk Res.
42:82–87. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jin J, Chen J, Suo S, Qian W, Meng H, Mai
W, Tong H, Huang J, Yu W, Wei J and Lou Y: Low-dose cytarabine,
aclarubicin and granulocyte colony-stimulating factor priming
regimen versus idarubicin plus cytarabine regimen as induction
therapy for older patients with acute myeloid leukemia. Leuk
Lymphoma. 56:1691–1697. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang Y, Li W, Chen S, Qiu H, Sun A and Wu
D: Salvage chemotherapy with low-dose cytarabine and aclarubicin in
combination with granulocyte colony-stimulating factor priming in
patients with refractory or relapsed acute myeloid leukemia with
translocation (8;21). Leuk Res. 35:604–607. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu L, Qu Q, Jiao W, Zhang Y, Li X, Ding C
and Wu D: Increasing aclarubicin dose in low-dose cytarabine and
aclarubicin in combination with granulocyte colony-stimulating
factor (CAG regimen) is efficacious as salvage chemotherapy for
relapsed/refractory mixed-phenotype acute leukemia. Leuk Res.
39:805–811. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ma X and Wang J, Xu Y, Zhang W, Liu J, Cao
X, He A, Wang F, Gu L, Lei B and Wang J: Dose-enhanced combined
priming regimens for refractory acute myeloid leukemia and
middle-and-high-risk myelodysplastic syndrome: A single-center,
retrospective cohort study. Onco Targets Ther. 9:3661–3669. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Miyauchi J, Kelleher CA, Wang C, Minkin S
and McCulloch EA: Growth factors influence the sensitivity of
leukemic stem cells to cytosine arabinoside in culture. Blood.
73:1272–1278. 1989.PubMed/NCBI
|
|
11
|
Tafuri A and Andreeff M: Kinetic rationale
for cytokine-induced recruitment of myeloblastic leukemia followed
by cycle-specific chemotherapy in vitro. Leukemia. 4:826–834.
1990.PubMed/NCBI
|
|
12
|
te Boekhorst PA, Löwenberg B, Vlastuin M
and Sonneveld P: Enhanced chemosensitivity of clonogenic blasts
from patients with acute myeloid leukemia by G-CSF, IL-3 or GM-CSF
stimulation. Leukemia. 7:1191–1198. 1993.PubMed/NCBI
|
|
13
|
Shen ZH, Zeng DF, Ma YY, Zhang X, Zhang C
and Kong PY: Are there any new insights for G-CSF and/or AMD3100 in
chemotherapy of haematological malignants? Med Oncol. 32:2622015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim CH and Broxmeyer HE: In vitro behavior
of hematopoietic progenitor cells under the influence of
chemoattractants: Stromal cell-derived factor-1, steel factor, and
the bone marrow environment. Blood. 91:100–110. 1998.PubMed/NCBI
|
|
15
|
Jo DY, Rafii S, Hamada T and Moore MA:
Chemotaxis of primitive hematopoietic cells in response to stromal
cell-derived factor-1. J Clin Invest. 105:101–111. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wright DE, Bowman EP, Wagers AJ, Butcher
EC and Weissman IL: Hematopoietic stem cells are uniquely selective
in their migratory response to chemokines. J Exp Med.
195:1145–1154. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Peled A, Petit I, Kollet O, Magid M,
Ponomaryov T, Byk T, Nagler A, Ben-Hur H, Many A, Shultz L, et al:
Dependence of human stem cell engraftment and repopulation of
NOD/SCID mice on CXCR4. Science. 283:845–848. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lapidot T and Kollet O: The essential
roles of the chemokine SDF-1 and its receptor CXCR4 in human stem
cell homing and repopulation of transplanted immune-deficient
NOD/SCID and NOD/SCID/B2m(null) mice. Leukemia. 16:1992–2003. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Broxmeyer HE, Cooper S, Kohli L, Hangoc G,
Lee Y, Mantel C, Clapp DW and Kim CH: Transgenic expression of
stromal cell-derived factor-1/CXC chemokine ligand 12 enhances
myeloid progenitor cell survival/antiapoptosis in vitro in response
to growth factor withdrawal and enhances myelopoiesis in vivo. J
Immunol. 170:421–429. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Broxmeyer HE, Kohli L, Kim CH, Lee Y,
Mantel C, Cooper S, Hangoc G, Shaheen M, Li X and Clapp DW: Stromal
cell-derived factor-1/CXCL12 directly enhances
survival/antiapoptosis of myeloid progenitor cells through CXCR4
and G(alpha)i proteins and enhances engraftment of competitive,
repopulating stem cells. J Leukoc Biol. 73:630–638. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zeng Z, Shi YX, Samudio IJ, Wang RY, Ling
X, Frolova O, Levis M, Rubin JB, Negrin RR, Estey EH, et al:
Targeting the leukemia microenvironment by CXCR4 inhibition
overcomes resistance to kinase inhibitors and chemotherapy in AML.
Blood. 113:6215–6224. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Saba F, Soleimani M, Kaviani S, Abroun S,
Sayyadipoor F, Behrouz S and Saki N: G-CSF induces up-regulation of
CXCR4 expression in human hematopoietic stem cells by
beta-adrenergic agonist. Hematology. 2014.
|
|
23
|
De La Luz, Sierra M, Gasperini P,
McCormick PJ, Zhu J and Tosato G: Transcription factor Gfi-1
induced by G-CSF is a negative regulator of CXCR4 in myeloid cells.
Blood. 110:2276–2285. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jadidi-Niaragh F, Yousefi M, Memarian A,
Hojjat-Farsangi M, Khoshnoodi J, Razavi SM, Jeddi-Tehrani M and
Shokri F: Increased frequency of CD8+ and
CD4+ regulatory T cells in chronic lymphocytic leukemia:
Association with disease progression. Cancer Invest. 31:121–131.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Filipazzi P, Huber V and Rivoltini L:
Phenotype, function and clinical implications of myeloid-derived
suppressor cells in cancer patients. Cancer Immunol Immunother.
61:255–263. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Raggi C, Mousa HS, Corrent M, Sica A and
Invernizzi P: Cancer stem cells and tumor-associated macrophages: A
roadmap for multitargeting strategies. Oncogene. 35:671–682. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zou LH, Barnett B, Safah H, LaRussa VF,
Evdemon-Hogan M, Mottram P, Wei S, David O, Curiel TJ and Zou W:
Bone marrow is a reservoir for CD4+CD25+ regulatory T cells that
traffic through CXCL12/CXCR4 signals. Cancer Res. 64:8451–8455.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tabe Y and Konopleva M: Advances in
understanding the leukaemia microenvironment. Br J Haematol.
164:767–778. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kowanetz M, Wu X, Lee J, Tan M, Hagenbeek
T, Qu X, Yu L, Ross J, Korsisaari N, Cao T, et al:
Granulocyte-colony stimulating factor promotes lung metastasis
through mobilization of Ly6G+Ly6C+ granulocytes. Proc Natl Acad Sci
USA. 107:21248–21255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bayne LJ, Beatty GL, Jhala N, Clark CE,
Rhim AD, Stanger BZ and Vonderheide RH: Tumor-derived
granulocyte-macrophage colony-stimulating factor regulates myeloid
inflammation and T cell immunity in pancreatic cancer. Cancer Cell.
21:822–835. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kohanbash G, McKaveney K, Sakaki M, Ueda
R, Mintz AH, Amankulor N, Fujita M, Ohlfest JR and Okada H: GM-CSF
promotes the immunosuppressive activity of glioma-infiltrating
myeloid cells through interleukin-4 receptor-α. Cancer Res.
73:6413–6423. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lin JP, Yang JS, Lu CC, Chiang JH, Wu CL,
Lin JJ, Lin HL, Yang MD, Liu KC, Chiu TH and Chung JG: Rutin
inhibits the proliferation of murine leukemia WEHI-3 cells in vivo
and promotes immune response in vivo. Leuk Res. 33:823–828. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wen YF, Yang JS, Kuo SC, Hwang CS, Chung
JG, Wu HC, Huang WW, Jhan JH, Lin CM and Chen HJ: Investigation of
anti-leukemia molecular mechanism of ITR-284, a carboxamide analog,
in leukemia cells and its effects in WEHI-3 leukemia mice. Biochem
Pharmacol. 79:389–398. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mohan S, Abdul AB, Abdelwahab SI,
Al-Zubairi AS, Sukari Aspollah M, Abdullah R, Taha MM, Beng NK and
Isa NM: Typhonium flagelliforme inhibits the proliferation of
murine leukemia WEHI-3 cells in vitro and induces apoptosis in
vivo. Leuk Res. 34:1483–1492. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mills KH: Regulatory T cells: Friend or
foe in immunity to infection? Nat Rev Immunol. 4:841–855. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wan YY and Flavell RA: TGF-beta and
regulatory T cell in immunity and autoimmunity. J Clin Immunol.
28:647–659. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Romano A, Parrinello NL, Vetro C, Tibullo
D, Giallongo C, La Cava P, Chiarenza A, Motta G, Caruso AL, Villari
L, et al: The prognostic value of the myeloid-mediated
immunosuppression marker Arginase-1 in classic Hodgkin lymphoma.
Oncotarget. 7:67333–67346. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Munder M: Arginase: An emerging key player
in the mammalian immune system. Br J Pharmacol. 158:638–651. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Meads MB, Hazlehurst LA and Dalton WS: The
bone marrow microenvironment as a tumor sanctuary and contributor
to drug resistance. Clin Cancer Res. 14:2519–2526. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tavor S and Petit I: Can inhibition of the
SDF-1/CXCR4 axis eradicate acute leukemia? Semin Cancer Biol.
20:178–185. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ye M, Zhang H, Yang H, Koche R, Staber PB,
Cusan M, Levantini E, Welner RS, Bach CS, Zhang J, et al:
Hematopoietic differentiation is required for initiation of acute
myeloid leukemia. Cell Stem Cell. 17:611–623. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Saito Y, Uchida N, Tanaka S, Suzuki N,
Tomizawa-Murasawa M, Sone A, Najima Y, Takagi S, Aoki Y, Wake A, et
al: Induction of cell cycle entry eliminates human leukemia stem
cells in a mouse model of AML. Nat Biotechnol. 28:275–280. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Konopleva MY and Jordan CT: Leukemia stem
cells and microenvironment: Biology and therapeutic targeting. J
Clin Oncol. 29:591–599. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wei G, Ni W, Chiao JW, Cai Z, Huang H and
Liu D: A meta-analysis of CA G(cytarabine, aclarubicin, G-CSF)
regimen for the treatment of 1029 patients with acute myeloid
leukemia and myelodysplastic syndrome. J Hematol Oncol. 4:462011.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu L, Zhang Y, Jin Z, Zhang X, Zhao G, Si
Y, Lin G, Ma A, Sun Y, Wang L and Wu D: Increasing the dose
ofaclarubicin in low-dose cytarabine and aclarubicin in combination
with granulocyte colony-stimulating factor (CAG regimen) can safely
andeffectively treat relapsed or refractory acute myeloid leukemia.
Int J Hematol. 99:603–608. 2014. View Article : Google Scholar : PubMed/NCBI
|