Introduction
Hepatocellular carcinoma (HCC) is one of the most
common malignant tumors and can be divided into two major
categories: Primary and secondary. Primary liver malignancy is a
malignant tumor which life-threatening and has a high mortality
rate in China (1,2). At present, the major treatments for HCC
are surgical resection, chemotherapy, radiation therapy and gene
therapy (3,4). Although there are obvious improvements
in the diagnostic approach and treatment of HCC, the curative ratio
remains low. Thus, identifying treatment strategies and gaining an
understanding of the underlying mechanism of HCC progression is
required.
miRNAs inhibit the expression of protein coding
genes by partially binding the 3-UTR mRNA (5). Mounting evidence has shown that miRNA
maladjustment is involved in the progression of many tumor diseases
(6–8),
including breast, prostate and lung cancers as well as HCC. For
instance, miR-196b upregulated HCC cell invasion and migration by
binding to the 3-UTR of FOXP2 (9).
Xue and Tian concluded that the miR-429/RAB23 axis provided a
potential target for treating HCC (10). miR-3613 and miR-1271 affected cell
proliferation and cycle of HCC via their target genes (11,12).
The deregulated expression of miR-106b plays an
abnormal role in regulating the development of various cancers
(13,14). miR-106b was found to be expressed at a
lower level in osteosarcoma cells and target HMGA2 to inhibit the
cell progression (15). However,
Zhang et al provided evidence that miR-106b was upregulated
in colorectal cancer and promoted cell invasion and migration by
targeting DLC1 (16). Furthermore, a
recent study has shown that in the progression of cervical cancer,
DAB2 was confirmed as a target of miR-106b (17). Another recent study showed that
miRNA-106b expression was markedly increased and regulated HCC
development by targeting mRNA (18,19).
However, whether miR-106b targeted DAB2 in the regulation of HCC
progression has yet to be reported.
Disabled homolog 2 (DAB2) is a member of the disable
gene family. DAB2/DOC-2 has been proven to function as a new tumor
suppressor that plays an important role in the occurrence and
development of tumors (20,21). A previous study reported that DAB2 was
downregulated in ovarian cancer (21). Subsequently, a lower DAB2 expression
was detected and cell development was regulated in various types of
cancer, including breast (22),
prostate (23), and non-small lung
cancer (24), nasopharyngeal
(25) and esophageal squamous cell
carcinoma (26). Previous findings
have shown that DAB2 expression is decreased in HCC cells (27,28) and
regulates the progression of HCC. However, the biological mechanism
of DAB2 in HCC cells regulated by miR-106b has not been reported
yet.
In the present study, to the best of our knowledge,
we showed for the first time that, DAB2 acted as a specific target
of miR-106b and confirmed the promotion effect of miR-106b in
regulating HCC cell proliferation and migration. miR-18a was
overexpressed in HCC, whereas DAB2 was expressed at a lower level
in HCC and miR-106b expression was negatively associated with DAB2
expression. The data suggested that miR-106b may promote HCC cell
viability and migration via inhibiting DAB2. This mechanism
provides a therapeutic strategy for treating HCC.
Materials and methods
Specimens and cells culture
We collected 50 HCC samples from patients who
underwent complete surgery at The Third People's Hospital of
Qingdao (Qingdao, China) from July, 2013 to September, 2017. The
samples were immediately placed in a liquid nitrogen tank and
stored in a refrigerator at −80°C. Written informed consent was
signed by all the patients and the study was approved by the Ethics
Committee of The Third People's Hospital of Qingdao.
We purchased the HCC cell lines (Hep3B, Huh7 and
Bel-7402) and normal liver cell L02 from the Cell Bank of Chinese
Academy of Sciences (Shanghai, China). The cell lines were cultured
in Dulbecco's modified Eagle's medium (DMEM; Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) containing 10% fetal
bovine serum, penicillin (100 U/ml) and streptomycin (100 µg/ml).
Subsequently, the cells were maintained in an incubator at 37°C
under a 5% CO2 atmosphere.
qPCR
Total RNA was isolated from HCC tissues and cells by
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
miR-106b expression was detected by miRNA First-Strand Synthesis
and miRNA Quantitation kits (Takara Biotechnology Co., Ltd.,
Dalian, China). DAB2 was examined by CellAmp™ Direct Prep kit for
RT-PCR and Protein Analysis (Takara Biotechnology Co., Ltd.).
Primer sequences used were: miR-106b-F: CTTCCTGTCATAAAGTGCTGAC
AGTGCAGATCTGCAGTCTGGAGTTTCA, miR-106b-R:
TGACAGGAAGTAAAGTGCTGACAGTGCAGATCGAGA TCTTGGGCCTCT. DAB2-F:
GTAGAAACAAGTGCAACC AATGG, DAB2-R: GCCTTTGAACCTTGCTAAGAGA. U6-F:
CTCGCTTCGGCAGCACA. U6-R: AACGCTTCAC GAATTTGCGT. GAPDH-F:
TGGTATCGTGGAAGGA CTC, GAPDH-R: AGTAGAGGCAGGGATGATG. U6 and GAPDH
were used as the internal references to standardize the miR-106b
and DAB2 relative expression respectively. The 2−ΔΔCq
method was used to detect the differential expression of miR-106b
and DAB2.
Western blot analysis
After transfection for 48 h, RIPA lysis containing
proteinase inhibitors (Beyotime Institute of Biotechnology, Haimen,
China) and phenylmethylsulfonyl fluoride were used to extract the
total protein from HCC cells. The protein concentrations were
tested with the BCA protein assay kit (Beyotime Institute of
Biotechnology). The total proteins (50 µg) were added into the
SDS-PAGE gels and performed electrophoresis at 60 V when
bromophenol blue ran out from the bottom. The proteins were then
transferred to nitrocellulose filter (NC) membranes. Then, skimmed
milk (5-10%) was used to block the proteins on the membranes at
room temperature for 2 h. Subsequently, the membranes were
incubated with the primary antibody: Rabbit polyclonal anti-DAB2
(ab76253; 1:1,000; Abcam, Cambridge, MA, USA) were added in to
incubate the samples at 4°C, and then horseradish
peroxidase-conjugated (HRP, 1:10,000). GAPDH primary antibody
(5174P; 1:5,000; Cell Signaling Technology, Inc., Danvers, MA, USA)
was chosen as the internal reference. After being washed three
times with 1X TBST, they were incubated with secondary antibody
goat anti-rabbit IgG-HRP (sc-2,004; 1:3,000; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) at room temperature for 2
h. Protein bands were detected using the chemiluminescence method
(ECL; Millipore, Billerica, MA, USA).
Cell transfection
We purchased miR-106b mimic and inhibitor from
GenePharma Co., Ltd. (Suzhou, China) and DAB2 vector from Shanghai
Genechem Co., Ltd. (Shanghai, China). Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) was used to transfect
miR-106b mimic, miR-106b inhibitor, DAB2 vector or both DAB2 vector
and miR-106b mimic, respectively, into Hep3B cells following the
manufacturers protocol.
MTT assay
An MTT assay was carried out to examine cell
viability to determine HCC cell proliferation. The cells
(5×103 cells/ml) treated with different transfection
were planted in 96-well plates and incubated for 48 h at 37°C with
5% CO2. Then, we added MTT reagent (20 ml) to each well
at 0, 1, 2, 3 and 4 days followed by incubation for another 4 h.
Dimethyl sulfoxide (150 ml) was added to dissolve the
crystallization. The absorbance value of cells was measured at 490
nm using enzyme-linked immunoassay.
Transwell assay
Cell migration was performed using the Transwell
assay. A Transwell chamber with 8 µm pore size polycarbonic
membrane (CoStar Group, Inc., Washington, DC, USA) was placed into
the 24-well plates to separate the upper and lower chambers. After
transfection for 48 h, the cells were added into the upper chamber
coated with gelatin and DMEM medium (600 ml) supplemented with 10%
fetal bovine serum was seeded in the lower chamber. Then, the cells
in each well were incubated at 37°C for 24 h. Cells migrating from
the upper to the lower chamber were fixed with 90% ethanol. Then,
0.05% crystal violet was added to stain the migrated cells for 15
min. Finally, cotton swab was used to gently scrape off the cells
that did not migrate. Images of the migration cells were captured
under a microscope (BX53; Olympus Corporation, Tokyo, Japan).
Luciferase assay
Relative luciferase ability was performed using the
recombinant pMIR-reporter luciferase vector (Guangzhou RiboBio Co.,
Ltd., Guangzhou, China). A site-directed mutagenesis kit (cat. no.
210518; Agilent Technologies, Inc., Santa Clara, CA, USA) was
carried out to introduce the site-directed mutagenesis into the
miR-106b binding site of DAB2 mRNA. The wild-type and mut-type
miR-106b putative targets on DAB2 3-UTR mRNA were cloned into the
downstream of pGL3 luciferase vector (Promega Corporation, Madison,
WI, USA). The HCC cells were transfected with miR-106b mimic using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
The Dual Luciferase Assay (Promega Corporation) was subsequently
used to analyze the luciferase activity values.
Statistical analysis
All the experiments were repeated in triplicate.
Data are expressed as mean ± SD. SPSS 19.0 software (SPSS, Inc.,
Chicago, IL, USA). One-way ANOVA was used to perform the
statistical analyses. The data were evaluated using the Students
t-test or Tukeys post-hoc test, with statistically significant
difference considered as P<0.05. Correlation between mRNA and
miRNA was estimated using the Spearmans correlation method.
Results
miR-106b is overexpressed in HCC
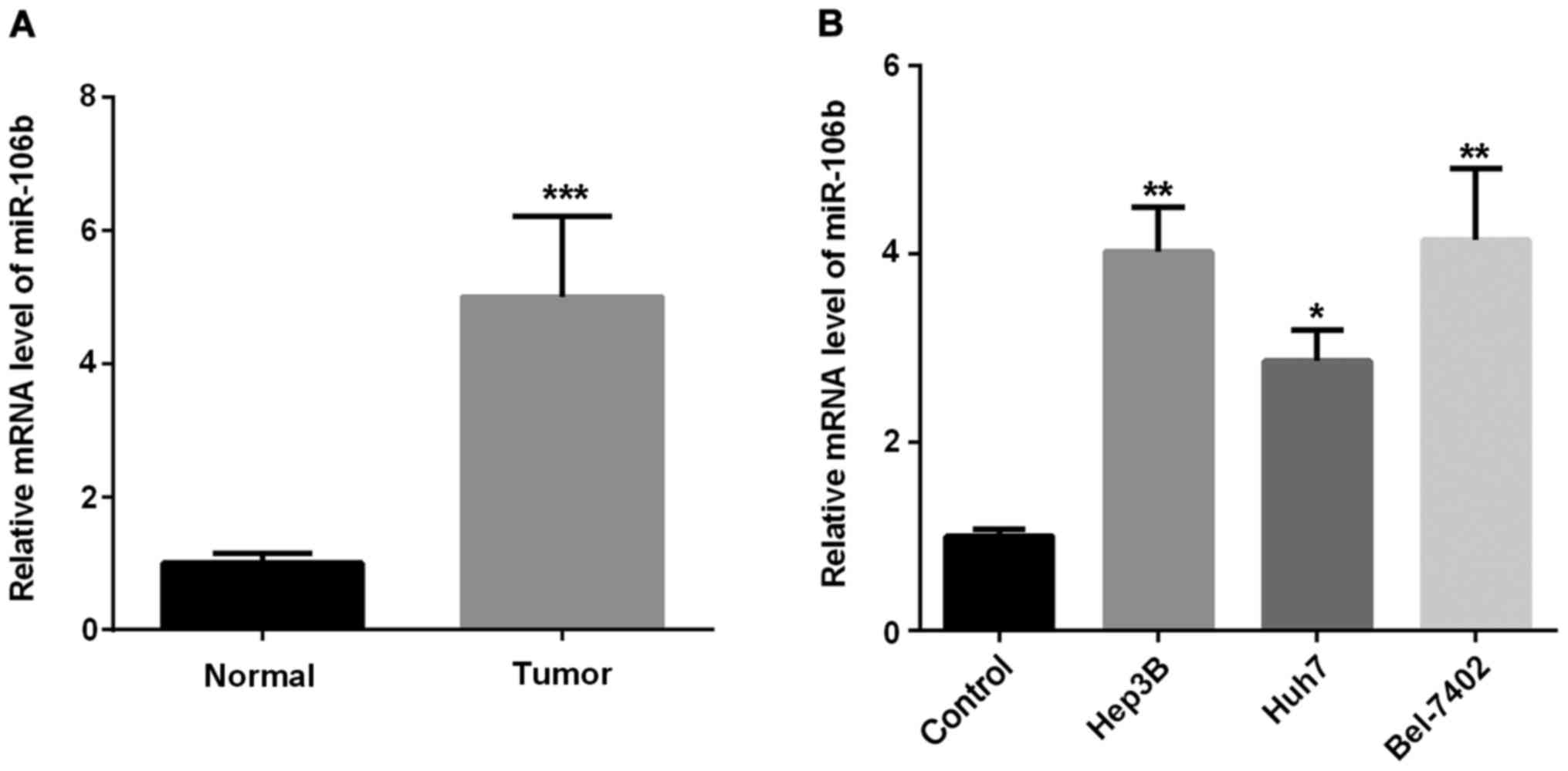
To understand the role miR-106b played in HCC
progression, we investigated miR-106b expression in HCC tissues and
cells. Firstly, we examined the expression of miR-106b in HCC
tissues. As shown in Fig. 1A, miR-106
expression was markedly higher in HCC tissues than that in the
adjacent normal tissues. Secondly, we examined miR-106b expression
in three HCC cell lines. As shown in Fig.
1B, miR-106b expression in HCC cells increased in different
degrees compared with the normal cells. Thus, miR-106b may function
as a tumor promoter in regulating HCC development.
miR-106b promotes HCC cell
proliferation and migration
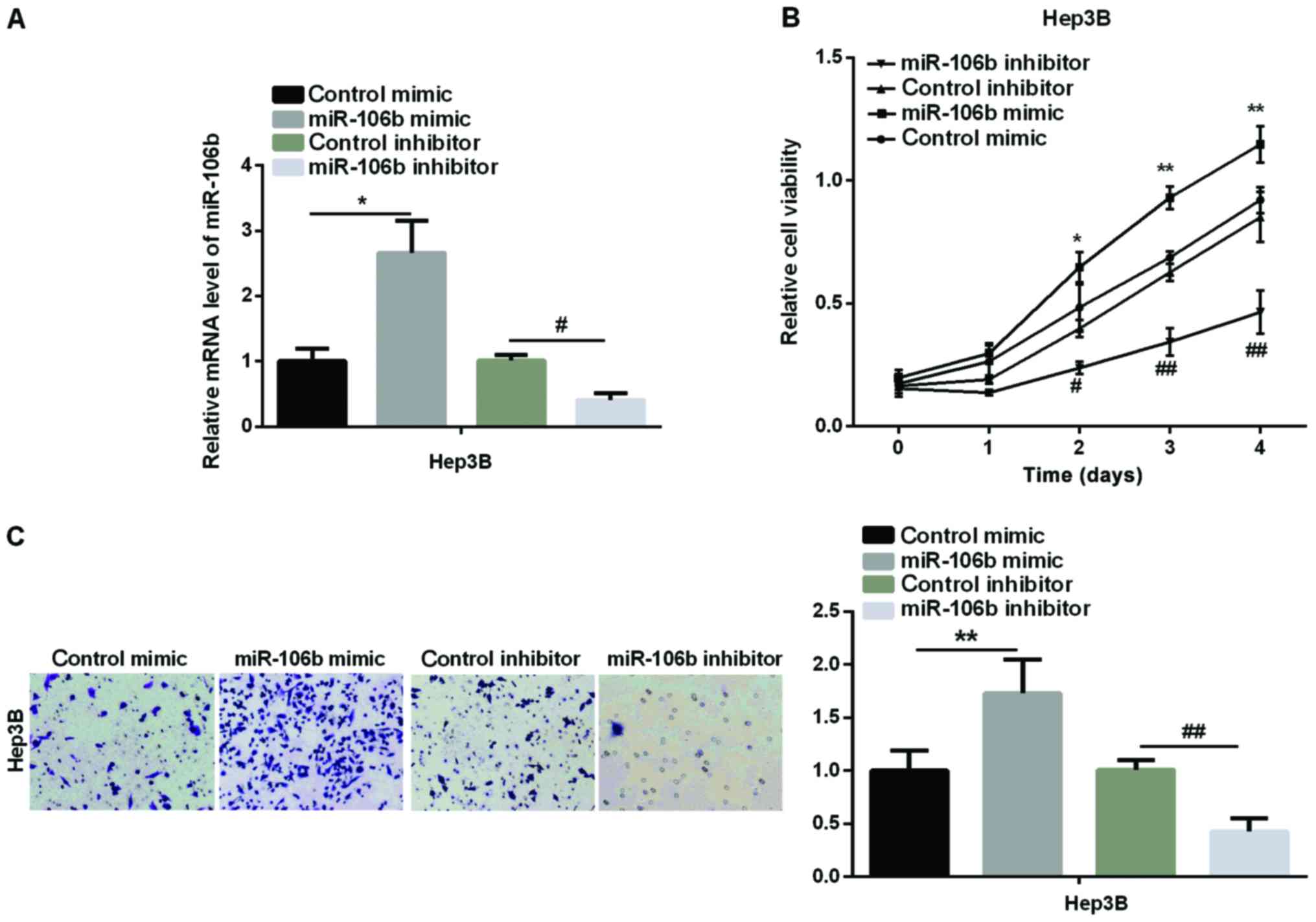
We then examined the effect of miR-106b on HCC cell
proliferation and migration. MTT and Transwell assays were
performed to test the cell viability and relative cell migration in
an HCC cell line (Hep3B). Firstly, we transfected Hep3B with
miR-106b mimic or miR-106b inhibitor to overexpress or silence
miR-106b. The transfection efficiency was detected by qPCR. As
shown in Fig. 2A, miR-106b expression
increased markedly in the miR-106b mimic group, but significantly
decreased in the miR-106b inhibitor group. Secondly, MTT assay
showed that the relative cell viability was obviously higher in
Hep3B after treatment with miR-106b mimic, whereas it was lower in
miR-106b inhibitor than in the control group (Fig. 2B). Finally, we used Transwell assay to
detect the change in HCC cell migration. The results in Fig. 2C revealed that relative cell migration
was significantly increased after the overexpression of miR-106b,
while it was inhibited by miR-106b silencing. The above results
clearly showed that miR-106b promotes the HCC development by
enhancing cell proliferation and migration due to its high-level
expression in HCC.
miR-106b specifically targets DAB2 in
HCC
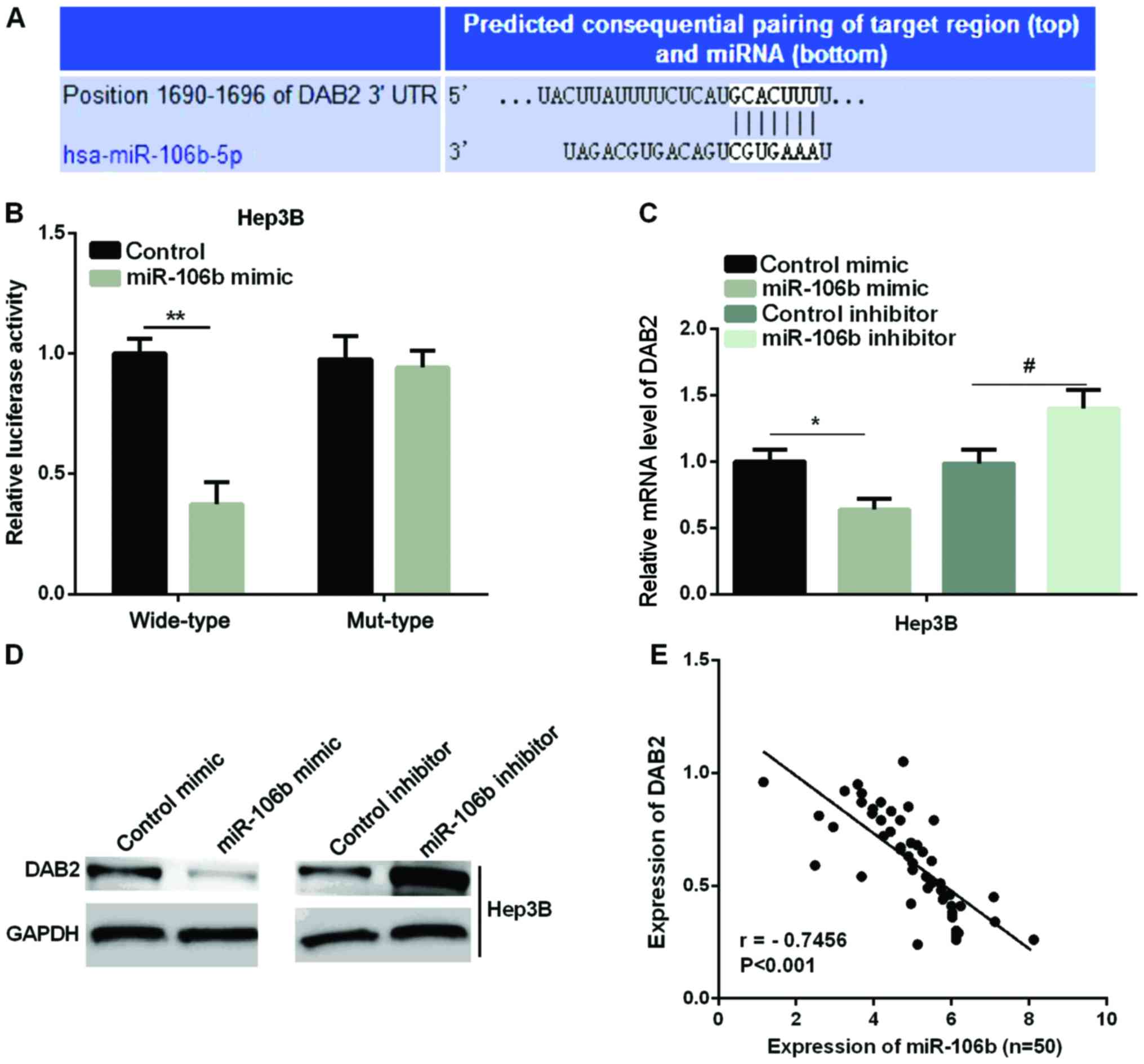
Previous studies have confirmed that DAB2 acts as a
tumor suppressor in HCC (28). We
predicted that DAB2 was a target of miR-106b in the regulation of
HCC cells. We first used TargetScanHuman 7.1 to validate this
prediction. The binding sites of DAB2 and miR-106b are shown in
Fig. 3A. A luciferase reporter assay
was to detect the luciferase ability in two HCC cell lines to
further determine the accuracy of this prediction. It was found
that the luciferase activity in the miR-106b mimic group was
significantly reduced compared with the control group in wild-type,
whereas there were no effects in mut-type in Hep3B (Fig. 3B). Furthermore, we examined the mRNA
and protein expression of DAB2 after the overexpression or
knockdown of miR-106b in the Hep3B cell line. As shown in Fig. 3C and D, both the miR-106b mRNA and
protein level were significantly reduced by the miR-106b mimic,
while it was increased by miR-106b inhibitor in Hep3B cell line.
Regression analysis showed that DAB2 expression and miR-106b
expression were negatively correlated in HCC tissues (Fig. 3E).
miR-106b upregulates HCC progression
by targeting DAB2
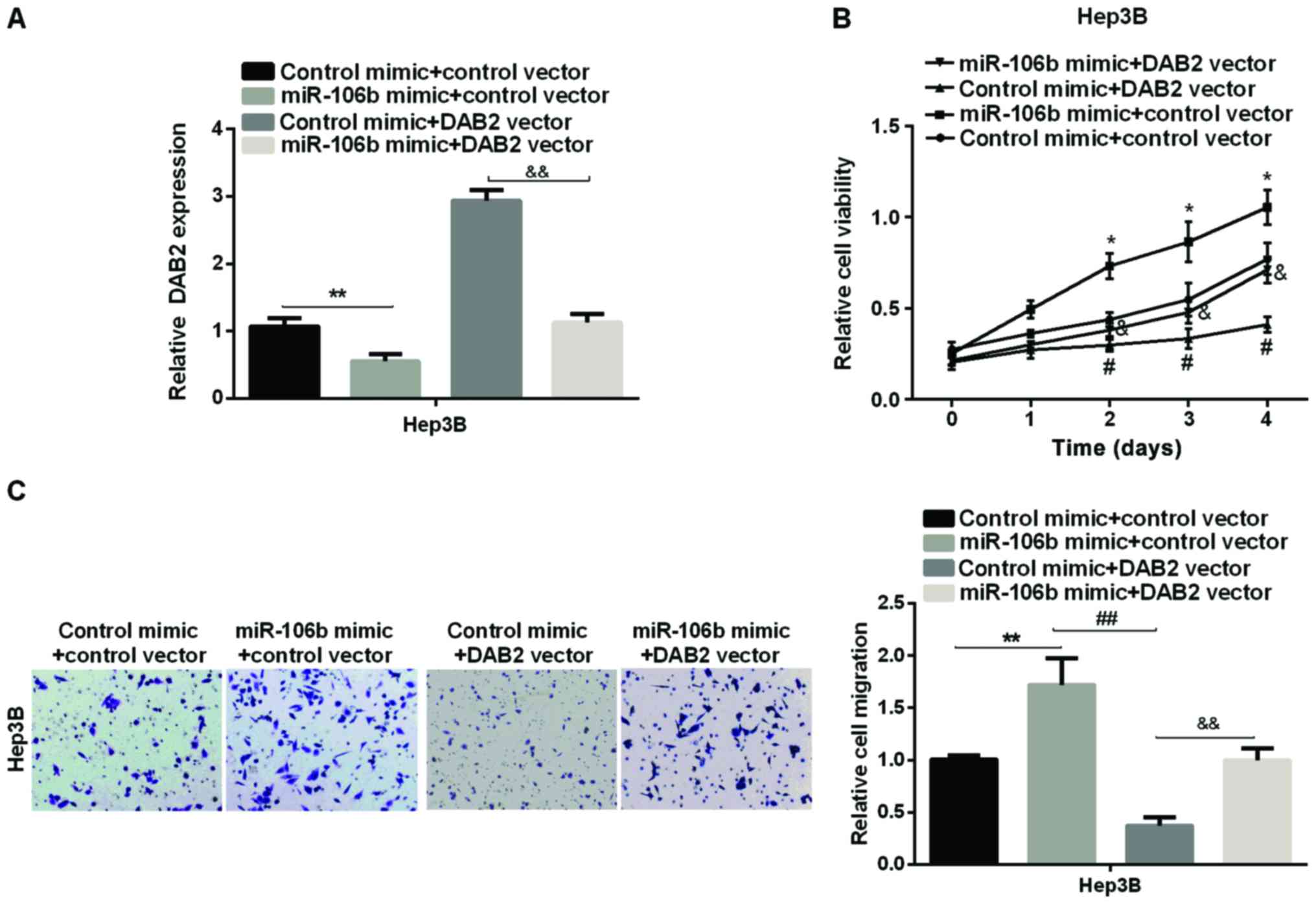
We first used the DAB2 vector to overexpress DAB2 in
Hep3B cell line (Fig. 4A) to
investigate whether DAB2 is the downstream mediator of miR-106b in
promoting HCC proliferation and migration. Subsequently, the effect
of DAB2 on relative cell viability and cell migration regulated by
miR-106 was investigated by MTT and Transwell assay. The results
showed that the overexpression of DAB2 reduced viability and
migration of HCC cells, and DAB2 may reverse the miR-106b-promoting
effect on HCC cell proliferation (Fig.
4B). The relative cell migration of HCC, which was increased by
miR-106b, was supressed by DAB2 re-expression in Hep3B cell line
(Fig. 4C). The data suggested that
miR-106b promoted HCC cell proliferation and migration via
targeting DAB2.
Discussion
Many studies have reported that the miRNAs were
abnormally expressed in HCC patients, such as miR-196b, miR-429,
miR-3613 and miR-1271 (9–12). Although the abnormal expression of
miRNAs in HCC was regarded as a cause of HCC, we need to further
explore the potential mechanism of the impact of miRNA on the
progression of HCC. Previous findings showed that miRNA-106b
expression is obviously increased in HCC and may provide a new
biomarker for the early diagnosis of HCC (18). Those findings are in line with those
of our study showing that miR-106b was overexpressed in HCC tissues
and cells. However, the underlying mechanism of miR-106 in
regulating HCC progression remains unclear.
Mounting evidence indicates that malignant tumor
cell proliferation, migration and invasion are the main causes of
human tumors (29). Previous findings
have shown that the different expression of miRNAs has different
effects on HCC cell development. It was proven that miR-765 mimic
may make the number of HCC cells increase by regulating INPP4B
(30). However, miR-340 may suppress
HCC cell proliferation by upregulating JAK1 as its expression was
downregulated in HCC (31). In the
present study, we revealed that the higher expression of miR-106
promoted HCC cell proliferation and migration, whereas the lower
expression of miR-106 repressed HCC cell development.
DAB2 is involved in the development of multiple
cancers, including prostate, non-small lung and breast cancers
(22–24). In HCC, a lower expression of DAB2 was
detected in HCC and inhibiting DAB2 may promote HCC cell
proliferation and invasion (28).
Furthermore, loss of DAB2 induced a significantly higher apoptosis
in HCC (32). Findings of those
studies are in agreement with our results, which showed DAB2
expression was lower in HCC and upregulation of DAB2 inhibited the
suppression effect on HCC cell proliferation and migration
ability.
In summary, miR-106b expression was higher while
that of DAB2 was lower in HCC and they were negatively correlated.
To the best of our knowledge, these are the first findings that
DAB2 was a direct target of miR-106b in regulating the progression
of HCC and DAB2 may partially reverse the promotion effect of
miR-106b in HCC, indicating that the miR-106b/DAB2 axis has a
potential for use in HCC diagnosis and therapy.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors contributions
CS conceived and designed this study. XY and QJ
performed the assay and interpreted the results. XS was involved in
drafting the manuscript and revising it critically for important
intellectual content. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
The Third People's Hospital of Qingdao (Qingdao, China). Patients
who participated in this study had complete clinical data. Signed
written informed consents were obtained from the patients and/or
guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Inokawa Y, Inaoka K, Sonohara F, Hayashi
M, Kanda M and Nomoto S: Molecular alterations in the
carcinogenesis and progression of hepatocellular carcinoma: Tumor
factors and background liver factors. Oncol Lett. 12:3662–3668.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fernández-Rodríguez CM and
Gutiérrez-García ML: Prevention of hepatocellular carcinoma in
patients with chronic hepatitis B. World J Gastrointest Pharmacol
Ther. 5:175–182. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li W, Man W, Guo H and Yang P: Clinical
study of transcatheter arterial chemoembolization combined with
microwave ablation in the treatment of advanced hepatocellular
carcinoma. J Cancer Res Ther. 12 Supplement:C217–C220. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Buendia MA and Neuveut C: Hepatocellular
carcinoma. Cold Spring Harb Perspect Med. 5:a0214442015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Di Leva G, Garofalo M and Croce CM:
MicroRNAs in cancer. Annu Rev Pathol. 9:287–314. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang P, Zou F, Zhang X, Li H, Dulak A,
Tomko RJ Jr, Lazo JS, Wang Z, Zhang L and Yu J: microRNA-21
negatively regulates Cdc25A and cell cycle progression in colon
cancer cells. Cancer Res. 69:8157–8165. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bulkowska M, Rybicka A, Senses KM, Ulewicz
K, Witt K, Szymanska J, Taciak B, Klopfleisch R, Hellmén E, Dolka
I, et al: MicroRNA expression patterns in canine mammary cancer
show significant differences between metastatic and non-metastatic
tumours. BMC Cancer. 17:7282017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu Z, Lin X, Tian M and Chang W: microRNA
196b promotes cell migration and invasion by targeting FOXP2 in
hepatocellular carcinoma. Oncol Rep. 39:731–738. 2018.PubMed/NCBI
|
|
10
|
Xue H and Tian GY: MiR-429 regulates the
metastasis and EMT of HCC cells through targeting RAB23. Arch
Biochem Biophys. 637:48–55. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qin A, Zhu J, Liu X, Zeng D, Gu M and Lv
C: MicroRNA-1271 inhibits cellular proliferation of hepatocellular
carcinoma. Oncol Lett. 14:6783–6788. 2017.PubMed/NCBI
|
|
12
|
Zhang D, Liu E, Kang J, Yang X and Liu H:
MiR-3613-3p affects cell proliferation and cell cycle in
hepatocellular carcinoma. Oncotarget. 8:93014–93028.
2017.PubMed/NCBI
|
|
13
|
Yu S, Qin X, Chen T, Zhou L, Xu X and Feng
J: MicroRNA-106b-5p regulates cisplatin chemosensitivity by
targeting polycystic kidney disease-2 in non-small-cell lung
cancer. Anticancer Drugs. 28:852–860. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Arias Sosa LA, Cuspoca Orduz AF and Bernal
Gómez BM: Deregulation of microRNAs in gastric cancer: Up
regulation by miR-21 and miR-106. Rev Gastroenterol Peru. 37:65–70.
2017.(In Spanish). PubMed/NCBI
|
|
15
|
He QY, Wang GC, Zhang H, Tong DK, Ding C,
Liu K, Ji F, Zhu X and Yang S: miR-106a-5p suppresses the
proliferation, migration, and invasion of osteosarcoma cells by
targeting HMGA2. DNA Cell Biol. 35:506–520. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang GJ, Li JS, Zhou H, Xiao HX, Li Y and
Zhou T: MicroRNA-106b promotes colorectal cancer cell migration and
invasion by directly targeting DLC1. J Exp Clin Cancer Res.
34:732015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Piao J, You K, Guo Y, Zhang Y, Li Z and
Geng L: Substrate stiffness affects epithelial-mesenchymal
transition of cervical cancer cells through miR-106b and its target
protein DAB2. Int J Oncol. 50:2033–2042. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Samal J, Kandpal M and Vivekanandan P:
HBeAg-induced miR-106b promotes cell growth by targeting the
retinoblastoma gene. Sci Rep. 7:143712017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu C, Shi L, Chen W, Fang P, Li J, Jin L,
Pan Z and Pan C: MiR-106b inhibitors sensitize TRAIL-induced
apoptosis in hepatocellular carcinoma through increase of death
receptor 4. Oncotarget. 8:41921–41931. 2017.PubMed/NCBI
|
|
20
|
Albertsen HM, Smith SA, Melis R, Williams
B, Holik P, Stevens J and White R: Sequence, genomic structure, and
chromosomal assignment of human DOC-2. Genomics. 33:207–213. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mok SC, Chan WY, Wong KK, Cheung KK, Lau
CC, Ng SW, Baldini A, Colitti CV, Rock CO and Berkowitz RS: DOC-2,
a candidate tumor suppressor gene in human epithelial ovarian
cancer. Oncogene. 16:2381–2387. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu S, Zhu J and Wu Z: Loss of Dab2
expression in breast cancer cells impairs their ability to deplete
TGF-β and induce Tregs development via TGF-β. PLoS One.
9:e917092014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xie Y, Zhang Y, Jiang L, Zhang M, Chen Z,
Liu D and Huang Q: Disabled homolog 2 is required for migration and
invasion of prostate cancer cells. Front Med. 9:312–321. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li C, Chen J, Chen T, Xu Z, Xu C, Ding C,
Wang Y, Lei Z, Zhang HT and Zhao J: Aberrant Hypermethylation at
Sites −86 to 226 of DAB2 Gene in Non-Small Cell Lung Cancer. Am J
Med Sci. 349:425–431. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu YF, Mao YP, Li YQ, Ren XY, He QM, Tang
XR, Sun Y, Liu N and Ma J: MicroRNA-93 promotes cell growth and
invasion in nasopharyngeal carcinoma by targeting disabled
homolog-2. Cancer Lett. 363:146–155. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li C, Ding C, Chen T, Chen J, Xu Z, Lei Z,
Xu C and Zhao J: Micro ribonucleic acid-93 promotes proliferation
and migration of esophageal squamous cell carcinoma by targeting
disabled 2. Thorac Cancer. 6:524–533. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Calvisi DF, Ladu S, Gorden A, Farina M,
Lee JS, Conner EA, Schroeder I, Factor VM and Thorgeirsson SS:
Mechanistic and prognostic significance of aberrant methylation in
the molecular pathogenesis of human hepatocellular carcinoma. J
Clin Invest. 117:2713–2722. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang X, Li N, Li X, Zhao W, Qiao Y, Liang
L and Ding Y: Low expression of DAB2IP contributes to malignant
development and poor prognosis in hepatocellular carcinoma. J
Gastroenterol Hepatol. 27:1117–1125. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xie BH, He X, Hua RX, Zhang B, Tan GS,
Xiong SQ, Liu LS, Chen W, Yang JY, Wang XN, et al: Mir-765 promotes
cell proliferation by downregulating INPP4B expression in human
hepatocellular carcinoma. Cancer Biomark. 16:405–413. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yuan J, Ji H, Xiao F, Lin Z, Zhao X, Wang
Z, Zhao J and Lu J: MicroRNA-340 inhibits the proliferation and
invasion of hepatocellular carcinoma cells by targeting JAK1.
Biochem Biophys Res Commun. 483:578–584. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Calvisi DF, Pinna F, Pellegrino R, Sanna
V, Sini M, Daino L, Simile MM, De Miglio MR, Frau M, Tomasi ML, et
al: Ras-driven proliferation and apoptosis signaling during rat
liver carcinogenesis is under genetic control. Int J Cancer.
123:2057–2064. 2008. View Article : Google Scholar : PubMed/NCBI
|