Introduction
Gastric cancer (GC) is the second highest cause of
global disease-associated mortality globally (1). With the progression in surgery resection
and chemotherapy, the clinical result of GC has been enhanced
gradually. However, the 5-year survival rate remains 20–30%
(2). It is assumed that poor
prognosis is associated with the multifaceted and malignant
characteristics of GC (3,4). Previous studies have reported that
various factors are associated with the pathogenesis of GC,
including bad eating habits, gastric ulcers and Helicobacter
pylori infection (5–7). However, the molecular mechanisms of GC
are not fully understood, and include a variety of tumor-associated
factors and genetic modifications of tumor suppressor factors.
Molecular studies investigating alterations in single genes have
provided evidence that GC progresses via different genetic pathways
(8–10). Therefore, the present study aimed to
decipher the molecular mechanism of GC, in order to establish
deeper understanding of GC and identify possible treatments for
patients with GC.
Peroxiredoxins (Prxs) exist in prokaryotes and
eukaryotes, and regulate the redox reaction in the body.
Researchers have demonstrated that Prxs are highly expressed in
different cancer tissues and immortalized cell lines (including
lung, renal and hepatocellular carcinoma cell lines), and promote
the progression of cancer (11,12). High
expression of Prxs is associated with the protection of tumors and
malignancy, which has been associated with resistance of cell lines
against certain chemotherapies and radiotherapies (13,14). As
one of the six components of the peroxiredoxin family, Prx II
serves essential roles in different tumors. Lehtonen et al
(15) reported that the expression of
Prx II was upregulated in bosom carcinoma. Soini et al
(16) demonstrated that the
expression of Prx II was associated with the development of renal
cancer. However, the effect of Prx II expression on GC growth
remains unclear. In the present study, the association between Prx
II expression and GC was investigated using GC tissues and
cells.
Epithelial-mesenchymal-transition (EMT) involves
changes in epithelial cells into ectomesenchymal cells under
particular conditions. It is involved in regulating tissue
development and repairing tissue injuries, and is associated with
the invasion and metastasis of tumors (17). The proteins associated with EMT,
including E-cadherin and N-cadherin, serve an important role in GC
(18,19). Matrix metalloproteinase (MMP)-2 and
MMP-9 are also associated with the metastasis of GC. The current
analysis aimed to investigate the effect of Prx II on GC cell
migration and proliferation by detecting the changes in MMP-2,
MMP-9, E-cadherin and N-cadherin expression in GC following the
downregulation of PrxII. It was revealed that Prx II promoted the
proliferation and migration of GC cells. Thus, Prx II may be a
promising target for treatment in GC.
Materials and methods
Patients and samples
Between January 2009 and December 2010, a total of
116 paraffin-embedded sections were collected from patients who
underwent gastrectomy at the Affiliated Hospital of Xuzhou Medical
College (Xuzhou, China). These samples were made into a tissue
microarray, and the expression of Prx II was investigated using
immunohistochemistry (IHC). The hospital routinely detects the
expression of Ki-67 in GC tissue following surgery. The positive
expression rate of Ki-67 was 62.1% (72/116) in GC tissues, which
indicated the proliferation of GC with higher Ki-67 expression was
significantly increased compared with those with lower Ki-67
expression. Additionally, 45 cases of patients with primary gastric
carcinoma receiving gastrectomy at the Affiliated Hospital of
Xuzhou Medical University between September 2015 and January 2016
were included. The 45 fresh frozen GC tissues and matched adjacent
non-cancerous tissues were utilized for western blot analysis or
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) investigation. The positive expression rate of Ki-67 was
73.3% (33/45) in the GC tissues. None of the patients had been
treated with chemotherapy or radiotherapy prior to the surgery, and
no other malignant tumors were identified. The patient's overall
survival time and disease-free survival time were statistically
analyzed. Overall survival was defined as the duration from the
time of diagnosis to the time of mortality from any cause.
Disease-free survival was defined as the duration from the time of
diagnosis to the time of mortality from disease progression. The
present study was approved by the Ethics Review Board of the
Affiliated Hospital of Xuzhou Medical College. All patients
provided written informed consent.
Cell culture
The human GC cell lines MKN-45, MGC-803 and typical
gastric cells (GES-1) were purchased from the Cell Bank of Chinese
Academy of Sciences (Shanghai, China). The cells were cultured in
the Dulbecco's modified Eagle medium (DMEM) (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) with 100 U/ml penicillin and
100 µg/ml streptomycin, supplemented with 10% heat-inactivated
fetal bovine serum (FBS; Beyotime Institute of Biotechnology,
Nanjing, China). These cells were cultured at 37°C in a humidified
environment with 5% CO2.
RNA interference and transfection
When cells had achieved 70% confluence, small
interfering (si)RNA sequences for Prx II and a scrambled
nonspecific sequence (Shanghai GenePharma Co., Ltd., Shanghai,
China) were transfected using Silentfect reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. The medium was replaced after 6–8 h of transfection in
order to prevent toxicity. The cells were then transfected for 48 h
prior to use in subsequent experiments. The Prx II siRNA sequences
were as follows: Forward primer, 5′-CCUAGAAGCUGAAUAGUGA-3′; reverse
primer, 5′-UCACUAUUCAGCUUCUAGG-3′. The scrambled siRNA sequences
were as follows: Forward primer, 5′-UUCUCCGAACGUGUCACGU-3′; reverse
primer, 5′-ACGUGACACGUUCGGAGAA-3′.
Western blot analysis
Cells or 60–80 mg tissues were lysed using radio
immunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology), and the concentration of protein was determined
using a bicinchoninic acid assay (Beyotime Institute of
Biotechnology). The protein was denatured by boiling at 100°C for 5
min. A total of 25 µg protein of each sample was subjected to
SDS-PAGE (8 and 10% gels) and transferred to polyvinylidene
fluoride membranes. The membranes were blocked with 5% skimmed milk
and Tris-buffered saline with Tween 20 at 37°C for 1.5 h and were
incubated with primary antibodies against Prx II (dilution,
1:3,000; cat no. ab15572; ABclonal Biotech Co., Ltd., Woburn, MA,
USA), E-cadherin, N-cadherin, MMP-2, MMP-9 or β-actin (dilution,
1:5,000; cat nos. 9961S, 5296S, 4061S, 3852S and 8457S,
respectively; Cell Signaling Technology, Inc., Danvers, MA, USA) at
4°C for 12 h, followed by incubation with monoclonal goat
anti-mouse or anti-rabbit fluorophore-conjugated secondary
antibodies (dilution, 1:10,000; cat no. RK-611-130-002; LI-COR
Biotechnology, Lincoln, NE, USA) at 37°C for 1.5 h. Enhanced
chemiluminescence detection reagent (Tanon Science and Technology
Co., Ltd., Shanghai, China) was used to visualize the bands with an
Odyssey® CLx Infrared Imaging system (cat no. 9140-00;
LI-COR Biotechnology).
Immunohistochemical staining
Immunostaining was performed on tissue microarray
samples. The microarray samples were processed in accordance with a
previously described method (20).
The microarrays were incubated with rabbit monoclonal antibody
against human Prx II (1:200; cat no. ab15572; ABclonal Biotech Co.,
Ltd.) for 12 h at 4°C, and biotinylated goat anti-rabbit serum IgG
was utilized as an secondary antibody (dilution, 1:5,000; cat no.
RK-611-130-002; LI-COR Biotechnology) at 37°C for 0.5 h. Following
washing with PBS (2.67 mM KCl, 1.47 mM
KH2PO4, 138.00 mM NaCl and 8.10 Mm
Na2HPO4), the slices were incubated with
diaminobenzidine substrate at 37°C for 2 min. The immunoreactivity
of Prx II was rated as 0, negative (0–10%); 1, weak (11–25%); 2,
moderate (26–50%) or 3, strong (>50%) according to the staining
area of positive tumor cells as previously described (20). Weakness to moderate signals (<50%)
of GC cells were considered as exhibiting a low expression of Prx
II. In contrast, strong staining of GC cells (>50%) was
considered to exhibit a high expression of Prx II. Positive and
negative biopsy samples were used as the positive and negative
control. Sections were observed at magnification, ×400 with a light
BHS/System Living microscope (Olympus Corporation, Tokyo, Japan)
and the outcomes were categorized according to the brown granules
in the cytoplasm.
RT-qPCR
According to the manufacturer's protocol, RNA was
isolated from tissue (Invitrogen; Thermo Fisher Scientific, Inc.).
The content of total RNA was measured and calculated using the
A260/A280 ratio. Then, 3 µg RNA was used for reverse transcription
using the TIANScript First Strand cDNA Synthesis kit (BioTeke
Corporation, Beijing, China) according to the manufacturer's
protocol. The cDNA was amplified using PCR using an ABI 7500 PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.) with
SYBR®Premix Ex Taq™ (Takara Bio Inc., Japan). The 20 µl
reaction comprised 10 µl SYBR Green premix and 10 µl primer mix
(including cDNA). The thermocycling conditions were as follows:
95°C for 30 sec (1 cycle); 95°C for 5 sec and 64°C for 34 sec (40
cycles); 95°C for 15 sec and 60°C for 60 sec, and 95°C for 15 sec
(1 cycle). The expression level of Prx II was normalized to GAPDH
and calculated using the 2−ΔΔCq method (21). The primers used were as follows: Prx
II forward, 5′-GTGTCCTTCGCCAGATCACT-3′ and reverse,
5′-AAACTTCCCCATGCTCGTCT-3′; GAPDH forward,
5′-GCAAATTCCATGGCACCGT-3′ and reverse,
5′-TCGCCCCACTTGATTTTGG-3′.
Cell proliferation assay
Cell Counting kit-8 (CCK-8) reagent was used to
examine cell proliferation (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). MGC-803 and MKN-45 cells were incubated in
96-well plates (3×103 cells/well) (Corning Incorporated,
Corning, NY, USA) in 100 µl DMEM. Following incubation for 24, 48,
72 and 96 h, the supernatant was extracted, and 100 µl serum-free
DMEM and 10 µl CCK-8 solution was added to each well, followed by
further incubation for 2 h at 37°C. SiRNA with an unspecific
scrambled sequence was used as a negative control (NC). SiRNA with
a positive sequence was used as a positive control (siRNA). The
positive sequence for Prx II was: Sense, 5′-CCUAGAAGCUGAAUAGUGA-3′
and antisense, 5′-UCACUAUUCAGCUUCUAGG-3′. The negative sequence for
Prx II was: Sense, 5′-UUCUCCGAACGUGUCACGU-3′ and antisense,
5′-ACGUGACACGUUCGGAGAA-3′. No added sequence was used as a blank
control (Blank). The absorbance at 450 nm was recorded with an
ELX-800 Absorbance reader (BioTek Instruments, Inc., Winooski, VT,
USA). Each experiment was repeated not ≥3 times under similar
conditions.
Cell migration and invasion assay
Transwell chambers (EMD Millipore, Billerica, MA,
USA) with an 8-µm pore polycarbonate membrane filter was covered
with 60 µl/well Matrigel (BD Biosciences, San Jose, CA, USA) for
the invasion assay or without Matrigel for a migration assay and
inserted in a 24-well cultured plate. A total of 600 µl DMEM with
10% fetal bovine serum was placed in the bottom compartment of the
chamber, 200 µl of serum-free medium containing ~2×104
cells were added into the upper compartment of the chamber. The
chamber was incubated at 37°C in a humidified chamber in an
atmosphere of 5% CO2 for 12 h (migration assay) or 24 h
(invasion assay). The evaluation of migration and invasion was
performed at different times as invasion experiments require gel
plating, thus cells need to be cultured for longer to pass through
the membrane.
Following incubation, the Transwell chambers were
removed. The non-invasive cells on the upper side of the chamber
were collected with a cotton swab. Cells that migrated into the
pores of the inserted filter were fixed with 95% methanol at 37°C
for 30 min and stained with 0.5% crystal violet at 37°C for 30 min.
Images of the cells in the lower chamber were captured using a
fluorescence microscope (Ti-U; Nikon Corporation, Tokyo, Japan)
equipped with NIS-Elements F3.2. software (Nikon Corporation) at
magnification, ×200. The number of cells was recorded by randomly
selecting five visual fields in the Transwell chamber.
Statistical analysis
All data were analyzed by using SPSS software
version 16.0 (SPSS, Inc., Chicago, IL, USA) and GraphPad Prism 5.0
(GraphPad, San Diego, CA, USA). Quantitative data is presented as
the mean ± standard deviation. The Student's t-test was used to
analyze data between two groups, and one-way analysis of variance
with post hoc contrast Student-Newman-Keuls test was applied for
multigroup comparison. χ2 was applied for the numeration
data. Survival data was examined by the Kaplan-Meier method, and
the contrast among the groups were analyzed by utilizing the
log-rank test. Independent prognostic indicators in the
multivariate investigation were evaluated by utilizing the Cox's
proportional hazard model. P<0.05 is considered to indicate
significant differences in statistics.
Results
Upregulated Prx II expression in GC
cells
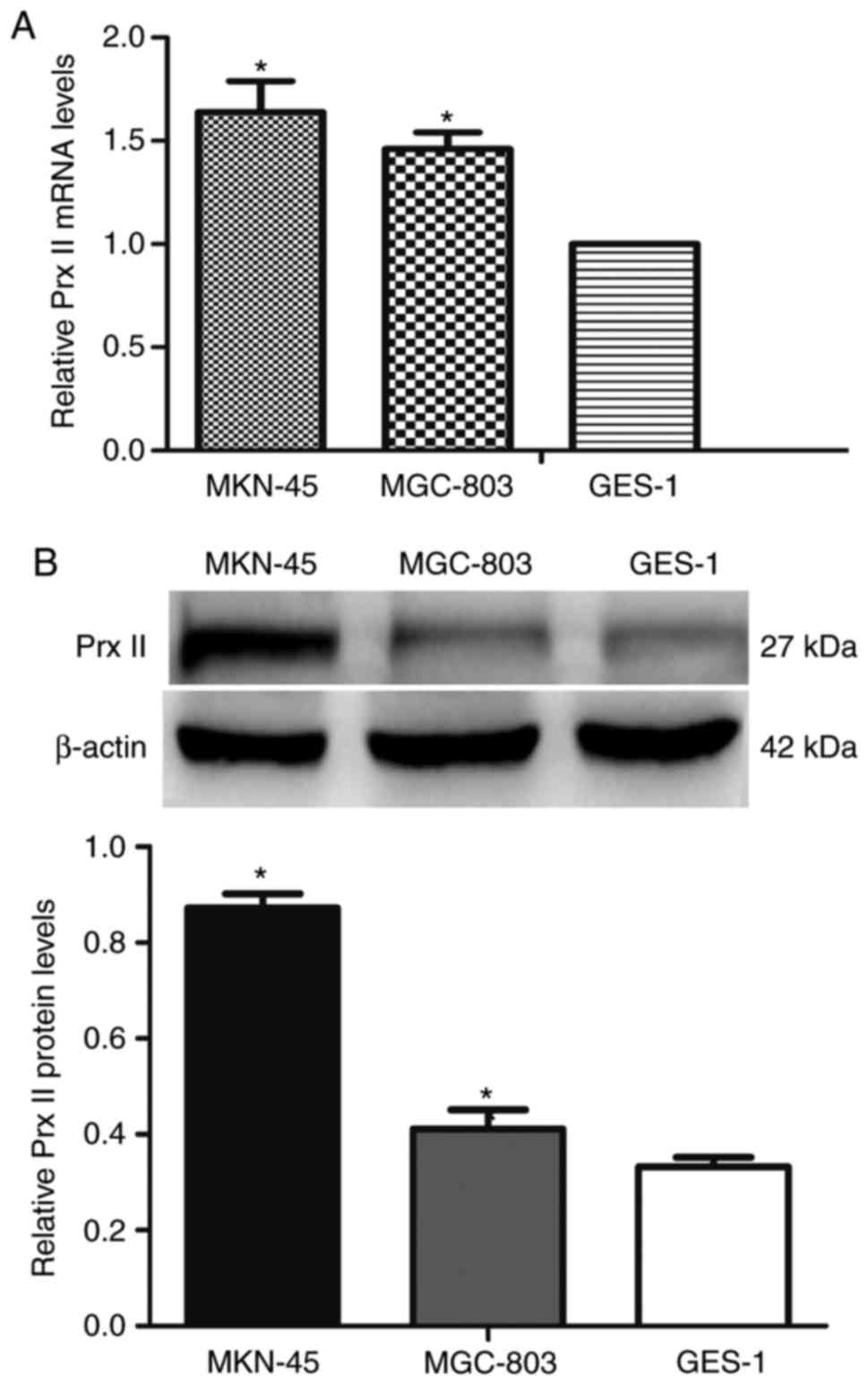
With the use of RT-qPCR, the mRNA expression level
of Prx II in GC cell lines (MGC-803 and MKN-45) were identified to
be significantly higher compared with that in normal cells of the
stomach (GES-1) (P<0.05; Fig. 1A).
For further confirmation, the protein expression of Prx II in cells
were examined (MGC-803, MKN-45, GES-1) by western blotting, which
demonstrated consistent results (P<0.05; Fig. 1B). Compared with GES-1 cells, the mRNA
and protein expression levels of Prx II exhibited no statistically
significant difference between different GC cell lines (MGC-803,
MKN-45; P>0.05).
Upregulated Prx II expression in GC
tissues
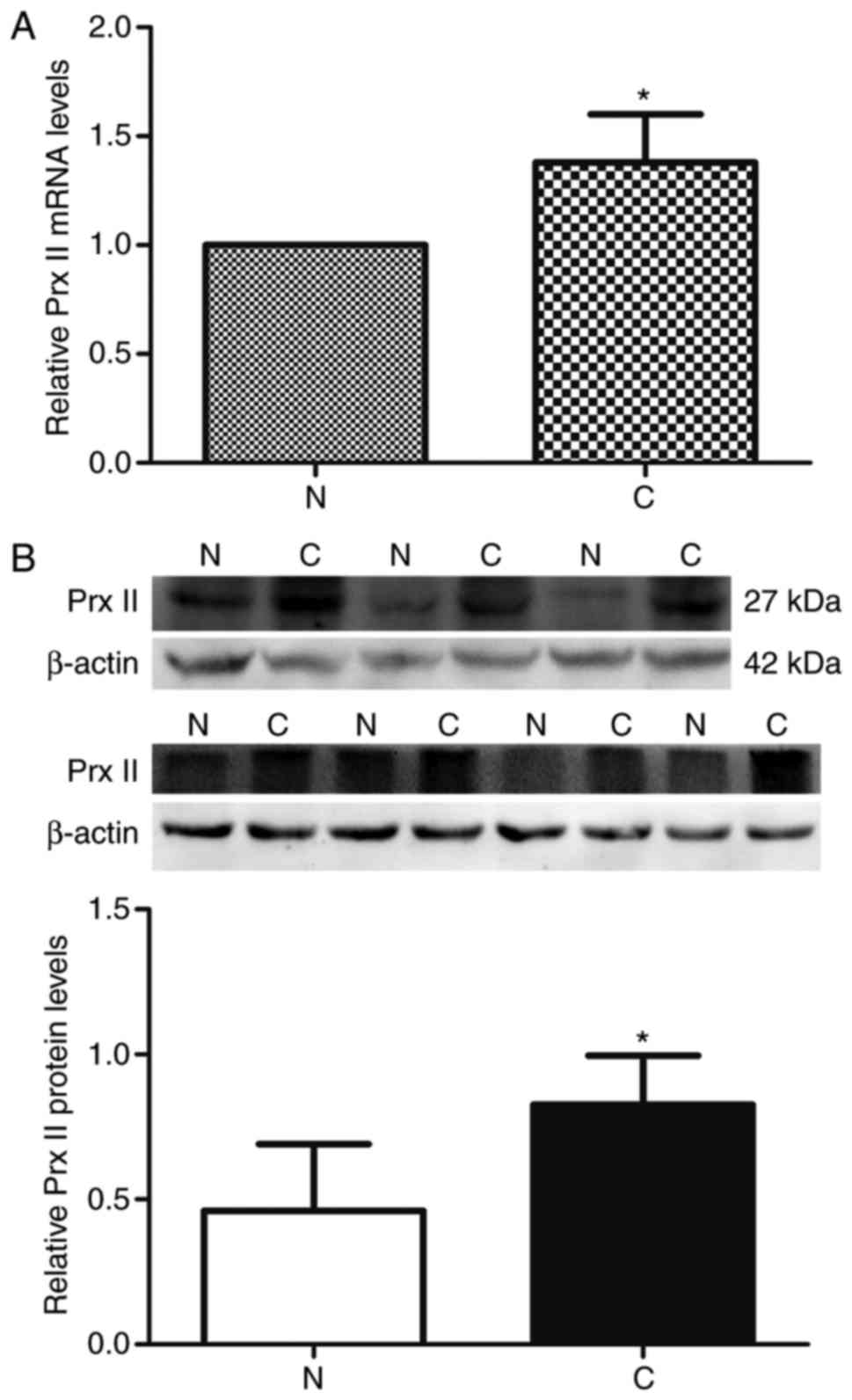
First, the mRNA expression level of Prx II in 45
paired GC and adjacent non-cancerous gastric tissues was analyzed
with RT-qPCR. The mean mRNA level of Prx II in the GC tissues
(1.38±0.22) was significantly higher compared with that in adjacent
tissues (1.00±0.00; P<0.01; Fig.
2A). Western blot analysis of clinical samples also verified a
significantly increased mRNA expression of Prx II in GC tissues
(P<0.01; Fig. 2B). In addition,
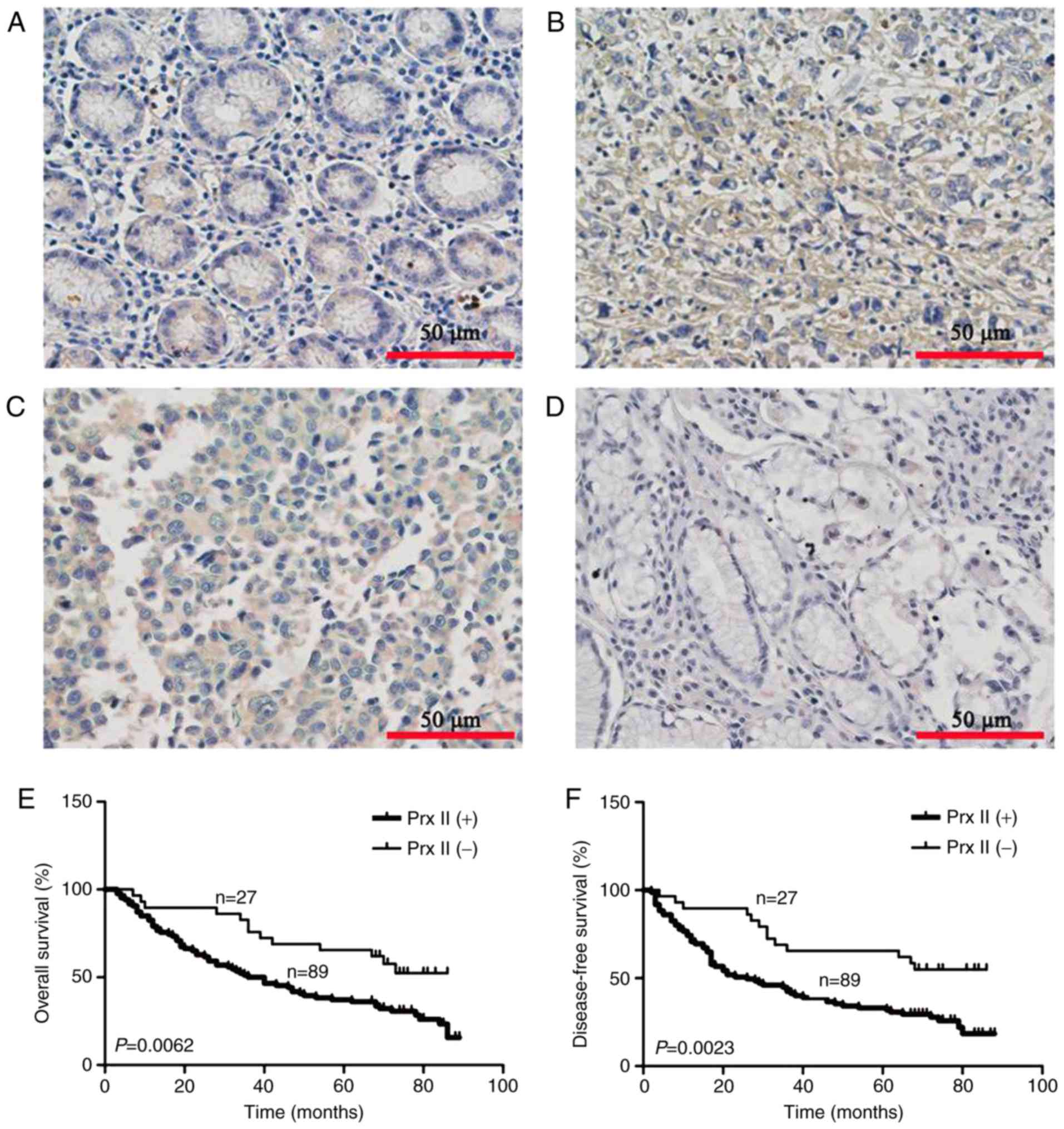
compared with poorly differentiated (Fig.
3B) and moderately differentiated GC tissues (Fig. 3C) the expression of Prx II in adjacent
tissues was significantly downregulated (Fig. 3A). There was a little difference
between the expression of Prx II in the highly differentiated GC
tissues (Fig. 3D) and adjacent
tissues. The positive expression rate of Prx II was 76.7% (89/116)
in the GC tissues and 30.1% (35/116) in the non-cancerous tissues
(χ2=50.516; P<0.01).
Levels of Prx II expression predicts
the survival of patients with GC
According to the results of immunohistochemistry,
116 GC patients were classified into two groups: High expression
(n=89) and low expression group (n=27). As presented in Table I, Prx II protein expression was
significantly associated with tumor size (P=0.029), histological
differentiation (P=0.024), depth of invasion (P<0.001), TNM
stage (P<0.001) and lymph node metastasis (P<0.001). No
significant associations were identified between the expression of
Prx II protein, and sex, age, location or distant metastasis
(P>0.05). Kaplan-Meier analysis demonstrated that patients with
low levels expression of Prx II had longer overall and disease-free
survival times compared with those with high Prx II levels
(P<0.01; Fig. 3E and F). Cox
multivariate analysis revealed that Prx II, depth of invasion,
lymph node metastasis and distant metastasis were independent
predictors of overall survival in patients with GC (P<0.05;
Table II). Similar results were also
obtained for disease-free survival in patients with GC (P<0.05;
Table III). Taken together, these
clinical findings indicated that Prx II expression was an important
indicator of survival for patients with GC among the analyzed
factors.
 | Table I.Association between the protein
expression of Prx II and clinicopathological parameters in GC. |
Table I.
Association between the protein
expression of Prx II and clinicopathological parameters in GC.
|
|
| Prx II (%) |
|
|---|
|
|
|
|
|
|---|
| Variable | No. of
patients | High | Low | P-value |
|---|
| Sex |
|
|
|
|
|
Male | 78 | 60 (76.9) | 18 (23.1) | 0.942 |
|
Female | 38 | 29 (76.3) | 9 (23.7) |
|
| Age, years |
|
|
|
|
|
<59 | 57 | 42 (73.7) | 15 (26.3) | 0.446 |
|
≥59 | 59 | 47 (79.7) | 12 (20.3) |
|
| Tumor size, cm |
|
|
|
|
|
<5 | 56 | 38 (67.9) | 18 (32.1) | 0.029 |
| ≥5 | 60 | 51 (85) | 9 (15) |
|
| Location |
|
|
|
|
| Other
location | 45 | 31 (68.9) | 14 (31.1) | 0.115 |
| Gastric
antrum | 71 | 58 (81.7) | 13 (18.3) |
|
| Depth of
invasion |
|
|
|
|
|
T1+T2 | 21 | 4 (19.0) | 17 (81.0) | <0.001 |
|
T3+T4 | 95 | 85 (90.4) | 10 (9.6) |
|
|
Differentiation |
|
|
|
|
|
Poor | 69 | 58 (84.1) | 11 (15.9) | 0.024 |
|
Moderate/high | 47 | 31 (66.0) | 16 (34.0) |
|
| TNM stage |
|
|
|
|
|
I+II | 39 | 17 (43.6) | 22 (56.4) | <0.001 |
|
III | 77 | 72 (93.5) | 5 (6.5) |
|
| LNM |
|
|
|
|
| No | 33 | 14 (43.6) | 19 (56.4) | <0.001 |
|
Yes | 83 | 75 (93.5) | 8 (6.5) |
|
| Distant
metastasis |
|
|
|
|
| No | 101 | 76 (75.2) | 25 (24.8) | 0.263 |
|
Yes | 15 | 13 (86.7) | 2 (13.3) |
|
| Table II.Univariable and multivariable
analysis of predictors of overall survival in patients with GC. |
Table II.
Univariable and multivariable
analysis of predictors of overall survival in patients with GC.
|
| Univariable
analysis |
| Multivariable
analysis |
|
|---|
|
|
|
|
|
|
|---|
| Variable | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Prx II
expression | 2.643
(1.395–5.007) | 0.003 | 2.034
(1.395–5.007) | 0.037 |
| Sex | 0.952
(0.593–1.530) | 0.839 | – | NA |
| Age | 0.796
(0.513–1.263) | 0.309 | – | NA |
| Tumor size | 1.571
(1.009–2.445) | 0.045 | 1.101
(0.687–1.765) | 0.690 |
| Location | 1.429
(0.914–2.232) | 0.117 | – | NA |
| Depth of
invasion | 3.338
(1.533–7.270) | 0.002 | 3.983
(1.039–14.919) | 0.044 |
|
Differentiation | 0.652
(0.409–1.038) | 0.031 | 0.894
(0.554–1.443) | 0.119 |
| TNM stage | 3.496
(2.029–6.025) | <0.001 | 1.305
(0.055–2.305) | 0.380 |
| LNM | 0.196
(0.103–0.373) | <0.001 | 0.106
(0.032–0.351) | <0.001 |
| Distant
metastasis | 0.353
(0.189–0.659) | 0.001 | 0.604
(0.288–1.053) | 0.042 |
| Table III.Univariable and multivariable
analysis of predictors of disease-free survival in patients with
GC. |
Table III.
Univariable and multivariable
analysis of predictors of disease-free survival in patients with
GC.
|
| Univariable
analysis |
| Multivariable
analysis |
|
|---|
|
|
|
|
|
|
|---|
| Variable | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Prx II
expression | 2.590
(1.908–4.776) | 0.006 | 2.168
(0.645–4.028) | 0.032 |
| Sex | 0.963
(0.600–1.547) | 0.454 | – | NA |
| Age | 0.775
(0.414–1.181) | 0.291 | – | NA |
| Tumor size | 1.575
(1.012–2.452) | 0.044 | 1.111
(0.692–1.783) | 0.664 |
| Location | 1.450
(0.928–2.265) | 0.103 | – | NA |
| Depth of
invasion | 3.793
(1.742–8.260) | <0.001 | 4.127
(1.073–15.867) | 0.039 |
|
Differentiation | 0.669
(0.171–2.244) | 0.040 | 0.839
(0.288–2.125) | 0.143 |
| TNM stage | 3.546
(2.05–6.118) | <0.001 | 1.286
(0.033–2.103) | 0.407 |
| LNM | 0.190
(0.100–0.363) | <0.001 | 0.095
(0.028–0.321) | <0.001 |
| Distant
metastasis | 0.377
(0.202–0.705) | 0.002 | 0.593
(0.309–1.137) | 0.039 |
Prx II knockdown suppresses GC cell
proliferation
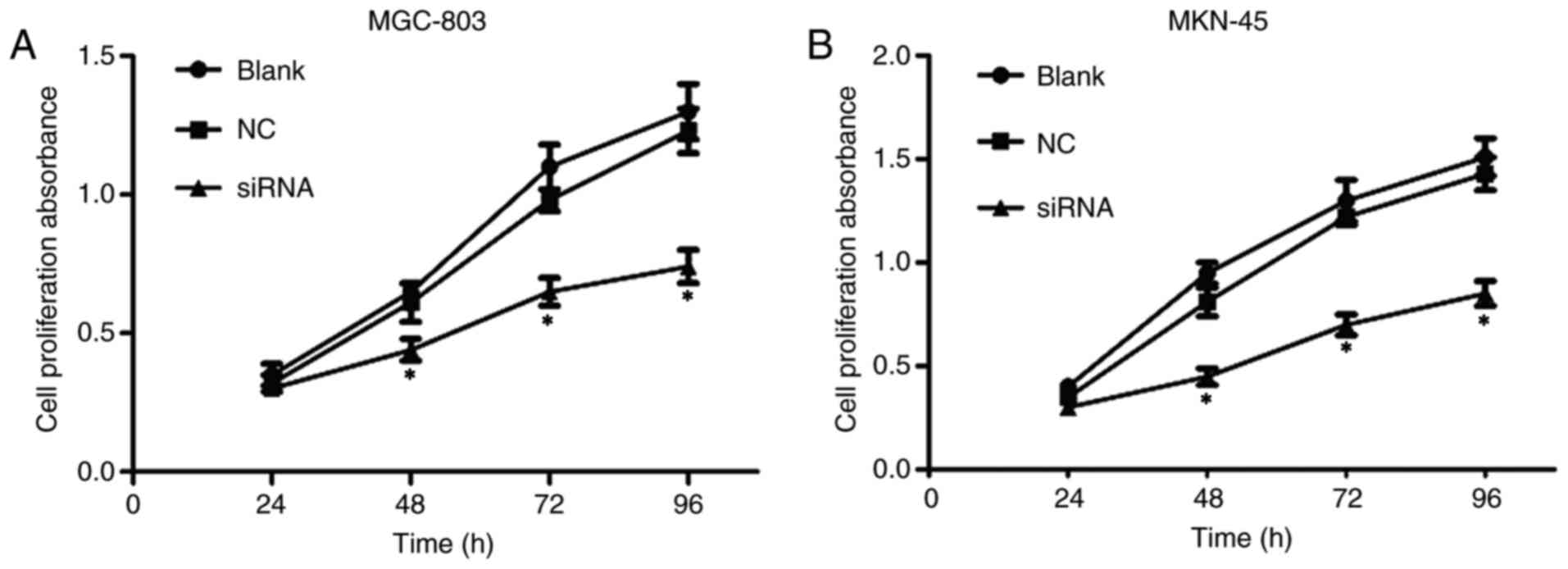
In order to investigate whether Prx II knockout
affected the proliferation of GC cells, a CCK-8 assay was used
performed. As presented in Fig. 4,
the proliferation rate in positive control group (siRNA) was
significantly suppressed at different times (48, 72 and 96 h),
particularly at 96 h (P<0.05). However, no significant
differences were observed between the normal cultured group (blank)
and the negative control group (NC) (P>0.05).
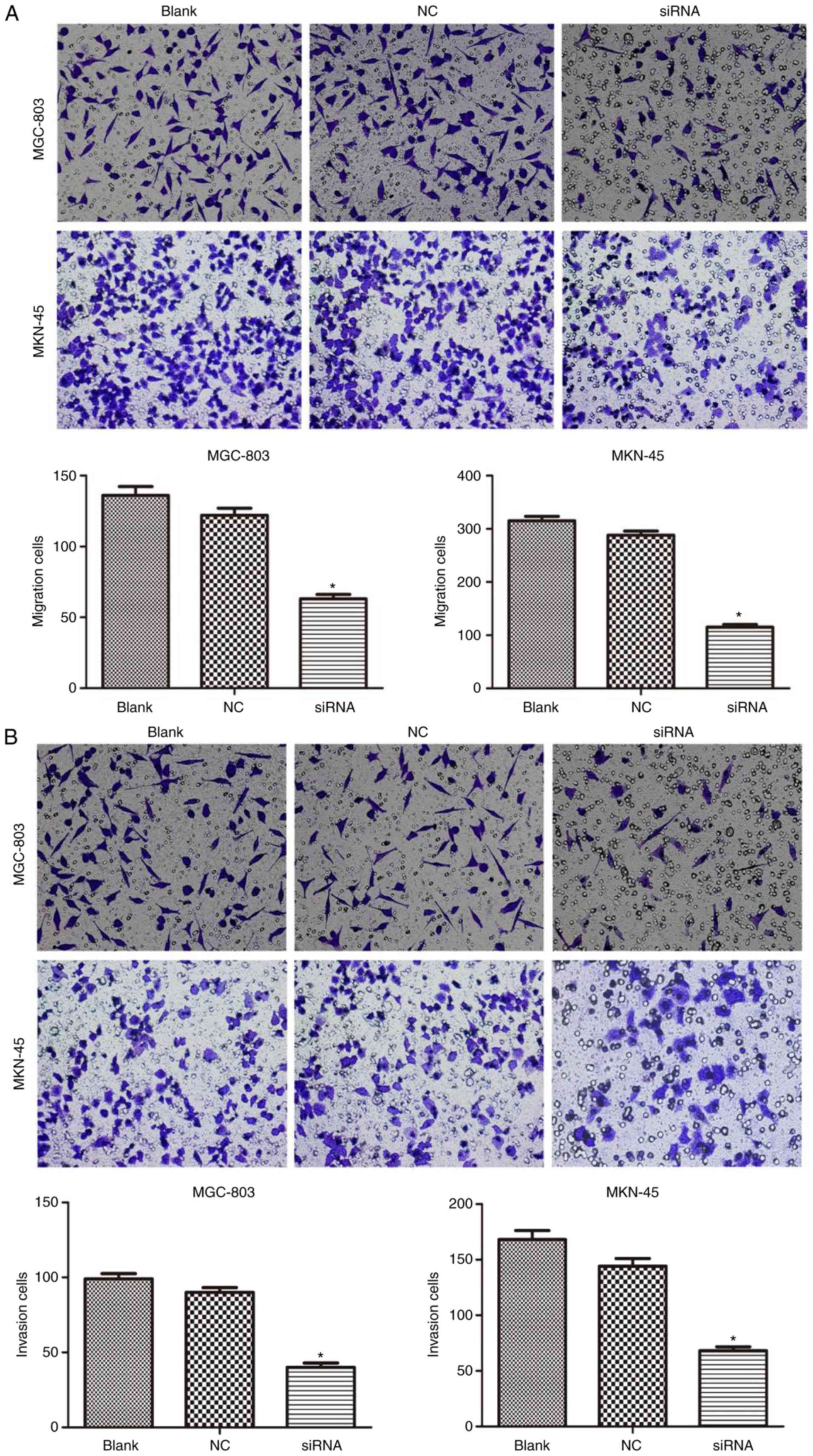
Prx II knockdown suppresses the
expression of E-cadherin, N-cadherin, MMP-2 and MMP-9, and GC cell
migration
As presented in Fig.
5, the western blot analysis revealed that silencing Prx II
resulted in significantly decreased protein expression levels of
MMP-2, MMP-9 and N-cadherin, and increased expression of E-cadherin
in MGC-803 and MKN-45 cells compared with the NC group (P<0.05).
The cell relocation measure showed that the transfection lessened
the cell movement capacity by 53.7% for MGC-803 and by 63.5% for
MKN-45 cells compared with the control group (Fig. 6A; P<0.05). Additionally, an
invasion assay was used quantify the invasive capacity of the cells
through channels covered with a reconstituted cellar membrane. It
was demonstrated that the transfection significantly reduced the
invasive capacity by 59.6% in MGC-803 and by 54.8% in MKN-45 cells
compared with the control group (Fig.
6B; P<0.05).
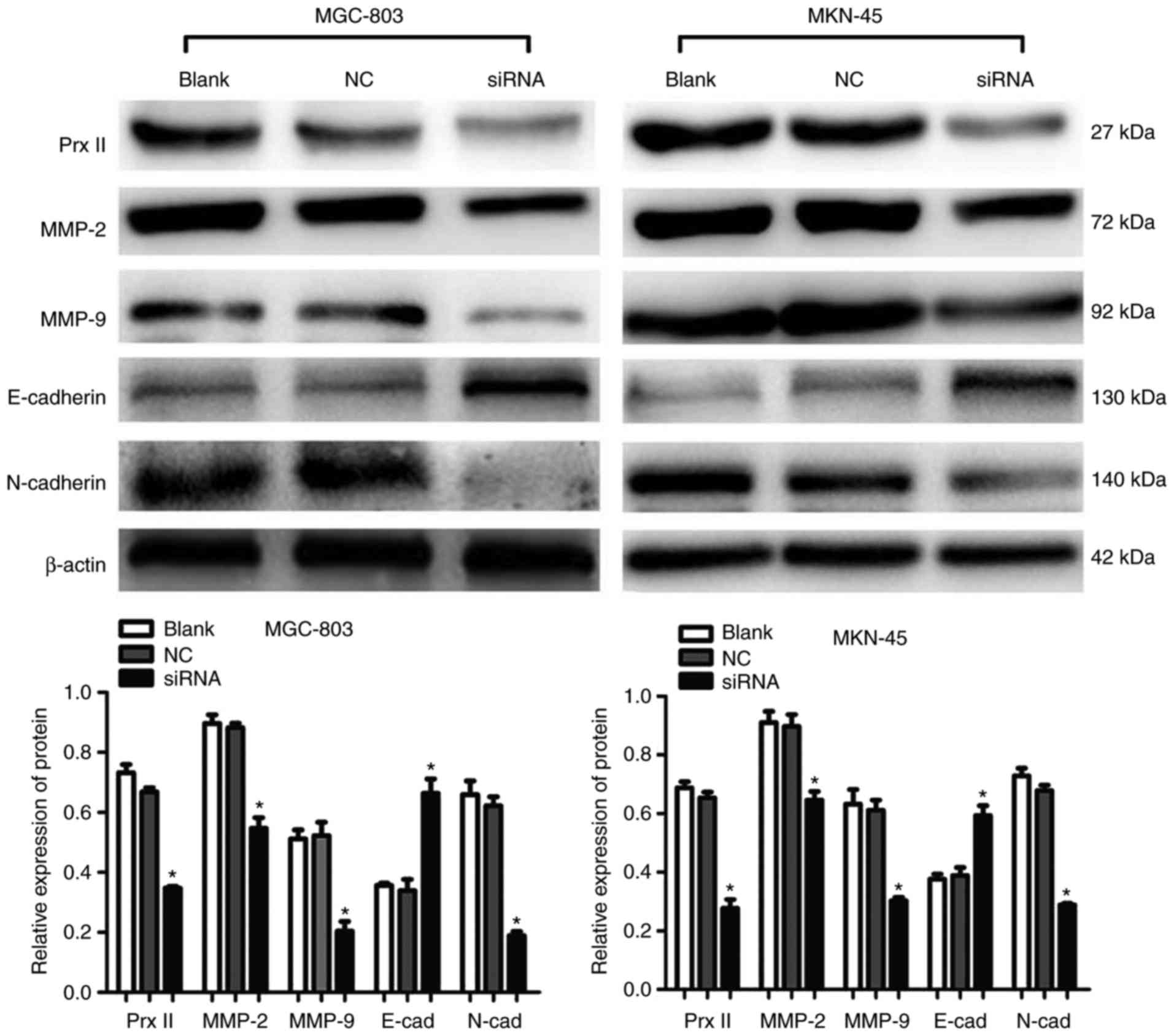 | Figure 5.Interaction between Prx II, and
E-cadherin, N-cadherin, MMP-2 and MMP-9 in gastric cancer MGC-803
and MKN-45 cells. The protein expression levels of Prx II,
E-cadherin, N-cadherin, MMP-2 and MMP-9 were determined by western
blot analysis. β-actin was used as a loading control. *P<0.05
vs. Blank and NC. Prx, peroxiredoxin; NC, negative control; siRNA,
small interfering RNA; MMP, matrix metalloproteinase; E-cad,
E-cadherin; N-cad, N-cadherin. |
Discussion
According to the cysteine residues present at the
carboxyl and N-terminus, Prxs may be divided into three categories:
1-Cys Prx, typical 2-Cys Prx and atypical 2-Cys Prx. Prx II belongs
to the typical 2-Cys Prx class (22,23). Prx
II is involved in regulating cell proliferation, differentiation,
apoptosis and signaling pathways, and they serve antioxidative
defense roles by reducing and detoxifying various reactive oxygen
species (ROS) that incorporate hydrogen peroxide
(H2O2). It has been demonstrated that high
levels of Prx II reduce H2O2-induced
apoptosis in thyroid cells (24).
Kang et al (25) reported that
the high expression of PrxII may participate in cell
differentiation and apoptosis through inhibiting the production of
NF-κB. In addition, knockdown of Prx II prompts advance
platelet-derived growth factor-induced cell division and metastasis
(26). Recent studies have shown that
PrxII participates in the activation of extracellular
signal-regulated kinase (ERK), but that it does not participate in
the activation of Janus kinase and p38 mitogen-activated protein
kinase (MAPK). In fact, overexpression of PrxII in activated T
cells increases the activation of the MAPK kinase (MEK)/ERK pathway
induced by CD3 (27). In vascular
endothelial cells, downregulation of PrxII can inhibit the
activation of ERK (28).
According to previous studies, Prx II expression is
increased in lung (15), renal
(16), hepatocellular (29) and bosom carcinoma (30). GC is one of the most devastating types
of tumor worldwide. In the present study, Prx II was upregulated in
GC cell lines and GC tissues from patients, and high expression of
Prx II was significantly associated with tumor size (P=0.029),
histological differentiation (P=0.024), depth of invasion
(P<0.001), TNM stage (P<0.001) and lymph node metastasis
(P<0.001). Notably, Kaplan-Meier analysis revealed that patients
with high Prx II expression had a poorer overall survival time
(P=0.0062) and disease-free survival time (P=0.0023) compared with
those with low Prx II. Cox multivariate analysis revealed that Prx
II, depth of invasion, lymph node metastasis and distant metastasis
were independent predictors of overall survival in patients with
GC. Thus, thus this indicates that Prx II may aid in improving
survival prediction of patients with GC.
It has been indicated that cells in a mild oxidation
state do not cause the upregulation or downregulation of Prx II,
while cells in a severe oxidation state promote the upregulation of
Prx II (31). Thus, the antioxidation
system serves an important role in regulating peroxide levels and
changes to redox states (31).
Therefore, high expression levels of Prx II may be associated with
a state of persistent oxidation in the tumor tissue
microenvironment. A chronic hypoxic state surrounds tumor tissues,
and may induce apoptosis of carcinoma cells (32). Thus, the role of the antioxidation
system of Prx II in the development of tumor cells is important.
ROS includes organic hydroperoxide and H2O2.
Increasing studies have demonstrated that ROS is associated with
the development of cancer in the human body (33). Wang et al (34) reported that high Prx II levels are
associated with the development of resistance of breast malignant
cells to certain radiotherapies, and it may result in to reduced
ROS levels. Thus, high Prx II expression levels may be associated
with ROS.
Cell proliferation and migration are associated with
various physiological processes. However, uncontrolled cell
proliferation and migration may induce and progress tumor
development (35). The present study
used Transwell migration and invasion assays, and western blot
analysis to detect the changes in invasion- and
migration-associated genes. In the present study, it was
demonstrated that Prx II knockdown significantly reduced the
proliferation and migration rates of GC cells.
EMT-associated proteins, including E-cadherin and
N-cadherin, serve an important role in GC. EMT is associated with
the metastasis of cancer, whereby epithelial cells become
mesenchymal cells; however, EMT also typically occurs in tissue
morphogenesis amid wound repair and tissue growth in healthy human
tissues (36,37). Altered expression levels of MMPs have
been associated with the metastasis of numerous types of cancer.
Degradation of the stromal cell layer is involved in the metastasis
of cancer (38). MMP-2, along with
MMP-9, is fit for debasing type IV collagen, which is the bottom of
the stromal cell layer. Thus, tumor metastasis capacity is
associated with E-cadherin, N-cadherin, MMP-2 and MMP-9. By
utilizing western blot analysis, it was revealed that
downregulating expression, the levels of MMP-2, MMP-9 and
N-cadherin protein were reduced, and that of E-cadherin was
upregulated in GC cells. However, a limitation of the present study
was that the activity levels of MMP-2 and −9 were not
evaluated.
There have been various studies investigating the
role of Prx II in the proliferation and metastasis of tumor. Lu
et al (39) reported that
downregulation of Prx II represses cell development in colorectal
cancer by influencing the Wnt/β-catenin signaling pathway. Park
et al (29) demonstrated that
Prx II increases hepatic tumorigenesis through collaboration with
the Ras/Forkhead box M1 signaling pathway. Furthermore, certain
studies have demonstrated that Prx II attenuates poly(ADP-ribose)
polymerase 1-induced and p53-induced cell apoptosis Prx II
transgenic mice (40). Low
concentrations of ROS advance cell proliferation, whereas high
concentrations of ROS advance cell apoptosis (41). Therefore, we hypothesized that Prx II
may improve the limitation of tumor metastasis by regulating the
level of ROS. As the present study was a preliminary study on the
role of Prx II in the development and progression of GC, no further
investigations into the role of ROS were performed. Therefore, it
is a limitation of this study.
In conclusion, the current results reveal that Prx
II is a necessary functional part of GC cell growth, but its
molecular mechanism remains unclear. In addition, further in
vitro and clinical studies on the effects of the Prx II gene in
tumorigenesis are required. The results of the present study
provide primary information and support the development of a novel
treatment strategy in the management of GC.
Acknowledgements
The present study was supported by Jiangsu Key
Laboratory of Biological Cancer Therapy. The authors would like to
thank their colleagues at the Jiangsu Key Laboratory of Biological
Cancer Therapy, Xuzhou Medical University for their technical
assistance.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YG, LN, AL and WX were responsible for the design of
the study. LN, AL, LY and WZ performed the experiments and
collected the analytical data. LN and AL wrote the manuscript.
Ethics approval and consent to
participate
Each patient was approved by the Ethics Review Board
of the Affiliated Hospital of Xuzhou Medical College. All
procedures performed in studies involving human participants were
in accordance with the Declaration of Helsinki. Informed consent
was obtained from all individual participants included in the
study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Calcagno DQ, de Arruda Cardoso Smith M and
Burbano RR: Cancer type-specific epigenetic changes: Gastric
cancer. Methods Mol Biol. 1238:79–101. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cheah IK, Lei F, Tang RMY, Lim KHC and
Halliwell B: Ergothioneine levels in an elderly population decrease
with age and incidence of cognitive decline; a risk factor for
neurodegeneration? Biochem Biophys Res Commun. 478:162–167. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Subramanya MS, Hossain MB, Khan S, Memon B
and Memon MA: Meta-analysis of D1 versus D2 gastrectomy for gastric
adenocarcinoma. Islamic Countries Conferecne on Statistical
Sciences. 2009.
|
|
5
|
Conteduca V, Sansonno D, Lauletta G, Russi
S, Ingravallo G and Dammacco F: H. pylori infection and gastric
cancer: State of the art (review). Int J Oncol. 42:5–18. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Venerito M, Vasapolli R, Rokkas T,
Delchier JC and Malfertheiner P: Helicobacter pylori,
gastric cancer and other gastrointestinal malignancies.
Helicobacter. 22 Suppl 1:2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sarker KK, Kabir MJ, Bhuyian AKMMU, Alam
MS, Chowdhury FR, Ahad MA, Rahman MA and Rahman MM: H. pylori
infection and gastric cancer in Bangladesh: A case-control study.
Int J Surg Oncol (N Y). 2:e442017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Grabsch HI and Tan P: Gastric cancer
pathology and underlying molecular mechanisms. Dig Surg.
30:150–158. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Canu V, Sacconi A, Lorenzon L, Biagioni F,
Lo Sardo F, Diodoro MG, Muti P, Garofalo A, Strano S, D'Errico A,
et al: MiR-204 down-regulation elicited perturbation of a gene
target signature common to human cholangiocarcinoma and gastric
cancer. Oncotarget. 8:29540–29557. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Abbas M, Habib M, Naveed M, Karthik K,
Dhama K, Shi M and Dingding C: The relevance of gastric cancer
biomarkers in prognosis and pre- and post-chemotherapy in clinical
practice. Biomed Pharmacother. 95:1082–1090. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gong F, Hou G, Liu H and Zhang M:
Peroxiredoxin 1 promotes tumorigenesis through regulating the
activity of mTOR/p70S6K pathway in esophageal squamous cell
carcinoma. Med Oncol. 32:4552015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Choi H, Chang JW and Jung YK:
Peroxiredoxin 6 interferes with TRAIL-induced death-inducing
signaling complex formation by binding to death effector domain
caspase. Cell Death Differ. 18:405–414. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Perkins A, Poole LB and Karplus PA: Tuning
of peroxiredoxin catalysis for various physiological roles.
Biochemistry. 53:7693–7705. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nyström T, Yang J and Molin M:
Peroxiredoxins, gerontogenes linking aging to genome instability
and cancer. Genes Dev. 26:2001–2008. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lehtonen ST, Svensk AM, Soini Y, Pääkkö P,
Hirvikoski P, Sang WK, Säily M and Kinnula VL: Peroxiredoxins, a
novel protein family in lung cancer. Int J Cancer. 111:514–521.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Soini Y, Kallio JP, Hirvikoski P, Helin H,
Kellokumpu-Lehtinen P, Kang SW, Tammela TL, Peltoniemi M,
Martikainen PM and Kinnula VL: Oxidative/nitrosative stress and
peroxiredoxin 2 are associated with grade and prognosis of human
renal carcinoma. APMIS. 114:329–337. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu Y and Zhou BP: Epithelial-Mesenchymal
Transition in Development and Diseases. Springer; New York:
2009
|
|
18
|
Petrova YI, Schecterson L and Gumbiner BM:
Roles for E-cadherin cell surface regulation in cancer. Mol Biol
Cell. 27:3233–3244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xian X, Huang L, Zhang B, Wu C, Cui J and
Wang Z: WIN 55,212-2 inhibits the epithelial mesenchymal transition
of gastric cancer cells via COX-2 signals. Cell Physiol Biochem.
39:2149–2157. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lv Z, Weng X, Du C, Zhang C, Xiao H, Cai
X, Ye S, Cheng J, Ding C, Xie H, et al: Downregulation of HDAC6
promotes angiogenesis in hepatocellular carcinoma cells and
predicts poor prognosis in liver transplantation patients. Mol
Carcinog. 55:1024–1033. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ow SH, Chua PJ and Bay BH: Epigenetic
regulation of peroxiredoxins: Implications in the pathogenesis of
cancer. Exp Biol Med (Maywood). 242:140–147. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rhee SG, Chae HZ and Kim K:
Peroxiredoxins: A historical overview and speculative preview of
novel mechanisms and emerging concepts in cell signaling. Free
Radic Biol Med. 38:1543–1552. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim H, Lee TH, Park ES, Suh JM, Park SJ,
Chung HK, Kwon OY, Kim YK, Ro HK and Shong M: Role of
peroxiredoxins in regulating intracellular hydrogen peroxide and
hydrogen peroxide-induced apoptosis in thyroid cells. J Biol Chem.
275:18266–18270. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kang SW, Chae HZ, Seo MS, Kim K, Baines IC
and Rhee SG: Mammalian peroxiredoxin isoforms can reduce hydrogen
peroxide generated in response to growth factors and tumor necrosis
factor-alpha. J Biol Chem. 273:6297–6302. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Choi MH, Lee IK, Kim GW, Kim BU, Han YH,
Yu DY, Park HS, Kim KY, Lee JS, Choi C, et al: Regulation of PDGF
signalling and vascular remodelling by peroxiredoxin II. Nature.
435:347–353. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kwon J, Devadas S and Williams MS: T cell
receptor-stimulated generation of hydrogen peroxide inhibits
MEK-ERK activation and lck serine phosphorylation. Free Radic Biol
Med. 35:406–417. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kang DH, Lee DJ, Lee KW, Park YS, Lee JY,
Lee SH, Koh YJ, Koh GY, Choi C, Yu DY, et al: Peroxiredoxin II Is
an essential antioxidant enzyme that prevents the oxidative
inactivation of VEGF receptor-2 in vascular endothelial cells. Mol
Cell. 44:545–558. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Park YH, Kim SU, Kwon TH, Kim JM, Song IS,
Shin HJ, Lee BK, Bang DH, Lee SJ, Lee DS, et al: Peroxiredoxin II
promotes hepatic tumorigenesis through cooperation with Ras
Forkhead box M1 signaling pathway. Oncogene. 35:3503–3513. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Karihtala P, Mäntyniemi A, Kang SW,
Kinnula VL and Soini Y: Peroxiredoxins in breast carcinoma. Clin
Cancer Res. 9:3418–3424. 2003.PubMed/NCBI
|
|
31
|
Lehtonen ST, Markkanen PM, Peltoniemi M,
Kang SW and Kinnula VL: Variable overoxidation of peroxiredoxins in
human lung cells in severe oxidative stress. Am J Physiol Lung Cell
Mol Physiol. 288:L997–L1001. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang WJ and Zou XY: Influence of
cilazapril and taurine on apoptosis of chronic hypoxic lung tissue.
Chin J Patho. 1999.
|
|
33
|
Marengo B, Nitti M, Furfaro AL, Colla R,
Ciucis CD, Marinari UM, Pronzato MA, Traverso N and Domenicotti C:
Redox homeostasis and cellular antioxidant systems: crucial players
in cancer growth and therapy. Oxid Med Cell Longev.
2016:62356412016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang T, Diaz AJ and Yen Y: The role of
peroxiredoxin II in chemoresistance of breast cancer cells. Breast
Cancer (Dove Med Press). 6:73–80. 2014.PubMed/NCBI
|
|
35
|
Ward E: Cancer statistics, 2010. Ca A
Cancer J Clin. 62:10–29. 2012.
|
|
36
|
Kim H, Yoo SB, Sun P, Jin Y, Jheon S, Lee
CT and Chung JH: Alteration of the E-Cadherin/β-Catenin Complex Is
an Independent Poor Prognostic Factor in Lung Adenocarcinoma.
Korean J Pathol. 47:44–51. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Brabletz T, Hlubek F, Spaderna S,
Schmalhofer O, Hiendlmeyer E, Jung A and Kirchner T: Invasion and
metastasis in colorectal cancer: epithelial-mesenchymal transition,
mesenchymal-epithelial transition, stem cells and beta-catenin.
Cells Tissues Organs. 179:56–65. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mook OR, Frederiks WM and Van Noorden CJ:
The role of gelatinases in colorectal cancer progression and
metastasis. Biochim Biophys Acta. 1705:69–89. 2004.PubMed/NCBI
|
|
39
|
Lu W, Fu Z, Wang H, Feng J, Wei J and Guo
J: Peroxiredoxin 2 knockdown by RNA interference inhibits the
growth of colorectal cancer cells by downregulating Wnt/β-catenin
signaling. Cancer Lett. 343:190–199. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Leak RK, Zhang L, Luo Y, Li P, Zhao H, Liu
X, Ling F, Jia J, Chen J and Ji X: Peroxiredoxin 2 battles
poly(ADP-ribose) polymerase 1- and p53-dependent prodeath pathways
after ischemic injury. Stroke. 44:1124–1134. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu SL, Shi DY, Pan XH and Shen ZH:
Inhibition of proliferation and expression of N-ras in hepatoma
cells by antioxidation treatment. Sheng Wu Hua Xue Yu Sheng Wu Wu
Li Xue Bao (Shanghai). 33:463–466. 2001.PubMed/NCBI
|