Introduction
Among all types of lung cancers, adenocarcinoma
accounts for ~40% and generally has both a poor prognosis and
increased potential for metastases (1). Currently, surgery is the primary
treatment for cancer. However, surgery itself can stimulate cell
growth (2), metastasis (3) and recurrence (4) of cancer. Anesthetic agents administered
during surgery might influence the cell activities of cancer
simultaneously (5).
Propofol (PPF) is a sedative-hypnotic agent, which
is widely used in operating rooms and intensive care units (ICU)
for smooth induction and rapid recovery from anesthesia. Potential
anticancer properties of PPF have been considered. PPF inhibits the
invasion and migration of the human lung adenocarcinoma epithelial
A549 cell line by regulating matrix metalloproteinases-2 (MMP-2)
and p38 MAPK signaling pathways (6).
Furthermore, it induces apoptosis in A549 cells through
extracellular signal-regulated kinase 1 and 2 (ERK1/2) pathways
(7). As a urinary trypsin inhibitor,
ulinastatin also has properties that suppress cancer cell growth,
proliferation, differentiation and migration (8–13). Studies
have shown that the anticancer drugs combined with ulinastatin
could offer therapeutic promise for cancer treatment (14–16).
At present, the effects of PPF in combination with
ulinastatin on post-perfusion lung syndrome (17) and acute lung injury (18) have been demonstrated. However,
antitumor effects associated with different ulinastatin and PPF
administration against A549 cells remain unclear and the delivery
of PPF (10, 20 and 30 µM) followed by 200 U/ml ulinastatin
treatments on cancer cells has not been studied. The aim of this
study is to evaluate the synergistic antitumor effect of PPF
followed by ulinastatin against A549 cells. The expression of
p-ERK1/2 and MMP-2 was detected to identify the mechanisms behind
the antitumor effects of PPF (10, 20 and 30 µM) followed by 200
U/ml ulinastatin.
Materials and methods
Cell culture
The A549 cell line was obtained from the Cancer
Research Institute of the Southern Medical University (Guangdong,
China). Cells were maintained at 37°C in a humidified atmosphere of
95% air and 5% CO2 in DMEM/F-12 with 10% fetal bovine
serum (FBS; both Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA), 100 units/ml penicillin and 100 ng/ml streptomycin
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany).
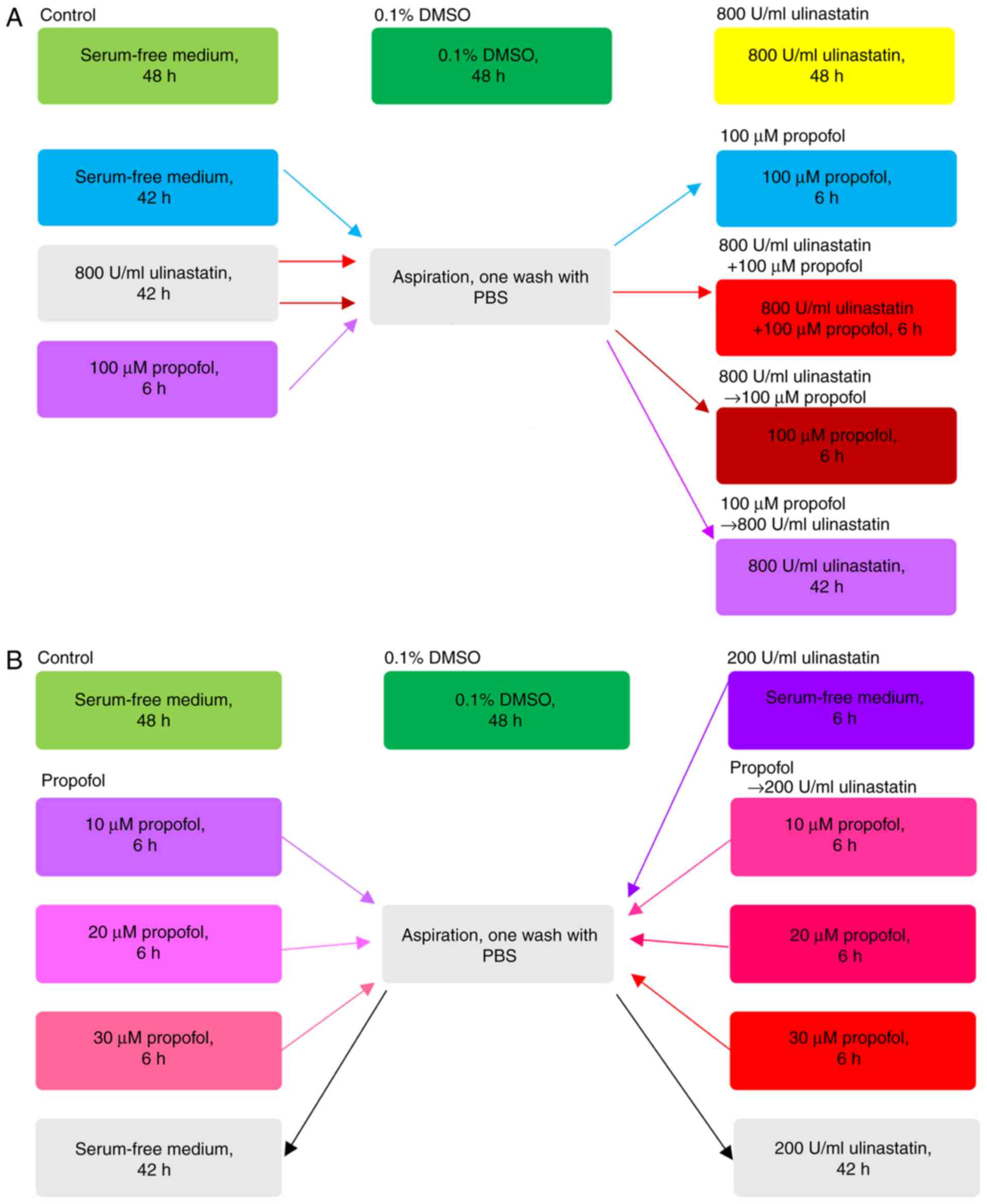
Different treatment schedules for A549
cells
To evaluate the antitumor effect of different
treatment schedules with PPF (Sigma-Aldrich; Merck KGaA) and
ulinastatin (Techpool Bio-Pharma, Guangzhou, China), cells were
treated with seven different treatments (Fig. 1A). According to the studies of
Kobayashi (8) and Song (7), the concentrations of 800 U/ml
ulinastatin and 100 µM PPF were selected as the optimum doses. The
control was defined as continuous treatment with serum-free medium
for 48 h; 0.1% DMSO was defined as continuous treatment with 0.1%
DMSO for 48 h; 800 U/ml ulinastatin was defined as continuous 800
U/ml ulinastatin treatment for 48 h; 100 µM PPF was defined as
pretreatment with serum-free medium for 42 h, followed by
aspiration, one wash with phosphate buffer saline (PBS), and PPF
(100 µM) treatment for 6 h; 800 U/ml ulinastatin +100 µM PPF was
defined as pretreatment with 800 U/ml ulinastatin for 42 h,
followed by aspiration, one wash with PBS, and concomitant
treatment with both 800 U/ml ulinastatin and 100 µM PPF for 6 h;
800 U/ml ulinastatin → 100 µM PPF was defined as pretreatment with
800 U/ml ulinastatin for 42 h, followed by aspiration, one wash
with PBS, and 100 µM PPF treatment for 6 h; 100 µM PPF → 800 U/ml
ulinastatin was defined as pretreatment with 100 µM PPF for 6 h,
followed by aspiration, one wash with PBS, and 800 U/ml ulinastatin
treatment for 42 h. The maximum concentration of DMSO
(Sigma-Aldrich; Merck KGaA) added to the medium in this study was
0.1%.
From the results of the first experimental block, we
found that the antitumor of the sequence PPF → ulinastatin at high
concentrations was the optimum sequence. To verify the synergistic
antitumor effect of PPF → ulinastatin at a low concentration, a
concentration gradient of PPF was generated: This increased from
10, 20, 30 µM, while the concentration of ulinastatin was 200 U/ml
(Fig. 1B). Control was defined as
continuous treatment with serum-free medium for 48 h; 0.1% DMSO was
defined as continuous treatment with 0.1% DMSO for 48 h; 200 U/ml
ulinastatin was defined as pretreatment with serum-free medium for
6 h, followed by aspiration, one wash with PBS, and 200 U/ml
ulinastatin treatment for 42 h; the PPF group was defined as
pretreatment with PPF (10, 20, 30 µM) for 6 h, followed by
aspiration, one wash with PBS, and incubation in serum-free medium
for 42 h; PPF → 200 U/ml ulinastatin groups were defined as
pretreatment with PPF (10, 20, 30 µM) for 6 h, followed by
aspiration, one wash with PBS, and 200 U/ml ulinastatin treatment
for 42 h.
Cell viability inhibition assay
The viability of PPF and ulinastatin against A549
cells was evaluated using the MTT assay. Twenty µl/well of 5 mg/ml
MTT solution (Sigma-Aldrich; Merck KGaA) was added to each well and
the cultures were further incubated for 4 h. Optical density (OD)
was measured at 490 nm on a multimode microplate reader (MDS;
SpectraMax M5, San Jose, CA, USA). Viability inhibition rate was
calculated as follows: Viability inhibition (%)=[1-(OD490 nm of
treated cells-blank/OD490 nm of control cells-blank)] ×100
(19). To determine whether the
sequential treatments with ulinastatin and PPF had a synergistic
effect, the combination index (CI) of each sequential treatment was
analyzed according to the method of Chou and Talaly (20). CI values of <1, 1 and >1
indicate synergistic, additive, and antagonistic effects,
respectively. By using CompuSyn 1.0 software (CompuSyn, Inc.,
Paramus, NJ, USA), the CI value was easily computed.
Cell proliferation cycle
detection
A cell proliferation cycle detection kit (KeyGEN Bio
TECH Ltd., Nanjing, China) was used to detect the cell
proliferation cycle. Pretreated cells were fixed in 70% ethanol at
4°C for 12 h. Cells were aspirated, gently washed twice with
ice-cold PBS, centrifuged at 2,000 × g for 5 min at 4°C, aspirated
once again, and resuspended in 1 ml PBS containing 50 µg/ml RNase A
for 30 min at 37°C. The cells were then incubated with propidium
iodide (PI) for 30 min at 4°C in the dark. The percentage of cells
with different DNA contents relating to different phases of the
cell cycle was measured by fluorescence-activated cell sorting
(FACS) analysis.
Trans-well assay for migration
Pretreated A549 cells (100 µl/chamber at a density
of 10×106 cells/ml) in serum-free medium were placed in
the upper chamber of the trans-well inserts with free Matrigel
matrix basement membrane. To attract cells, medium containing 10%
FBS was placed in the bottom of the chamber. Cells in the upper
membranes were wiped using a cotton swab after incubation for 24 h.
Migratory cells were treated with different treatments:
Pre-fixation with methanol for 20 min, aspiration, one wash with
PBS, followed by 0.1% crystal violet staining for 10 min, before
three washes with PBS. Cells were photographed in 9 predetermined
fields under an inverted microscope (IX71; Olympus, Center Valley,
PA, USA) at magnification, ×200 and images were scored using
CompuSyn software.
Trans-well assay for invasion
Falcon cell culture inserts (pore size of 8 µm;
Corning Inc., Corning, NY, USA) were pretreated with Matrigel
matrix basement membrane (Corning Inc., Corning, NY, USA).
Pretreated A549 cells (100 µl/chamber at a density of
10×106 cells/ml) in serum-free medium were placed in the
upper chamber of the trans-well inserts. To attract cells, medium
containing 10% FBS was placed in the bottom of the chamber. Cells
in the upper membrane were wiped using a cotton swab after
incubation for 24 h. Invasive cells were treated with different
treatments: Pre-fixation with methanol for 20 min, aspiration, one
wash with PBS, followed by 0.1% crystal violet staining for 10 min,
before three washes with PBS. Cells were photographed in 9
predetermined fields under an inverted microscope (IX71; Olympus,
Center Valley, PA, USA) at magnification, ×200 and images were
scored using CompuSyn software.
Annexin V-FITC/PI staining assay for
apoptosis detection
The Annexin V-FITC apoptosis detection kit (Merck,
Darmstadt, Germany) was used to detect apoptosis. Pretreated cells
were harvested before centrifugation at 1,000 × g for 5 min at
18–24°C. Then, cells were resuspended in 500 µl 1× binding buffer,
before incubation with Annexin V for 15 min at 18–24°C in the dark.
The cells were gently resuspended in 500 µl 1X binding buffer. PI
was added in the dark. The number of healthy viable cells,
apoptotic, and necrotic cells were immediately measured by FACS
analysis. The apoptosis rate was calculated as follows: The
apoptosis rate (%)=(number of apoptotic cells)/(number of total
cells observed) ×100 (21).
Western blot analysis
Pretreated cells were washed three times with
ice-cold PBS and lysed with RIPA lysis buffer (1% Triton X-100, 1%
sodium deoxycholate, 0.1% sodium dodecyl sulfate, sodium salt,
phosphatase inhibitor, and phenylmethanesulfonyl fluoride) (CW2333;
CW Bio, Beijing, China). The cell extracts were collected,
incubated for 30 min on ice, and centrifuged for 20 min at 12,000 ×
g at 4°C. The supernatants were used as cell lysates. The cell
lysates were subjected to SDS-polyacrylamide gel electrophoresis
and transferred to polyvinylidene fluoride (PVDF) membranes after
the bicinchoninic acid (BCA) method was used for protein
quantification and equitable application of proteins to the gel.
The membranes were blocked with 5% bovine serum albumin (BSA) in
Tris-buffered saline (TBS) for 1 h at room temperature. After
washing 3 times with TBS, the membranes were incubated in rabbit
polyclonal antibodies (Cell Signaling Technology, Inc., Danvers,
MA, USA) against ERK1/2, p-ERK1/2, and MMP-2 diluted with TBS
containing 0.1% Tween-20 with 5% BSA (TBST) before being gently
agitated overnight at 4°C. After washing 3 times with TBST, the
membranes were incubated in fluorophore-conjugated secondary
antibody (LI-COR Biosciences, Nebraska, USA) dilution buffer (TBS
containing 0.1% Tween-20 with 5% skimmed dry milk) with gentle
agitation for 1 h at room temperature. After washing 3 times with
TBST, the Odyssey Infrared Imaging System (Licor, Lincoln, NE, USA)
was used to detect proteins. The results were analyzed using ImageJ
1.42q software (Wayne Rasband National Institutes of Health,
Bethesda, MD, USA).
Statistical analysis
Data were expressed as mean ± standard deviation
(SD) of three independent experiments. After determination of
variance homogeneity of variance test, the
Least-Significant-Difference and Dunnett' T3 were used to assess
statistical significance, with P<0.05 considered to indicate a
statistically significant difference
Results
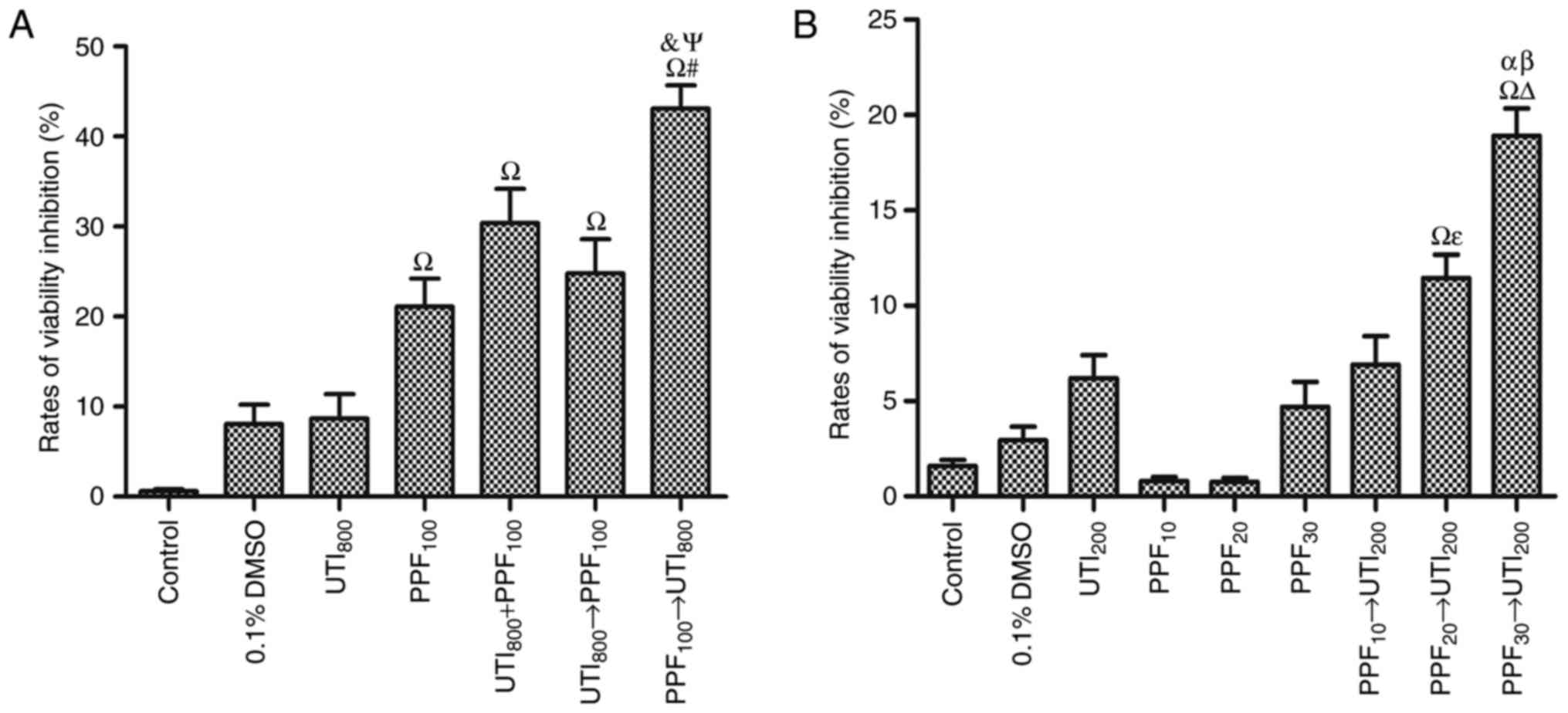
PPF followed by ulinastatin
synergistically inhibited the viability of A549 cells
Fig. 2A shows that 100
µM PPF inhibited the viability of A549 cells. However, 800 U/ml
ulinastatin had no statistically significant in inhibiting the
viability of A549 cells. 100 µM PPF → 800 U/ml ulinastatin was the
optimum sequence in inhibiting the viability of A549 cells. There
was an antagonistic effect when A549 cells were treated with 800
U/ml ulinastatin → 100 µM PPF, with the CI >1. Furthermore,
there was an additive effect when A549 cells were treated with 800
U/ml ulinastatin + 100 µM PPF, with the CI=1. When A549 cells were
treated with 100 µM PPF → 800 U/ml ulinastatin, the CI <1, which
indicates a synergistic effect.
From the results of the first experimental block, we
found that the antitumor effect of the sequence PPF → ulinastatin
at high concentration was the optimum sequence. To verify the
synergistic antitumor effect of PPF → ulinastatin at low
concentration, the results of the second experimental block were as
follows. Fig. 2B demonstrates 200
U/ml ulinastatin and PPF groups (10, 20, 30 µM) did not inhibit the
viability of A549 cells. 10 µM PPF → 200 U/ml ulinastatin did not
significantly inhibit the viability of A549 cells. PPF (20 and 30
µM) → 200 U/ml ulinastatin synergistically inhibited the viability
of A549 cells in a dose-dependent manner associated with PPF
stimulation. There was a demonstrable antagonistic effect when A549
cells were treated with 10 µM PPF → 200 U/ml ulinastatin, where CI
>1, while there was a synergistic effect when A549 cells were
treated with 20 µM PPF →200 U/ml ulinastatin and 30 µM PPF→200 U/ml
ulinastatin, where CI <1.
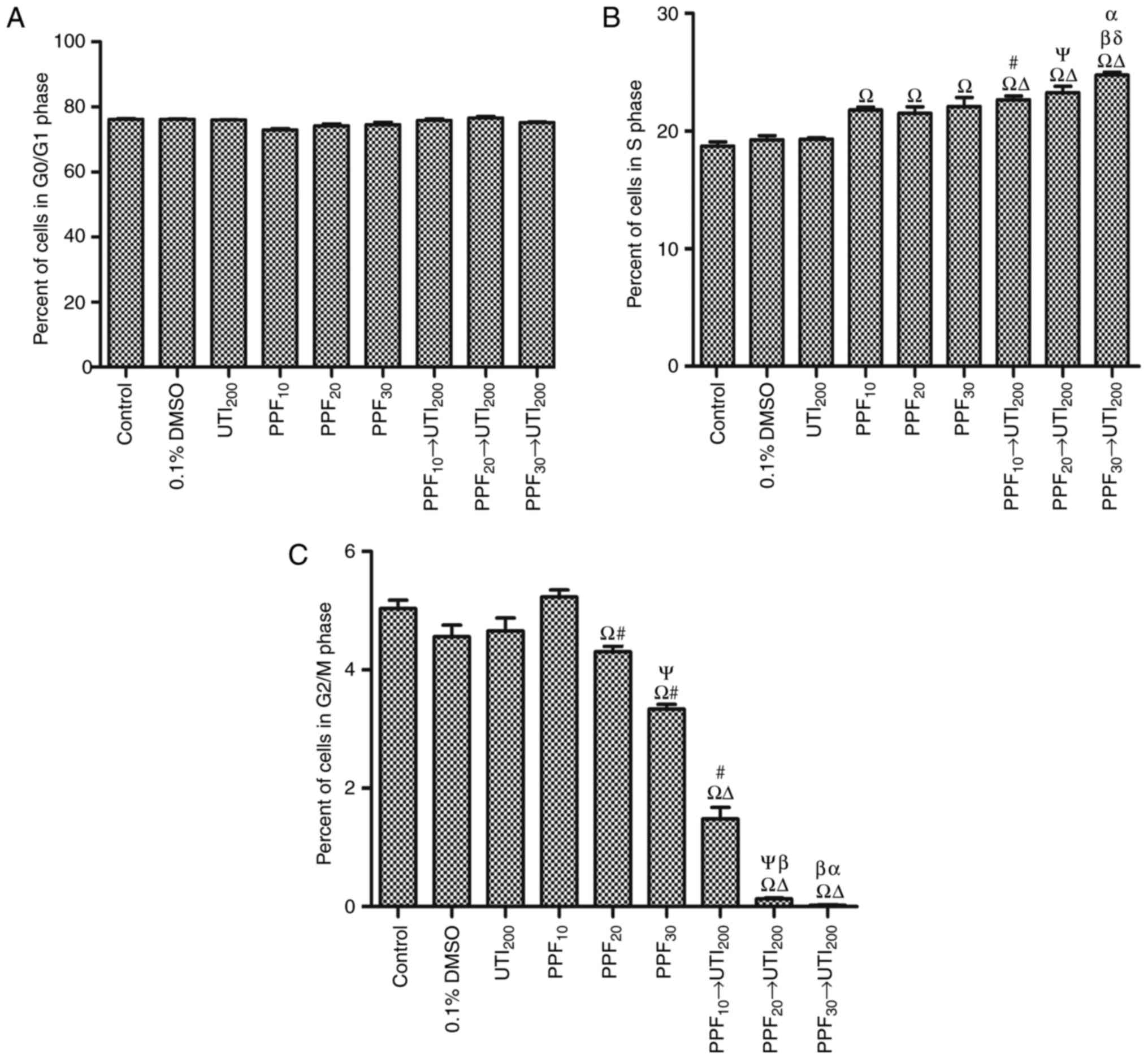
PPF followed by ulinastatin
synergistically increased the number of S cells and reduced the
number of G2/M cells in a PPF dose-dependent manner
There were no statistically significant differences
with respect to the number of G0/G1 cells among groups (Fig. 3A). As shown in Fig. 3B, PPF groups (10, 20, 30 µM) and PPF →
200 U/ml ulinastatin groups significantly increased the number of S
cells, but 0.1% DMSO and 200 U/ml ulinastatin did not significantly
increase the number of S cells. PPF (10, 20 and 30 µM) → 200 U/ml
ulinastatin significantly increased the number of S cells
respectively compared with PPF (10, 20 and 30 µM) and 200 U/ml
ulinastatin in a PPF dose-dependent manner.
Fig. 3C shows the PPF
groups (20, 30 µM) and PPF (10, 20, 30 µM) → 200 U/ml ulinastatin
groups significantly reduced the number of G2/M cells. However,
0.1% DMSO and 200 U/ml ulinastatin did not significantly reduce the
number of G2/M cells. PPF → 200 U/ml ulinastatin groups
synergistically reduced the number of G2/M cells compared with 200
U/ml ulinastatin.
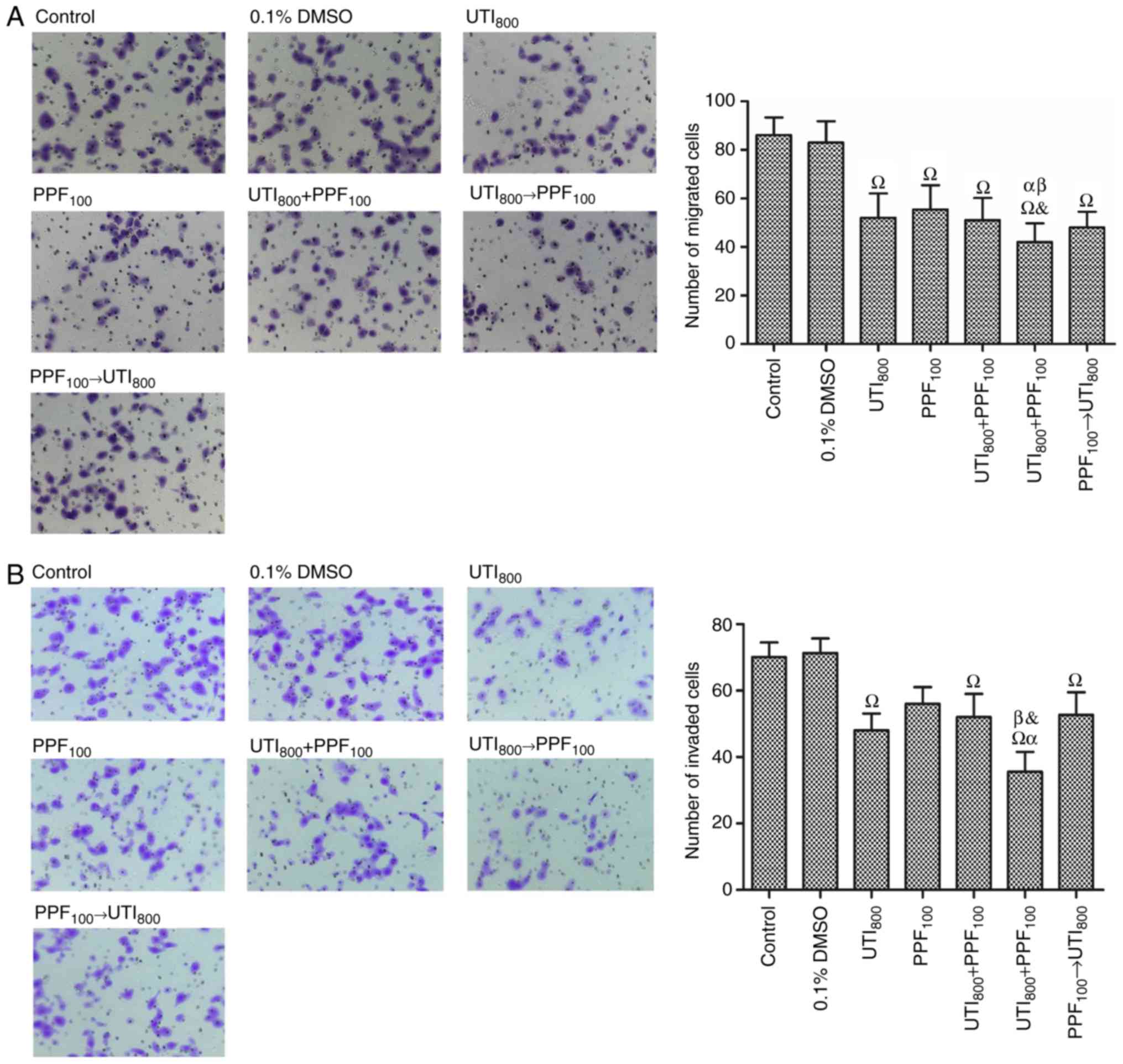
PPF → ulinastatin treatments did not
synergistically inhibit the migration and invasion of A549
cells
A549 cells were harvested and assayed for migration
(Fig. 4A). 800 U/ml ulinastatin → 100
µM PPF significantly reduced the migration of A549 cells compared
with 100 µM PPF, 800 U/ml ulinastatin + 100 µM PPF, and 100 µM
PPF→800 U/ml ulinastatin. There was no statistically significant
difference in the inhibition of migration of A549 cells treated
with 800 U/ml ulinastatin+ 100 µM PPF, 800 U/ml ulinastatin→100 µM
PPF, and 100 µM PPF→800 U/ml ulinastatin, compared with 800 U/ml
ulinastatin.
A549 cells were harvested and assayed for invasion
(Fig. 4B). The group of 100 µM PPF
treated A549 cells (56±5.0) was better than control (70.1±4.4). 800
U/ml ulinastatin→100 µM PPF significantly reduced the invasion of
A549 cells compared with 100 µM PPF, 800 U/ml ulinastatin + 100 µM
PPF, and 100 µM PPF→ 800 U/ml ulinastatin. 100 µM PPF→ 800 U/ml
ulinastatin and 800 U/ml ulinastatin + 100 µM PPF did not
significantly reduce the invasion of A549 cells compared with 100
µM PPF and 800 U/ml ulinastatin.
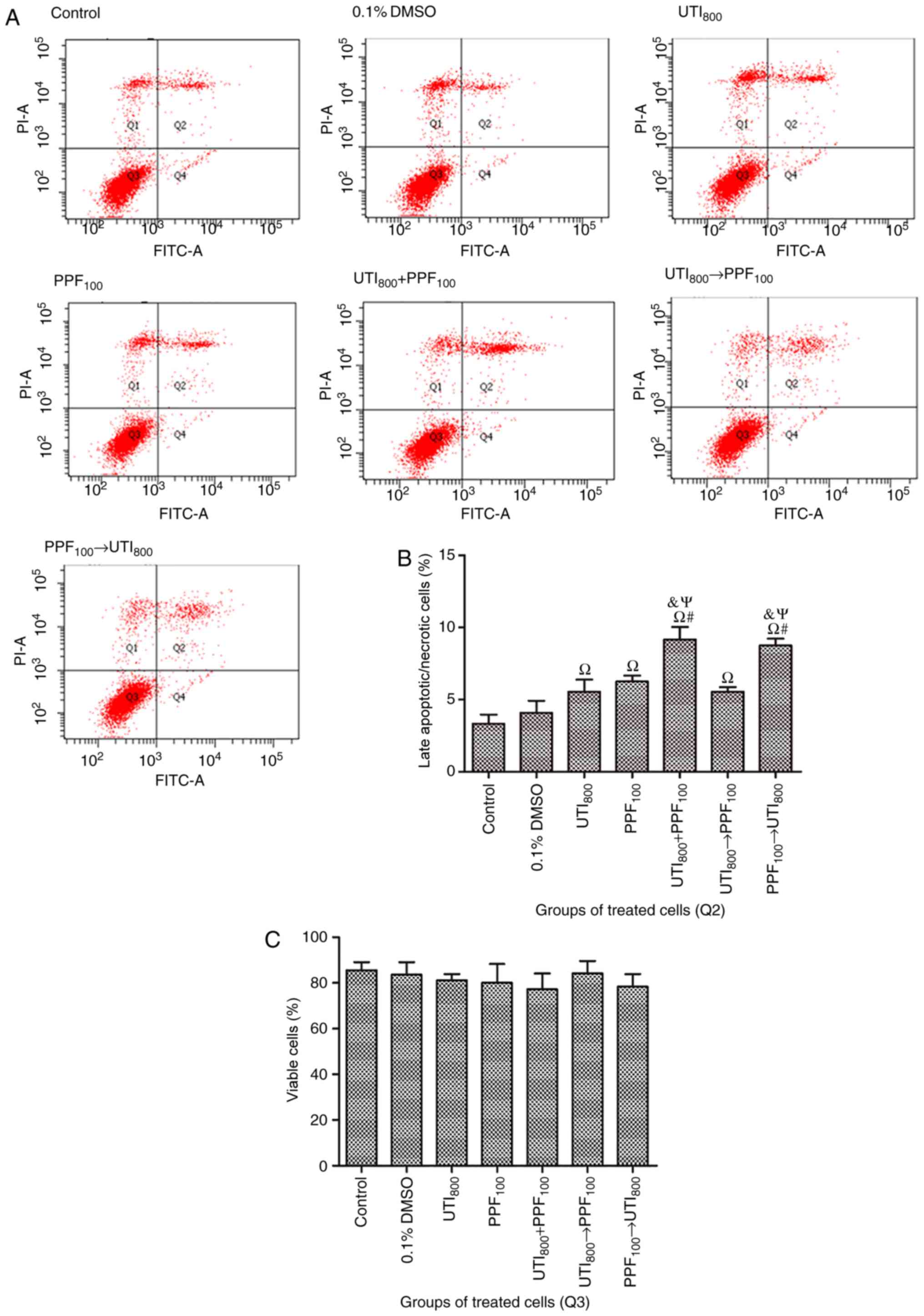
PPF followed by ulinastatin
synergistically stimulated late apoptosis or necrosis in A549
cells
As shown in Fig. 5,
compared with control, 0.1% DMSO did not significantly stimulate
late apoptosis or necrosis in A549 cells. However, 800 U/ml
ulinastatin, 100 µM PPF, 800 U/ml ulinastatin + 100 µM PPF, 800
U/ml ulinastatin → 100 µM PPF, and 100 µM PPF → 800 U/ml
ulinastatin stimulated apoptosis or necrosis in A549 cells. 800
U/ml ulinastatin + 100 µM PPF and 100 µM PPF → 800 U/ml ulinastatin
stimulated late apoptosis or necrosis in A549 cells to a
significantly greater extent than 800 U/ml ulinastatin, 100 µM PPF,
and 800 U/ml ulinastatin → 100 µM PPF treatments. There were no
statistically significant differences on the viability of treated
groups with respect to early apoptotic cells.
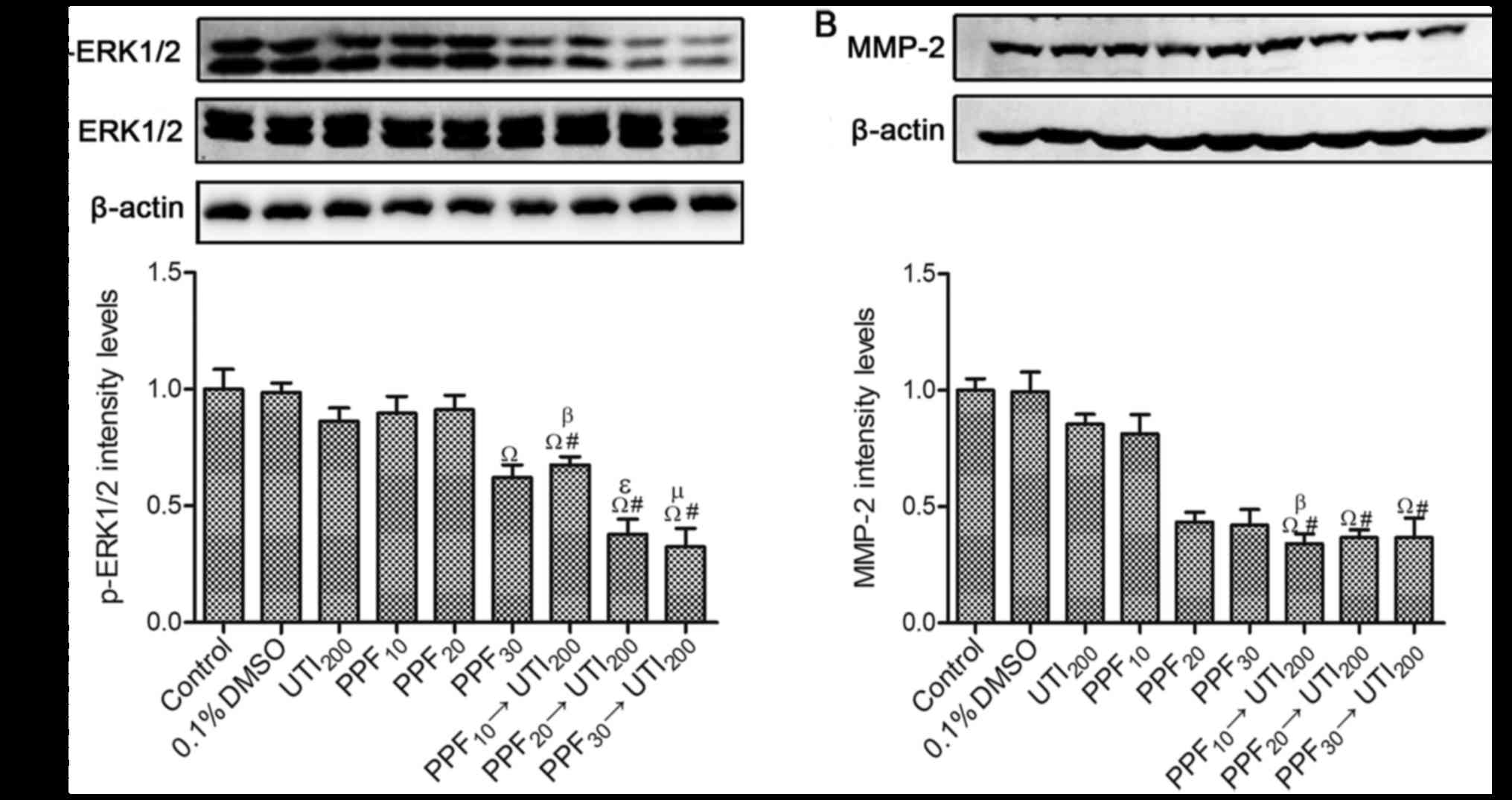
Effects of PPF → ulinastatin at low
concentrations on the expression of p-ERK1/2 and MMP-2
The expression of total ERK1/2 was not significantly
different among all groups. However, the expression of p-ERK1/2 was
different from total ERK1/2 (Fig.
6A). 200 U/ml ulinastatin, 10 µM PPF, and 20 µM PPF did not
downregulate the expression of p-ERK1/2 in A549 cells. However, 30
µM PPF and PPF (10, 20, 30 µM) → 200 U/ml ulinastatin significantly
reduced the expression of p-ERK1/2. The expression of p-ERK1/2 was
synergistically downregulated by PPF (10, 20, 30 µM) → 200 U/ml
ulinastatin.
0.1% DMSO, 200 U/ml ulinastatin, and PPF groups (10,
20, 30 µM) did not downregulate the expression of MMP-2 (Fig. 6B). Compared with 200 U/ml ulinastatin
alone, the expression of MMP-2 was significantly downregulated
after cells were treated with PPF (10, 20, 30 µM) →200 U/ml
ulinastatin. 10 µM PPF → 200 U/ml ulinastatin downregulated the
expression of MMP-2 compared with 10 µM PPF. However, there was no
statistically significant difference when cells were treated with
20 µM PPF→200 U/ml ulinastatin or 30 µM PPF→200 U/ml ulinastatin,
compared with 20 µM PPF or 30 µM PPF.
Discussion
Among all types of lung cancers, adenocarcinoma
accounts for ~40% of cancer and generally has a poor prognosis
(1). The A549 cell line is the
typical cell line in human lung adenocarcinoma and surgery is the
primary treatment for lung cancer. The immunosuppressive effects of
surgery are well known with respect to cancer progression (22). Surgery can also generate a
microenvironment that is abundant in inflammatory cells and growth
factors, including potent angiogenic, lymphangiogenic growth
factors, cytokines, and proteases (23). Granov et al found that cancer
patients were susceptible in developing acute lung lesions (ALL)
and adult respiratory distress syndrome (ARDS) postoperatively
(24). Use of ulinastatin and PPF may
benefit ARDS patients through different mechanisms (17). PPF exhibits protective effects
including an antiinflammatory effect, enhancement of antitumor
immunity, reduction of the concentration of cytokines (IL-1, TNF-α
and IL-6) and natural killer cell function preservation (25–28).
Ulinastatin also improves the immunosuppressive state during
surgery for malignancy (29). In view
of the clinical translation of our results, the optimum
administration protocol (PPF → ulinastatin) may benefit ARDS
patients and inhibit lung adenocarcinoma cells, which can improve
the postoperative prognosis of lung adenocarcinoma patients. PPF →
ulinastatin synergistic antitumor effects may be importantly
related to the immune microenvironment. As ERK1/2 phosphorylation
is an important step for cytokine secretion such as TNF-α (30) and IL-1β (31), PPF → ulinastatin may synergistically
reduce cytokine secretion of TNF-α and IL-1β by inhibiting ERK1/2
phosphorylation in A549 cells.
With respect to clinical application, 100 µM PPF and
800 U/ml ulinastatin was more potent but PPF (6.2–33.7 µM)
administered through Target Controlled Infusion (TCI) is widely
used in clinical applications (e.g., the maintenance of general
anesthesia), and 200 U/ml ulinastatin is introduced in a pharmacy.
To verify the synergistic effect of PPF → ulinastatin at a clinical
concentration, we tested several concentration gradients of PPF
using TCI (10, 20, 30 µM) and a clinical concentration of
ulinastatin (200 U/ml). We demonstrated that PPF → ulinastatin
treatments effectively inhibited the viability of A549 cells and
stimulated late apoptosis or necrosis cells. However, PPF →
ulinastatin treatments did not synergistically inhibit the
migration and invasion of A549 cells. From the results, we found
that the molecular mechanisms regulating the viability and late
apoptosis or necrosis of A549 cells might share common properties
from which regulating the migration and invasion of A549 cells was
different.
In our investigation, the MTT assay clearly
indicated that PPF → ulinastatin treatments had a synergistic
effect at high and low concentrations in inhibiting A549 cell
viability. PPF → ulinastatin synergistically inhibited A549 cell
viability, which could be attributed to the different timing events
in the cell cycle: PPF → ulinastatin treatments synergistically
increased the number of S cells and synergistically reduced the
number of G2/M cells in a PPF dose-dependent manner. The G2/M DNA
damage checkpoint serves to prevent the cell from entering M-phase,
which can result in genomic damage. DNA damage can activate the
DNA-PK/ATM/ATR kinases, which result in two parallel cascades that
ultimately serve to inactivate the cyclin B-cdc2 kinase. The first
cascade rapidly inhibits progression into mitosis: The Chk kinases
phosphorylate and inactivate cdc25, which prevents activation of
cdc2 (32,33). Phosphorylated ERK1/2 (p-ERK1/2)
activates cdc25, which promotes the cell from entering M-phase
(34). ERK1/2 is activated through
phosphorylation, which plays an important role in the regulation of
fundamental cellular processes including proliferation, survival,
differentiation, migration (35–37), and
apoptosis (38). It has been reported
that 100 µM (7) PPF can downregulate
the expression of p-ERK1/2 in A549 cells. Liposoluble PPF is liable
to pass through the A549 cytomembrane into the cytoplasm and
nucleus, which may inactivate ERK1/2 and/or promote DNA damage in
A549 cells. This DNA damage may improve the dosing of soluble
ulinastatin in the cytoplasm and nucleus of A549 cells, which
synergistically inactivate p-ERK1/2, cdc2, and/or cdc25. The
DNA-PK/ATM/ATR kinases from DNA damage are inactivated by
ulinastatin, which counteracts the DNA damage in A549 cells caused
by PPF. We believe these were the reasons why the treatment with
PPF → ulinastatin was more effective than both ulinastatin → PPF
and the simultaneous combination.
The suppression of ERK1/2 and hypoxia pathways
resulted in the suppression of MMP-2, MMP-9, and MMP-7 expression
in A549 cell metastasis (39).
Metalloproteinases, particularly MMP-2, play an important role in
the regulation of fundamental cancer cellular processes including
cell growth, invasion, inflammation and angiogenesis (40). PPF suppresses the invasion and
migration of A549 human lung adenocarcinoma epithelial cells by
downregulating the expression of MMP-2 and p38 MAPK signaling
(6). It has been reported that a
urinary trypsin inhibitor-like inhibitor can be isolated from human
lung cancer tissue (41). In our
study, we suspected that ulinastatin did not statistically inhibit
the viability of A549 cells because of the presence of the
inhibitor from human lung cancer tissues or the low concentration
of ulinastatin. Two hundred U/ml of ulinastatin alone did not
effectively inhibit the expression of MMP-2 due to the inhibitor or
the low concentration of ulinastatin. PPF alone did not effectively
inhibit the expression of MMP-2 due to the low concentration of
PPF. However, the expression of MMP-2 after PPF → ulinastatin
treatment was synergistically downregulated as pretreated A549
cells with PPF could regulate expression of the inhibitor.
In our study, we have partly elucidated the
underlying mechanisms of the synergistic antitumor effect of PPF →
ulinastatin at clinical concentrations, and detected p-ERK1/2 and
MMP-2. DNA-PK/ATM/ATR, Cyclin B-cdc2, and Chk kinases in A549 cells
will be detected to verify the mechanism of G2/M DNA damage and how
this relates to synergistic suppression of the human lung
adenocarcinoma epithelial A549 cell line with PPF treatment
followed by ulinastatin.
In summary, we conclude that PPF (20, 30 µM)
followed by 200 U/ml ulinastatin treatments synergistically
stimulated a significant proportion of A549 cells in S phase,
synergistically reduced the percent of A549 cells in G2/M phase,
and synergistically suppressed viability, which could possibly be
related to regulating the expression of p-ERK1/2 and MMP-2 in A549
cells.
Acknowledgements
Not applicable.
Funding
The present study was supported in part by the
Scheme of the Guandong Province Sciences Research (grant no.
2012A030400014).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
PL and PG assisted in the design of the study,
conducedt the study, analyze the data and wrote the manuscript. CL
assisted in the analysis of the statistics, interpreted the data
and graphic illustrations and drafted the manuscript. MH, XZ, CL,
JT, WW and WL collaborated in the design of the study and assisted
in writing the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yan KH, Lee LM, Yan SH, Huang HC, Li CC,
Lin HT and Chen PS: Tomatidine inhibits invasion of human lung
adenocarcinoma cell A549 by reducing matrix metalloproteinases
expression. Chem Biol Interact. 203:580–587. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tyzzer EE: Factors in the production and
growth of tumor metastases. J Med Res. 28:309–332.1.
1913.PubMed/NCBI
|
|
3
|
Demicheli R, Retsky MW, Hrushesky WJ, Baum
M and Gukas ID: The effects of surgery on tumor growth: a century
of investigations. Ann Oncol. 19:1821–1828. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Allen-Mersh TG, McCullough TK, Patel H,
Wharton RQ, Glover C and Jonas SK: Role of circulating tumour cells
in predicting recurrence after excision of primary colorectal
carcinoma. Br J Surg. 94:96–105. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cassinello F, Prieto I, del Olmo M, Rivas
S and Strichartz GR: Cancer surgery: How may anesthesia influence
outcome? J Clin Anesth. 27:262–272. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu KC, Yang ST, Hsia TC, Yang JS, Chiou
SM, Lu CC, Wu RS and Chung JG: Suppression of cell invasion and
migration by propofol are involved in down-regulating matrix
metalloproteinase-2 and p38 MAPK signaling in A549 human lung
adenocarcinoma epithelial cells. Anticancer Res. 32:4833–4842.
2012.PubMed/NCBI
|
|
7
|
Song J, Shen Y, Zhang J and Lian Q: Mini
profile of potential anticancer properties of propofol. PLoS One.
9:e1144402014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kobayashi H, Shinohara H, Takeuchi K, Itoh
M, Fujie M, Saitoh M and Terao T: Inhibition of the soluble and the
tumor cell receptor-bound plasmin by urinary trypsin inhibitor and
subsequent effects on tumor cell invasion and metastasis. Cancer
Res. 54:844–849. 1994.PubMed/NCBI
|
|
9
|
Kobayashi H, Gotoh J, Fujie M and Terao T:
Characterization of the cellular binding site for the urinary
trypsin inhibitor. J Biol Chem. 269:20642–20647. 1994.PubMed/NCBI
|
|
10
|
Kobayashi H, Gotoh J, Kanayama N,
Hirashima Y, Terao T and Sugino D: Inhibition of tumor cell
invasion through matrigel by a peptide derived from the domain II
region in urinary trypsin inhibition. Cancer Res. 55:1847–1852.
1995.PubMed/NCBI
|
|
11
|
Kobayashi H, Shinohara H, Fujie M, Gotoh
J, Itoh M, Takeuchi K and Terao T: Inhibition of metastasis of
Lewis lung carcinoma by urinary trypsin inhibitor in experimental
and spontaneous metastasis models. Int J Cancer. 63:455–462. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kobayashi H: Mechanism of tumor
cell-induced extracellular matrix degradation-inhibition of
cell-surface proteolytic activity might have a therapeutic effect
on tumor cell invasion and metastasis. Nihon Sanka Fujinka Gakkai
zasshi. 48:623–632. 1996.(In Japanese). PubMed/NCBI
|
|
13
|
Yoshioka I, Tsuchiya Y, Aozuka Y, Onishi
Y, Sakurai H, Koizumi K, Tsukada K and Saiki I: Urinary trypsin
inhibitor suppresses surgical stress-facilitated lung metastasis of
murine colon 26-L5 carcinoma cells. Anticancer Res. 25:815–820.
2005.PubMed/NCBI
|
|
14
|
Kobayashi H, Shinohara H, Gotoh J, Fujie
M, Fujishiro S and Terao T: Anti-metastatic therapy by urinary
trypsin inhibitor in combination with an anti-cancer agent. Br J
Cancer. 72:1131–1137. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Song B, Bian Q, Shao CH, Li G, Liu AA,
Jing W, Liu R, Zhang YJ, Zhou YQ, Hu XG and Jin G: Ulinastatin
reduces the resistance of liver cancer cells to epirubicin by
inhibiting autophagy. PLoS One. 10:e01206942015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gao F, Sun Z, Sun X, Zhang Y, Wang H,
Zhong B, Luo J and Zhao X: Ulinastatin exerts synergistic effects
with taxotere and inhibits invasion and metastasis of breast cancer
by blocking angiogenesis and the epithelial-mesenchymal transition.
Cancer Biother Radiopharm. 28:218–225. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yuan SM: Postperfusion lung syndrome:
Respiratory mechanics, respiratory indices and biomarkers. Ann
Thorac Med. 10:151–157. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu HL and Qian YN: Protective effects of
propofol combined with ulinastatin on acute lung injury induced by
endotoxin in rats. Zhongguo Ying Yong Sheng Li Xue Za Zhi.
29:56–57. 622013.(In Chinese). PubMed/NCBI
|
|
19
|
Vinodhkumar R, Song YS, Ravikumar V,
Ramakrishnan G and Devaki T: Depsipeptide a histone deacetlyase
inhibitor down regulates levels of matrix metalloproteinases 2 and
9 mRNA and protein expressions in lung cancer cells (A549). Chem
Biol Interact. 165:220–229. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ashton JC: Drug combination studies and
their synergy quantification using the Chou-Talalay method-letter.
Cancer Res. 75:24002015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yousef BA, Hassan HM, Guerram M, Hamdi AM,
Wang B, Zhang LY and Jiang ZZ: Pristimerin inhibits proliferation,
migration and invasion, and induces apoptosis in HCT-116 colorectal
cancer cells. Biomed Pharmacother. 79:112–119. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Page GG: Surgery-induced immunosuppression
and postoperative pain management. AACN Clin Issues. 16:302–309;
quiz 416–418. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Coussens LM and Werb Z: Inflammation and
cancer. Nature. 420:860–867. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Granov AM, Rozenberg OA, Tsybul'kin EK,
Erokhin VV, Khubulava GG, Likhvantsev VV, Osovskikh VV, Bautin AE,
Gavrilin SV, Kazennov VV, et al: Critical state medicine.
Surfactant therapy of adult respiratory distress syndrome (results
of multicenter studies). Vestn Ross Akad Med Nauk. 34–38. 2001.(In
Russian). PubMed/NCBI
|
|
25
|
Ke JJ, Zhan J, Feng XB, Wu Y, Rao Y and
Wang YL: A comparison of the effect of total intravenous
anaesthesia with propofol and remifentanil and inhalational
anaesthesia with isoflurane on the release of pro- and
anti-inflammatory cytokines in patients undergoing open
cholecystectomy. Anaesth Intensive Care. 36:74–78. 2008.PubMed/NCBI
|
|
26
|
Kushida A, Inada T and Shingu K:
Enhancement of antitumor immunity after propofol treatment in mice.
Immunopharmacol Immunotoxicol. 29:477–486. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou W, Fontenot HJ, Wang SN and Kennedy
RH: Propofol-induced alterations in myocardial beta-adrenoceptor
binding and responsiveness. Anesth Analg. 89:604–608. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
González-Correa JA, Cruz-Andreotti E,
Arrebola MM, López-Villodres JA, Jódar M and De La Cruz JP: Effects
of propofol on the leukocyte nitric oxide pathway: In vitro and ex
vivo studies in surgical patients. Naunyn Schmiedebergs Arch
Pharmacol. 376:331–339. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hosokawa T, Hori Y, Nakagawa H, Nakagawa
M, Hashimoto T and Miyazaki M: Effect of urinastatin on immunity
during anesthesia and surgery for malignant disease. Masui.
38:1341–1348. 1989.PubMed/NCBI
|
|
30
|
Guzmán-Mejía F, López-Rubalcava C and
González-Espinosa C: Stimulation of nAchRα7 receptor inhibits TNF
synthesis and secretion in response to LPS treatment of mast cells
by targeting ERK1/2 and TACE activation. J Neuroimmune Pharmacol.
13:39–52. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mezzasoma L, Antognelli C and Talesa VN: A
novel role for brain natriuretic peptide: Inhibition of IL-1β
secretion via downregulation of NF-kB/Erk 1/2 and
NALP3/ASC/caspase-1 activation in human THP-1 monocyte. Mediators
Inflamm. 2017:58583152017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Al-Ejeh F, Kumar R, Wiegmans A, Lakhani
SR, Brown MP and Khanna KK: Harnessing the complexity of DNA-damage
response pathways to improve cancer treatment outcomes. Oncogene.
29:6085–6098. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Okumura E, Fukuhara T, Yoshida H, Hanada
Si S, Kozutsumi R, Mori M, Tachibana K and Kishimoto T: Akt
inhibits Myt1 in the signalling pathway that leads to meiotic
G2/M-phase transition. Nat Cell Biol. 4:111–116. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yan Y, Black CP and Cowan KH:
Irradiation-induced G2/M checkpoint response requires ERK1/2
activation. Oncogene. 26:4689–4698. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cristea S and Sage J: Is the canonical
RAF/MEK/ERK signaling pathway a therapeutic target in SCLC? J
Thorac Oncol. 11:1233–1241. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Frémin C, Saba-El-Leil MK, Lévesque K, Ang
SL and Meloche S: Functional redundancy of ERK1 and ERK2 MAP
kinases during development. Cell Rep. 12:913–921. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fey D, Matallanas D, Rauch J, Rukhlenko OS
and Kholodenko BN: The complexities and versatility of the
RAS-to-ERK signalling system in normal and cancer cells. Semin Cell
Dev Biol. 58:96–107. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Persaud SD, Park SW, Ishigami-Yuasa M,
Koyano-Nakagawa N, Kagechika H and Wei LN: All trans-retinoic acid
analogs promote cancer cell apoptosis through non-genomic Crabp1
mediating ERK1/2 phosphorylation. Sci Rep. 6:223962016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lee SH, Jaganath IB, Manikam R and Sekaran
SD: Inhibition of Raf-MEK-ERK and hypoxia pathways by Phyllanthus
prevents metastasis in human lung (A549) cancer cell line. BMC
Complement Altern Med. 13:2712013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: Regulators of the tumor microenvironment. Cell.
141:52–67. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Okumichi T, Nishiki M, Takasugi S, Toki N
and Ezaki H: Isolation of urinary trypsin inhibitor-like inhibitor
from human lung cancer tissue. Cancer Res. 44:2011–2015.
1984.PubMed/NCBI
|