Introduction
As a type of malignant tumor that develops in the
esophagus, esophageal cancer affects ~0.3% of females and 0.8% of
males (1). Currently, esophageal
cancer is one of the least studied but one of the deadliest types
of malignant tumor due to a low survival rate and the extremely
aggressive nature of esophageal cancer cells (2,3). The
progression of esophageal cancer is a complex process involving
various internal and environment factors (1,3). Previous
studies have demonstrated that risk factors for esophageal cancer
include drinking hot water, poor oral health, smoking, consumption
of red meat, alcohol, low socioeconomic status and numerous other
factors (3,4). With the development of treatments for
malignant tumors, the overall survival rates of patients with a
number of different of cancer types have been significantly
improved over the past decades (5).
However, the treatment of esophageal cancer is unreliable for
providing satisfactory outcomes due to an unclear understanding of
its pathogenesis (1,6). Therefore, understanding the mechanisms
underlying esophageal cancer to improve the treatment of this
disease is of great clinical significance.
Mitotic spindle checkpoint signaling, which is
essential for normal cell division, has been indicated to be
involved in the development of a variety of cancer types (7). A previous study indicated that the
alternative reading frame tumor suppressor may maintain mitotic
checkpoint fidelity to prevent chromosomal instability (8), indicating the crosstalk between mitotic
checkpoint fidelity and tumor development. As a mitotic spindle
checkpoint gene, baculoviral IAP repeat containing 5 (BIRC5) was
determined to have the ability to regulate cell apoptosis (9). BIRC5 was also indicated to be involved
in the onset and development of various types of malignant tumors,
including breast (10), colorectal
(11) and gastric cancer (12). Therefore, BIRC5 may also serve
important roles in the development of esophageal cancer. The level
of BIRC5 expression was reported to be increased in esophageal
squamous cell carcinoma tissues compared with normal human tissues
(13). However, the specific roles of
BIRC5 in the pathogenesis of esophageal cancer have not been well
studied.
In the present study, expression levels of BIRC5 in
patients with different stages of esophageal cancer and in
different esophageal cancer cell lines were detected. The effect of
BIRC5 on tumor cell migration and invasion was explored. The
association between BIRC5 and angiogenesis-associated factor
vascular endothelial growth factor (VEGF), brain-specific
angiogenesis inhibitor 1 (BAI1) and methyl-CpG binding domain
protein 2 (MBD2), and the phosphatidylinositol 3-kinase (PI3K)/Akt
pathway was also investigated.
Materials and methods
Patients
A total of 178 patients with esophageal cancer
diagnosed via pathological examination and imaging methods,
including mini-probe ultrasonography, spiral computed tomography
and magnetic resonance imaging were selected at The Fourth
Affiliated Hospital of Hebei Medical University (Shijiazhuang,
China) from May 2015 to September 2016. The patients included 81
males and 97 females, with an age range of 35–89 years old and a
mean age of 54±7.8 years. The inclusion criteria are as follows: i)
Diagnosis of malignant esophageal tumor and ii) signing of informed
consent. The exclusion criteria are as follows: i) Patients with
other types of malignant tumors; ii) patients with severe
coagulation dysfunction and iii) patients who refused to sign the
informed consent. The present study was approved by the Ethics
Committee of The Fourth Affiliated Hospital of Hebei Medical
University. According to the pathological examination and imaging
results, the patients were divided into different stages of
esophageal cancer according to the following criteria, as
previously described (14): T0, no
detection of primary tumor (n=30); Tis, high-grade dysplasia
(n=25); T1, muscularis mucosae, lamina propria or submucosa invaded
by the tumor (n=29); T2, muscularis propria invaded by the tumor
(n=29); T3, adventitia invaded by the tumor (n=32); T4, adjacent
structures invaded by the tumor (n=33). No significant difference
in age, sex and other basic information was determined among
patients in different stages of esophageal cancer.
Cell lines and cell culture
Homo sapiens esophageal normal cell line
Het-1A, esophageal cancer cell line KYSE510 and EC9706 were
purchased from American Type Culture Collection (Manassas, VA,
USA). All cells were cultured in RPMI-1640 medium (HyClone; GE
Healthcare Life Sciences, Logan, UT, USA) containing 10% fetal
bovine serum (FBS; Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) in an incubator at 37°C with an atmosphere
containing 5% CO2. FBS was not added for cells used for
treatment with the PI3K/Akt pathway activator SC79 (10 µM).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from tumor tissue and in
vitro cultured cells using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), and cDNA was
synthesized using SuperScript III reverse transcriptase kit (Thermo
Fisher Scientific, Inc.). Reverse transcription conditions were: 50
min at 50°C and 5 min at 85°C. SYBR®-Green Real-Time PCR
Master Mix (Thermo Fisher Scientific, Inc.) was used to prepare PCR
reaction system, and PCR reactions were performed on a CFX96
Touch™ Real-Time PCR Detection system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The following primers were
used in PCR: BIRC5, forward, 5′-TGCCTGGCAGCCCTTTC-3′ and reverse,
5′-CCTCCAAGAAGGGCCAGTTC-3′; VEGF, forward,
5′-ATCTTCAAGCCATCCTGTGTGC-3′ and reverse,
5′-CAAGGCCCACAGGGATTTTC-3′; BAI1, forward,
5′-GCAAACCAAGTTCTGCAACAT-3′ and reverse,
5′-GCAAACCAAGTTCTGCAACAT-3′; MBD2, forward,
5′-CCCTGCTGTTTGGCTTAACACATCTC-3′ and reverse,
5′-TGCGTACTTGCTGTACTCGCTCTTC-3′; GAPDH, forward,
5′-GAGTCAACGGATTTGGTCGT-3′ and reverse, 5′-TTGATTTTGGAGGGATCTCG-3′
for GAPDH. The PCR reaction conditions were: 95°C for 30 sec,
followed by 40 cycles of 95°C for 10 sec and 50°C for 45 sec. The
data were analyzed using the 2−∆∆Cq method (15), and the relative expression level of
each gene was normalized to endogenous control GAPDH.
Small-interfering (si)RNA
transfection
BIRC5 siRNA (catalog no. 121294) was purchased from
Thermo Fisher Scientific, Inc. Silencer® negative
control #1 siRNA (Thermo Fisher Scientific, Inc.) was used as a
negative control. Oligofectamine™ reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) was used to transfect 40 nM siRNA
into 5×105 cells by incubation with the cells at 37°C in
a CO2 incubator for 4 h, followed by incubation in
RPMI-1640 medium (Hyclone; GE Healthcare Life Sciences, Logan, UT,
USA) at 37°C for 48 h prior to harvest.
Transwell migration and invasion
assays
Cell migratory ability of each cell line was
measured using Transwell cell migration assay (polyester inserts,
BD Biosciences, Franklin Lakes, NJ, USA). In brief,
~5×104 cells were transferred to the upper chamber
containing RPMI-1640 medium without FCS, while RPMI-1640 medium
(Thermo Fisher Scientific, Inc.) containing 20% FCS (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) was used to fill the lower chamber.
The membrane was collected 24 h later, followed by staining for 20
min with 0.5% crystal violet at room temperature. Following
staining, the cells on the membrane were counted under a light
microscope (magnification, ×20; Olympus Corporation, Tokyo, Japan).
Cell invasion assay was performed using the same method except that
Matrigel (cat no. 356234; EMD Millipore, Billerica, MA, USA) was
used to coat the upper chamber prior to experiment. Each experiment
was repeated 3 times to calculate the mean value.
Western blot analysis
Total protein was extracted from cells using Cell
Lysis Solutions (Thermo Fisher Scientific, Inc.), and BCA method
was used to measure protein concentration. Following denaturation,
30 µg protein (mass per lane) from each sample was subjected to
electrophoresis using 10% SDS-PAGE gel, followed by transfer to
polyvinylidene fluoride membranes. Following washing with TBS with
Tween 20 (TBST) buffer three times for 10 min, the membrane was
blocked with 5% skimmed milk for 2 h at room temperature. The
membranes were then incubated with corresponding primary antibodies
including rabbit anti-BIRC5 polyclonal antibody (dilution, 1:1,000;
catalog no. MBS151070; MyBioSource, Inc., San Diego, CA, USA),
rabbit anti-VEGF polyclonal antibody (dilution, 1:1,000; catalog
no. bs-0279R; Biomathematics and Statistics Scotland, Edinburgh,
Scotland), rabbit anti-BAI1 polyclonal antibody (dilution, 1:1,000;
catalog no. MBS8242484; MyBioSource, Inc.), rabbit anti-MBD2
polyclonal antibody (dilution, 1:1,000; catalog no. MBS126361;
MyBioSource, Inc.), rabbit anti-phospho (p)-Akt polyclonal antibody
(dilution, 1:1,000; catalog no. MBS330024; MyBioSource, Inc.) and
rabbit anti-GAPDH polyclonal antibody (dilution, 1:1,000; catalog
no. MBS9216842; MyBioSource, Inc.) overnight at 4°C. The membrane
was then washed with TBST three times (10 min each time), followed
by incubation with goat anti-rabbit IgG-horseradish peroxidase
secondary antibody (dilution, 1:1,000; catalog no. MBS435036;
MyBioSource, Inc.) at room temperature for 2 h. Following washing,
chemiluminescence method was used to detect the signal, and the
relative expression level of each protein was calculated by
normalizing to the endogenous control GAPDH using the ImageJ 1.48
software (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
SPSS statistical software (version 19.0; IBM Corp.,
Armonk, NY, USA) was used for statistical analysis. All data are
expressed as the mean ± standard deviation. Comparisons between two
groups of data were performed using an unpaired Student's t-test.
Comparisons among multiple groups were performed using one-way
analysis of variance, followed by the least significant difference
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Expression level of BIRC5 mRNA in
patients with different stages of esophageal cancer
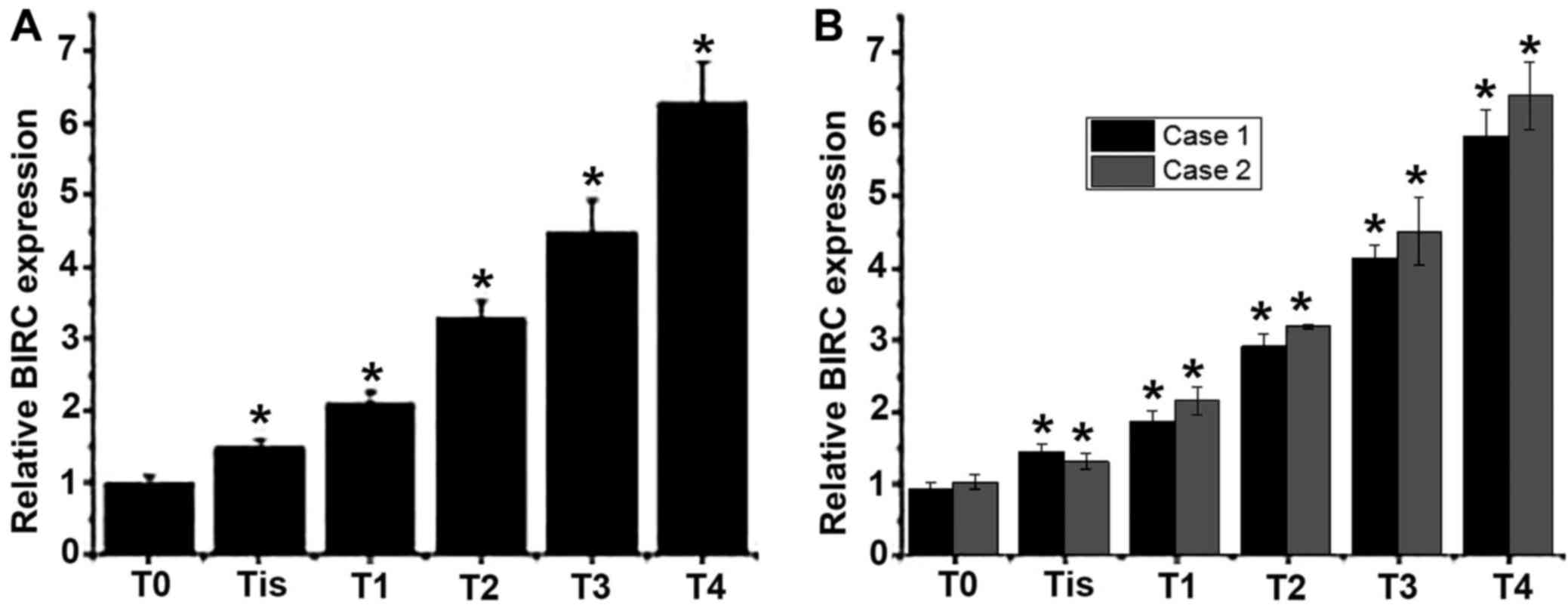
Total RNA was extracted from tumor tissues collected
from patients with different stages of esophageal cancer to detect
the mRNA expression of BIRC5. As presented in Fig. 1A and B, an increased level of BIRC5
mRNA expression was observed with the progression of esophageal
cancer. Those results indicated that BIRC5 may be involved in the
development of esophageal cancer.
Expression of BIRC5 gene in different
cell lines
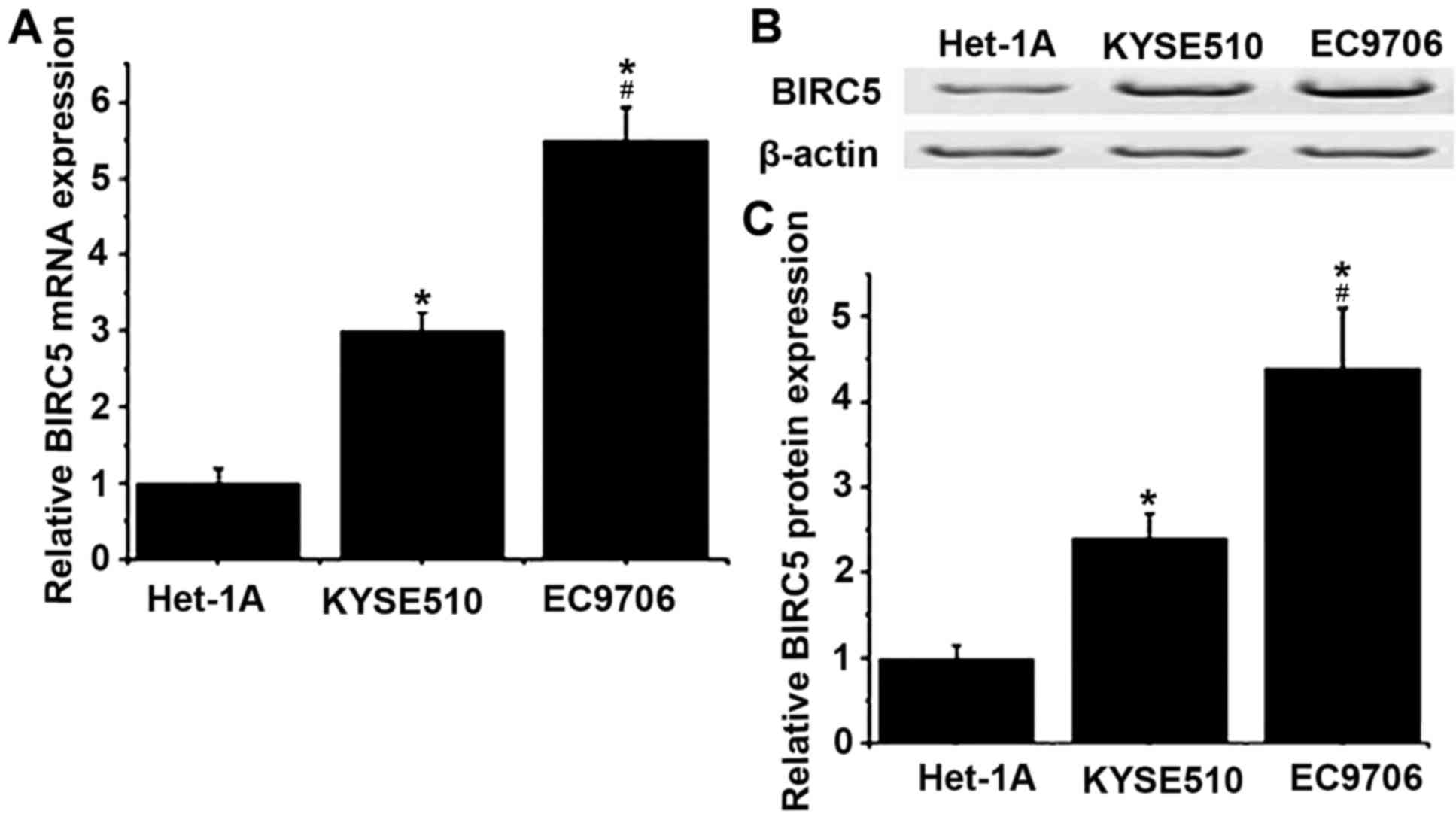
The expression levels of BIRC5 in Homo
sapiens normal esophageal cell line Het-1A, and esophageal
cancer cell lines KYSE510 and EC9706 were detected by RT-qPCR and
western blotting at the level of mRNA and protein, respectively. As
presented in Fig. 2, the levels of
mRNA and protein of BIRC5 were determined to be significantly
increased in KYSE510 and EC9706 cells compared with Het-1A cells
(P<0.05). Compared with KYSE510, mRNA and protein expression
levels of BIRC5 were significantly increased in EC9706 cells
(P<0.05).
Effects of BIRC5 siRNA silencing on
the expression levels of angiogenesis-associated factor VEGF, BAI1
and MBD2 proteins in different cell lines
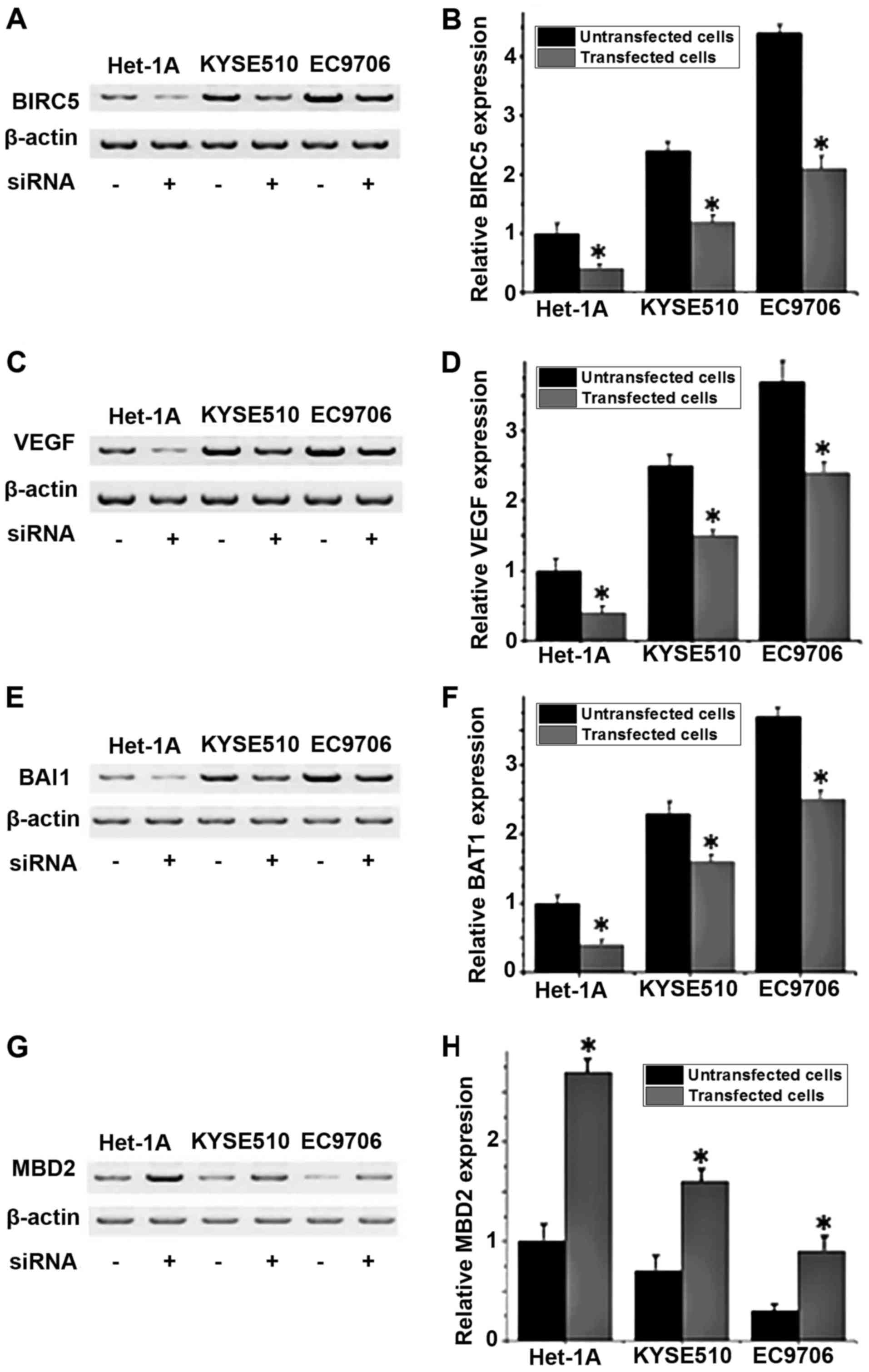
The protein expression levels of VEGF, BAI1 and MBD2
in different cell lines with or without BIRC5 silencing were
detected by western blot analysis. No significant differences in
expression of those proteins were detected between the control
group (untransfected cells) and negative control group
(Silencer® Negative Control #1 siRNA; data not shown).
As depicted in Fig. 3A and B, the
expression level of BIRC5 was significantly decreased in all three
cell lines following siRNA transfection compared with untransfected
cells (P<0.05), indicating that BIRC5 expression was
successfully silenced. Compared with untransfected cells, the
levels of VEGF and BAI1 protein expression significantly increased
(P<0.05; Fig. 3C-F), and the level
of MBD2 protein expression significantly decreased in
BIRC5-silenced cells (P<0.05; Fig. 3G
and H). These results indicated that the expression level of
BIRC5 is positively associated with the expression levels of VEGF
and BAI1 and negatively associated with the expression level of
MBD2.
Effects of BIRC5 silencing on
migration and invasion in different cell lines
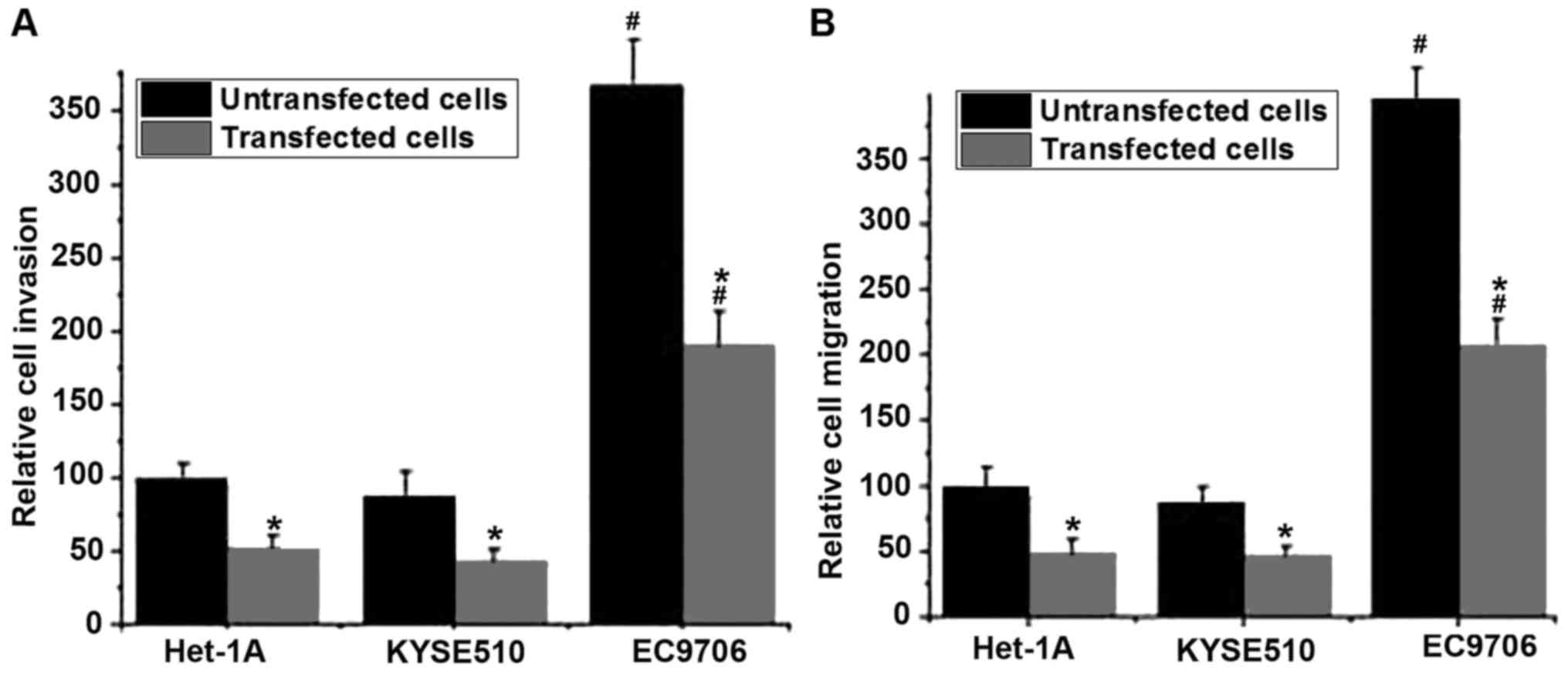
The migratory and invasive abilities of cells in
different cell lines were detected using Transwell migration and
invasion assays. No significant differences in migratory and
invasive abilities of those cell lines were detected between the
control group (untransfected cells) and the negative control group
(Silencer® negative control #1 siRNA; data not shown).
As presented in Fig. 4A, the invasive
abilities of the cells were significantly decreased following BIRC5
silencing compared with untransfected cells (P<0.05). Similar
results were observed in the cell migration assay. The silencing of
BIRC5 was able to significantly reduce the migratory ability of all
three cell lines compared with untransfected cells (P<0.05).
Those results indicated that the silencing of BIRC5 was able to
inhibit the migration and invasion of normal and esophageal cancer
cells. In addition, the migratory and invasive abilities of the
esophageal cancer cell line EC9706 with or without BIRC5 silencing
were significantly higher compared with the esophageal cancer cell
line KYSE510 with the same treatment (P<0.05; Fig. 4A and B).
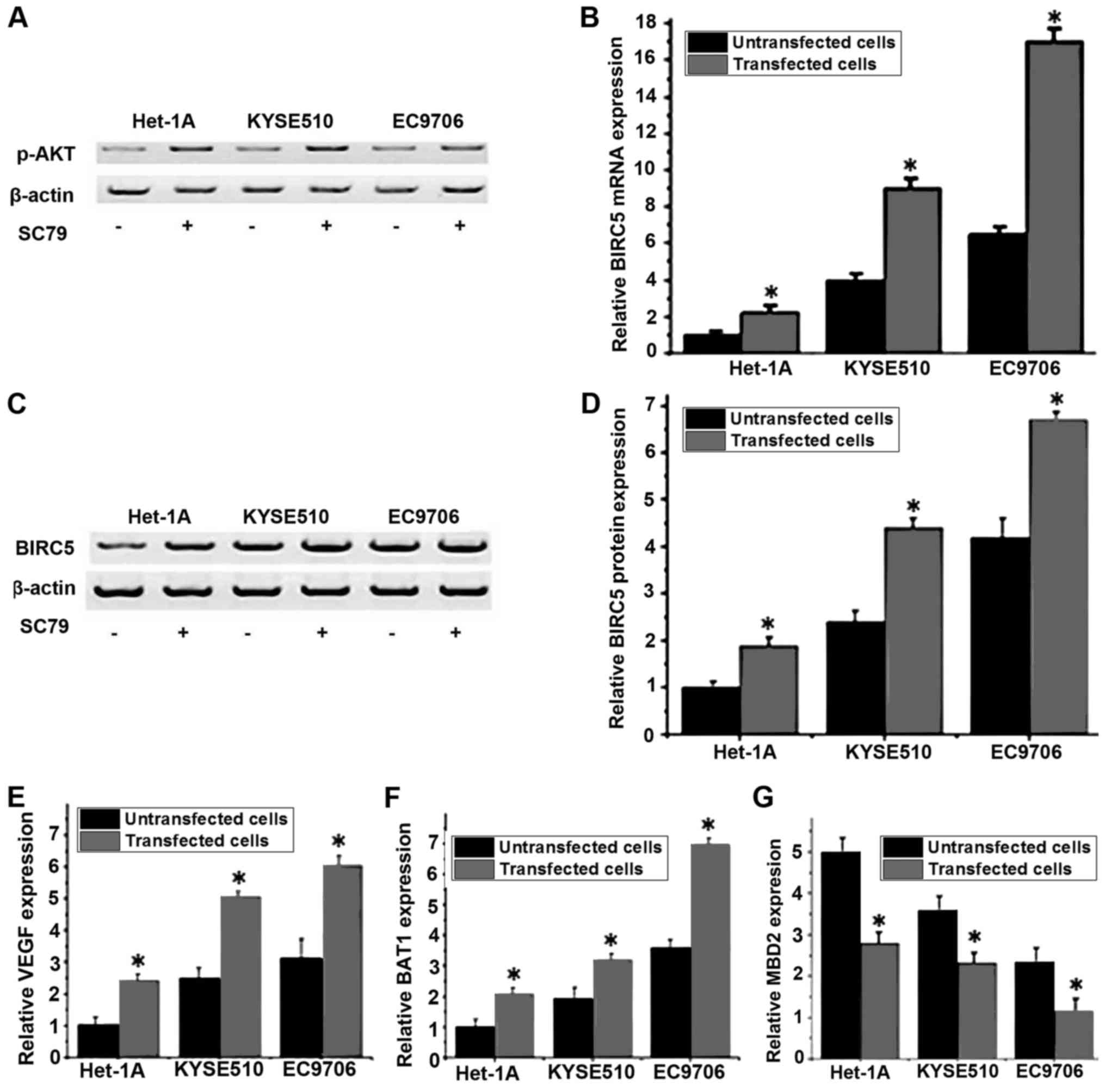
Effect of the PI3K/Akt signaling
pathway activator SC79 on the expression level of BIRC5
A previous study has demonstrated that the
expression and accumulation of BIRC5 was regulated via the PI3K/Akt
pathway (16). The PI3K/Akt pathway
activator SC79 was used to treat different cell lines, and the mRNA
and protein expression levels of BIRC5 in cells that were treated
with or without SC79 were detected by RT-qPCR and western blot
analysis, respectively. As presented in Fig. 5A, the expression level of p-Akt was
increased in all three cell lines following SC79 treatment,
indicating the activation of the PI3K/Akt pathway. As presented in
Fig. 5B, compared with untreated
cells, the mRNA expression level of BIRC5 was significantly
increased in cells that were treated with SC79 in the same cell
line (P<0.05). Similar results were detected for the levels of
BIRC5 protein expression, where the treatment with SC79 was able to
significantly increase the protein expression level of BIRC5
(Fig. 5C and D). Furthermore, RT-qPCR
was performed to detect the mRNA expression of VEGF, BAI1 and MBD2
following SC79 treatment. The results indicated that the expression
levels of VEGF (Fig. 5E) and BAI1
(Fig. 5F) were significantly
increased and the expression level of MBD2 (Fig. 5G) was significantly decreased
following treatment with SC79. These results indicated that the
activation of the PI3K/Akt signaling pathway may upregulate the
mRNA and protein expression level of BIRC5, which in turn promote
the expression of VEGF and BAI1 and inhibit the expression of
MBD2.
Discussion
BIRC5, also termed survivin, is an anti-apoptosis
gene that has been demonstrated to be involved in the progression
of a variety of malignant tumor types (17). The expression of BIRC5 was indicated
to increase in tumor tissues, compared with normal healthy tissues
(12). In a study on breast cancer,
Zu et al (18) reported that
the expression of BIRC5 was significantly upregulated by the
pro-oncogene zinc finger and BTB-domain containing 7A, which in
turn promoted the progression of breast cancer. The expression
level of BIRC5 was also determined to be significantly increased in
colon cancer, and the increased expression of BIRC5 was maintained
during the development of tumor (19). A previous study reported that BIRC5
expression level was continuously increased with the development of
cervical cancer (20). In a study on
esophageal cancer, Kato et al (21) determined that the expression level of
BIRC5 was increased to different extents in patients with
esophageal cancer, and patients with higher expression levels of
BIRC5 were determined to have a reduced survival time compared with
those with lower expression levels, indicating that the expression
level of BIRC5 may be used as a predictor for the survival of
patients.
In the present study, the expression level of BIRC5
was indicated to increase with the progression of esophageal
cancer. In addition, the expression level of BIRC5 was also
determined to be significantly increased in esophageal cancer cell
lines KYSE510 and EC9706, compared with the Homo sapiens
normal esophageal cell line Het-1A, which further confirms that
BIRC5 may be involved in the development of esophageal cancer.
Previous studies have demonstrated that BIRC5 is able to promote
the migration and invasion of a number of different types of tumor
cells (22). In the present study,
the silencing of BIRC5 by siRNA was determined to be able to
significantly reduce the migratory and invasive abilities of the
normal esophageal and two esophageal cancer cell lines. In addition
compared with KYSE510, a significantly higher expression level of
BIRC5 mRNA, and increased migratory and invasive abilities were
detected in the EC9706 cell line, indicating that the expression
level of BIRC5 may be positively associated with the migration and
invasion abilities of esophageal cancer cells.
Angiogenesis serves pivotal roles in the development
of cancer (23), and anti-angiogenic
therapy has been indicated to be a promising strategy in the
treatment of various cancer types (24). VEGF and BAI1 are two factors that can
promote angiogenesis, while MBD2 can interact with the tumor
suppressor gene BAI1 to inhibit its anti-angiogenic function
(25). It is understood that BIRC5
serves pivotal roles in angiogenesis during tumor development
(26). In a study on glioma, BIRC5
was determined to promote angiogenesis through the interactions
with VEGF in vivo and in vitro (26). In the present study, the silencing of
BIRC5 by siRNA in three different cell lines were determined to
reduce the expression levels of VEGF and BAI1 protein and increase
the expression level of MBD2 protein, indicating that the
downregulation of BIRC5 may be able to inhibit angiogenesis in
esophageal cancer.
The PI3K/AKT signaling pathway, which serves key
roles in a variety of physiological and biochemical processes,
including cell proliferation, migration and invasion, is usually
activated in different types of human cancer (27). Treatment strategies that aimed to
inhibit the PI3K/AKT signaling pathway has been frequently used in
the treatment of various types of human cancer, including
endometrial (28) and colorectal
cancer (29). A previous study has
demonstrated that PI3K is able to regulate the expression of BIRC5
in the human ovarian cancer cell line OVCAR-3 via the activation of
Akt. In the present study, the expression level of BIRC5 was
significantly increased following the activation of PI3K/AKT
signaling pathway. Furthermore, the expression levels of VEGF and
BAI1 were significantly increased and the expression level of MBD2
was significantly decreased following treatment with SC79. Those
results indicated that the activation of the PI3K/Akt signaling
pathway may upregulate the levels of BIRC5 mRNA and protein
expression, which in turn promote the expression of VEGF and BAI1,
and inhibit the expression of MBD2.
In conclusion, BIRC5 may be able to promote the
development of esophageal cancer via regulating the expression of
angiogenesis-associated factor and increasing the migration and
invasion of tumor cells. The activation of the PI3K/Akt signaling
pathway is able to upregulate the expression level of BIRC5 to
participate in development of esophageal cancer.
References
|
1
|
Enzinger PC and Mayer RJ: Esophageal
cancer. N Engl J Med. 349:2241–2252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lin Y, Totsuka Y, He Y, Kikuchi S, Qiao Y,
Ueda J, Wei W, Inoue M and Tanaka H: Epidemiology of esophageal
cancer in Japan and China. J Epidemiol. 23:233–242. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang Y: Epidemiology of esophageal
cancer. World J Gastroenterol. 19:5598–5606. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu M, Zhao JK, Zhang ZF, Han RQ, Yang J,
Zhou JY, Wang XS, Zhang XF, Liu AM, van't Veer P, et al: Smoking
and alcohol drinking increased the risk of esophageal cancer among
Chinese men but not women in a high-risk population. Cancer Causes
Control. 22:649–657. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
LeVea C: Pathogenesis of Esophageal Cancer
[M]/Minimally Invasive Foregut Surgery for Malignancy. Springer
International Publishing; pp. 1–9. 2015
|
|
7
|
Weaver BA and Cleveland DW: Decoding the
links between mitosis, cancer, and chemotherapy: The mitotic
checkpoint, adaptation, and cell death. Cancer Cell. 8:7–12. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Britigan EM, Wan J, Zasadil LM, Ryan SD
and Weaver BA: The ARF tumor suppressor prevents chromosomal
instability and ensures mitotic checkpoint fidelity through
regulation of Aurora B. Mol Biol Cell. 25:2761–2773. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li F, Ambrosini G, Chu EY, Plescia J,
Tognin S, Marchisio PC and Altieri DC: Control of apoptosis and
mitotic spindle checkpoint by survivin. Nature. 396:580–584. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gu Y, Li P, Peng F, Zhang M, Zhang Y,
Liang H, Zhao W, Qi L, Wang H, Wang C and Guo Z: Autophagy-related
prognostic signature for breast cancer. Mol Carcinog. 55:292–299.
2016. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang R, Kang Y, Löhr CV, Fischer KA,
Bradford CS, Johnson G, Dashwood WM, Williams DE, Ho E and Dashwood
RH: Reciprocal regulation of BMF and BIRC5 (Survivin) linked to
Eomes overexpression in colorectal cancer. Cancer Lett.
381:341–348. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lim T, Lee I, Kim J and Kang WK:
Synergistic effect of simvastatin plus radiation in gastric cancer
and colorectal cancer: Implications of BIRC5 and connective tissue
growth factor. Int J Radiat Oncol Biol Phys. 93:316–325. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vaĭshlia NA, Zinov'eva MV, Sass AV,
Kopantsev EP, Vinogradova TV and Sverdlov ED: Increase of BIRC5
gene expression in non-small cell lung cancer and esophageal
squamous cell carcinoma does not correlate with expression of genes
SMAC/DIABLO and PML encoding its inhibitors. Mol Biol (Mosk).
42:652–661. 2007.(In Russian).
|
|
14
|
Rice TW and Blackstone EH: Esophageal
cancer staging: Past, present, and future. Thorac Surg Clin.
23:461–469. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao P, Meng Q, Liu LZ, You YP, Liu N and
Jiang BH: Regulation of survivin by PI3K/Akt/p70S6K1 pathway.
Biochem Biophys Res Commun. 395:219–224. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ambrosini G, Adida C and Altieri DC: A
novel anti-apoptosis gene, survivin, expressed in cancer and
lymphoma. Nat Med. 3:917–921. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zu X, Ma J, Liu H, Liu F, Tan C, Yu L,
Wang J, Xie Z, Cao D and Jiang Y: Pro-oncogene Pokemon promotes
breast cancer progression by upregulating survivin expression.
Breast Cancer Res. 13:R262011. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hernandez JM, Farma JM, Coppola D, Hakam
A, Fulp WJ, Chen DT, Siegel EM, Yeatman TJ and Shibata D:
Expression of the antiapoptotic protein survivin in colon cancer.
Clin Colorectal Cancer. 10:188–193. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu D, Qian J, Yin X, Xiao Q, Wang C and
Zeng Y: Expression of PTEN and survivin in cervical cancer:
Promising biological markers for early diagnosis and prognostic
evaluation. Br J Biomed Sci. 69:143–146. 2012.PubMed/NCBI
|
|
21
|
Kato J, Kuwabara Y, Mitani M, Shinoda N,
Sato A, Toyama T, Mitsui A, Nishiwaki T, Moriyama S, Kudo J and
Fujii Y: Expression of survivin in esophageal cancer: Correlation
with the prognosis and response to chemotherapy. Int J Cancer.
95:92–95. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kogo R, How C, Chaudary N, Bruce J, Shi W,
Hill RP, Zahedi P, Yip KW and Liu FF: The microRNA-218~Survivin
axis regulates migration, invasion, and lymph node metastasis in
cervical cancer. Oncotarget. 6:1090–1100. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Carmeliet P and Jain RK: Angiogenesis in
cancer and other diseases. Nature. 407:249–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shojaei F: Anti-angiogenesis therapy in
cancer: Current challenges and future perspectives. Cancer Lett.
320:130–137. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu D, Hunter SB, Vertino PM and Van Meir
EG: Overexpression of MBD2 in glioblastoma maintains epigenetic
silencing and inhibits the antiangiogenic function of the tumor
suppressor gene BAI1. Cancer Res. 71:5859–5870. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang P, Zhen H, Zhang J, Zhang W, Zhang R,
Cheng X, Guo G, Mao X, Wang J and Zhang X: Survivin promotes glioma
angiogenesis through vascular endothelial growth factor and basic
fibroblast growth factor in vitro and in vivo. Mol Carcinog.
51:586–595. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Karar J and Maity A: PI3K/AKT/mTOR pathway
in angiogenesis. Front Mol Neurosci. 4:512011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Slomovitz BM and Coleman RL: The
PI3K/AKT/mTOR pathway as a therapeutic target in endometrial
cancer. Clin Cancer Res. 18:5856–5864. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pandurangan AK: Potential targets for
prevention of colorectal cancer: A focus on PI3K/Akt/mTOR and Wnt
pathways. Asian Pac J Cancer Prev. 14:2201–2205. 2013. View Article : Google Scholar : PubMed/NCBI
|