Introduction
Esophageal cancer is a common malignant tumor that
occurs frequently in China, the incidence rate ranks fifth and
mortality rate fourth in malignant tumors (1), dominated by squamous carcinoma. At
present, the main clinical treatment methods include surgery,
radiotherapy and chemotherapy. Even so, patients with esophageal
squamous cell carcinoma have not only a poor prognosis, but also a
low 5-year survival rate of less than 15% (2), so study of effective therapeutic drugs
is urgent.
The existing clinical treatment often cause great
adverse reactions, which not only bring great pain to the
patient's, but also seriously reduce their life quality.
Traditional Chinese medicine combined with conventional medicine
has many advantages, and the active ingredients have high
efficiency and low toxicity; traditional Chinese medicine has
attracted increasingly more attention of scientists in China and
worldwide because of these irreplaceable advantages. It has been
reported that the active ingredients in traditional Chinese
medicine and its extracts play significant roles in inhibiting the
proliferation or inducing apoptosis of cancer cells, and reducing
adverse reactions to radiotherapy and chemotherapy (3). Zerumbone, extracted from Zingiber
zerumbeta, has been proven by many studies to have antitumor,
anti-inflammation and other pharmacological activities; in
particular, the anticancer effect of zerumbone and its mechanism
have been investigated by a large number of scientists (4,5).
This study aimed to assess the effects of zerumbone
on the proliferation and apoptosis of esophageal cancer EC-109
cells and the possible relevant mechanism during the apoptosis
process, so as to provide a basis for the clinical treatment of
esophageal cancer with zerumbone.
Materials and methods
Materials and reagents
Esophageal cancer EC-109 cells (Cell Bank of Chinese
Academy of Sciences, Shanghai, China); zerumbone (Sigma, New York,
NY, USA); methyl thiazolyl tetrazolium (MTT) (Sigma, St. Louis, MO,
USA); rabbit anti-human P53 and Bcl-2 poluclonal primary antibodies
and goat anti-rabbit horseradish peroxidase (HRP)-labeled secondary
polyclonal antibody (cat. nos. 10442-1-AP, 12789-1-AP and
SA00001-2; Wuhan Sanying Biotechnology, Wuhan, China; Dulbecco's
modified Eagle's medium (DMEM) (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA); TdT-mediated dUTP nick end-labeling
(TUNEL) apoptosis kit, TRIzol and reverse transcription-polymerase
chain reaction (RT-PCR) kits (Invitrogen; Thermo Fisher Scientific,
Inc.); primer synthesis (Takara, Dalian, China).
Cell culture
The cells were cultured using DMEM cell culture
fluid containing 10% fetal bovine serum, amino acid and double
antibodies (100 kU/l penicillin and 0.1% streptomycin) in an
incubator at 37°C under 5% CO2 and saturated humidity
for continuous subculture. The culture fluid was regularly
replaced, and the cells were digested with trypsin for follow-up
experiments when they grew until covering 80% of the bottom of
bottle.
Detection of cell proliferation
inhibition rate via Cell Counting Kit-8 (CCK-8) method
The esophageal cancer EC-109 cells in logarithmic
growth phase was taken and counted using the cell counter after
digestive treatment. Single-cell suspension (100 µl)
(1×105) was added into the 96-well plate. After the
cells adhered to the wall, the original culture fluid was removed
and the drugs with corresponding concentration were added according
to the experimental grouping, and the volume was 100 µl. After 24,
48 and 72 h, 10 µl CCK-8 solution was added into the 96-well plate.
After 4 h, the optical density (OD) value at the wavelength of 450
nm of each well was measured using the microplate reader.
Inhibition rate = (1 - OD value in experimental group/OD value in
blank control group) × 100%.
Detection of apoptosis of EC-109 via
TUNEL method
EC-109 cells were inoculated in a laser confocal
culture dish and fixed with 4% paraformaldehyde for 15 min after
drug treatment. After being washed with phosphate buffered saline
(PBS) for 5 min ×2 times, tissues were treated with 100 µl 20 µg/ml
proteinase K at room temperature for 10–30 min, and then washed
with PBS for 3 min. Cells in treatment group were mixed evenly with
1 µl rTdT + 1 µl biotin-labeled dUTP + 98 µl equilibrium liquid;
rTdT was replaced with tri-distilled water in negative control
group; 100 µl DNase I buffer was added into the positive control
group for incubation for 5 min, and 100 µl DNase I (10 U/ml) was
added for enzyme digestion for 10 min after the liquid was removed;
finally, cells were rinsed with deionized water 4 times and washed
with PBS for 5 min; 100 µl TUNEL reaction mixture was added onto
the specimen, and the specimen was covered with cover glass or
sealed with sealing film for reaction in the dark wet box at 37°C ×
1 h; enzyme-labeled reaction: 100 µl streptavidin-labeled HRP
(diluted by 1:500 PBS) was added for 30 min. Finally, cells were
observed under a laser confocal microscope (Olympus Corporation,
Tokyo, Japan), and photographed in the randomly-selected visual
field.
Detection of the mRNA expression of
P53 and Bcl-2 via RT-PCR
The cultured cells were inoculated into the 6-well
plate with 104 cells per well. After 24 h, the
supernatant was discarded and zerumbone in the concentration of 30,
40 and 50 µg/ml was administered, respectively. After the reaction
for 48 h, the EC-109 cells in each group were collected and used
for the total RNA extraction with TRIzol (Invitrogen; Thermo Fisher
Scientific, Inc.). Then cDNA was synthesized via reverse
transcription with the qualified total RNA as the template. The
specific reaction conditions were as follows: incubation at 42°C
for 15 min and incubation at 95°C for 3 min. Then, cDNA was cooled
on ice and stored at −80°C for later use. The qPCR amplification
system is 25 µl: SYBR Premix Ex Taq II (2×) 12.5 µl, PCR Forward
Prime (10 µM) 1 µl, DNA Templates 2 µl, DDW 8.52 µl. The reaction
conditions are as follows: denatured at 95°C for 30 sec and
annealed at 55°C for 60 sec, extension at 72°C, 30 cycles. Routine
amplification was performed according to the primer sequences in
Table I.
 | Table I.RT-PCR primer sequences of Fas and
FasL mRNA. |
Table I.
RT-PCR primer sequences of Fas and
FasL mRNA.
| Gene name | Primer sequence |
|---|
| P53 | Forward:
5′-GGAAATCTCACCCCATCCCA-3′ |
|
| Reverse:
5′-CAGTAAGCCAAGATCACGCC-3′ |
| Bcl-2 | Forward:
5′-GGCCTGTGTCTCCTTGTGAT-3′ |
|
| Reverse:
5′-TGCCAGCTCCTTCTGAAGTA-3′ |
Detection of the protein expression of
P53 and Bcl-2 via western blotting
The cultured cells were inoculated into the 6-well
plate with 104 cells per well. After 24 h, the
supernatant was discarded and zerumbone in the concentration of 30,
40 and 50 µg/ml was administered, respectively. After the reaction
for 48 h, the cells in each group were collected and the total
protein was extracted and the protein concentration was determined.
After the samples were treated, 50 µg protein was isolated via
sodium dodecyl sulfate polyacrylamide gel electrophoresis
(SDS-PAGE) and the protein isolated was electronically transferred
onto the polyvinylidene fluoride (PVDF) membrane. The membrane was
sealed using the blocking solution at room temperature for 1 h and
incubated with primary antibodies (1:500) at 4°C overnight. After
the membrane was fully washed with Tween Tris base buffer solution
(TTBS), the secondary antibody (1:2,000) was added for incubation
at room temperature for 1 h, followed by washing with TTBS, color
development and photography.
Statistical analysis
The data were presented as mean ± standard
deviation, and processed with SPSS 19.0 (SPSS, Inc., Chicago, IL,
USA). One-way analysis of variance (ANOVA) was used for the
statistical analysis of data and the post hoc test was Dunnett's
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effect of zerumbone on the inhibition
of EC-109 proliferation
Compared with that in control group, the activity of
EC-109 cells could be inhibited in administration groups with the
zerumbone concentration of 30, 40 and 50 µmol/l. With the increase
of concentration and time, the inhibition rate of proliferation was
significantly increased in a significant concentration-dependent
manner. The results in this study showed that the inhibition rate
of EC-109 growth and proliferation was 49.82% under the action of
zerumbone in the concentration of 40 µmol/l for 48 h. Therefore, 40
µmol/l and 48 h were selected as the administration concentration
and dosage in this experiment (Table
II).
| Table II.Effects of zerumbone in different
concentrations on the inhibition of EC-109 proliferation (mean ±
SD, %). |
Table II.
Effects of zerumbone in different
concentrations on the inhibition of EC-109 proliferation (mean ±
SD, %).
|
| Proliferation
inhibition rate (%) |
|---|
| Concentration
(µmol/l) | 24 h | 48 h | 72 h |
|---|
| Control group | 0 | 0 | 0 |
| 30 |
3.35±0.31a |
16.21±2.31a |
28.25±3.43a |
| 40 |
6.68±1.52a |
49.82±3.58a |
59.43±4.25a |
| 50 |
15.54±1.12a |
59.77±3.56a |
70.53±4.79a |
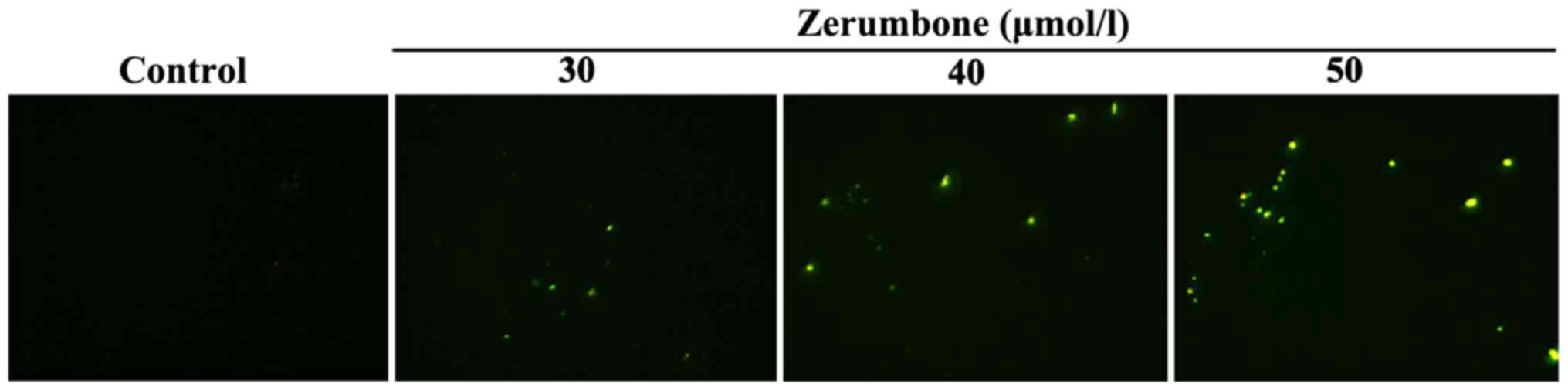
Effect of zerumbone on apoptosis of
EC-109 cells
Compared with that in control group, the yellow
green-stained cells in nucleus were apoptotic cells under the
action of zerumbone in the concentration of 30, 40 and 50 µmol/l
for 48 h. The results revealed that the number of apoptotic cells
increased gradually with the increase of administration
concentration (Fig. 1).
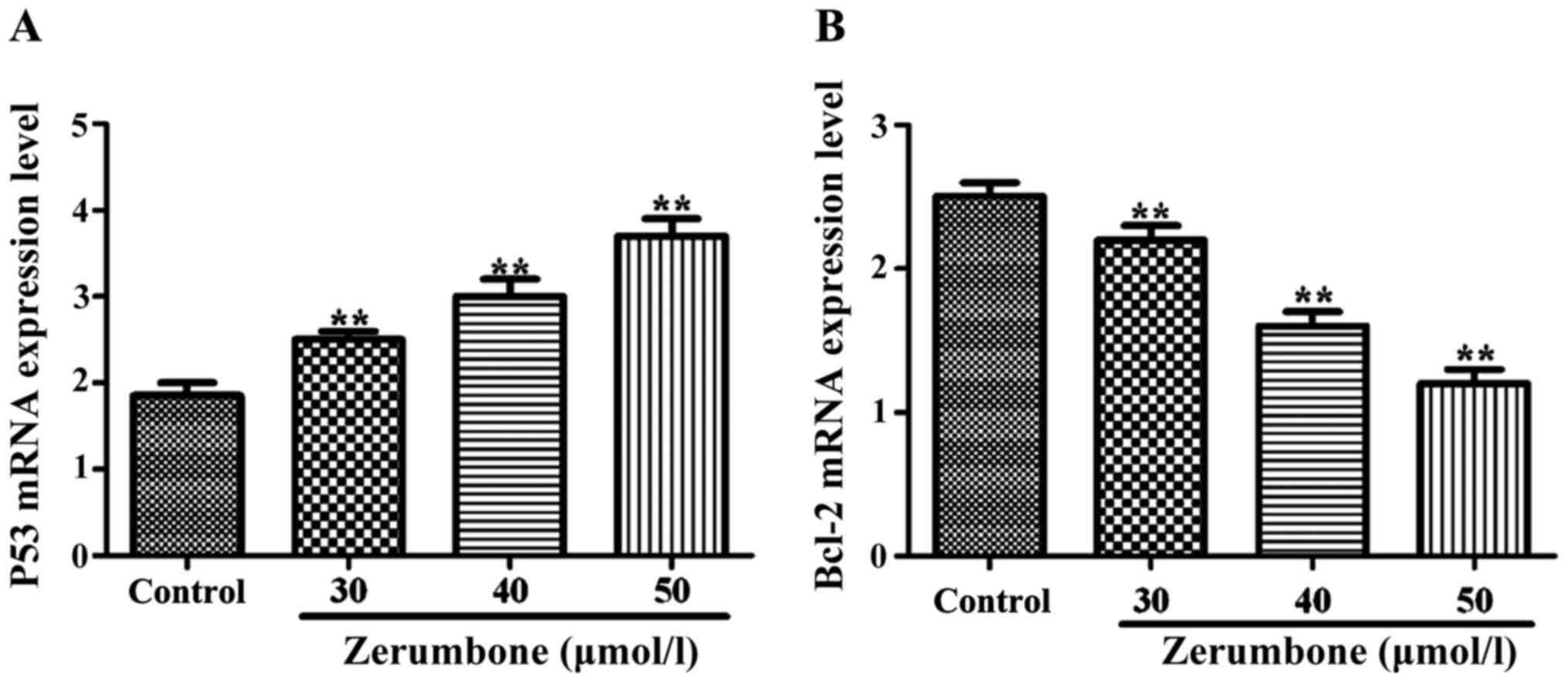
Effects of zerumbone on mRNA levels of
P53 and Bcl-2
Compared with those in control group, under the
action of zerumbone in the concentration of 30, 40 and 50 µmol/l
for 48 h, the mRNA expression level of P53 in each group was
obviously increased (P<0.05), but the mRNA expression level of
Bcl-2 was significantly decreased (P<0.01) in a significant
concentration-dependent manner (Fig.
2).
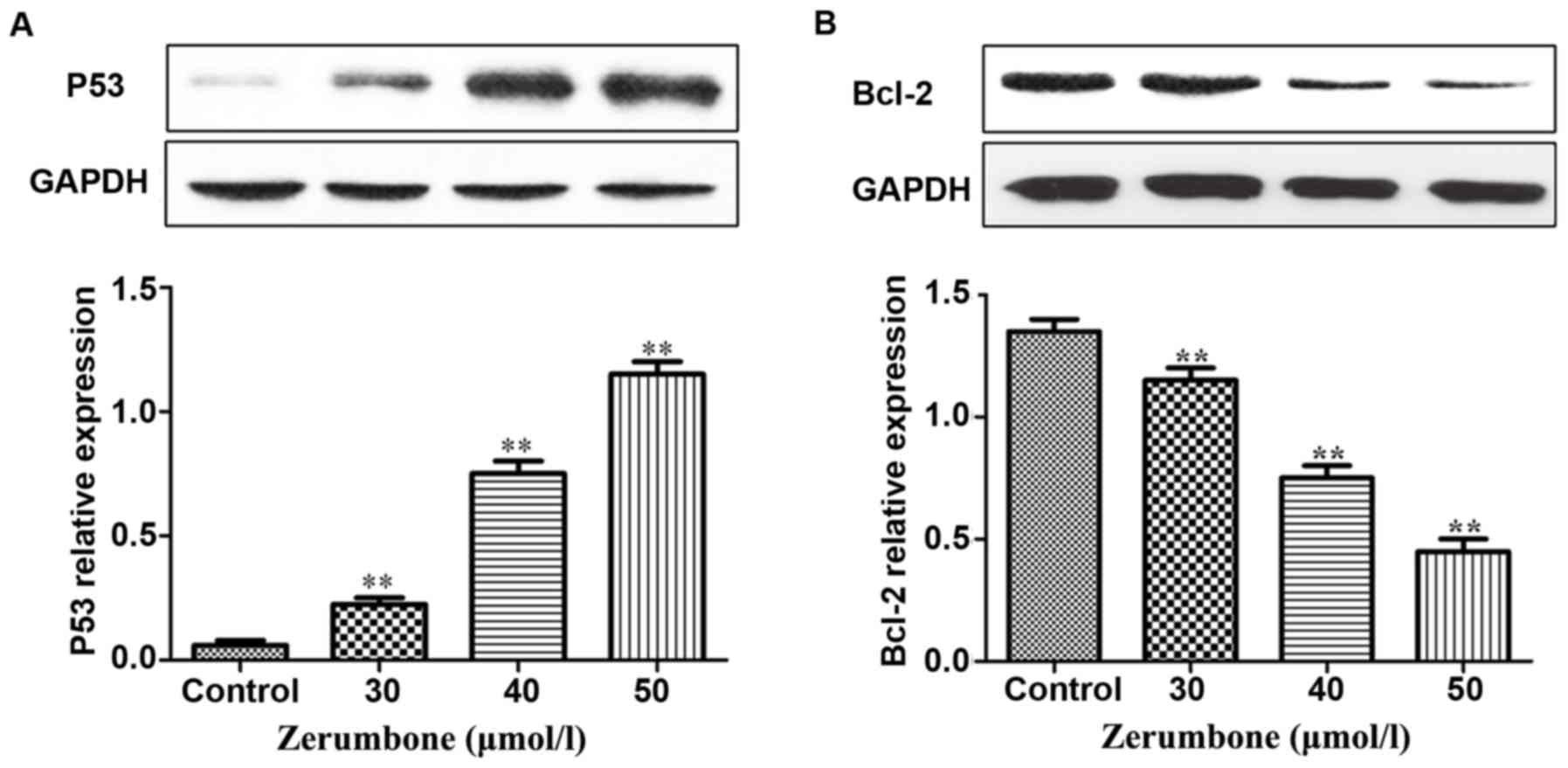
Effects of zerumbone on protein
expression levels of P53 and Bcl-2
Compared with those in control group, under the
action of zerumbone in the concentration of 30, 40 and 50 µmol/l
for 48 h, the protein expression level of P53 in each group was
remarkably increased with the increase of zerumbone concentration,
but the protein expression level of Bcl-2 was remarkably decreased
in a significant concentration-dependent manner (Fig. 3).
Discussion
Esophageal cancer has a high incidence rate in
certain regions in China, and the clinical surgical treatment,
combined with radiotherapy and chemotherapy, is still the preferred
choice for the treatment of esophageal cancer, but its curative
effect still needs improvement. Zerumbone is a kind of
sesquiterpene substance extracted from the rhizome of wild
Zingiber zerumbeta with antitumor, anti-inflammation and
other pharmacological activities. It has been reported that
zerumbone is effective for many cancer cells, such as colon cancer,
lung cancer (6), leukemia (7) and liver cancer (8), but there is little research on the
biological activity of zerumbone on esophageal cancer. In this
experiment, the effects of zerumbone on the proliferation and
apoptosis of esophageal cancer EC-109 cells and the possible
mechanism in apoptosis process were studied preliminarily.
The wild-type P53 gene can monitor the abnormalities
of cell genome and play a negative regulatory role in the cell
growth process. When the DNA in cells or cells tend to be
cancerous, the P53 gene can remove them in time, thus playing an
anticancer effect (9–11); but if the P53 gene mutation occurs, it
will lose such a regulatory effect and mutate from the cancer
cell-removing gene into the oncogene, thus being transformed from
normal cells into cancer cells. Studies have shown that P53 gene
locus mutation occurs in more than 60% of gastric cancer; the
wild-type P53 protein is unstable in the normal physiological
conditions, whose half-life is very short, so it is difficult to be
detected via ordinary immunological detection. On the contrary,
after the P53 protein mutation, its half-life is extended, and its
stability is also increased significantly, so that the
immunological detection can detect the mutant P53 protein (12–14). A
large number of studies have found that the P53 protein is
overexpressed in colorectal cancer, lung cancer, gastric cancer,
liver cancer and other tumor cells; moreover, the P53 gene is also
widely used in the gene therapy at present (15–18). Bcl-2
gene has the biological function of inhibiting apoptosis (19), and plays an important role in the
mechanism of apoptosis, which can protect cells from various forms
of death and improve cell survival, thus increasing the number of
cells. In some tumor cells, when the expression of Bcl-2 gene is
inhibited, it will cause tumor cell apoptosis (20), indicating that Bcl-2 gene has a very
close relationship with tumors.
The results of CCK-8 showed that different
concentrations of zerumbone could inhibit the activity of EC-109
compared with that in control group, and the proliferation
inhibition rate was significantly increased with the increase of
concentration in a concentration-independent manner. TUNEL staining
revealed that the cell apoptosis began to occur gradually in the
administration group, and the number of apoptotic cells was
increased with the increase of concentration in a
concentration-independent manner. The results of RT-PCR detection
showed that with the increase of zerumbone concentration, the mRNA
expression level of P53 was gradually increased, but that of Bcl-2
was gradually decreased. Moreover, the results of western blotting
showed that with the increase of zerumbone concentration, the
protein expression level of P53 was gradually increased, but that
of Bcl-2 was gradually decreased, suggesting that zerumbone can
upregulate the protein expression of P53 and downregulate the
protein expression of Bcl-2, thus inducing apoptosis. Similar to
this study, antitumor drugs significantly increase the expression
of P53 in lung cancer cells (21). Xi
et al (22) studied and
revealed that some antitumor drugs can induce tumor cell apoptosis
through downregulating the Bcl-2 expression. For example, carnosol
can decrease the Bcl-2 protein expression in leukemia cells by
34–53%.
In conclusion, it was proved in this study that
zerumbone can inhibit the proliferation of esophageal cancer EC-109
cells and induce the occurrence of apoptosis. Moreover, its
induction of apoptosis may be realized by upregulating the
expression of P53 and downregulating the expression of Bcl-2.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SM was a major contributor in writing the manuscript
and CCK-8 test. YL made contributions to cell culture. LZ helped
with data analysis. JW performed TUNEL method. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, He Y, Zheng R, Zhang S, Zeng H,
Zou X and He J: Esophageal cancer incidence and mortality in China,
2009. J Thorac Dis. 5:19–26. 2013.PubMed/NCBI
|
|
2
|
Jemal A, Murray T, Ward E, Samuels A,
Tiwari RC, Ghafoor A, Feuer EJ and Thun MJ: Cancer statistics,
2005. CA Cancer J Clin. 55:10–30. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Olaku O and White JD: Herbal therapy use
by cancer patients: A literature review on case reports. Eur J
Cancer. 47:508–514. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Murakami A, Tanaka T, Lee JY, Surh YJ, Kim
HW, Kawabata K, Nakamura Y, Jiwajinda S and Ohigashi H: Zerumbone,
a sesquiterpene in subtropical ginger, suppresses skin tumor
initiation and promotion stages in ICR mice. Int J Cancer.
110:481–490. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Abdul AB, Abdelwahab SI, Bin Jalinas J,
Al-Zubairi AS and Taha MM: Combination of zerumbone and cisplatin
to treat cervical intraepithelial neoplasia in female BALB/c mice.
Int J Gynecol Cancer. 19:1004–1010. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim M, Miyamoto S, Yasui Y, Oyama T,
Murakami A and Tanaka T: Zerumbone, a tropical ginger
sesquiterpene, inhibits colon and lung carcinogenesis in mice. Int
J Cancer. 124:264–271. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xian M, Ito K, Nakazato T, Shimizu T, Chen
CK, Yamato K, Murakami A, Ohigashi H, Ikeda Y and Kizaki M:
Zerumbone, a bioactive sesquiterpene, induces G2/M cell cycle
arrest and apoptosis in leukemia cells via a Fas- and
mitochondria-mediated pathway. Cancer Sci. 98:118–126. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sakinah SA, Handayani ST and Hawariah LP:
Zerumbone induced apoptosis in liver cancer cells via modulation of
Bax/Bcl-2 ratio. Cancer Cell Int. 7:42007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wiman KG: Pharmacological reactivation of
mutant p53: From protein structure to the cancer patient. Oncogene.
29:4245–4252. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Farnebo M, Bykov VJ and Wiman KG: The p53
tumor suppressor: A master regulator of diverse cellular processes
and therapeutic target in cancer. Biochem Biophys Res Commun.
396:85–89. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Oren M and Rotter V: Mutant p53
gain-of-function in cancer. Cold Spring Harb Perspect Biol.
2:a0011072010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Machado-Silva A, Perrier S and Bourdon JC:
p53 family members in cancer diagnosis and treatment. Semin Cancer
Biol. 20:57–62. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chari NS, Pinaire NL, Thorpe L, Medeiros
LJ, Routbort MJ and McDonnell TJ: The p53 tumor suppressor network
in cancer and the therapeutic modulation of cell death. Apoptosis.
14:336–347. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vazquez A, Bond EE, Levine AJ and Bond GL:
The genetics of the p53 pathway, apoptosis and cancer therapy. Nat
Rev Drug Discov. 7:979–987. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tang NP, Wu YM, Wang B and Ma J:
Systematic review and meta-analysis of the association between P53
codon 72 polymorphism and colorectal cancer. Eur J Surg Oncol.
36:431–438. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tam CW, Liu VW, Leung WY, Yao KM and Shiu
SY: The autocrine human secreted PDZ domain-containing protein 2
(sPDZD2) induces senescence or quiescence of prostate, breast and
liver cancer cells via transcriptional activation of p53. Cancer
Lett. 271:64–80. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Oguztüzun S, Aydin M, Demirag F, Yazici U,
Ozhavzali M, Kiliç M and Işcan M: The expression of GST isoenzymes
and p53 in non-small cell lung cancer. Folia Histochem Cytobiol.
48:122–127. 2010.PubMed/NCBI
|
|
18
|
Baker L, Quinlan PR, Patten N, Ashfield A,
Birse-Stewart-Bell LJ, McCowan C, Bourdon JC, Purdie CA, Jordan LB,
Dewar JA, et al: p53 mutation, deprivation and poor prognosis in
primary breast cancer. Br J Cancer. 102:719–726. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cory S, Huang DCS and Adams JM: The Bcl-2
family: Roles in cell survival and oncogenesis. Oncogene.
22:8590–8607. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Packham G and Cleveland JL: c-Myc and
apoptosis. Biochim Biophys Acta. 1242:11–28. 1995.PubMed/NCBI
|
|
21
|
Dworakowska D, Jassem E, Jassem J, Boltze
C, Wiedorn KH, Dworakowski R, Skokowski J, Jaśkiewicz K and
Czestochowska E: Prognostic value of cyclin D1 overexpression in
correlation with pRb and p53 status in non-small cell lung cancer
(NSCLC). J Cancer Res Clin Oncol. 131:479–485. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xi S, Dyer KF, Kimak M, Zhang Q, Gooding
WE, Chaillet JR, Chai RL, Ferrell RE, Zamboni B, Hunt J, et al:
Decreased STAT1 expression by promoter methylation in squamous cell
carcinogenesis. J Natl Cancer Inst. 98:181–189. 2006. View Article : Google Scholar : PubMed/NCBI
|