Introduction
Epithelial ovarian cancer (EOC) is the most common
histological type of ovarian cancer, and is the gynecological
malignancy with the highest mortality rate. It was estimated that
globally, 21,980 new cases and 14,270 mortalities occurred in 2014
as a consequence of EOC (1). The main
reasons for the high mortality rate include a lack of diagnostic
methods appropriate for early stage detection and a lack of
effective treatment strategies (2).
Therefore, understanding the molecular pathogenesis and uncovering
the associated molecular biomarkers of EOC may facilitate earlier
detection and improve the survival of patients with ovarian
cancer.
In addition to serving a crucial function in the
regression of the Müllerian duct and the regulation of
folliculogenesis (3,4), the anti-Müllerian hormone (AMH) has been
previously studied for the potential expansion of its application
against ovarian cancer. Accumulating evidence has demonstrated that
AMH exhibits an inhibitory effect on the proliferation, invasion,
metastasis and drug resistance of ovarian cancer in vitro
and in vivo, through binding to its specific receptor, named
AMH type II receptor (AMHRII) (5,6). In
particular, ovarian cancer stem cells, which facilitate resistance
to chemotherapeutic agents in addition to serving an integral
function in tumor recurrence, are inhibited by AMH (7–9). In order
to examine its toxicity, AMH was tested in vivo using female
nude mice for a total of 20 weeks, and significant inhibition of
tumor growth was observed without notable toxicity (10). Therefore, AMH may be a promising
anticancer drug for this gynecological malignancy.
An increasing number of studies have confirmed that
stem cell factor (SCF) serves a key function in tumor occurrence,
development, migration and recurrence, through activating
downstream signaling molecules, following interaction with its
receptor, c-kit (11–14). SCF interacting with c-kit via
autocrine or paracrine signaling has been identified in the
majority of ovarian cancer types, and results in the enhanced
proliferation, invasion, anti-apoptosis and drug resistance of
cancer cells (12–14). In addition, with respect to the
development of follicles, AMH was demonstrated to be a modulatory
factor that decreases the expression of SCF via the cyclic
adenosine monophosphate (cAMP)/protein kinase A (PKA) pathway
(15).
The underlying mechanism of AMH-mediated growth
inhibition in EOC remains unclear. The present study was performed
to investigate if AMH inhibits proliferation and induces apoptosis
in EOC cells via regulating the cell cycle and the downregulation
of SCF.
Materials and methods
Cell culture
Human EOC cell lines (OVCAR3 and OVCAR8) purchased
from the Institute of Biochemistry and Cell Biology at the Chinese
Academy of Sciences (Shanghai, China) were cultured in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(Invitrogen; Thermo Fisher Scientific, Inc.). No antibiotics were
used. The cells were incubated in a humidified atmosphere
containing 5% CO2 at 37°C. Recombinant human AMH (rhAMH;
R&D Systems, Inc., Minneapolis, MN, USA) was diluted with 10%
fetal bovine serum to 10 µg/ml for subsequent experiments.
Cell proliferation assay
An MTT assay was used to detect cell proliferation.
Briefly, EOC OVCAR3 and OVCAR8 cell lines (1,500 cells/well) were
seeded into 96-well plates, incubated in a humidified atmosphere
containing 5% CO2 at 37°C overnight to allow for cell
attachment and recovery, and then exposed to 10 µg/ml rhAMH, or
fetal bovine serum without rhAMH. The cells were cultured for the
indicated intervals (48 and 72 h) and then stained with sterile MTT
dye (0.5 mg/ml; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany).
After 4 h of incubation at 37°C, the supernatant was aspirated, and
dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA) was added to
dissolve the purple formazan. The absorbance was measured at 490
nm. All experiments were performed in triplicate.
Cell apoptosis detection
Two EOC OVCAR3 and OVCAR8 cell lines in each group
were seeded into 6-well plates at a density of 1×106/ml.
After 48 and 72 h of exposure to rhAMH at 37°C, cells were
harvested and washed twice with pre-chilled PBS followed by
centrifugation (100 × g) at 4°C for 5 min. Approximately
5×105 cells were collected. For apoptosis analysis, a
Dead Cell Apoptosis kit with Annexin V/fluorescein isothiocyanate
(FITC) and propidium iodide (PI) for flow cytometry (BD
Biosciences, Franklin Lakes, NJ, USA) was used according to the
manufacturer's protocol. Briefly, following washing twice with PBS,
cells were re-suspended in 1× Annexing binding buffer at a density
of 1×106 cells/ml. Then, 5 µl Annexin V/FITC and 1 µl
100 µg/ml PI were added to each 100 µl cell suspension, and the
samples were incubated at room temperature for 15 min in the dark.
Following incubation, apoptosis was analyzed using
fluorescence-activated cell sorting (FACS; BD Biosciences, Franklin
Lakes, NJ, USA) using Cell-Quest software version 5.1 (BD
Biosciences) within 1 h. The cells undergoing apoptosis were those
that were Annexin V FITC-positive and PI-negative. All experiments
were repeated three times.
Cell cycle analysis
Two EOC OVCAR3 and OVCAR8 cell lines in each group
were seeded in six-well plates with 1×106/ml. After 48
or 72 h exposure to rhAMH at 37°C, cells were harvested and washed
twice with pre-chilled PBS, then fixed in pre-chilled 70% ethanol
at 4°C overnight. The fixed cells were washed with 1 ml PBS, 100
µg/ml RNase A was then added, and then incubated at 37°C for 30
min. Approximately 20,000-30,000 cells were counted using a flow
cytometer (BD Biosciences), detecting red fluorescence at an
excitation wavelength of 488 nm.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA from two EOC OVCAR3 and OVCAR8 cell lines
was extracted using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). RT-qPCR was performed using the Moloney Murine
Leukemia Virus reverse transcriptase (Promega Corporation, Madison,
WI, USA) and platinum SYBR Green qPCR SuperMix-UDG reagents
(Invitrogen; Thermo Fisher Scientific, Inc.). Primers were designed
for human AMHRII (forward, 5′-TCCCAAGGCCAATATAAACC-3′ and reverse,
5′-TTATCCAGAGAACTCACTTCCA-3′), human SCF (forward,
5′-TCCACCATGGGAGAGCCTAA-3′ and reverse,
5′-AGTAACCTAGTTTTCAACACCCA-3′) and β-actin (forward,
5′-CTCTGGCCGTACCACTGGC-3′ and reverse, 5′-GTGAAGCTGTAGCCGCGC-3′).
β-actin was used as the internal control. qPCR was performed with
an initial denaturation step at 95°C for 30 sec, followed by 40
cycles of 95°C for 5 sec and 60°C for 40 sec. The PCR products were
run on a 1% agarose Tris-acetate-EDTA gel. The relative fold
changes in mRNA expression were calculated using the
2−ΔΔCq method (16).
ELISA
The protein levels of SCF were detected in OVCAR8
cell line culture supernatants and tumor homogenate using a Human
SCF ELISA kit (DCK00; R&D Systems, Inc.) according to the
manufacturer's protocol. All samples were assayed in
triplicate.
Statistical analysis
Measurement data were expressed as the mean ±
standard deviation. Comparisons between two independent samples
were performed using a Student's t-test. Additionally, one-way
analysis of variance followed by Fisher's least significant
difference test was applied for comparisons among multiple groups.
SPSS v.16.0 statistical software (SPSS, Inc., Chicago, IL, USA) was
used for statistical analysis. P<0.05 was considered to indicate
a statistically significant difference.
Results
AMHRII detected by RT-qPCR in EOC cell
lines
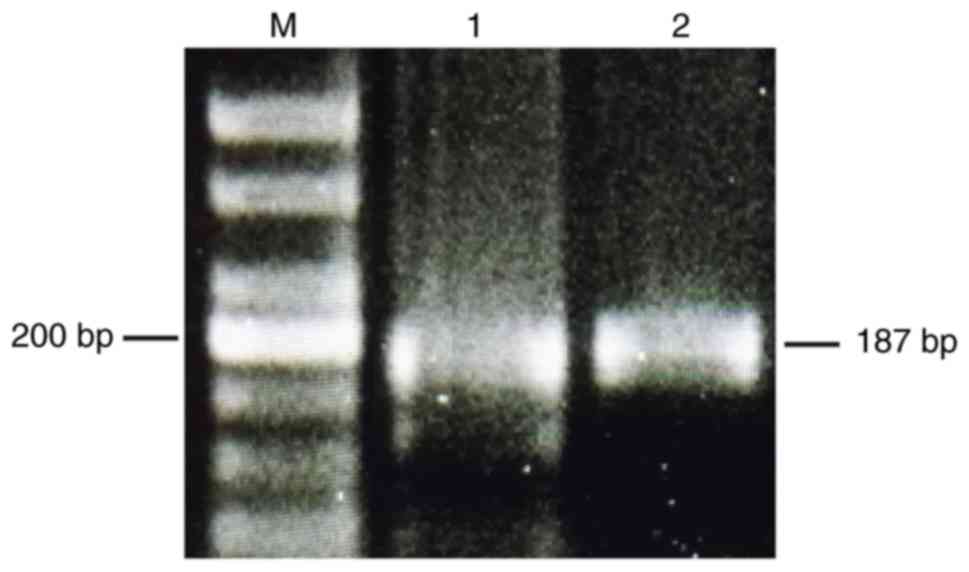
OVCAR3 and OVCAR8 cell lines were revealed to
express AMHRII mRNA. Electrophoretic gel examination of the PCR
products confirmed an appropriate band of the predicted length of
~200 bp for AMHRII in the two cell lines (Fig. 1).
RhAMH inhibits EOC cell
proliferation
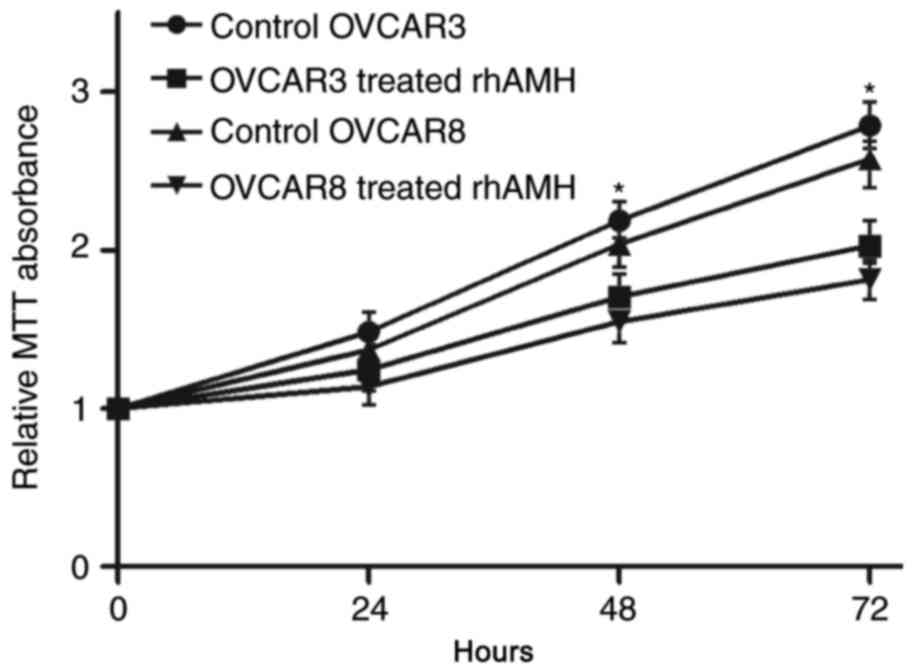
To identify the role of rhAMH in the proliferation
of EOC cells, an MTT assay was conducted using OVCAR3 and OVCAR8
cell lines. Compared with the control group (treated with fetal
bovine serum without rhAMH), the exogenous addition of rhAMH (10
µg/ml) at 37°C significantly reduced the proliferation of the two
cell lines (P<0.01; Fig. 2). At 72
h after exposure, the inhibition was greater compared with at 48
h.
RhAMH induces EOC cell apoptosis
Cell apoptosis was analyzed using FACS. Annexin
V-FITC staining was used to detect early-stage apoptosis in cells
treated with rhAMH. The results revealed that the apoptosis rate in
the rhAMH treated group (48 h) was significantly increased compared
with the control group (OVCAR3, P=0.035; OVCAR8, P=0.020). The
apoptosis rate increased at 72 h, but was not statistically
significantly different when compared with the 48 h group (OVCAR3,
P=0.145; OVCAR8, P=0.296; Fig. 3A and
B).
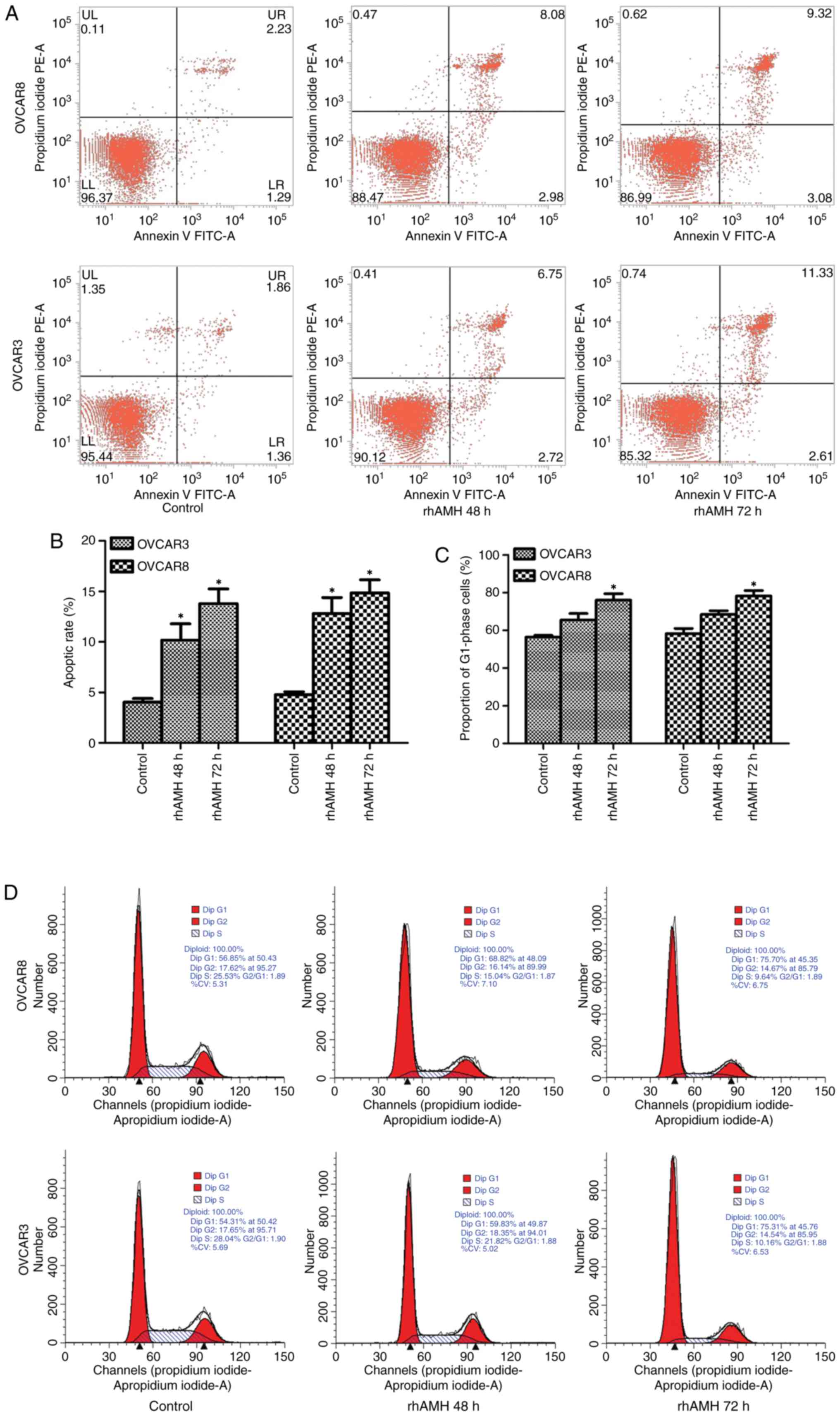 | Figure 3.OVCAR3 and OVCAR8 cell apoptosis
detection by fluorescence-activated cell sorting and cell cycle
analysis by flow cytometry. (A) Cell apoptosis was analyzed using
Annexin V FITC and (B) quantified. The apoptosis rate in the rhAMH
treated group (48 h) was significantly increased compared with the
control group (OVCAR3, P=0.035; OVCAR8, P=0.020). The apoptosis
rate increased at 72 h but was not significantly different compared
with the 48 h group (OVCAR3, P=0.145; OVCAR8, P=0.296). (C)
Quantified flow cytometry revealed the number of cells at the
G1 phase. The percentage of cells at G1 phase
in the rhAMH treated group (48 h) increased, but was not
significantly different when compared with the control group
(OVCAR3, P=0.070; OVCAR8, P=0.051). A significant difference was
identified at 72 h compared with the control group (OVCAR3,
P=0.016; OVCAR8, P=0.019). (D) Flow cytometry results. *P<0.01
vs. the control. RhAMH, recombinant human anti-Müllerian hormone;
FITC, fluorescein isothiocyanate. |
RhAMH induces G1/S cell cycle arrest
in EOC cells
Exogenous addition of rhAMH (10 µg/ml) substantially
increased the percentage of cells at the G1 phase and
decreased the percentage of cells in the S phase. The percentage of
cells in the G1 phase in the rhAMH treated group (48 h)
increased but was not significantly different when compared with
the control group (OVCAR3, P=0.070; OVCAR8, P=0.051). However,
there was a significant difference at 72 h compared with the
control group (OVCAR3, P=0.016; OVCAR8, P=0.019; Fig. 3C). The result of the single test via
flow cytometry is shown in Fig. 3D.
These results suggested that rhAMH may induce G1/S-phase
arrest in EOC cells.
RhAMH enhanced the transcriptional
activity and protein expression of SCF
The interaction between SCF and c-kit enhances cell
proliferation and suppresses apoptosis (11–14). To
further investigate the molecular mechanisms underlying the
rhAMH-mediated effects on proliferation, apoptosis and the cell
cycle, the levels of SCF in culture supernatants and tumor
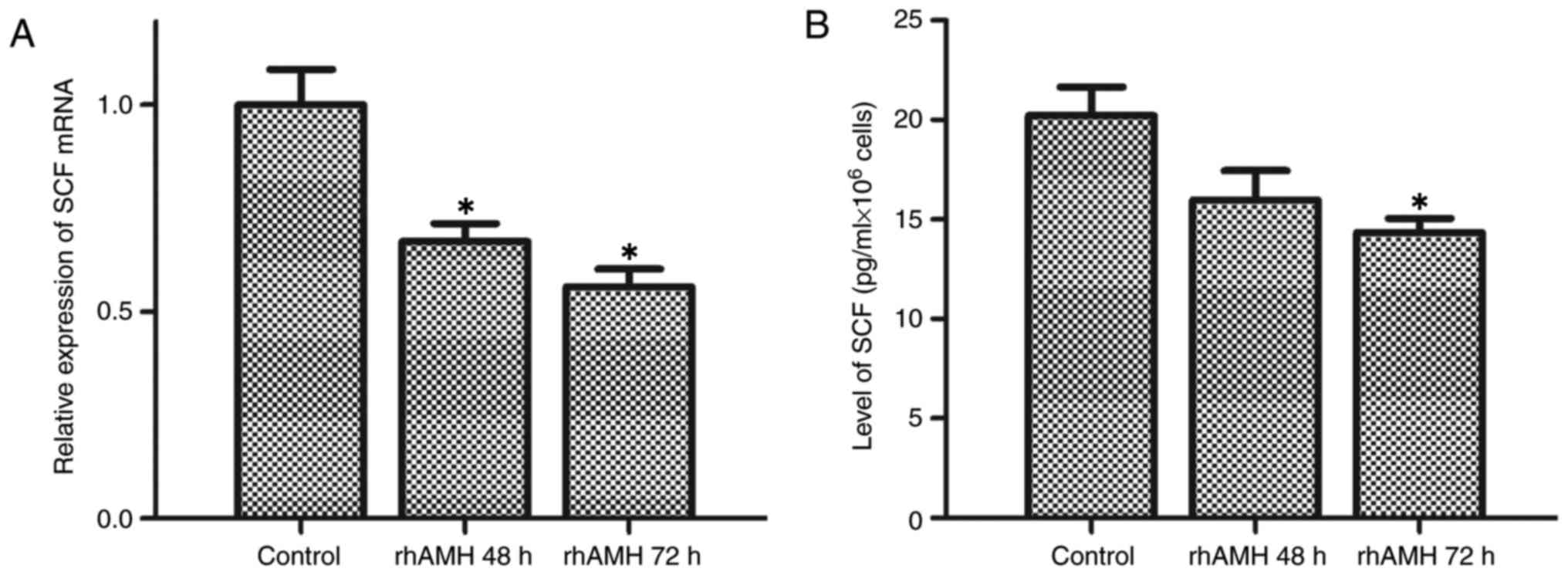
homogenates were detected using RT-qPCR and ELISA. As presented in
Fig. 4A and B, SCF was downregulated
following treatment with rhAMH in OVCAR8 cells. Treatment with
rhAMH for 48 h resulted in the significant inhibition of SCF mRNA
expression levels (P=0.008) but did not significantly affect the
protein expression level (P=0.101) compared with the control, while
the mRNA and protein expression levels were significantly inhibited
at 72 h (mRNA expression levels, P=0.005; protein expression
levels, P=0.036).
Discussion
Although the biological characteristics of ovarian
malignant tumor types have been widely investigated in previous
decades, the prognosis remains poor, with the 5-year survival rate
of advanced ovarian cancer being <30% (2). Therefore, there is an urgent requirement
to develop effective drugs that reduce the risk of recurrence
subsequent to first-line treatment and inhibit drug-resistant
cancer. Previous studies have revealed that the physiological
hormone AMH may not only specifically suppress the proliferation,
invasion and metastasis of AMHRII-positive ovarian cancer, but that
it may additionally reduce drug resistance (17,18),
suggesting that AMH may have potential therapeutic uses against
ovarian cancer. In the present study, a commonly used low dose (10
µg/ml) of rhAMH was administered to OVCAR3 and OVCAR8 cells in
vitro. The results revealed that, compared with the control
group, rhAMH inhibited EOC cell proliferation, induced apoptosis
and resulted in a time-dependent G1/S-phase cell cycle
arrest. These results suggest that the suppression of proliferation
and the induction of apoptosis by AMH may occur via blocking the
transition from the G1 to the S phase.
One previous study has revealed that exogenous AMH
may activate the Smad family member 1/5 signaling pathway in
ovarian granulosa cell tumor types, leading to the activation of
caspase-3 and consequently to apoptosis (19). G1/S-phase cell cycle arrest
may be exerted via upregulating the cyclin-dependent kinase
inhibitor p16 through an AMHRII-mediated mechanism and inhibiting
growth in the absence of detectable or inactive retinoblastoma (Rb)
protein (20). Prolonged treatment
with rhAMH downregulates the Rb-associated protein p130 and
increases the Rb family-regulated transcription factor E2F
transcription factor 1, the overexpression of which inhibits the
growth of EOC (20).
In the present study, the biological agent (rhAMH)
mediating the afore-described functions was investigated. SCF
serves an important function in the recurrence, development,
invasion, metastasis and drug resistance of a variety of malignant
tumor types (12–14). Previous studies have also revealed
that rhAMH may inhibit the expression of SCF in normal ovarian
epithelial cells and granular cells (15,21).
Preliminary experiments in the present study detected the
expression of SCF in the OVCAR8 cell line. Subsequently, the mRNA
and protein expression levels of SCF in the OVCAR8 cell line at 48
and 72 h following rhAMH treatment were examined. The results
demonstrated that rhAMH reduced SCF mRNA transcription and protein
levels in OVCAR8 cells, suggesting that SCF may be involved in the
process of AMH-mediated inhibition of ovarian cancer. The
inhibition was increased further at 72 h, suggesting that rhAMH may
exert a time-dependent suppression of SCF secretion.
One previous study by Zhang et al (22) demonstrated that SCF may increase mast
cell numbers and microvascular density in a tumor. The expression
of von Willebrand factor, an endothelial cell marker, was also
increased, suggesting that SCF modulates tumor growth and
angiogenesis via the involvement of mast cells (22). A study by Huang et al (23) further revealed that SCF may not only
effectively recruit mast cells, which express multiple
proinflammatory factors and increase interleukin-17 expression in a
tumor, but that it may also exacerbate tumor immunosuppression by
releasing adenosine and increasing T-regulatory cells, which
augment the suppression of T cells and natural killer cells in a
tumor. Additionally, bone marrow-derived suppression cells may in
turn stimulate tumor cells to secrete a soluble form of SCF,
further activating mast cells through the secretion of matrix
metalloproteinase 9 (24). A study by
Shaw and Vanderhyden (25) revealed
that exogenous SCF may induce drug-resistance and apoptosis
resistance in ovarian cancer cells, while the c-kit neutralizing
antibody anti-c-kit antibody and c-kit inhibitor imatinib mesylate
may bind precisely to c-kit, enhancing the chemosensitivity and
inducing the apoptosis of ovarian cancer cells. Taken together with
the results of the present study, it may be proposed that the joint
administration of rhAMH and imatinib reduce the side effects and
improve the antitumor effects for patients with ovarian cancer.
Another study also revealed that treatment of EOC cells with rhAMH
combined with calcitriol resulted in the inhibition of cell growth
and survival (26). The combined
treatment significantly suppressed cell growth, downregulated the
expression of B-cell lymphoma 2, and upregulated the expression of
Bcl-2-associated X protein, caspase-3 and caspase-9 through the
extracellular signal-regulated kinase signaling pathway (26). Researchers have further investigated
the underlying pathways involving the function of SCF in the
inhibition of AMHRII-positive ovarian cancer cells. Hu et al
(15) revealed that rhAMH may reduce
the mRNA transcription and protein expression levels of SCF through
the cAMP/PKA pathway in human granulosa cells. A study by Katiyar
et al (27) identified that
the proto-oncogene c-Jun may induce SCF protein, mRNA and promoter
activity. Induction of the SCF promoter required the c-Jun
DNA-binding domain (27).
A number of limitations of the present study should
be addressed. Firstly, EOC cells may have a mutation of AMHRII,
which affect the level and function of AMHRII. The study was
divided into the control group and rhAMH-treated (AMHR+) group.
However, there was no rhAMH-treated (AMHR-) cells designed.
Secondly, OVCAR3 and OVCAR8 were cultured for considerable periods
of time and may be unreliable. However, the present study primarily
focused on the AMHRII expression of EOC cells according to previous
studies (26,28). No inconsistent or variable responses
of these cell lines were observed. In future studies the study
design will be improved by using a greater number of EOC cell
lines. Thirdly, the rhAMH was purchased from R&D Systems, Inc.,
which has been previously demonstrated to be inactive in a
Müllerian duct regression assay (29). The same product, purchased from
alternative companies, will be used in future studies. Finally, the
working concentration of rhAMH was diluted to 10 µg/ml according to
previous experiments (30,31). It would have been ideal to instead
perform a dose-response analysis to identify the optimal working
concentration.
To conclude, the present study revealed that rhAMH
may be able to inhibit the proliferation and induce the apoptosis
of EOC cells via G1/S-phase cell cycle arrest and
decreased SCF secretion. In addition, it was demonstrated that
rhAMH may inhibit the synthesis of SCF in EOC cells at the
transcription and translation levels. Future studies will evaluate
the effects of a higher concentration of rhAMH, applied to a
greater number of ovarian cancer cell lines. Silencing or
upregulating SCF via cell transfection may illuminate the direct
function of SCF in ovarian cancer, and the various pathways
involved. In addition, the number of immunocytes in tumor tissue,
the microvascular density and the levels of associated inflammatory
factors in EOC will be evaluated.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Female Cancer
Drug Program of Guangdong Province Pharmaceutical Society (grant
no. 2014D08) awarded to Dr Jian Gu (Department of Gynecology, The
Third Affiliated Hospital of Sun Yat-Sen University, Guangdong,
China).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TZ and JG conceived and designed the study. QX
performed the flow cytometry assay. TZ and LD performed all other
experiments. SS carried out statistical analysis and interpreted
results. All drafts of the reports were written by TZ and LD. All
authors read and approved the final paper.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AMH
|
anti-Müllerian hormone
|
|
AMHRII
|
anti-Müllerian hormone type II
receptor
|
|
EOC
|
epithelial ovarian cancer
|
|
rhAMH
|
recombinant human anti-Müllerian
hormone
|
|
SCF
|
stem cell factor
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lowe KA, Chia VM, Taylor A, O'Malley C,
Kelsh M, Mohamed M, Mowat FS and Goff B: An international
assessment of ovarian cancer incidence and mortality. Gynecol
Oncol. 130:107–114. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Durlinger AL, Gruijters MJ, Kramer P,
Karels B, Kumar TR, Matzuk MM, Rose UM, de Jong FH, Uilenbroek JT,
Grootegoed JA and Themmen AP: Anti-Müllerian hormone attenuates the
effects of FSH on follicle development in the mouse ovary.
Endocrinology. 142:4891–4899. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Durlinger AL, Kramer P, Karels B, de Jong
FH, Uilenbroek JT, Grootegoed JA and Themmen AP: Control of
primordial follicle recruitment by anti-Müllerian hormone in the
mouse ovary. Endocrinology. 140:5789–5796. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Masiakos PT, MacLaughlin DT, Maheswaran S,
Teixeira J, Fuller AF Jr, Shah PC, Kehas DJ, Kenneally MK,
Dombkowski DM, Ha TU, et al: Human ovarian cancer, cell lines, and
primary ascites cells express the human Müllerian inhibiting
substance (MIS) type II receptor, bind, and are responsive to MIS.
Clin Cancer Res. 5:3488–3499. 1999.PubMed/NCBI
|
|
6
|
Stephen AE, Pearsall LA, Christian BP,
Donahoe PK, Vacanti JP and MacLaughlin DT: Highly purified
Müllerian inhibiting substance inhibits human ovarian cancer in
vivo. Clin Cancer Res. 8:2640–2646. 2002.PubMed/NCBI
|
|
7
|
Meirelles K, Benedict LA, Dombkowski D,
Pepin D, Preffer FI, Teixeira J, Tanwar PS, Young RH, MacLaughlin
DT, Donahoe PK and Wei X: Human ovarian cancer stem/progenitor
cells are stimulated by doxorubicin but inhibited by Müllerian
inhibiting substance. Proc Natl Acad Sci USA. 109:2358–2363. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wei X, Dombkowski D, Meirelles K,
Pieretti-Vanmarcke R, Szotek PP, Chang HL, Preffer FI, Mueller PR,
Teixeira J, MacLaughlin DT and Donahoe PK: Müllerian inhibiting
substance preferentially inhibits stem/progenitors in human ovarian
cancer cell lines compared with chemotherapeutics. Proc Natl Acad
Sci USA. 107:18874–18879. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Szotek PP, Pieretti-Vanmarcke R, Masiakos
PT, Dinulescu DM, Connolly D, Foster R, Dombkowski D, Preffer F,
Maclaughlin DT and Donahoe PK: Ovarian cancer side population
defines cells with stem cell-like characteristics and Müllerian
Inhibiting Substance responsiveness. Proc Natl Acad Sci USA.
103:11154–11159. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pieretti-Vanmarcke R, Donahoe PK, Szotek
P, Manganaro T, Lorenzen MK, Lorenzen J, Connolly DC, Halpern EF
and MacLaughlin DT: Recombinant human Müllerian inhibiting
substance inhibits long-term growth of MIS type II
receptor-directed transgenic mouse ovarian cancers in vivo. Clin
Cancer Res. 12:1593–1598. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liang J, Wu YL, Chen BJ, Zhang W, Tanaka Y
and Sugiyama H: The C-kit receptor-mediated signal transduction and
tumor-related diseases. Int J Biol Sci. 9:435–443. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tonary AM, Macdonald EA, Faught W,
Senterman MK and Vanderhyden BC: Lack of expression of c-KIT in
ovarian cancers is associated with poor prognosis. Int J Cancer.
89:242–250. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wilczynski SP, Chen YY, Chen W, Howell SB,
Shively JE and Alberts DS: Expression and mutational analysis of
tyrosine kinase receptors c-kit, PDGFRalpha, and PDGFRbeta in
ovarian cancers. Hum Pathol. 36:242–249. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Arber DA, Tamayo R and Weiss LM: Paraffin
section detection of the c-kit gene product (CD117) in human
tissues: Value in the diagnosis of mast cell disorders. Hum Pathol.
29:498–504. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu R, Wang FM, Yu L, Luo Y, Wu X, Li J,
Zhang XM, Oehninger S and Bocca S: Antimüllerian hormone regulates
stem cell factor expression in human granulosa cells. Fertil
Steril. 102:1742–1750.e1. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Magee JA, Piskounova E and Morrison SJ:
Cancer stem cells: Impact, heterogeneity, and uncertainty. Cancer
Cell. 21:283–296. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Foster R, Buckanovich RJ and Rueda BR:
Ovarian cancer stem cells: Working towards the root of stemness.
Cancer Lett. 338:147–157. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Anttonen M, Färkkilä A, Tauriala H,
Kauppinen M, Maclaughlin DT, Unkila-Kallio L, Bützow R and
Heikinheimo M: Anti-Müllerian hormone inhibits growth of AMH type
II receptor-positive human ovarian granulosa cell tumor cells by
activating apoptosis. Lab Invest. 91:1605–1614. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ha TU, Segev DL, Barbie D, Masiakos PT,
Tran TT, Dombkowski D, Glander M, Clarke TR, Lorenzo HK, Donahoe PK
and Maheswaran S: Müllerian inhibiting substance inhibits ovarian
cell growth through an Rb-independent mechanism. J Biol Chem.
275:37101–37109. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nilsson E, Rogers N and Skinner MK:
Actions of anti-Müllerian hormone on the ovarian transcriptome to
inhibit primordial to primary follicle transition. Reproduction.
134:209–221. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang W, Stoica G, Tasca SI, Kelly KA and
Meininger CJ: Modulation of tumor angiogenesis by stem cell factor.
Cancer Res. 60:6757–6762. 2000.PubMed/NCBI
|
|
23
|
Huang B, Lei Z, Zhang GM, Li D, Song C, Li
B, Liu Y, Yuan Y, Unkeless J, Xiong H and Feng ZH: SCF-mediated
mast cell infiltration and activation exacerbate the inflammation
and immunosuppression in tumor microenvironment. Blood.
112:1269–1279. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu J, Zhang Y, Zhao J, Yang Z, Li D,
Katirai F and Huang B: Mast cell: Insight into remodeling a tumor
microenvironment. Cancer Metastasis Rev. 30:177–184. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shaw TJ and Vanderhyden BC: AKT mediates
the pro-survival effects of KIT in ovarian cancer cells and is a
determinant of sensitivity to imatinib mesylate. Gynecol Oncol.
105:122–131. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jung YS, Kim HJ, Seo SK, Choi YS, Nam EJ,
Kim S, Kim SW, Han HD, Kim JW and Kim YT: Anti-proliferative and
apoptotic activities of Müllerian inhibiting substance combined
with calcitriol in ovarian cancer cell lines. Yonsei Med J.
57:33–40. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Katiyar S, Jiao X, Wagner E, Lisanti MP
and Pestell RG: Somatic excision demonstrates that c-Jun induces
cellular migration and invasion through induction of stem cell
factor. Mol Cell Biol. 27:1356–1369. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Barbie TU, Barbie DA, MacLaughlin DT,
Maheswaran S and Donahoe PK: Mullerian Inhibiting Substance
inhibits cervical cancer cell growth via a pathway involving p130
and p107. Proc Natl Acad Sci USA. 100:15601–15606. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kano M, Sosulski AE, Zhang L, Saatcioglu
HD, Wang D, Nagykery N, Sabatini ME, Gao G, Donahoe PK and Pépin D:
AMH/MIS as a contraceptive that protects the ovarian reserve during
chemotherapy. Proc Natl Acad Sci USA. 114:E1688–E1697. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Park SH, Chung YJ, Song JY, Kim SI, Pépin
D, MacLaughlin DT, Donahoe PK and Kim JH: Müllerian inhibiting
substance inhibits an ovarian cancer cell line via β-catenin
interacting protein deregulation of the Wnt signal pathway. Int J
Oncol. 50:1022–1028. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chang HL, Pieretti-Vanmarcke R, Nicolaou
F, Li X, Wei X, MacLaughlin DT and Donahoe PK: Mullerian inhibiting
substance inhibits invasion and migration of epithelial cancer cell
lines. Gynecol Oncol. 120:128–134. 2011. View Article : Google Scholar : PubMed/NCBI
|