Introduction
Bladder cancer is the most common malignancy of the
urinary tract (1). There were an
estimated 16,000 mortalities due to bladder cancer in 2015 in the
United States alone (2). A notable
risk factor for the development of bladder cancer is the
occupational exposure to aromatic amines (e.g., benzidine,
2-naphthylamine and 4-aminobiphenyl). Benzidine is commonly
encountered in the industrial dye and rubber industries, as well as
in hair dyes, paints, fungicides, motor vehicle exhaust fumes and
industrial pollutant emissions (3).
Thus, benzidine and other aromatic compounds are notable hazards
for human health. Although there have been a number of
epidemiological investigations about the association of benzidine
exposure with bladder cancer and other carcinomas (4), including our previous study, in which it
was demonstrated that benzidine could induce the
epithelial-mesenchymal transition in human uroepithelial cells
through the extracellular regulated protein kinases 1 and 2
(ERK1/2) pathway (5), limited
research has been conducted to investigate whether benzidine
exposure promotes cell proliferation and the underlying mechanisms
of this process.
The cell cycle is the process through which living
cells replicate genetic information and generate progeny cells. The
process can be divided into two highly regulated phases: The
interphase and mitotic phase. The former involves three phases, the
G1, S and G2 phases. Complex mechanisms are associated with the
modulation of the cell cycle, which is strictly regulated in normal
cells; modifications to cell cycle regulation may lead to disease,
including tumor formation (6). Cell
cycle dysregulation is necessary for cancer initiation and
progression (7).
The mitogen-activated protein kinases (MAPKs), a
family of enzymes that catalyze the phosphorylation of specific
serine and threonine residues on target substrates in order to
convert extracellular signals to intracellular, include four major
subfamilies: ERK1/2, the Jun N-terminal kinases (JNKs), p38 and
ERK5 (8,9). MAPKs serve important functions in many
life processes, including proliferation, differentiation and
apoptosis (10). The activation of
the ERK1/2, p38, and JNK/MAPK pathways is associated with the
induction of many transcription factors, resulting in the altered
expression of various genes associated with tumor cell
proliferation, apoptosis, angiogenesis, metastasis and the
progression of various types of cancer, including bladder cancer
(11,12).
In the present study, the mechanism of
benzidine-induced SV-HUC-1 cell proliferation was investigated,
including the role of MAPKs in the proliferation process. The aim
was to elucidate a potential mechanism for benzidine-induced
carcinogenesis.
Materials and methods
Chemicals and reagents
Benzidine (4,4′-diaminobiphenyl; ≥98.0%), dimethyl
sulfoxide (DMSO), MTT, methanol, glutaraldehyde and all other
chemicals and dyes of the highest purity were purchased from Merck
KGaA (Darmstadt, Germany). The kinase inhibitors U0126 (against
ERK1/2), SB203580 (against p38) and SP600125 (against JNK) were
obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Ham's F12 medium, fetal bovine serum (FBS), PBS, antibiotics and
trypsin were obtained from HyClone (GE Healthcare, Chicago, IL,
USA). Antibodies to ERK1/2 (cat. no. BS-2637R; 1:1,000), p38 (cat.
no. 12; 1:1,000), JNK (cat. no. AR2045; 1:1,000), phosphorylated
(p)-ERK1/2 (cat. no. RS-2637S; 1:1,000), p-JNK (cat. no. AF3318;
1:1,000), p-p38 (cat. no. 9212; 1:1,000), p-c-Jun (cat. no.
AF-3095; 1:1,000), p-c-Fos (cat. no. 5348; 1:1,000), cyclin D1
(cat. no. 2978; 1:1,000), proliferating cell nuclear antigen (PCNA;
cat. no. 10205-2-AP; 1:1,000) and P21 (cat. no. 10355-1-AP;
1:1,000) were purchased from Cell Signaling Technology, Inc. The
GAPDH (cat. no. 60004-1-Ig; 1:1,000) antibody was obtained from
Biogot Technology Co., Ltd. (Nanjing, China). Monoclonal rabbit Jun
D antibody (cat. no. sc-74; 1:1,000), Jun B antibody (cat. no.
10486-1-AP; 1:1,000), Fra-1 antibody (cat. no. D80B4; 1:1,000) and
FosB antibody (cat. no. AF5010; 1:1,000) was purchased from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA). Mouse anti-rabbit IgG
secondary antibodies (cat. no. bs-0295M; 1:10,0000) were purchased
from Beyotime Institute of Biotechnology (Nanjing, China).
Cell culture and treatments
SV-40 immortalized human uroepithelial cells
(SV-HUC-1) were obtained from the American Type Culture Collection
(Manassas, VA, USA) and grown in 25-cm2 flasks (initial
density, 1×105 cells/ml). Cells were maintained in Ham's
F-12 medium supplemented with 10% FBS, 100 units/ml penicillin and
100 units/ml streptomycin at 37°C with 5% CO2 in a
humidified incubator. Following culture for 12 h, cells were
exposed to different concentrations of benzidine (including 0,
0.0001, 0.001, 0.005, 0.01, 0.05, 0.1, 1, 10, 50, 75, 100, 125, 150
and 200 µM) diluted with DMSO, and/or treated with U0126 (10 µM),
SB203580 (10 µM) or SP600125 (5 µM), and were passaged daily for 6
days. All experiments were performed three times.
Cell proliferation assay
Cell viability was assayed by MTT conversion to
formazan. Subsequent to growing to 80% confluence on a 10
cm2 plate, 5×104 SV-HUC-1 cells per well were
seeded in 96-well plates. Then cells were treated with 100 µl
growth medium with 0.1% DMSO or benzidine (0.001-200 µM) for 2 or 6
days. MTT solution (10 µl of 5 mg/ml) was added to each well and
the plates incubated for an additional 4 h at 37°C. The medium was
removed and DMSO was added to each well to solubilize precipitants.
Absorbance was measured at 490 nm using a microplate reader. All
measurements were performed in triplicate.
Western blotting
For the western blot analysis, 6×106
SV-HUC-1 cells per dish were seeded in 100 mm plastic tissue
culture dishes. Following culture for 12 h as previously described,
cells were either exposed to different concentrations of benzidine
(0, 0.001, 0.005, 0.01, 0.05 or 0.1 µM) or treated with U0126,
SB203580 or SP600125 for 6 days. Cells were harvested, washed with
ice-cold PBS, and lysed in RIPA buffer (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). Concentrations of the precipitated
proteins in cell lysates were measured with BCA Protein Assay
(Pierce; Thermo Fisher Scientific, Inc.). Then, proteins (50 g per
lane) were separated by 10% SDS-PAGE and transferred onto a
polyvinylidene difluoride membrane (EMD Millipore, Billerica, MA,
USA). Subsequent to blocking in 5% fat-free dry milk in
Tris-buffered saline with Tween-20 (TBST), membranes were incubated
with primary antibodies (1:500, diluted with 5% milk) overnight at
4°C, washed in TBST and then incubated with goat anti-rabbit
peroxidase-conjugated secondary antibodies (1:500, diluted with 5%
milk) for 1 h at room temperature. The blots were subsequently
developed using an enhanced chemiluminescence detection kit
(Amersham; GE Healthcare) and exposed to film (Kodak, Rochester,
NY, USA). GAPDH served as the loading control. For densitometric
analyses, protein bands on the blots were measured using the Eagle
Eye II software (Agilent Technologies, Inc., Santa Clara, CA,
USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
For mRNA analysis, 6×106 SV-HUC-1 cells
per dish were seeded in 100-mm plastic tissue culture dishes.
Following culture for 12 h, cells were exposed to 0, 0.001, 0.005,
0.01, 0.05 or 0.1 µM benzidine. Following a further 6 days of
culture, cells were harvested and total RNA was isolated with
RNAiso Plus (Takara Bio, Inc., Otsu, Japan) following the
manufacturer's protocol. Total RNA was transcribed into cDNA using
AMV reverse transcriptase (Takara Bio, Inc.) according to the
manufacturer's protocol.
qPCR was performed using the Power SYBR-Green Master
Mix (Takara Bio, Inc.) and an Applied Biosystems 7300 Real-Time PCR
Detection System (Thermo Fisher Scientific, Inc.). The 20 µl total
reaction mixture included 10 µl SYBR premix, 0.4 µl Rox, 7.8 µl
dH2O, 0.4 µl forward and 0.4 µl reverse primers, and 1
µl cDNA sample. Each sample was repeated three times.
All primers were synthesized by Invitrogen (Thermo
Fisher Scientific, Inc.). The primers used were: Cyclin D1 forward,
5′-CGTGGCCTCTAAGATGAAGG-3′ and reverse, 5′-TGCGGATGATCTGTTTGTTC-3′;
p21 forward, 5′-GACACCACTGGAGGGTGACT-3′ and reverse,
5′-CAGGTCCACATGGTCTTCCT-3′; PCNA forward,
5′-CTGAAGCCGAAACCAGCTAGACT-3′ and reverse,
5′-TCGTTGATGAGGTCCTTGAGTGC-3′; GAPDH forward,
5′-GCTGCCCAACGCACCGAATA-3′ and reverse, 5′-GAGTCAACGGATTTGGTCGT-3′.
The PCR program included an initial denaturation step at 95°C for
15 sec, followed by 40 cycles of amplification and quantification
at 95°C for 10 sec, 60°C for 30 sec, and 72°C for 30 sec. At the
end of the program, a melting curve analysis was performed. Fold
changes in the expression of each gene were calculated using the
comparative quantitation cycle (Cq) method using the formula
2−∆∆Cq (13).
Cell cycle analysis
SV-HUC-1 (1×106 cells/well) were grown in
6-well plates followed by treatment with 0, 0.001, 0.005, 0.01,
0.05 or 0.1 µM benzidine. Following 6 days of growth, cells were
trypsinized, washed twice with cold PBS, and centrifuged (500 × g
for 5 min at 4°C). The cell pellet was resuspended in 500 µl cold
PBS and fixed in 2-3 ml 70% ethanol at 4°C for 1-14 days. Cells
were centrifuged (500 × g for 5 min at 4°C) and resuspended in 500
µl PBS. Propidium iodide staining buffer was added in the dark at
room temperature for 30 min and cells were then analyzed with flow
cytometry (FACStar cytofluorometer; BD Biosciences, Franklin Lakes,
NJ, USA). Each assay was repeated three times.
Statistical analysis
Statistical analyses were performed with SPSS 17.0
(SPSS, Inc., Chilcago, IL, USA). All data are expressed as mean ±
standard deviation. One-way analysis of variance or the
Kruskal-Wallis test were used to analyze differences among groups.
In case of comparison between two groups, an unpaired Student's
t-test was used. Results are expressed as mean ± standard deviation
from ≥3 independent experiments. P<0.05 was considered to
indicate statistically significant differences.
Results
Benzidine enhances the proliferation
of SV-HUC-1 cells
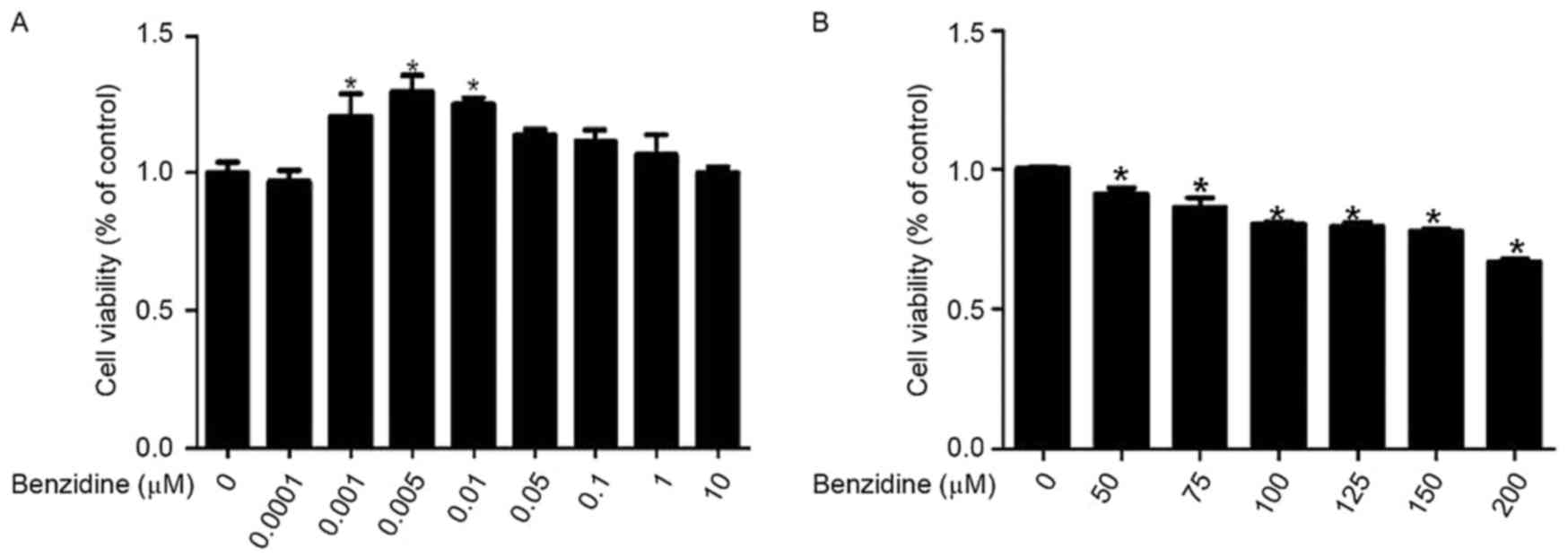
The effect of benzidine on the proliferation of
SV-HUC-1 cells was determined by an MTT assay. The data revealed
that treatment with benzidine at the concentration range of
0.001-0.01 µM for 6 days significantly increased the cell viability
of SV-HUC-1 cells compared with the 0 µM control group (P<0.05;
Fig. 1A), whereas treatment with
benzidine at doses >10 µM induced a toxic effect on SV-HUC-1
cells (Fig. 1B). Therefore, benzidine
at doses from 0.001-0.1 µM was used in subsequent experiments.
Benzidine facilitates SV-HUC-1 cell
transition from G1 to S phase
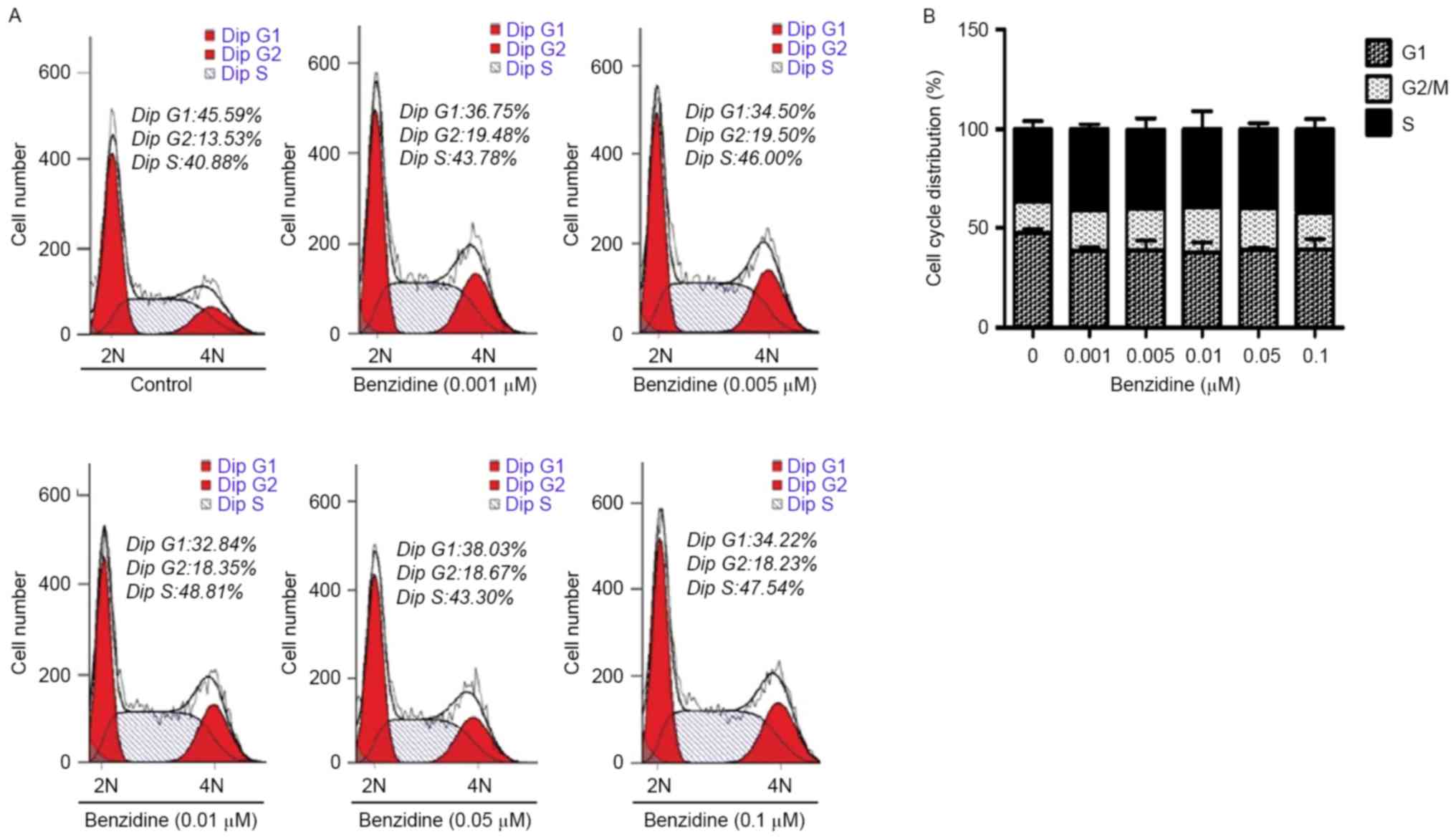
To ascertain that benzidine treatment induced
SV-HUC-1 cell proliferation as demonstrated by the MTT assay data,
flow cytometry was applied to detect alterations in the cell cycle
distribution. Following 6 days of treatment with benzidine, an
increased percentage of cells in the S and G2 phases was observed
(Fig. 2A and B). The fraction of
cells in the S phase increased from 40.88 to 48.81% (P=0.036) and
the fraction of cells in the G2 or M phase increased from 13.53 to
19.50% (P=0.018); the population of cells in the G1 phase decreased
from 45.59 to 34.22% (P<0.01).
Benzidine alters cell cycle associated
marker expression in SV-HUC-1 cells
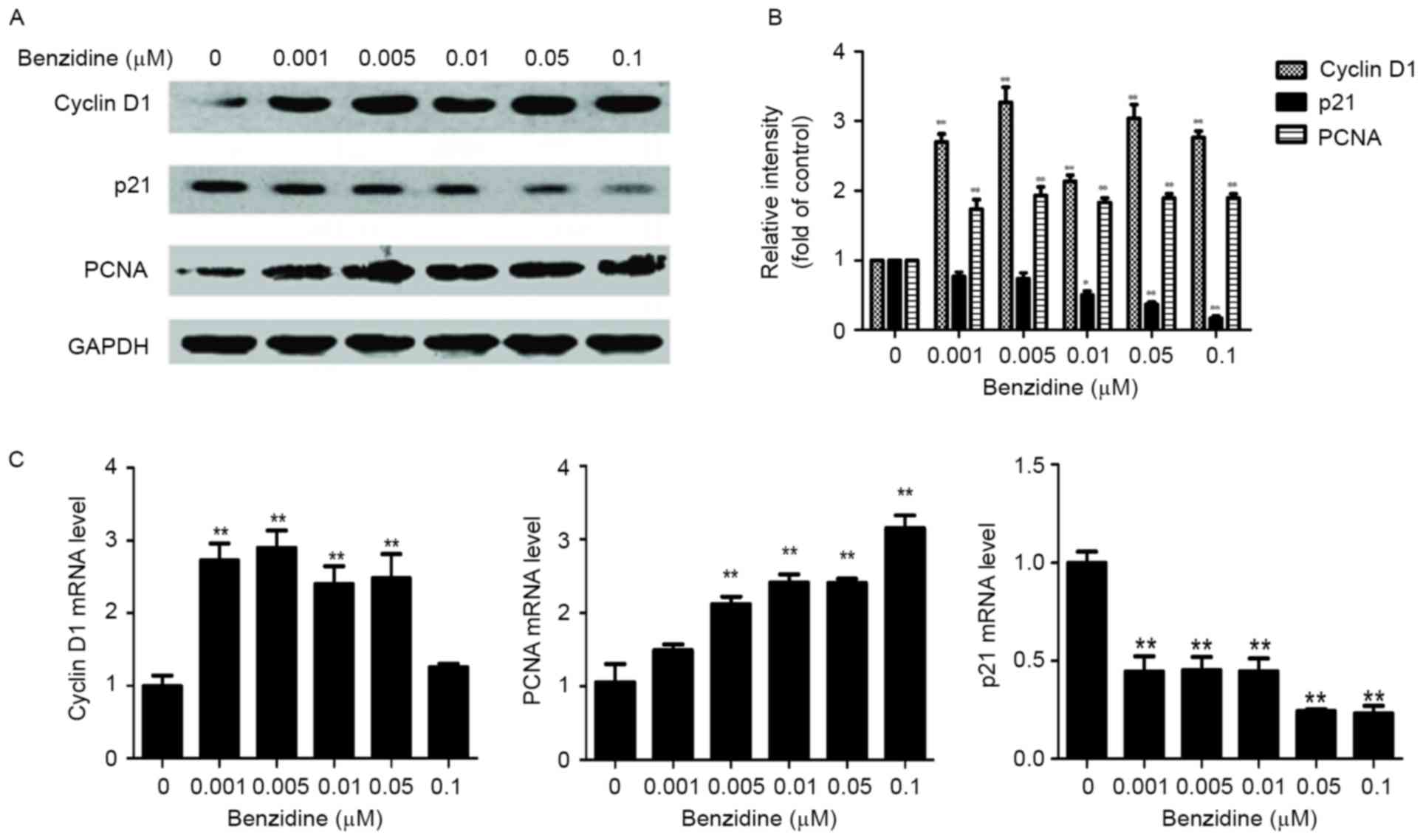
The protein and mRNA levels of cyclin D1, p21 and
PCNA were measured as cell cycle-specific markers. The results
demonstrated that exposure to benzidine significantly increased the
protein levels of cyclin D1 and PCNA (both P<0.01, 0.001-0.1
µM), whereas the p21 protein level was decreased (P<0.01,
0.05-0.1 µM; Fig. 3A and B). The mRNA
levels of cyclin D1 (P<0.01, 0.001-0.05 µM) and PCNA (P<0.01,
0.005-0.1 µM) were also significantly elevated, whereas the p21
mRNA was downregulated (P<0.01, 0.001-0.1 µM; Fig. 3C).
Exposure to benzidine induces
MAPK/AP-1 activation
To determine whether MAPK/AP-1 signaling pathways
were activated in benzidine-mediated SV-HUC-1 proliferation, MAPK
and AP-1 markers were measured with western blotting. Increases in
p-ERK, p-p38 and p-JNK protein levels were observed, whereas total
ERK1/2, p38 and JNK protein levels remained unchanged in cells
treated with benzidine (P<0.05 vs control group) (Fig. 4A and B). The protein levels of members
of the Jun family, including p-c-Jun, JunB and JunD (Fig. 4C and D), and members of the Fos
family, including p-c-Fos, Fos-like antigen 1 and FosB (Fig. 4E and F), were also observed to be
increased (P<0.05).
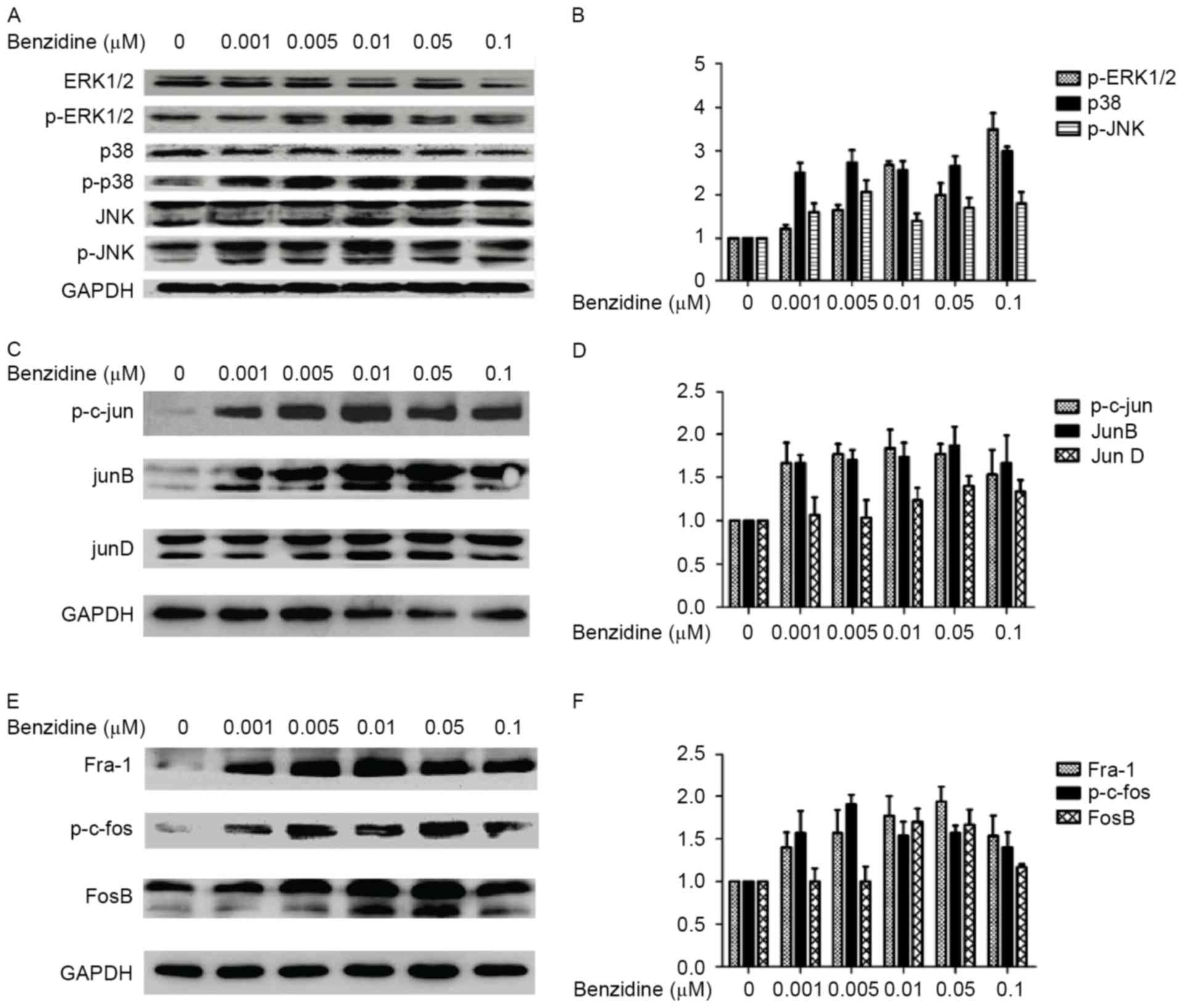 | Figure 4.Effects of benzidine on the expression
and activation of mitogen-activated protein kinases and activating
protein-1 monomers. (A) Total and phosphorylated ERK1/2, p38 and
JNK protein levels were determined by western blotting. The levels
of p-ERK1/2, p-p38 and p-JNK increased without any significant
changes to total ERK1/2, p38 or JNK levels, indicating that
benzidine exposure activated ERK1/2, p38, and JNK. The alterations
to p-ERK1/2, p-p38 and p-JNK protein level occurred particularly at
concentrations of 0.005 or 0.01 µM benzidine. (B) Densitometric
quantification of the data from A. (C) Western blotting analysis of
Jun family proteins. Significant increases in p-c-Jun and JunB
levels were observed, whereas the JunD level was not significantly
increased. (D) Densitometric quantification of the data from C. (E)
The Fos family, including p-c-Fos, FosB and Fra-1, were all
upregulated. (F) Densitometric quantification of the data from E.
Densitometric data are expressed as the mean ± standard deviations
of three independent experiments. p-, phosphorylated; ERK1/2,
extracellular regulated protein kinases 1 and 2; JNK, Jun
N-terminal kinase; Fra-1, Fos-like antigen 1. |
Benzidine-mediated SV-HUC-1 cell
proliferation is suppressed by MAPK-specific inhibitors
MAPK specific inhibitors (including U0126 for
ERK1/2, SB203580 for p38 and SP600125 for JNK) were used to confirm
the association between the activation of MAPKs and
benzidine-induced cell proliferation. When the cells were treated
with the inhibitors, benzidine-elevated cell viability was reversed
(Fig. 5A). The protein level of MAPKs
and cell cycle-associated proteins following exposure to benzidine
and each of the inhibitors was also assessed. The repression of
MAPKs, AP-1 monomers and cell cycle protein expression was detected
(Fig. 5B-D). The results indicated
the importance of MAPK/AP-1 signaling in benzidine-mediated
SV-HUC-1 proliferation.
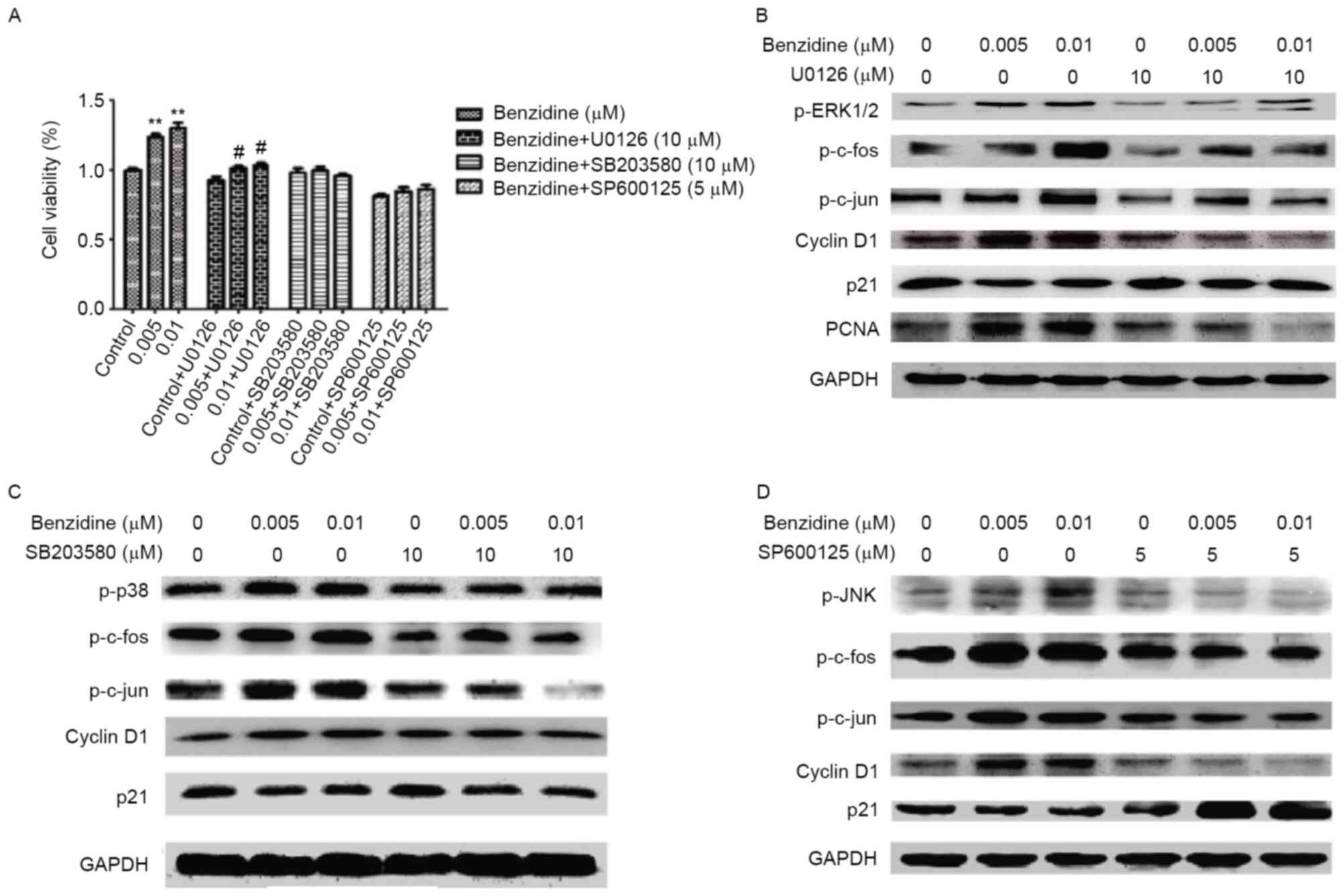 | Figure 5.Proliferation-inducing effect of
benzidine on SV-HUC-1 cells was reversed by MAPK inhibitors. (A)
The relative cell proliferation was suppressed by MAPK pathway
inhibitors. The effect of the p38 inhibitor was the most distinct,
as no increase in cell proliferation following the treatment with
benzidine was observed. Western blot of SV-HUC-1 cells treated with
benzidine and (B) U0126, an ERK1/2 inhibitor, (C) SB203580, a p38
inhibitor or (D) SP600125, a Jun N-terminal kinase inhibitor, for 6
days. Following the treatment with the MAPK inhibitors, the effect
of benzidine treatment on cyclin D1 and p21 expression levels was
suppressed; however, the increase in PCNA protein levels were
inhibited only by U0126. Data is expressed as the means ± standard
deviation of three independent experiments for each treatment.
**P<0.01 vs. control; #P<0.05 vs. control+U0126.
SV-HUC-1, SV-40 immortalized human uroepithelial cells; MAPK,
mitogen-activated protein kinase; ERK1/2, extracellular regulated
protein kinases 1 and 2; PCNA, proliferating cell nuclear
antigen. |
Discussion
Bladder cancer is a major cause of cancer-associated
mortality worldwide (2). The
occupational exposure to benzidine has been established as one of
the risk factors for bladder cancer (3). The mechanism for the occurrence and
development of benzidine-induced bladder cancer is not yet fully
characterized. In the present study, low concentrations of
benzidine enhanced SV-HUC-1 cell proliferation. Activation of
MAPK/AP-1 signaling was also detected. Furthermore, the regulative
role of MAPK signaling in benzidine-induced cell proliferation was
confirmed.
The regulation of the cell cycle is necessary for
cell growth. The escape from the regulation of the cell cycle may
cause unlimited proliferation, which serves a critical role in the
initiation and progression of tumorigenesis (14,15). In
the present study, it was demonstrated for the first time that low
concentrations of benzidine exposure enhanced SV-HUC-1 cell
proliferation, elevated cyclin D1 and PCNA expression, decreased
the expression of p21, and provoked the transition of cells from G1
to S and G2 phase. Cyclin D1 is a nuclear protein that serves a
pivotal role in cell proliferation and the transition from the G1
to S phase. Its expression has been implicated in cancer from a
number of tissue types and in cultured cells (16). Sun et al (17) demonstrated that CyclinD1 protein may
serve a different role in modulating chemoresponses in MCF7 and
MDA-MB231 cells. Additionally, Guo et al (18) indicated that Cyclin D1 is a cell cycle
machine, a sensor of extracellular signals and serves an important
role in G1-S phase progression; their research demonstrated that
cyclinD1 is an activator of cell cycle initiation and
progression.
PCNA is a non-histone nuclear protein that is
necessary for DNA synthesis, and its expression is well documented
as enhancing cancer cell proliferation (19). The PCNA gene contains AP-1 sites in
the promoter region and its expression is regulated by AP-1
activity. The association of PCNA with cancer transformation
resulted in the use of PCNA as a diagnostic and prognostic cell
cycle marker for tumors (20). p21, a
cyclin dependent kinase inhibitor in the G1/S transition, is a
downstream mediator of tumor suppressor p53. It is a
well-characterized partner of PCNA that has been identified as
occurring in a complex of PCNA, cyclin D1 and cyclin-dependent
kinases (CDKs). The p21 protein has two inhibitory effects on the
entry of a cell into S-phase, including the inhibition of CDK
kinase activity and the inhibition of DNA replication via
interactions with PCNA (21). A
previous report demonstrated that p21 expression was associated
with a poor prognosis in patients with bladder cancer (22). Consistent with previous observations,
the results of the present study revealed that benzidine-induced
SV-HUC-1 cell proliferation was associated with the upregulation of
cyclin D1 and PCNA, and the downregulation of p21.
Multiple signaling pathways are associated with the
regulation of the cell cycle. MAPK pathways, which include a series
of protein kinase cascades, serve important roles in various
biological processes, including cell proliferation. The pathway
associated with ERK1/2, MAPK family members, may induce the
initiation and progression of cancer (23,24). In
the present study, it was identified that not only ERK1/2, but also
p38 and JNK, were activated in benzidine-induced SV-HUC-1 cell
proliferation. In addition, cell proliferation was reversed when
MAPK-specific inhibitors were used together with benzidine,
indicating the pivotal role of MAPK activation in benzidine-induced
SV-HUC-1 cell proliferation.
AP-1 is a transcription-activating heterodimer
composed of members of the Jun and Fos families. It is associated
with cell proliferation and differentiation, and the invasion and
metastasis of cancer (25). A
previous study revealed that upregulation of AP-1 enhanced
anaplastic large cell lymphoma progression and dissemination
(26). In the present study,
benzidine promoted the activation of AP-1 monomers and AP-1 was
downregulated following the inhibition of MAPKs, with
benzidine-induced cell proliferation reversed simultaneously. The
results revealed that MAPKs regulated the benzidine-induced
SV-HUC-1 cell proliferation via the regulation of AP-1.
In conclusion, the present study demonstrated that
low concentrations of benzidine lead to increased cell
proliferation via the upregulation of the MAPK/AP-1 pathway in
SV-HUC-1 cells. The inhibition of MAPKs reversed benzidine-induced
SV-HUC-1 proliferation. These findings indicate the role of MAPK
pathways in benzidine-induced pathologies, including tumorigenesis,
and may provide novel insights into the molecular mechanisms that
underlie pathologies induced by benzidine or other aromatic amine
compounds.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81373005,
81072330 and 81202194) and by the Priority Academic Program
Development of Jiangsu Higher Education Institutions.
Funding
The National Natural Science Foundation of China
(Beijing, China; grant nos. 81373005, 81072330 and 81202194).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DXY and CYZ conceived and designed the study. LZ,
TZ, HG, ZQL, ZFL, ZQZ and JM performed the experiments. LZ wrote
the paper. LZ, TZ, HG, ZQL, ZFL, DXY and CYZ reviewed and edited
the manuscript. All authors read and approved the manuscript.
Ethics approval and consent to
participate
The Medical Ethics Committee of Anhui Medical
University gave approval for this study, and all participants gave
informed consent.
Consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
SV-HUC-1
|
SV-40 immortalized human uroepithelial
cell
|
References
|
1
|
Cheung G, Sahai A, Billia M, Dasgupta P
and Khan MS: Recent advances in the diagnosis and treatment of
bladder cancer. BMC Med. 11:132013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Fedewa SA, Miller KD,
Goding-Sauer A, Pinheiro PS, Martinez-Tyson D and Jemal A: Cancer
statistics for Hispanics/Latinos, 2015. CA Cancer J Clin.
65:457–480. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yu MC, Skipper PL, Tannenbaum SR, Chan KK
and Ross RK: Arylamine exposures and bladder cancer risk. Mutat Res
506-507. 1–28. 2002.
|
|
4
|
Letašiová S, Dušinská M, Volkovová K,
Mosoiu C and Bartonva A: Bladder cancer, a review of the
environmental risk factors. Environ Health. 11 Suppl 1:S112012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhao L, Geng H, Liang ZF, Zhang ZQ, Zhang
T, Yu DX and Zhong CY: Benzidine induces epithelial-mesenchymal
transition in human uroepithelial cells through ERK1/2 pathway.
Biochem Biophys Res Commun. 459:643–649. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Correia I, Alonso-Monge R and Pla J: MAPK
cell-cycle regulation in Saccharomyces cerevisiae and Candida
albicans. Future Microbiol. 5:1125–1141. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Thomasova D and Anders HJ: Cell cycle
control in the kidney. Nephrol Dial Transplant. 30:1622–1630. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chang LF and Karin M: Mammalian MAP kinase
signalling cascades. Nature. 410:37–40. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sangrar W, Shi C, Mullins G, LeBrun D,
Ingalls B and Greer PA: Amplified Ras-MAPK signal states correlate
with accelerated EGFR internalization, cytostasis and delayed HER2
tumor onset in Fer-deficient model systems. Oncogene. 34:4109–4117.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dhanasekaran DN and Johnson GL: MAPKs:
Function, regulation, role in cancer and therapeutic targeting.
Oncogene. 26:3097–3099. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ansari KM and Das M: Skin tumor promotion
by argemone oil/alkaloid in mice: Evidence for enhanced cell
proliferation, ornithine decarboxylase, cyclooxygenase-2 and
activation of MAPK/NF-kappaB pathway. Food Chem Toxicol.
48:132–138. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu T, Zhou M, Peng L, Kong S, Miao R, Shi
Y, Sheng H and Li L: Upregulation of CD147 promotes cell invasion,
epithelial-to-mesenchymal transition and activates MAPK/ERK
signaling pathway in colorectal cancer. Int J Clin Exp Pathol.
7:7432–7441. 2014.PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baldi A, De Luca A, Esposito V, Campioni
M, Spugnini EP and Citro G: Tumor suppressors and cell-cycle
proteins in lung cancer. Pathol Res Int. 2011:6050422011.
View Article : Google Scholar
|
|
15
|
Sobus SL and Warren GW: The biologic
effects of cigarette smoke on cancer cells. Cancer. 120:3617–3626.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li X, Gong X, Chen J, Zhang J, Sun J and
Guo M: miR-340 inhibits glioblastoma cell proliferation by
suppressing CDK6, cyclin-D1 and cyclin-D2. Biochem Biophys Res
Commun. 460:670–677. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun Y, Luo D and Liao DJ: CyclinD1 protein
plays different roles in modulating chemoresponses in MCF7 and
MDA-MB231 cells. J Carcinog. 11:122012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guo ZY, Hao XH, Tan FF, Pei X, Shang LM,
Jiang XL and Yang F: The elements of human cyclin D1 promoter and
regulation involved. Clin Epigenetics. 2:63–76. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Malmström PU, Wester K, Vasko J and Busch
C: Expression of proliferative cell nuclear antigen (PCNA) in
urinary bladder carcinoma. Evaluation of antigen retrieval methods.
APMIS. 100:988–992. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Russo G, Zamparelli A, Howard CM, Minimo
C, Bellan C, Carillo G, Califano L, Leoncini L, Giordano A and
Claudio PP: Expression of cell cycle-regulated proteins pRB2/p130,
p107, E2F4, p27, and pCNA in salivary gland tumors: Prognostic and
diagnostic implications. Clin Cancer Res. 11:3265–3273. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Waga S, Hannon GJ, Beach D and Stillman B:
The p21 inhibitor of cyclin-dependent kinases controls DNA
replication by interaction with PCNA. Nature. 369:574–578. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shariat SF, Kim J, Raptidis G, Ayala GE
and Lerner SP: Association of p53 and p21 expression with clinical
outcome in patients with carcinoma in situ of the urinary bladder.
Urology. 61:1140–1145. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang YQ, Wei XL, Liang YK, Chen WL, Zhang
F, Bai JW, Qiu SQ, Du CW, Huang WH and Zhang GJ: Over-expressed
twist associates with markers of epithelial mesenchymal transition
and predicts poor prognosis in breast cancers via ERK and Akt
activation. PloS One. 10:e01358512015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li T, Zhang C, Ding Y, Zhai W, Liu K, Bu
F, Tu T, Sun L, Zhu W, Zhou F, et al: Umbilical cord-derived
mesenchymal stem cells promote proliferation and migration in MCF-7
and MDA-MB-231 breast cancer cells through activation of the ERK
pathway. Oncol Rep. 34:1469–1477. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Han C and Wan F: Research progress on
AP-1. Chin J Cell Biol. 2017.
|
|
26
|
Schiefer AI, Vesely P, Hassler MR, Egger G
and Kenner L: The role of AP-1 and epigenetics in ALCL. Front
Biosci (Schol Ed). 7:226–235. 2015. View
Article : Google Scholar : PubMed/NCBI
|