Introduction
Insulin, insulin-like growth factors (IGFs), and
IGF-binding proteins (IGFBs) are involved in numerous biological
processes such as cellular growth, proliferation, metabolism,
glucose homeostasis, cell differentiation, and apoptosis; they are
implicated in the autocrine/paracrine stimulation of a variety of
malignancies (1,2). Insulin receptor substrates (IRSs) are
adaptors of the insulin/IGF signaling pathways, with the ability to
modulate and coordinate multiple signaling cascades, transmitting
upstream signals to intracellular pathways, including the
PI3K/AKT/mTOR and MAP kinase pathways (3). Deregulation of these pathways might
increase the risk of several types of cancers, including prostate,
colon, liver, pancreas, kidney, lung, and breast cancers (2,4–6). In vivo and in vitro
experimental models have highlighted the role of increased insulin
and IGF signaling in enhancing tumorigenesis (7). High IGF-1 and low IGFBP3 plasma levels
indicate an increased risk of cancers (8). IGFBP3, a mediator of apoptosis, which
has opposing effects to those of IGF1, can inhibit the growth of
human breast cancer cells with HER2 overexpression (9,10).
IGF-1 receptors are widely distributed in many human
malignancies (11). The upregulation
of IGF-1 receptor signaling may contribute to resistance to
therapies (12). Following the
activation of IGF-1 receptors, IRSs are rapidly phosphorylated on
tyrosine residues, revealing the docking site for multiple
SH-2-containing proteins such as p85, Nck, Crk, Fyn, Syp, and SHP2
at the carboxy terminus (13). The
phosphorylation of IRSs leads to the transmission of mitogenic,
anti-apoptotic, and anti-differentiation signals to tumor cells.
Among the six family members of IRSs, IRS1 and IRS2 are widely
expressed in human tissues (14). An
increasing amount of evidence has revealed that they are involved
in tumor progression, including cell proliferation, adhesion, and
migration (15,16). The pathological mechanisms
contributing to malignancy could be a function of gene
amplification, influenced by the feedback loops of other mutations
and constitutively phosphorylated proteins. The overexpression of
IRS1 or IRS2 may result in palpable tumors of the mammary glands,
which exhibit a unique histopathology associated with the
activation of β-catenin in murine models (17,18). In
breast cancer cells, the overexpression of IRS1 and IRS2 increases
tumor proliferation and motility, respectively (19). IRS1 is implicated in IGF-mediated
proliferation (15), whereas IRS2 is
predominantly involved in tumor adhesion and migration following
IGF-1 stimulation (3,15,19–21). The
metastatic potential of tumors might be impeded by IRS deletion
(22).
IRS2 has been reported to be overexpressed in
hepatocellular carcinoma (HCC) as an early event or implicated in
the later stages of tumorigenesis (4). IRS2 overexpression may promote the
survival of tumor cells independently of IRS1 (4). The protective effect of IRS2
overexpression against apoptosis is also implicated in liver tumor
progression. Our previous data have demonstrated that IRS2
overexpression is correlated with copy number amplification and
associated with poor disease-free survival (DFS) in intrahepatic
cholangiocarcinoma (iCCA) (23). By
inducing IRS2 overexpression, the mobility potential of iCCA cells
can be increased. However, further studies are needed to elucidate
the role of IRS2 overexpression in migration and metastasis.
In the present study, we examined the
clinicopathological features of IRS2 expression in 86 cases of iCCA
and assessed the relationship between IRS2 overexpression and
epithelial-mesenchymal transition (EMT) in cell lines. We aimed to
examine the clinicopathological significance of IRS2 overexpression
and whether this aberration correlates with EMT in iCCA.
Materials and methods
Tumor materials
We selected 86 formalin-fixed, paraffin-embedded
tumor samples from the collection at the Department of Pathology,
Chang Gung Memorial Hospital, (Kaohsiung, Taiwan). Then, we
reviewed hematoxylin and eosin-stained slides and medical records,
defined survival time, and constructed tissue microarrays for IRS2
immunostaining as previously described (23). The study was approved by the
Institutional Review Board of Chang Gung Medical Foundation, in
accordance with the Helsinki Declaration (IRB 103-0818C,
103-4961B).
Immunohistochemical (IHC)
analysis
IHC staining was performed as previously described
(23). The primary antibody against
IRS2 (1:150; Abcam, Cambridge, UK) was used with the PicTureTM-Plus
kit (ZYMED® 2nd Generation Polymer Detection System;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). The labeling
intensities were 0 (negative), 1 (weak), 2 (moderate), and 3
(strong), and the percentages of tumor cells with cytoplasmic
immunoreactivity for IRS2 were counted in 5% increments. The two
scores were multiplied to calculate the expression index. Only
cases containing two or more analyzable cores were scored, and the
scores of multiple cores from the same patient were averaged to
obtain the mean expression index. Whole sections were used for IHC
staining in cases with non-informative tissue cores (no tumor, or
analyzable cores <2). The expression index values were evaluated
by two pathologists blinded to the clinicopathological data and
averaged. After testing a series of cutoff values, the IRS2 protein
was regarded as overexpressed when the expression index was
>150.
Cell lines and stable
transfection
The iCCA cell lines (RBE, SNU1079, and SSP25) were
purchased from the Korean Cell Line Bank (Seoul, South Korea) and
the Riken BRC Cell Bank (Koyadai, Japan). Tumor cell lines were
cultured in RPMI medium (Gibco; Thermo Fisher Scientific, Inc.) as
previously described (23). Cells
were transfected with pCMV-IRS2 entry vector using Lipofectamine
2000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according
to the manufacturer's instructions. Then, the cells were selected
with complete medium containing G418 (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). The medium was changed every fourth day.
Positive clones were selected through resistance against G418 and
characterized for DDK and IRS2 expression by western blot
analysis.
Western blot analysis
Western blot analysis was performed with a sodium
dodecyl sulfate-polyacrylamide gel electrophoresis system as
previously described (23).
Immunoblot analysis was performed by incubation with primary
antibodies at 25°C for 2 h (Table I).
The blots were then washed and incubated with a 1:2,000 dilution of
horseradish peroxidase (HRP)-conjugated secondary antibody
(Jackson, West Grove, PA, USA), followed by three washes with
Tris-buffered saline-Tween. An enhanced chemiluminescent HRP
substrate (Pierce, Rockford, IL, USA) was used for detection
according to the manufacturer's instructions.
 | Table I.Description of western blot
antibodies. |
Table I.
Description of western blot
antibodies.
| Antibody | Vendor | Clone | WB (dilution) |
|---|
| IRS2 | Abcam | Monoclonal | 1:3,000 |
| DDK | Origene | Monoclonal | 1:2,000 |
| E-cadherin | Upstate | Polyclonal | 1:3,000 |
| N-cadherin | Abcam | Polyclonal | 1:3,000 |
| Vimentin | Abcam | Monoclonal | 1:3,000 |
| Fibronectin | Abcam | Polyclonal | 1:3,000 |
| GAPDH | GeneTex | Polyclonal | 1:10,000 |
Cell migration and invasion
assays
Cell migration and invasion were assessed as
previously described (23). Briefly,
200 ml of cell suspension was added to the top wells of the chamber
with 8 µm pores coated with 0.1 ml of diluted Matrigel matrix
coating solution (Corning Inc., Corning, NY, USA) for the invasion
assay, or left uncoated for the migration assay. Average cell
mobility was determined by counting three random high-powered
fields at magnification, ×100. Three independent experiments were
performed for both invasion and migration assays.
Statistical analysis
All statistical analyses were performed using SPSS
for Windows 17.0 software (SPSS Inc., Chicago, IL, USA). The
significance of the association between IRS2 expression and
histopathological variables was determined by chi-square and
Fisher's exact tests. Comparisons between the two groups were
conducted using Student's t-test. Overall survival was calculated
from the date of diagnosis to death as a result of all causes. DFS
was computed from the time of surgery to recurrence in the liver or
distant metastasis. The Kaplan-Meier method was used for univariate
survival analysis, and the difference between survival curves was
tested by a log-rank test. For all analyses, two-sided tests of
significance were used with P<0.05 considered to indicate a
statistically significant difference.
Results
Correlation between IHC findings and
clinicopathological variables
The cohort consisted of 48 men and 38 women, with a
median age of 56.5 years (range, 30–84; mean, 56.8 years).
Immunoexpression of the IRS2 protein was observed in 86 cases
(Table II). IRS2 was overexpressed
in 29 cases (33.7%; Fig. 1) and was
significantly more frequent in cases with large tumor size
(P=0.033), classified as advanced stage by the American Joint
Committee on Cancer (P=0.046). In addition, IRS2-overexpressing
iCCA demonstrated a marginal difference in DFS. When IRS2 was
overexpressed in the tumors, DFS was shorter (mean 31.8 vs. 51.1
months; P=0.302).
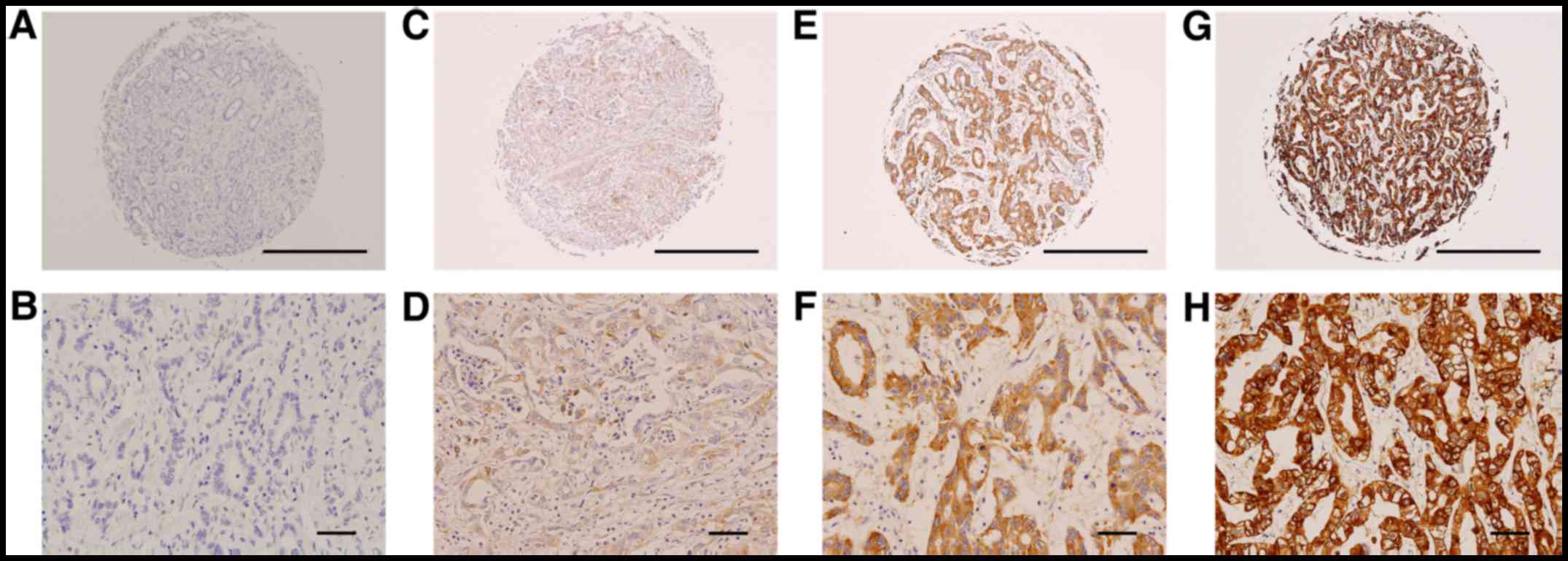 | Figure 1.Representative photographs of IRS2
immunostaining in iCCA. (A, C, E, and G) Tissue microarray cores at
magnification, ×40 (scale bar, 500 µm); (B, D, F, and H) selected
areas from (C, E, and G) at a higher magnification (×200; scale
bar, 50 µm). Expression index values were calculated by multiplying
the percentage of positive tumor cells by the average intensity. (A
and B) The negative control incubated with secondary antibody only.
(C and D) Weak staining (1+) with 15% positive tumor cells. (E and
F) Moderate staining (2+) with 85% positive tumor cells. (G and H)
Strong staining (3+) with 95% positive tumor cells. iCCA,
intrahepatic cholangiocarcinoma. |
| Table II.Clinicopathological characteristics
and associations with IRS2 immunoexpression in 86 intrahepatic
cholangiocarcinoma. |
Table II.
Clinicopathological characteristics
and associations with IRS2 immunoexpression in 86 intrahepatic
cholangiocarcinoma.
|
|
| IRS2
expression |
|
|---|
|
|
|
|
|
|---|
| Parameters | No. of
patients | Positive | Negative | P-value |
|---|
| Age, years |
|
|
| NS |
|
≤60 | 49 | 19 | 30 |
|
|
>60 | 37 | 10 | 27 |
|
| Sex |
|
|
| NS |
|
Male | 48 | 13 | 35 |
|
|
Female | 38 | 16 | 22 |
|
| Gross pattern |
|
|
| NS |
| MF | 52 | 14 | 38 |
|
|
MF+PI | 33 | 14 | 19 |
|
| Tumor N |
|
|
| NS |
|
Solitary | 69 | 23 | 46 |
|
|
Multiple | 16 | 6 | 10 |
|
| Tumor size, cm |
|
|
| 0.033a |
| 5 | 46 | 11 | 35 |
|
|
>5 | 40 | 18 | 22 |
|
| Necrosis |
|
|
| NS |
|
≤10% | 62 | 24 | 38 |
|
|
>10% | 24 | 5 | 19 |
|
| VI |
|
|
| NS |
| No | 52 | 18 | 34 |
|
|
Yes | 34 | 11 | 23 |
|
| NI |
|
|
| NS |
| No | 55 | 18 | 37 |
|
|
Yes | 31 | 11 | 20 |
|
| H grade |
|
|
| NS |
| I | 26 | 9 | 17 |
|
|
II+III | 60 | 20 | 40 |
|
| Stage |
|
|
| 0.046a |
|
I+II | 45 | 11 | 34 |
|
|
III+IV | 41 | 18 | 23 |
|
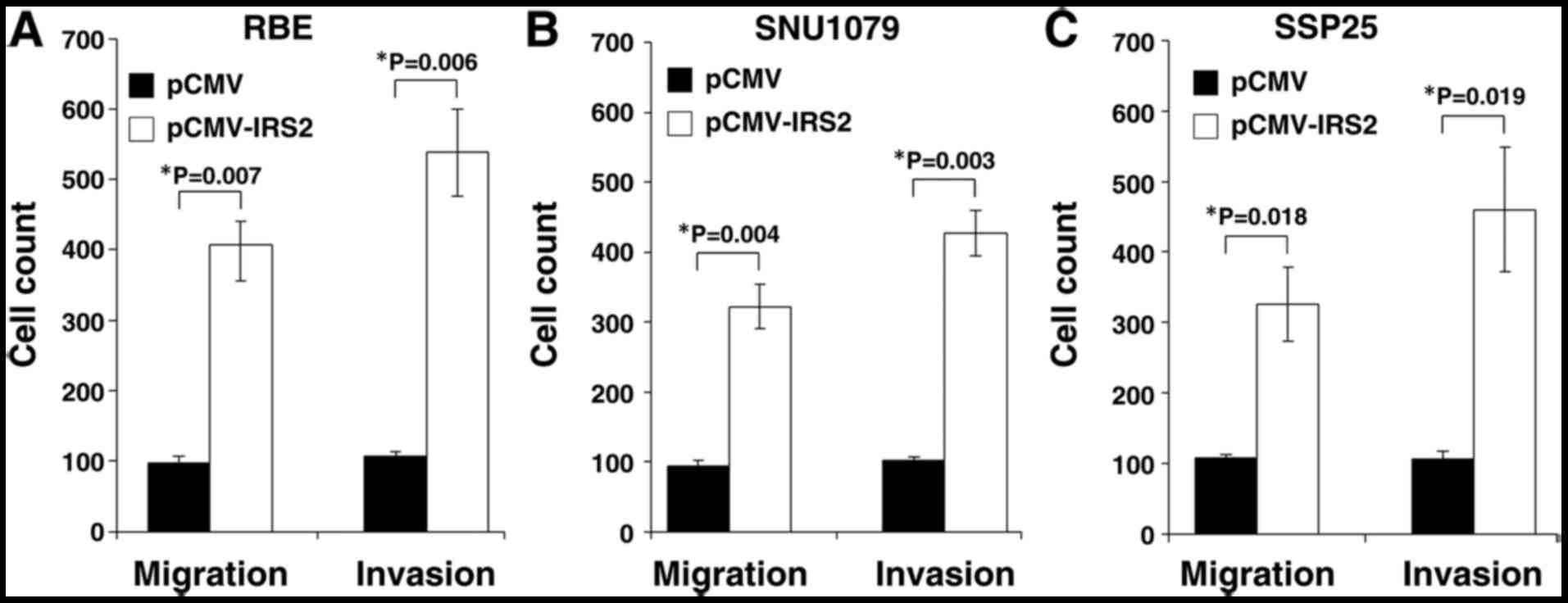
IRS2 overexpression affected the
migratory and invasive capacities of iCCA cells through EMT in
vitro
To assess the oncogenic activity of IRS2 in
cell lines, we established stable IRS2 expression in RBE, SNU1079,
and SSP25 cells (designated as RBE-IRS2, SNU1079-IRS2, and
SSP25-IRS2, respectively). IRS2-transfected cells were compared
with the corresponding control cells with empty vectors; the number
of migratory cells was significantly increased (RBE: P=0.007;
SNU1079: P=0.004; SSP25: P=0.018; Fig.
2). The penetration function of IRS2 during metastasis was
assessed. The cell invasion ability increased following ectopic
expression of IRS2 (RBE: P=0.006; SNU1079: P=0.003; SSP25: P=0.019;
Fig. 2). In comparison with control
cells, all the SSP25-IRS2 and SNU1079-IRS2 cells had higher numbers
of migratory and invasive cells.
Furthermore, IRS2-trasfected cells lost
intercellular cohesion and displayed a more spindle-like
fibroblastic appearance (Fig. 3A).
Expression analysis of EMT markers demonstrated decreased
epithelial marker levels (E-cadherin) and increased mesenchymal
marker levels (N-cadherin and fibronectin) in RBE-IRS2, SSP25-IRS2,
and SNU1079-IRS2 cells compared with their corresponding control
cells (Fig. 3B). The data suggest
that IRS2 overexpression could promote EMT despite the conflicting
results of vimentin expression in RBE-IRS2 and SNU1079-IRS2
cells.
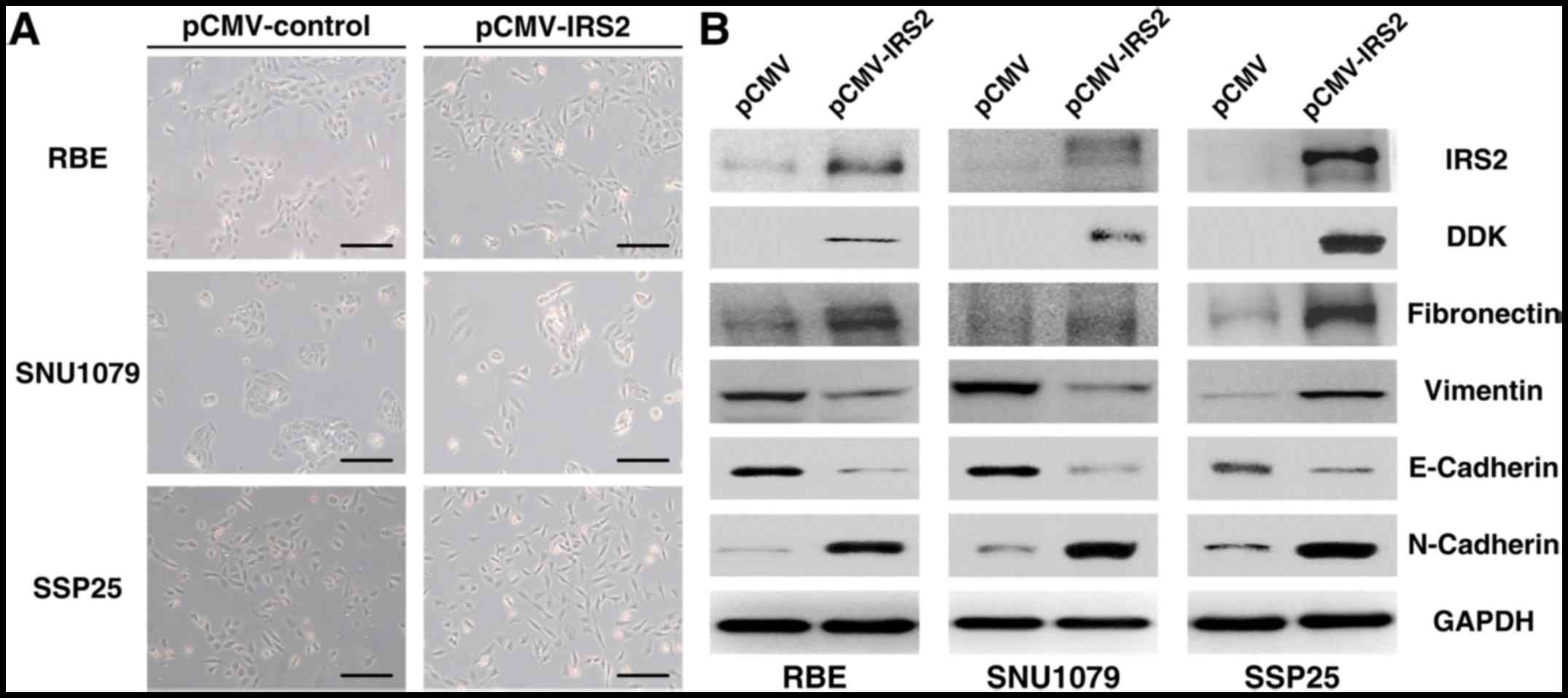 | Figure 3.Effect of pCMV-IRS2 transfection on
the EMT in RBE, SNU1079, and SSP25 cells. (A) After transfection
with pCMV-IRS2 vectors, morphological changes from a
cobblestone-like (left) to a spindle-like fibroblastic (right)
appearance (magnification, ×50; scale bar, 100 µm). (B) RBE,
SNU1079, and SSP25 cell extracts were subjected to 10% SDS-PAGE and
western blot analysis with the respective primary antibodies
against IRS2, vimentin, N-cadherin, fibronectin, and E-cadherin.
GAPDH was used as an internal control. EMT, epithelial-mesenchymal
transition; IRS2, insulin receptor substrate 2. |
Discussion
In this study, we investigated the
clinicopathological roles of IRS2 expression in 86 iCCA samples.
IRS2 overexpression was associated with large tumor size and
advanced tumor stage. Furthermore, the transfection of IRS2 in iCCA
cells promoted migration, invasion, and EMT in vitro. Taking
together all the findings, it can be surmised that IRS2 plays a
major role in regulating EMT in iCCA. The upregulation of IRS2 may
be a late event, promoting tumor progression and possibly
accounting for its more frequent presence in large-sized
tumors.
The putative role of IRS2 in tumor metastasis is
supported by other studies. Day et al demonstrated that an
increase in IRS2 expression is associated with disease progression
through the stages of colorectal carcinoma formation (15). A metastatic phenotype, conferring
tumor cells with the ability to invade and survive in foreign
environments, has been identified in IRS2-overexpressing mammary
tumors (24). Therefore, the
upregulation of IRS2 levels and activity may contribute to tumor
metastasis (24), as opposed to the
effects of IRS2 gene deletion (22). The relationship between IRS2
expression and liver cancer has not been explored in detail.
Boissan et al reported that high IRS2 levels can promote
tumor survival and progression in HCC (4). Consistent with other studies,
IRS2-overexpressing iCCA demonstrated increased migratory and
invasive capacities, which may lead to an advanced tumor stage and
adverse prognosis. Meanwhile, the knockdown of IRS2 has been
found to inhibit the mobility potential (23).
The present study revealed a function of IRS2 in
iCCA metastasis through EMT regulation. For migration, cancer cells
must first overcome programmed cell death and decrease
proliferation rates when the contact with the surrounding
extracellular matrix is lost. With the loss of the expression of
E-cadherin, a key marker of the epithelial phenotype, followed by
the expression of mesenchymal markers such as N-cadherin, vimentin,
and fibronectin, the invasive ability of cancer cells is increased.
Several transcription factors are upregulated and involved in EMT;
Snail, ZEB, and Twist induce epigenetic silencing at the E-cadherin
promoter, decreasing E-cadherin expression in the development of
the mesenchymal phenotype. Recently, large-scale evidence has
revealed that IGF-I/IGF-IR signaling is involved in EMT-associated
tumor metastasis and drug resistance (25–28). Our
study showed that iCCA cells with stable IRS2 expression exhibited
mesenchymal phenotypes characterized by the increased expression of
N-cadherin and fibronectin in all cell lines. Moreover, the
expression of E-cadherin, responsible for the epithelial phenotype,
was decreased. Thus, IRS2 overexpression is a factor involved in
the regulation of EMT in iCCA mainly through N-cadherin,
fibronectin, and E-cadherin.
Ligand activation of IGF-IR activates two main
signaling pathways, the IRS/PI3K/Akt and Ras/Raf/ERK pathways,
which regulate the transcription factors of EMT, including members
of ZEB, Snail, and Twist families. Crosstalk between other
signaling pathways and IGF signaling are also involved in the EMT
process, including the crosstalk and positive feedback loop between
IGF-I signaling and Wnt/β-catenin signaling and the activation of
Notch signaling (29). As for the
discrepancy of IRS2-induced change in vimentin expression between
different cell lines, we speculate that the overexpression of IRS2
in different cell lines might have direct and indirect impacts on
vimentin expression. IRS2 overexpression directly increased
vimentin expression in the SSP25 cell line through the IRS/PI3K/Akt
pathway, while IRS2 indirectly decreased vimentin through the
crosstalk of the EMT pathway. Further investigation to disclose the
genetic background related to the IRS/PI3K/Akt pathway and EMT
pathway in different cell lines will help elucidate the
discrepancy.
Although IRS2 is associated with metastatic
behaviors, it is not required for tumor initiation and growth. By
using the polyomavirus middle T antigen (PyV-MT) mouse model of
mammary tumorigenesis, Nagle et al found no differences in
mammary tumor onset or growth between Irs2-null and wild-type mice
(22). The ability of PyV-MT-derived
tumor cells to metastasize to the lungs was also significantly
impaired (22). Despite the close
structural homology of IRS1 with IRS2, IRS1 may regulate cancer
metastasis in a different manner. Ma et al demonstrated that
IRS1 expression was selectively inhibited in metastatic mammary
tumors (24). IRS1 may function as a
metastasis suppressor, given that IRS2 overexpression, as a
compensation for the loss of IRS1, promotes mammary tumor
metastasis (24). Similar malignant
tumor phenotypes affected by IRS1 and IRS2 have been found in other
tumor cells: HCC, neuroblastoma, mesothelioma, and prostate
carcinoma (4,30,31). Our
data are in line with previous findings and indicate that IRS2 is a
candidate prognostic marker for iCCA metastasis.
Considering that the insulin/IGF signaling pathways
are commonly known for fine-tuning numerous biological processes in
cancer, targeting these pathways is a promising strategy in cancer
therapy. Some therapeutic strategies include small-molecule
tyrosine kinase inhibitors, receptor blockage with neutralizing
antibodies, and IGFBs, which have been developed and evaluated in
preclinical and clinical phases (1).
IGF-1R and IR tyrosine kinase inhibitors can inhibit constitutive
or ligand-dependent phosphorylation, as well as the downstream
PI3K/AKT/mTOR and Erk MAP kinase pathways (32,33).
Neutralizing antibodies are monoclonal antibodies developed to
target the extracellular domain of IGF-1R and IR, subsequently
blocking the downstream signaling pathways (1). Both strategies result in the inhibition
of cell proliferation and stimulation of apoptosis in cancer cells
(1). Recently described antibodies
against IGF to neutralize the ligand have been found to be
effective in inhibiting the tumor metastasis of a variety of cancer
cells (34,35). A unique family of small molecules,
IRS-targeted agents that can lead to Ser phosphorylation and the
destruction of IRS1 and IRS2, results in the long-term inhibition
of IR/IGF-IR signaling and strong inhibition of tumor cell growth
(12). Therefore, targeting IGF-1R
and IR can be a promising strategy in cancer therapy.
In conclusion, our present results showed that iCCA
patients with IRS2 overexpression had large tumor size and advanced
tumor stage compared to patients without IRS2 overexpression.
According to in vitro studies, the stable IRS2-introdued
cells lines had a higher mobility potential, owing to the
association with EMT pathways. The regulatory mechanisms between
IRS2 expression and EMT pathways should be further investigated. We
propose that IRS2 is a valuable predictive indicator of metastasis
and may provide new directions for targeted therapy in iCCA.
Acknowledgements
Not applicable.
Funding
The study was supported by the Ministry of Science
and Technology, Taiwan (grant nos. MOST 104-2320-B-182A-010 and
MOST 105-2320-B-182A-016) and Chang Gung Memorial Hospital (grant
nos. CMRPG8C0591-2, CMRPG8E1471-2, CMRPG8B1251-3, CMRPG8D1321 and
CMRPG8E0921). The funders had no role in the manuscript design,
data collection and analysis, decision to publish, or preparation
of the manuscript.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Author's contributions
WTH conceived and designed the experiments. HLY, TTL
and CHC performed the experiments. SWW, YCW and HLE analyzed the
data. HLY and TTL wrote the manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Chang Gung
Medical Foundation Institutional Review Board (approval no. IRB
103-0818C, 103-4961B). Informed consent for participation was
waived by the International Review Board on the basis that all
samples and medical data used in the present study has been
irreversibly anonymized. Furthermore, it was a retrospective study
using archived material.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
IRS2
|
insulin receptor substrate 2
|
|
iCCA
|
intrahepatic cholangiocarcinoma
|
|
EMT
|
epithelial-mesenchymal transition
|
|
IGFs
|
insulin-like growth factors
|
|
IGFBs
|
IGF-binding proteins
|
|
DFS
|
disease-free survival
|
|
HCC
|
hepatocellular carcinoma
|
|
IHC
|
immunohistochemical
|
|
PyV-MT
|
polyomavirus middle T antigen
|
References
|
1
|
Singh P, Alex JM and Bast F: Insulin
receptor (IR) and insulin-like growth factor receptor 1 (IGF-1R)
signaling systems: Novel treatment strategies for cancer. Med
Oncol. 31:8052014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kasper JS and Giovannucci E: A
meta-analysis of diabetes mellitus and the risk of prostate cancer.
Cancer Epidemiol Biomarkers Prev. 15:2056–2062. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dearth RK, Cui X, Kim HJ, Hadsell DL and
Lee AV: Oncogenic transformation by the signaling adaptor proteins
insulin receptor substrate (IRS)-1 and IRS-2. Cell Cycle.
6:705–713. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boissan M, Beurel E, Wendum D, Rey C,
Lécluse Y, Housset C, Lacombe ML and Desbois-Mouthon C:
Overexpression of insulin receptor substrate-2 in human and murine
hepatocellular carcinoma. Am J Pathol. 167:869–877. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Esposito DL, Verginelli F, Toracchio S,
Mammarella S, De Lellis L, Vanni C, Russo A, Mariani-Costantini R
and Cama A: Novel insulin receptor substrate 1 and 2 variants in
breast and colorectal cancer. Oncol Rep. 30:1553–1560. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rozengurt E, Sinnett-Smith J and Kisfalvi
K: Crosstalk between insulin/insulin-like growth factor-1 receptors
and G protein-coupled receptor signaling systems: A novel target
for the antidiabetic drug metformin in pancreatic cancer. Clin
Cancer Res. 16:2505–2511. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hsing AW, Sakoda LC and Chua S Jr:
Obesity, metabolic syndrome, and prostate cancer. Am J Clin Nutr.
86:s843–s857. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dziadziuszko R, Camidge DR and Hirsch FR:
The insulin-like growth factor pathway in lung cancer. J Thorac
Oncol. 3:815–818. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jerome L, Alami N, Belanger S, Page V, Yu
Q, Paterson J, Shiry L, Pegram M and Leyland-Jones B: Recombinant
human insulin-like growth factor binding protein 3 inhibits growth
of human epidermal growth factor receptor-2-overexpressing breast
tumors and potentiates herceptin activity in vivo. Cancer Res.
66:7245–7252. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nahta R, Yu D, Hung MC, Hortobagyi GN and
Esteva FJ: Mechanisms of disease: Understanding resistance to
HER2-targeted therapy in human breast cancer. Nat Clin Pract Oncol.
3:269–280. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ouban A, Muraca P, Yeatman T and Coppola
D: Expression and distribution of insulin-like growth factor-1
receptor in human carcinomas. Hum Pathol. 34:803–808. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Reuveni H, Flashner-Abramson E, Steiner L,
Makedonski K, Song R, Shir A, Herlyn M, Bar-Eli M and Levitzki A:
Therapeutic destruction of insulin receptor substrates for cancer
treatment. Cancer Res. 73:4383–4394. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Taniguchi CM, Emanuelli B and Kahn CR:
Critical nodes in signalling pathways: Insights into insulin
action. Nat Rev Mol Cell Biol. 7:85–96. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cai D, Dhe-Paganon S, Melendez PA, Lee J
and Shoelson SE: Two new substrates in insulin signaling, IRS5/DOK4
and IRS6/DOK5. J Biol Chem. 278:25323–25330. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Day E, Poulogiannis G, McCaughan F,
Mulholland S, Arends MJ, Ibrahim AE and Dear PH: IRS2 is a
candidate driver oncogene on 13q34 in colorectal cancer. Int J Exp
Pathol. 94:203–211. 2013.PubMed/NCBI
|
|
16
|
Yuan TL and Cantley LC: PI3K pathway
alterations in cancer: Variations on a theme. Oncogene.
27:5497–5510. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rosner A, Miyoshi K, Landesman-Bollag E,
Xu X, Seldin DC, Moser AR, MacLeod CL, Shyamala G, Gillgrass AE and
Cardiff RD: Pathway pathology: Histological differences between
ErbB/Ras and Wnt pathway transgenic mammary tumors. Am J Pathol.
161:1087–1097. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dearth RK, Cui X, Kim HJ, Kuiatse I,
Lawrence NA, Zhang X, Divisova J, Britton OL, Mohsin S, Allred DC,
et al: Mammary tumorigenesis and metastasis caused by
overexpression of insulin receptor substrate 1 (IRS-1) or IRS-2.
Mol Cell Biol. 26:9302–9314. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Byron SA, Horwitz KB, Richer JK, Lange CA,
Zhang X and Yee D: Insulin receptor substrates mediate distinct
biological responses to insulin-like growth factor receptor
activation in breast cancer cells. Br J Cancer. 95:1220–1228. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gibson SL, Ma Z and Shaw LM: Divergent
roles for IRS-1 and IRS-2 in breast cancer metastasis. Cell Cycle.
6:631–637. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mardilovich K and Shaw LM: Hypoxia
regulates insulin receptor substrate-2 expression to promote breast
carcinoma cell survival and invasion. Cancer Res. 69:8894–8901.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nagle JA, Ma Z, Byrne MA, White MF and
Shaw LM: Involvement of insulin receptor substrate 2 in mammary
tumor metastasis. Mol Cell Biol. 24:9726–9735. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu TT, You HL, Weng SW, Wei YC, Eng HL
and Huang WT: Recurrent amplification at 13q34 targets at CUL4A,
IRS2, and TFDP1 As an independent adverse prognosticator in
intrahepatic cholangiocarcinoma. PLoS One. 10:e01453882015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ma Z, Gibson SL, Byrne MA, Zhang J, White
MF and Shaw LM: Suppression of insulin receptor substrate 1 (IRS-1)
promotes mammary tumor metastasis. Mol Cell Biol. 26:9338–9351.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao H, Desai V, Wang J, Epstein DM,
Miglarese M and Buck E: Epithelial-mesenchymal transition predicts
sensitivity to the dual IGF-1R/IR inhibitor OSI-906 in
hepatocellular carcinoma cell lines. Mol Cancer Ther. 11:503–513.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Denduluri SK, Idowu O, Wang Z, Liao Z, Yan
Z, Mohammed M, Ye J, Wei Q, Wang J, Zhao L and Luu HH: Insulin-like
growth factor (IGF) signaling in tumorigenesis and the development
of cancer drug resistance. Genes Dis. 2:13–25. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Motallebnezhad M, Aghebati-Maleki L,
Jadidi-Niaragh F, Nickho H, Samadi-Kafil H, Shamsasenjan K and
Yousefi M: The insulin-like growth factor-I receptor (IGF-IR) in
breast cancer: Biology and treatment strategies. Tumour Biol.
37:11711–11721. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang R, Li H, Guo X, Wang Z, Liang S and
Dang C: IGF-I induces epithelial-to-mesenchymal transition via the
IGF-IR-Src-MicroRNA-30a-E-cadherin pathway in nasopharyngeal
carcinoma cells. Oncol Res. 24:225–231. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li H, Batth IS, Qu X, Xu L, Song N, Wang R
and Liu Y: IGF-IR signaling in epithelial to mesenchymal transition
and targeting IGF-IR therapy: Overview and new insights. Mol
Cancer. 16:62017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hoang CD, Zhang X, Scott PD, Guillaume TJ,
Maddaus MA, Yee D and Kratzke RA: Selective activation of insulin
receptor substrate-1 and −2 in pleural mesothelioma cells:
Association with distinct malignant phenotypes. Cancer Res.
64:7479–7485. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim B, van Golen CM and Feldman EL:
Insulin-like growth factor-I signaling in human neuroblastoma
cells. Oncogene. 23:130–141. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wittman M, Carboni J, Attar R,
Balasubramanian B, Balimane P, Brassil P, Beaulieu F, Chang C,
Clarke W, Dell J, et al: Discovery of a
(1H-benzoimidazol-2-yl)-1H-pyridin-2-one (BMS-536924) inhibitor of
insulin-like growth factor I receptor kinase with in vivo antitumor
activity. J Med Chem. 48:5639–5643. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pitts TM, Tan AC, Kulikowski GN, Tentler
JJ, Brown AM, Flanigan SA, Leong S, Coldren CD, Hirsch FR,
Varella-Garcia M, et al: Development of an integrated genomic
classifier for a novel agent in colorectal cancer: Approach to
individualized therapy in early development. Clin Cancer Res.
16:3193–3204. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Miyamoto S, Nakamura M, Shitara K,
Nakamura K, Ohki Y, Ishii G, Goya M, Kodama K, Sangai T, Maeda H,
et al: Blockade of paracrine supply of insulin-like growth factors
using neutralizing antibodies suppresses the liver metastasis of
human colorectal cancers. Clin Cancer Res. 11:3494–3502. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Goya M, Miyamoto S, Nagai K, Ohki Y,
Nakamura K, Shitara K, Maeda H, Sangai T, Kodama K, Endoh Y, et al:
Growth inhibition of human prostate cancer cells in human adult
bone implanted into nonobese diabetic/severe combined
immunodeficient mice by a ligand-specific antibody to human
insulin-like growth factors. Cancer Res. 64:6252–6258. 2004.
View Article : Google Scholar : PubMed/NCBI
|