Introduction
Gastric cancer is one of the most common malignant
tumor types globally. The morbidity of gastric carcinoma in China,
particularly in rural areas, is one of the top leading causes of
cancer mortality, which poses a huge threat to human health
(1). The majority of patients with
early gastric carcinoma are able to survive >5 years or even
achieve remission following surgical treatments. However, the onset
of gastric carcinoma is latent, symptoms and physical signs at the
early stages of the disease are not significant, and the malignancy
develops rapidly (2). For these
reasons, >50% of patients present with advanced stages of the
disease at the point of diagnosis, and radical surgical treatment
is unsuitable for this group of patients. Consequently, the 5-year
survival rate decreases to 5% (2).
Therefore, access to tumor markers of early gastric carcinoma with
high sensitivity and specificity is of great importance for the
improvement of the diagnostic and survival rates of gastric
carcinoma.
MicroRNAs (miRNAs), a type of highly conserved
non-protein-coding endogenous small RNAs, bind to the
3′-untranslated region area of the target mRNAs by specific base
pairing, which result in the degradation of target mRNAs or
inhibition of protein translation. miRNAs participate in
posttranscriptional regulation and repress the expression of target
genes. miRNAs exhibit tissue specificity and are abnormally
expressed in the majority of tumors, including gastric, colorectal
and pancreatic carcinoma (3).
miR-155 is a type of miRNA, the expression of which
is increased in gastric carcinoma tissues compared with adjacent
tissues, and has been associated with lymphatic metastasis
(4,5).
Previous studies investigating the effect of miR-155 on the
behavior of tumor cells have focused primarily on proliferation and
apoptosis, as opposed to metastasis and invasion of tumor cells.
Concurrently, there has been a study suggesting that signal
transduction and activators of transcription 3 (STAT3) binds to the
promoter of miR-155, and that small hairpin RNA of STAT3 is able to
downregulate the expression of miR-155 (6). STAT3, and its phosphorylated form, have
been reported to be highly expressed or exhibit increased levels of
activity in a number of human malignancies, including gastric
carcinoma (7). The expression of
phosphorylated (p)-STAT3 has also been closely associated with the
metastasis, invasion and prognosis of gastric carcinoma (8–12). It has
also been reported that miR-155 is able to regulate the metastasis
and invasion of Panc-1 and Capan-2 pancreatic carcinoma cells, and
the proliferation and invasion of Hep-2 cells in hepatic carcinoma,
through the STAT3 signaling pathway (13,14). It
has also been demonstrated that miR-155 may affect the metastasis
and invasion of gastric carcinoma cells (15).
Therefore, based on these aforementioned findings,
the present study detected the expression of miR-155 in gastric
carcinoma cell lines and normal gastric epithelium cell lines using
reverse transcription-polymerase chain reaction (RT-PCR), and
transfected a miR-155 inhibitor into gastric carcinoma cells to
investigate the inhibitory effect of miR-155 on the metastasis as
well as invasive ability of gastric carcinoma cells, and to
determine whether this effect was mediated through the STAT3
signaling pathway.
Materials and methods
Cell lines
Human gastric cancer cell lines BGC-823, NCI-N87,
SGC-7901, AGS, MKN-45 and immortalized gastric mucosa epithelial
cell line GES-1 were purchased from The Cell Bank of Type Culture
Collection of Chinese Academy of Sciences (Shanghai, China).
Main reagents and instruments
Anti-rabbit matrix metalloproteinase (MMP) 2, MMP9
monoclonal antibodies were purchased from Epitomics Inc., Abcam
(Cambridge, MA, USA). Anti-rabbit STAT3, p-STAT3, vascular
endothelial growth factor (VEGF) and suppressor of cytokine
signaling 1 (SOCS1) polyclonal antibodies were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA); MTT was obtained
from Gibco; Thermo Fisher Scientific, Inc. (Waltham, MA, USA);
fetal bovine serum (FBS) and Dulbecco's modified Eagle's medium
(DMEM) culture medium were purchased from Hyclone; GE Healthcare
Life Sciences (Logan, UT, USA) and Transwell Chambers from Corning
Incorporated (Corning, NY, USA); mini double vertical
electrophoresis apparatus, mini transfer electrophoresis apparatus,
ChemiDoc™ XRS Gel Imaging System were purchased from
Bio-Rad Laboratories, Inc. (Hercules, CA, USA); and TE2000
fluorescence inverted microscope was obtained from Nikon
Corporation (Tokyo, Japan).
RT-PCR validation of miR-155
expression
Total RNA was extracted using the TRIzol®
reagent kit (Invitrogen; Thermo Fisher Scientific, Inc.) in the
presence of RNAse inhibitory reagents from the cells of BGC-823,
NCI-N87, SGC-7901, AGS, MKN-45 and GES-1. Primer sequences are as
follows: miR-155 forward, 5′-GTCGTATCCAGTGCAGGGTCCGAGG-3′; reverse,
5′-TATTCGCACTGGATACGACCCCCTA-3′; GADPH forward,
5′-AGCCACATCGCTCAGACA-3′; reverse, 5′-TGGACTCCACGACGTACT-3′. The
RNA was reverse transcribed into cDNA 42°C for 10 min (HiFiScript
cDNA, cat. no. CW2569M; Jjiangsu Kangwei Biotech Co., Ltd.,
Jiangsu, China; http://www.cwbiotech.com/article/list/25.jhtml) and
PCR amplification was conducted using the One-step RT-PCR kit
(UltraSYBR Mixture, cat. no. CW0957M; CWBIO). The primers were
added into a 25 µl PCR reaction system. The thermocycling
conditions were as follows: 94°C for 45 sec, 59°C for 45 sec and
72°C for 60 sec, for 35 cycles and 72°C for 10 min. The relative
expression of miR-155 in each group was detected by RT-qPCR and
calculated with GAPDH as internal reference using the
2−ΔΔCq method (16). A
total of ~5 µl amplification products were used in the next step,
which involved detected of DNA fragments using a 2% agarose gel.
Electrophoresis bands were detected, and the images were captured
using an ultraviolet spectrophotometer.
Cell migration assay
Gastric carcinoma AGS and MKN-45 cells
(2×105 cells) were inoculated in 6-well plates,
respectively. When the confluence of the cells reached 50%, 50 nM
miR-155 inhibitor and miR-155 negative control (NC; Shanghai
Sangong Pharmaceutical Co., Ltd., Shanghai, China) were transfected
into the cells by using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). After transfection for 48 h at
37°C, the cells were digested and added into the upper Transwell
chamber, and the lower chamber continued to be cultured for 24 h in
DMEM medium with 5% with FBS. Then, the Transwell chamber was
removed, washed and fixed with 4% paraformaldehyde at 4°C for 30
min. The cells were stained with 0.1% crystal violet at room
temperature for 5 min, and the number of cells that had passed
through the membrane in five fields of view was counted under an
inverted optical microscope (×200). The migratory ability of the
cells was assessed by calculating the average number of cells per
field of view.
Cell invasion assay
The Matrigel gel was evenly spread on the micro-film
of the Transwell chamber. The remaining steps were the same as the
cell migration assay as mentioned previously. Then, the number of
SGC-7901 cells that had passed through the membrane in five fields
of view were counted under an inverted optical microscope with a
magnification of ×200. The invasive ability of the cells was
assessed by calculating the average number of cells per field of
view.
Western blot analysis
The AGS and MKN-45 cells (2×106 cells)
were inoculated in 6-well plates at 37°C for 2 h. When the
confluence of the cells reached 50%, miR-155 inhibitor and miR-155
NC were transfected into the cells by Lipofectamine 2000. After
transfection for 48 h, the cells were scraped and centrifuged with
2,200 × g for 2 min at room temperature. Subsequent to the addition
of the 200 µl radioimmunoprecipitation assay lysate buffer (cat.
no. 89900; Thermo Fisher Scientific, Inc.), the cells were vortexed
for 30 sec. After 40 min, the cells were centrifuged at 4°C at
11,180 × g for 10 min, and then the supernatant was carefully
removed to obtain the total protein. The protein concentration was
measured using the BCA kit (cat. no. CW0014S; CWBIO). The proteins
were analyzed using 10% SDS-PAGE, and then transferred to a
polyvinylidene fluoride membrane. Following blocking for 2 h at
37°C with 5% skimmed milk powder, the membrane was immersed and
incubated in primary antibody (Stat3 rabbit mAb; cat. no. 12640;
dilution, 1:1,000; Cell Signaling Technology, Inc.) solution
overnight at 4°C and washed by Tris Buffered Saline Tween three
times. Following rinsing, the membrane was immersed and incubated
in the secondary antibody solution (goat anti-rat IgG; cat. no.
ZB-2305; dilution, 1:2,000; OriGene Technologies, Inc., Rockville,
MD, USA) at room temperature for 1–2 h. The membrane was removed
and washed by Tris Buffered Saline Tween for 4 times, and then the
ECL solution (cat. no. RJ239678; Thermo Fisher Scientific, Inc.)
was added. Subsequently, the membrane was exposed in the gel
imaging system (ChemiDoc XRS+; Bio-Rad Laboratories, Inc.). The
gray value of each antibody band was detected with Quantity One
software version 4.62 (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Statistical analysis
Mean and standard deviation (SD) values were used to
summarize continuous variables. Independent t-tests were used to
determine the differences between groups. P<0.05 was considered
to indicate a statistically significant difference. All analyses
were performed using SPSS software, version 17.0 (SPSS, Inc.,
Chicago, IL, USA).
Results
Expression of miR-155 in gastric
carcinoma cells
The expression levels (mean ± SD) of miR-155 in
BGC-823, NCI-N87, SGC-7901, AGS and MKN-45 cells were 0.22±0.03,
0.36±0.03, 0.43±0.02, 0.86±0.05 and 0.94±0.04, respectively, which
were significantly increased compared with the GES-1 cell line
(0.13±0.02; all P<0.05; data not shown), as confirmed by RT-PCR.
AGS and MKN-45 were selected for subsequent experiments due to the
high expression levels of miR-155.
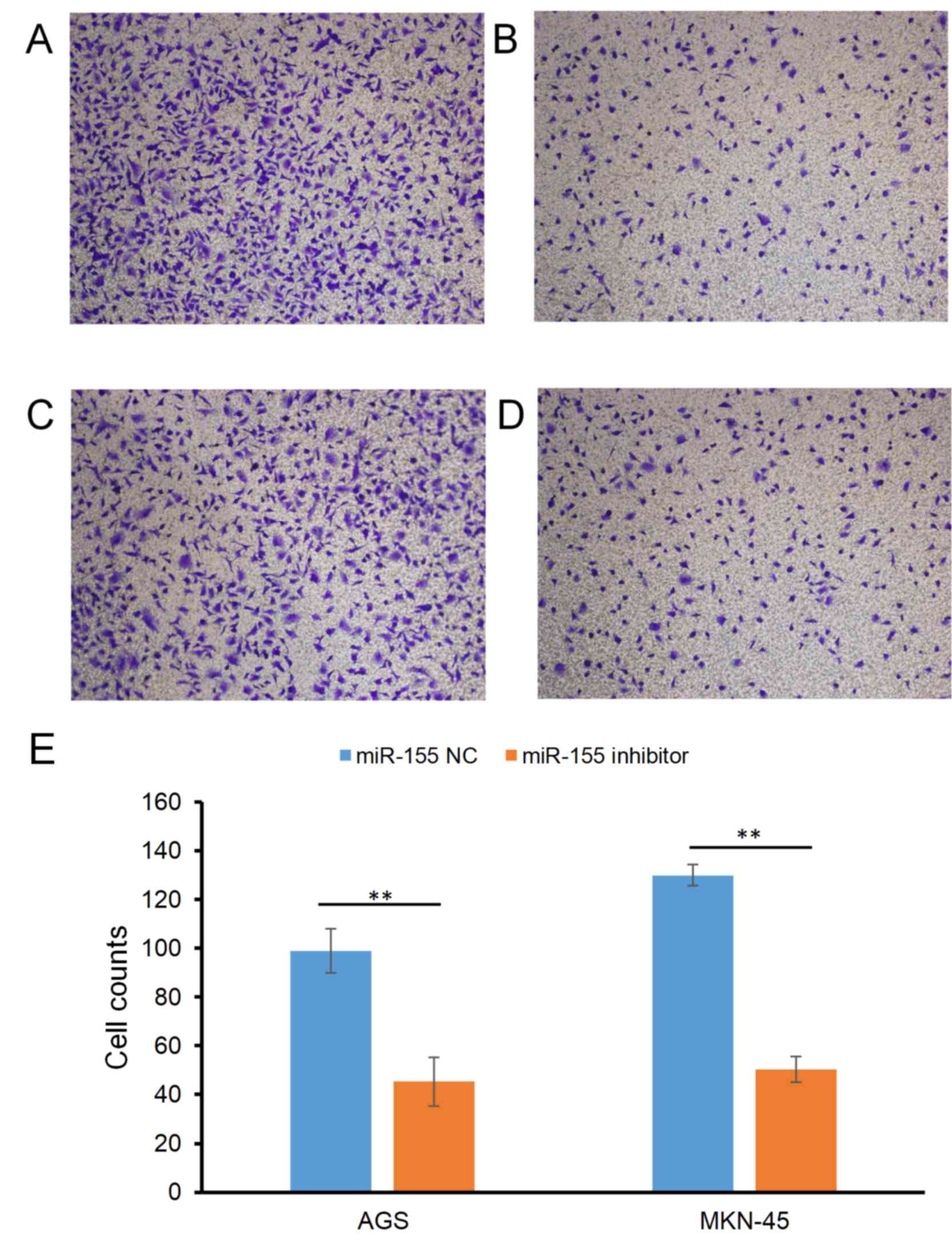
Effect of miR-155 on the migratory
ability of gastric carcinoma cells
The effect of miR-155 on the migration of gastric
carcinoma cells is presented in Fig.
1. The cell counts (mean ± SD) of the AGS cell line were
98.99±9.13 in the miR-155 NC group and 45.32±4.32 in the miR-155
inhibitor group (P<0.01; Fig. 1A and
B); For the MKN-45 cell line, the counts were 129.99±10.12 and
50.36±5.2 in the miR-155 NC and miR-155 inhibitor groups,
respectively (P<0.01; Fig.
1C-E).
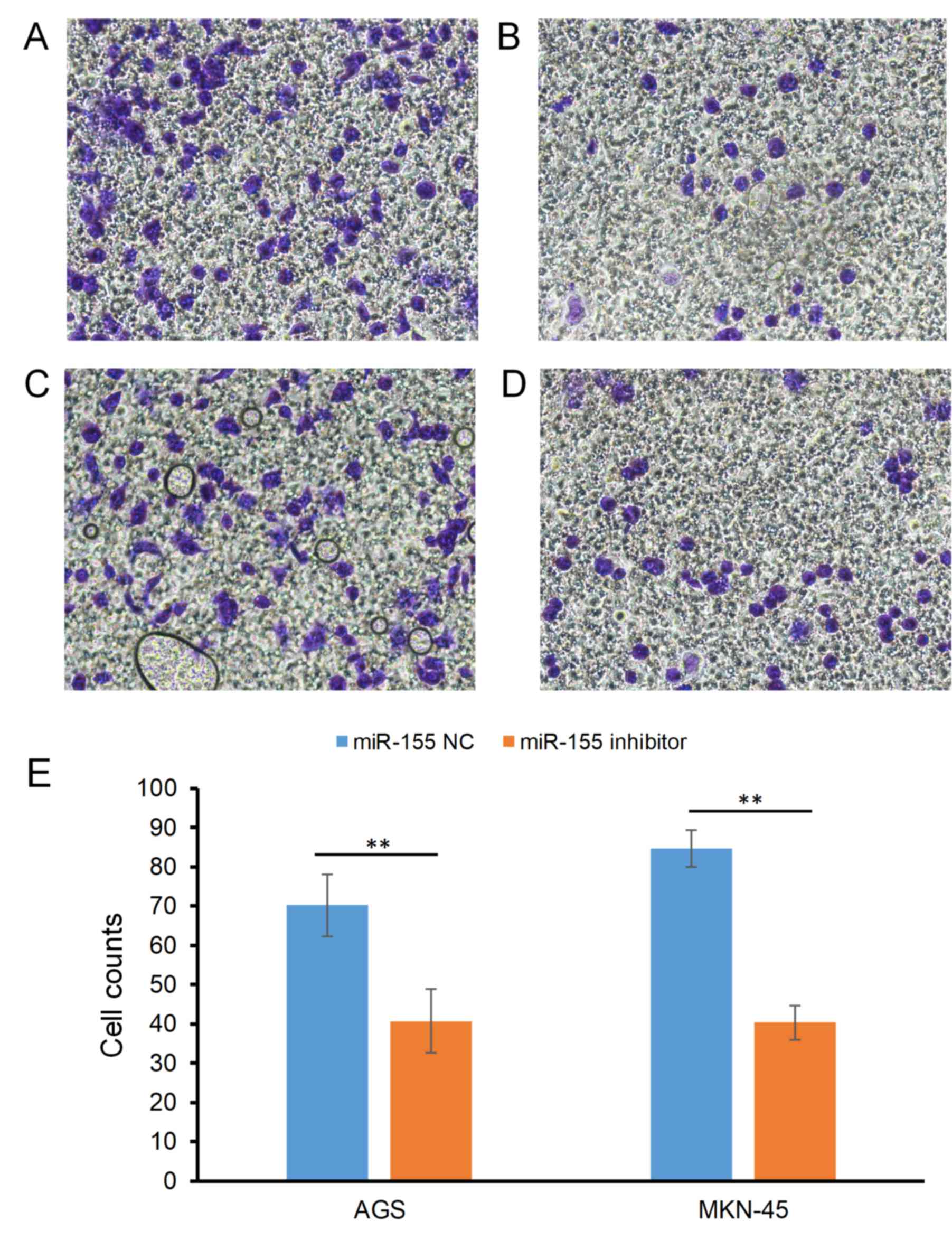
Effect of miR-155 on the invasive
ability of gastric carcinoma cells
The effect of miR-155 on the invasion of gastric
carcinoma cells is indicated in Fig.
2. The cell counts of the AGS cell line were 70.25±7.94 in the
miR-155 NC group and 40.68±4.73 in the miR-155 inhibitor group
(P<0.05; Fig. 2A and B); For the
MKN-45 cell line, the counts were 84.63±8.12 and 40.35±4.29 in the
miR-155 NC and miR-155 inhibitor groups, respectively (P<0.05;
Fig. 2C-E).
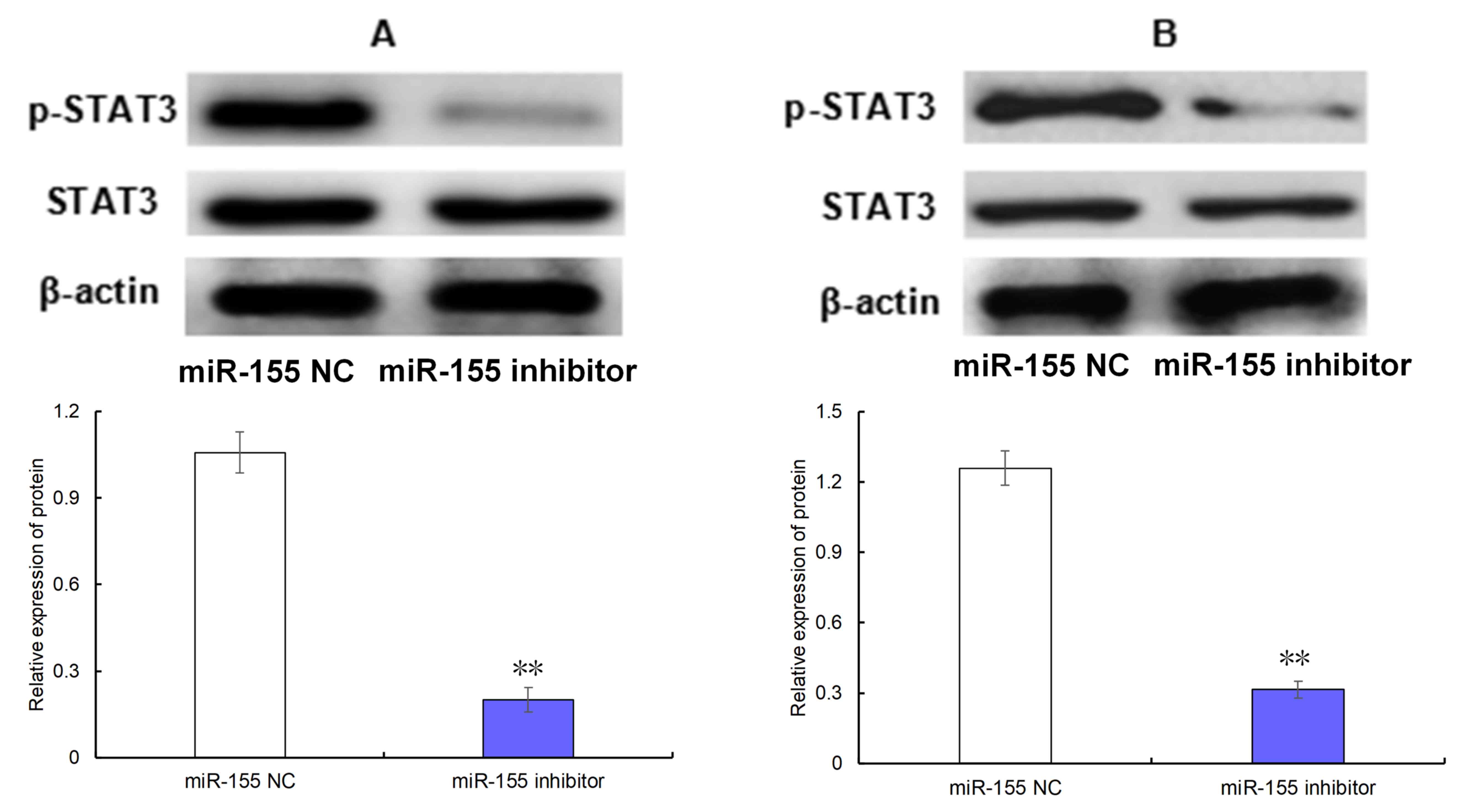
Effect of miR-155 on the level of
phosphorylated (p-)STAT3
As observed in Fig. 3,
transfection of the miR-155 inhibitor was able to significantly
decrease the level of p-STAT3 in AGS (Fig. 3A) and MKN-45 cells (Fig. 3B) compared with the cells transfected
with miR-155 NC (both P<0.05).
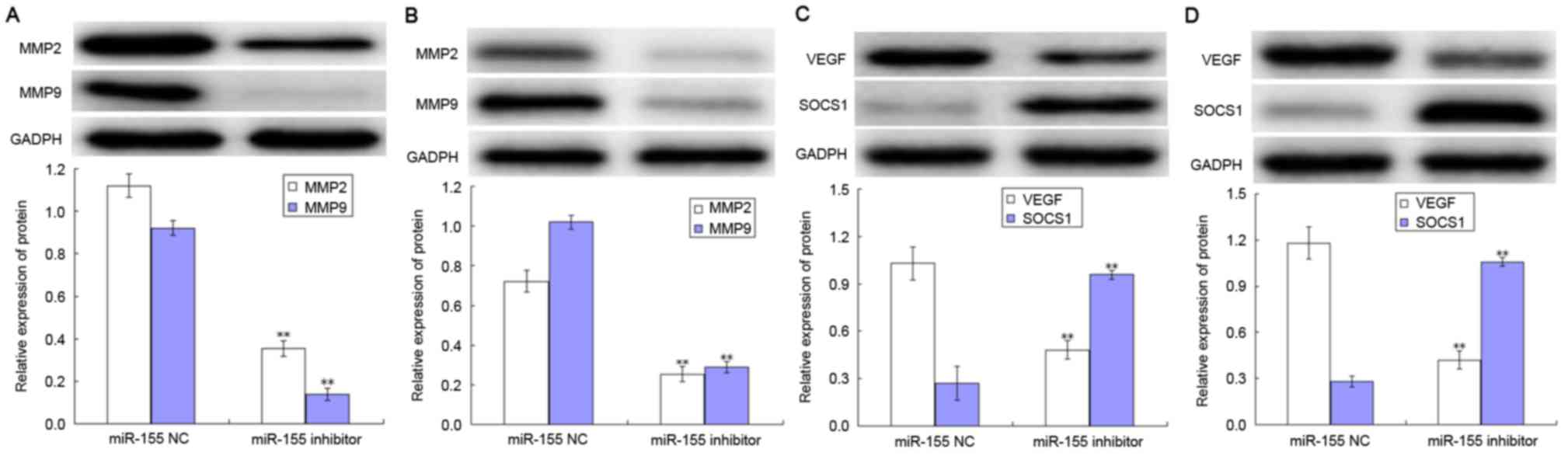
Effect of miR-155 on the expression
levels of MMPs, VEGF and SOCS1
The levels of MMP-2, MMP-9, VEGF and SOCS1
expression in AGS and MKN-45 cells are presented in Fig. 4. The expression levels of MMP2 and
MMP9 were decreased following transfection with miR-155 in the AGS
and MKN-45 cell lines, compared with respective negative control
group (both P<0.05; Fig. 4A and
B). Notably, transfection with the miR-155 inhibitor was able
to decrease the level of VEGF expression in AGS and MKN-45 cells,
whilst increasing the level of SOCS1 expression compared with
negative control group (all P<0.05; Fig. 4C and D).
Discussion
The abnormal expression of miRNA is closely
associated with various types of human tumors (2). miRNA is a key factor which affects the
formation and development of tumors. miRNA is a type of
multifunctioning RNA molecule, located in the non-coding region of
chromosome q21 (17). miRNAs
participate in a number of biological processes, including
hematopoiesis, inflammation and regulation of immune responses
(18,19). Previous studies had hypothesized that
miR-155 exhibited a carcinogenic effect, and high expression of
miR-155 has been reported in breast, hepatic, pancreatic and
colorectal carcinoma (3). Liu et
al (5) observed that the
expression of miR-155 was increased in gastric carcinoma tissues
compared with corresponding non-tumor normal tissues using RT-PCR
analysis. Song et al (4)
additionally verified that the expression of miR-155 was increased
in cases of gastric carcinoma with lymphatic metastasis, and the
expression level of miR-155 was independent of the gender, age,
tumor size, level of invasion, tumor node metastasis (TNM) stage
and vascular invasion, but miR-155 expression was associated with
lymphatic metastasis. Therefore, they concluded that miR-155 is
closely associated with the formation and development of gastric
carcinoma (4). Based on the
aforementioned findings, the present study focused on the
expression of miR-155 in 6 gastric carcinoma (BGC-823, NCI-N87,
SGC-7901, AGS and MKN-45) cell lines and the normal GES-1 cell
line. The results indicated that miR-155 expression was the highest
in MKN-45 cells, which was in accordance with the results of Liu
et al (5) and Song et
al (4). The unfavorable prognosis
and the low five-year survival rate are primarily based on the
invasion and metastasis rates of the carcinoma cells. Therefore,
the present study also investigated the effect of miR-155 on the
invasive and metastatic ability of gastric carcinoma cells.
The active form of STAT3 binds to the gene promoter
of miR-155 in chronic lymphocytic leukemia (6). It was reported that the STAT3 small
hairpin RNA may decrease the expression of miR-155 (6). Huang et al (14) suggested that miR-155 regulated the
migration and invasion of pancreatic carcinoma Panc-1 and Capan-2
cells via the STAT3 signaling pathway. Zhao et al (13) demonstrated that miR-155 promoted the
proliferation and invasion of hepatic carcinoma Hep-2 cells through
increasing the activation of the STAT3 signaling pathway.
Therefore, the present study hypothesized that miR-155 may affect
the migratory and invasive abilities of gastric carcinoma via the
STAT3 signaling pathway. Located on chromosome 12, STAT3 is one of
the members of the STAT family. STAT3 is activated by
phosphorylation of a tyrosine residue, which is induced by the
binding of cytokines or growth factors or activation of oncogenes
(20). STAT3 binds with the tyrosine
residue of p-STAT3 and forms a dimer through the Src homolog 2
domain. Subsequently, the dimer is translocated into the nucleus to
bind to the promoter region of the target genes and regulates the
transcription of these genes. Consequently, proliferation is
promoted and apoptosis is blocked. Other effects include the
induction of immune evasion, promotion of angiogenesis and
induction of invasion and metastasis of tumor cells. STAT3 is
involved in the initiation and development of tumors (9). p-STAT3 is an independent prognostic
factor of gastrointestinal tumors, the expression of which is high
in tumor cells, and is closely associated with low overall survival
and disease-free survival rates (10,12). The
expression of p-STAT3 in gastric carcinoma tissues is significantly
increased compared with adjacent tissues, and is associated with
lymphatic metastasis (8). STAT3
expression increases in severe atypical hyperplasia tissues,
gastric carcinoma tissues and lymph metastasis (21). This finding suggests that there is a
positive correlation between STAT3 and histodifferentiation and
lymphatic metastasis.
A meta-analysis which included 5,757 patients with
gastric cancer indicated that the 5-year survival rate of patients
with high expression levels of STAT3 and MMP-9 was decreased,
compared with the low expression levels of STAT3 and MMP-9 and was
associated with lymphatic metastasis, distant metastasis,
differentiation, tumor size and high TNM stage, suggesting that the
expression level of p-STAT3 is closely associated with the
formation and development of gastric carcinoma, and is positively
correlated with lymphatic metastasis (11). The hypothesis is that the decreasing
the level of p-STAT3 may inhibit the metastasis and invasion of
gastric carcinoma cells. The results of the present study indicated
that decreasing the expression of miR-155 may decrease the level of
p-STAT3, and consequently decreases the metastatic and invasive
abilities of gastric carcinoma AGS and MKN-45 cells. Therefore,
miR-155 affects the metastatic and invasive abilities of gastric
carcinoma cells via the STAT3 signaling pathway.
The STAT3 signaling pathway is regulated by negative
feedback by SOCS (22). SOCS1 binds
to Janus kinase and then inhibits its activity, which results in
the decrease of STAT3 activity or the level of p-STAT3. Souma et
al (23) demonstrated that
adenovirus-expressing SOCS1 may decrease the level of p-STAT3 in
gastric carcinoma NUGC-3 and AGS cell lines and inhibit the
proliferation and metastasis of carcinoma cells. miR-155 may
promote the proliferation and invasion of Hep-2 hepatic carcinoma
cells by increasing the expression of STAT3 and decreasing the
expression of SOCS1 (13). The result
of the present study indicated that decreasing the expression of
miR-155 in gastric carcinoma cell lines may significantly increase
the expression of SOCS1, suggesting that miR-155 inhibition weakens
the invasive and metastatic abilities of AGS and MKN-45 cells by
increasing the expression of SOCS1 and decreasing the level of
p-STAT3.
STAT3 promotes the metastasis, angiogenesis and
invasion of tumors by increasing the expression of B-cell lymphoma
2-like protein 1, myeloid cell leukemia 1, survivin, cyclinD1, VEGF
and MMP-2 (24–27). Niu et al (25) and Wei et al (24) revealed the presence of STAT3 binding
sites on the VEGF promoter, and that a mutation in that site
resulted in a loss of activity of the VEGF promoter, which is
mediated by STAT3. This finding suggested that VEGF was a
downstream target gene of STAT3.
The formation and development of tumor depends on
the nutrient supply from angiogenesis (28). VEGF serves a pivotal role in that
process, and also mediates the invasion and metastasis of tumor
cells. The expression of STAT3, p-STAT3 and VEGF-D in gastric
carcinoma tissues was increased compared with adjacent tissues and
GES-1 (29). The downregulated
expression of STAT3 may decrease the expression of VEGF-D and
thereby inhibit lymphatic metastasis. Xie et al (27) reported the presence of STAT3 binding
sites in the MMP2 promoter. It was demonstrated that STAT3 was able
to increase the expression of MMP2, to promote the invasion and
metastasis of melanoma cells in nude mice (30).
MMPs are the most important proteinase in this
process, as they are the key enzymes involved in the invasion and
metastasis of tumor cells (31).
MMP-2 is able to digest the collagen IV component of gelatin in
extracellular matrix to assist tumor cells to invade through the
damaged basement membrane (32).
MMP-2 is also able to induce capillary hyperplasia, which is
characteristic of tumor cell invasion and metastasis. MMP-9 is a
type of proteolytic enzyme secreted by a various types of cells,
and it has one of the highest molecular weight among all members of
the MMP family (31). MMP-9 digests
the extracellular matrix and basement membrane, which consequently
increases the motility ability of cells and promotes the spread and
metastasis of tumor cells (31).
There is a positive association between the expression of p-STAT3
in SNU-638 and MKN1 cells, and MMP9, suggesting that decreasing the
level of phosphorylated STAT3 may decrease the expression of VEGF,
MMP2 and MMP9, thereby inhibiting the invasion and metastasis of
gastric carcinoma cells (33). The
results of the present study indicate that the downregulation of
miR-155 expression in gastric carcinoma cell lines was able to
significantly decrease the expression of VEGF, MMP2 and MMP9,
thereby inhibiting the invasion and metastasis of gastric carcinoma
cells.
In conclusion, miR-155 is highly expressed in
gastric carcinoma cell lines. The downregulation of miR-155
expression may significantly decrease the level of p-STAT3 and the
expression of VEGF, MMP2 and MMP9, as well as increasing the
expression of SOCS1.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Ethics approval and consent to
participate
Not applicable.
Authors' contributions
ZMS conceived and designed the experiments. Data
collection and experiments were performed by HW, YL and QN. HW and
YL analyzed the data and all authors contributed to the writing of
the manuscript.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Xia Y, Zhang M, Zhang X and Liu X: A
systematic review and meta-analysis of runt-related transcription
factor 3 gene promoter hypermethylation and risk of gastric cancer.
J Cancer Res Ther. 10 Suppl:S310–S313. 2014. View Article : Google Scholar
|
|
2
|
Jiang C, Chen X, Alattar M, Wei J and Liu
H: MicroRNAs in tumorigenesis, metastasis, diagnosis and prognosis
of gastric cancer. Cancer Gene Ther. 22:291–301. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wu LT, Cui L, Wang YL and Huang LM:
Research progress on miR-155, EMT and tumour invasion/metastasis.
Biotechnol Bull. 30:55–59. 2014.
|
|
4
|
Song J, Wang H, Fu HX, Meng S, Xu YX and
Xu W: MiR-155 expression in gastric cancer tissues and its clinical
significance. Chin J Gen Surg. 21:1236–1239. 2012.(In Chinese).
|
|
5
|
Liu L, Chen Q, Lai R, Wu X, Wu X, Liu F,
Xu G and Ji Y: Elevated expression of mature miR-21 and miR-155 in
cancerous gastric tissues from Chinese patients with gastric
cancer. J Biomed Res. 24:187–197. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li P, Grgurevic S, Liu Z, Harris D,
Rozovski U, Calin GA, Keating MJ and Estrov Z: Signal transducer
and activator of transcription-3 induces microRNA-155 expression in
chronic lymphocytic leukemia. PLoS One. 8:e646782013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ji K, Zhang L, Zhang M, Chu Q, Li X and
Wang W: Prognostic value and clinicopathological significance of
p-stat3 among gastric carcinoma patients: A systematic review and
meta-analysis. Medicine (Baltimore). 95:e26412016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang XM, Zhou C, Gu H, Yan L and Zhang
GY: Correlation of RKIP, STAT3 and cyclin D1 expression in
pathogenesis of gastric cancer. Int J Clin Exp Pathol. 7:5902–5908.
2014.PubMed/NCBI
|
|
9
|
Li MX, Bi XY, Huang Z, Zhao JJ, Han Y, Li
ZY, Zhang YF, Li Y, Chen X, Hu XH, et al: Prognostic role of
phospho-STAT3 in patients with cancers of the digestive system: A
systematic review and meta-analysis. PLoS One. 10:e01273562015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Inokuchi M, Murayama T, Hayashi M, Takagi
Y, Kato K, Enjoji M, Kojima K, Kumagai J and Sugihara K: Prognostic
value of co-expression of STAT3, mTOR and EGFR in gastric cancer.
Exp Ther Med. 2:251–256. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen J, Liu X, Jiao H, Peng L, Huo Z, Yang
W, Shen Q, Li T and Liu Q: Prognostic and clinical significance of
STAT3 and MMP9 in patients with gastric cancer: A meta-analysis of
a Chinese cohort. Int J Clin Exp Med. 8:546–557. 2015.PubMed/NCBI
|
|
12
|
Liu Y, Deng J, Luo X, Pan Y, Zhang L,
Zhang R and Liang H: Overexpression of SMYD3 was associated with
increased STAT3 activation in gastric cancer. Med Oncol.
32:4042015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao XD, Zhang W, Liang HJ and Ji WY:
Overexpression of miR-155 promotes proliferation and invasion of
human laryngeal squamous cell carcinoma via targeting SOCS1 and
STAT3. PLoS One. 8:e563952013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang C, Li H, Wu W, Jiang T and Qiu Z:
Regulation of miR-155 affects pancreatic cancer cell invasiveness
and migration by modulating the STAT3 signaling pathway through
SOCS1. Oncol Rep. 30:1223–1230. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wan J, Xia L, Xu W and Lu N: Expression
and function of miR-155 in diseases of the gastrointestinal tract.
Int J Mol Sci. 17:E7092016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Seddiki N, Brezar V, Ruffin N, Lévy Y and
Swaminathan S: Role of miR-155 in the regulation of lymphocyte
immune function and disease. Immunology. 142:32–38. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jurkovicova D, Magyerkova M, Kulcsar L,
Krivjanska M, Krivjansky V, Gibadulinova A, Oveckova I and Chovanec
M: miR-155 as a diagnostic and prognostic marker in hematological
and solid malignancies. Neoplasma. 61:241–251. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Buettner R, Mora LB and Jove R: Activated
STAT signaling in human tumors provides novel molecular targets for
therapeutic intervention. Clin Cancer Res. 8:945–954.
2002.PubMed/NCBI
|
|
21
|
Zhang L, Li J, Wang Q, Meng G, Lv X, Zhou
H, Li W and Zhang J: The relationship between microRNAs and the
STAT3-related signaling pathway in cancer. Tumour Biol.
39:10104283177198692017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alexander WS, Starr R, Metcalf D,
Nicholson SE, Farley A, Elefanty AG, Brysha M, Kile BT, Richardson
R, Baca M, et al: Suppressors of cytokine signaling (SOCS):
Negative regulators of signal transduction. J Leukoc Biol.
66:588–592. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Souma Y, Nishida T, Serada S, Iwahori K,
Takahashi T, Fujimoto M, Ripley B, Nakajima K, Miyazaki Y, Mori M,
et al: Antiproliferative effect of SOCS-1 through the suppression
of STAT3 and p38 MAPK activation in gastric cancer cells. Int J
Cancer. 131:1287–1296. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wei D, Le X, Zheng L, Wang L, Frey JA, Gao
AC, Peng Z, Huang S, Xiong HQ, Abbruzzese JL and Xie K: Stat3
activation regulates the expression of vascular endothelial growth
factor and human pancreatic cancer angiogenesis and metastasis.
Oncogene. 22:319–329. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Niu G, Wright KL, Huang M, Song L, Haura
E, Turkson J, Zhang S, Wang T, Sinibaldi D, Coppola D, et al:
Constitutive Stat3 activity up-regulates VEGF expression and tumor
angiogenesis. Oncogene. 21:2000–2008. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li S, Priceman SJ, Xin H, Zhang W, Deng J,
Liu Y, Huang J, Zhu W, Chen M, Hu W, et al: Icaritin inhibits
JAK/STAT3 signaling and growth of renal cell carcinoma. PLoS One.
8:e816572013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xie TX, Huang FJ, Aldape KD, Kang SH, Liu
M, Gershenwald JE, Xie K, Sawaya R and Huang S: Activation of stat3
in human melanoma promotes brain metastasis. Cancer Res.
66:3188–3196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gupta MK and Qin RY: Mechanism and its
regulation of tumor-induced angiogenesis. World J Gastroenterol.
9:1144–1155. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Deng J, Cui J, Jiang N, Zhang R, Zhang L,
Hao X and Liang H: STAT3 regulation the expression of VEGF-D in
HGC-27 gastric cancer cell. Am J Transl Res. 6:756–767.
2014.PubMed/NCBI
|
|
30
|
Fofaria NM and Srivastava SK: Critical
role of STAT3 in melanoma metastasis through anoikis resistance.
Oncotarge. 5:7051–7064. 2014. View Article : Google Scholar
|
|
31
|
Deryugina EI and Quigley JP: Tumor
angiogenesis: MMP-mediated induction of intravasation- and
metastasis-sustaining neovasculature. Matrix Biol. 44-46:1–112.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mook OR, Frederiks WM and Van Noorden CJ:
The role of gelatinases in colorectal cancer progression and
metastasis. Biochim Biophys Acta. 1705:69–89. 2004.PubMed/NCBI
|
|
33
|
Yoon J, Cho SJ, Ko YS, Park J, Shin DH,
Hwang IC, Han SY, Nam SY, Kim MA, Chang MS, et al: A synergistic
interaction between transcription factors nuclear factor-κB and
signal transducers and activators of transcription 3 promotes
gastric cancer cell migration and invasion. BMC Gastroenterol.
13:292013. View Article : Google Scholar : PubMed/NCBI
|