Introduction
Melanoma, one of the most aggressive cancer types,
accounts for <5% of all skin cancer types but is responsible for
the majority of incidences of skin cancer-associated mortality
globally (1). It has been
hypothesized that melanoma stem cells (MSCs) possess the capacity
for self-renewal, differentiation, chemoresistance and immune
evasion (2,3). Thus, understanding the mechanism
underlying melanoma progression may contribute to the targeted
killing of MSCs, which may improve the efficacy of anticancer
therapies and decrease the risk of relapse and progression.
Cytotoxic T lymphocyte antigen-4 (CTLA-4) is a
co-stimulatory molecule that is expressed on activated T cells and
competes with T-cell-specific surface glycoprotein CD28 for binding
to B7 and inhibits T-cell proliferation, activation and immune
response (4,5). CTLA-4 is also expressed on regulatory T
cells with immunosuppressive properties (6,7). Studies
have demonstrated that the human CTLA-4 blocking antibody,
ipilimumab, which was approved by the United States Food and Drug
Administration for the monotherapy of advanced melanoma in March
2011, has a response rate of 11% and prolongs overall survival in
22% of patients (8,9). As a subset of patients treated with
CTLA-4 blockade, ipilimumab, exhibited an objective clinical
response, it may be advantageous to investigate the molecular
mechanisms underlying CTLA-4 anti-tumour activity and identify
patients who are most likely to respond.
CTLA-4 has also been implicated to serve functions
in neoplastic cells, including B cell chronic lymphocytic leukaemia
(10), non-Hodgkin's lymphoma
(11), breast cancer (12), lung cancer (13), melanoma (14), gastric cancer (15), colorectal cancer (15), cervical cancer (16) and oesophageal carcinoma (17). Prior studies indicated that CTLA-4
expression in tumour cells is associated with poor prognosis
(12,17). CTLA-4 engagement with B7 ligands
induces tumour cell death through apoptosis (18), and ipilimumab triggers effector
lymphocytes and tumour necrosis factor release to directly
eliminate CTLA-4+ melanomas (14). These studies have indicated the
potential function of CTLA-4 in tumour cells. Furthermore, CTLA-4
was overexpressed in allogeneic mesenchymal stem cells, which
resulted in the suppression of the immune response and the
promotion of osteogenic differentiation (19). It has been indicated that CTLA-4 is
involved in regulating the biological characteristics of stem cells
(19). Extensive clinical evidence
supports the use of CTLA-4 blockade against advanced melanoma
(8,9).
Nevertheless, the involvement of the CTLA-4 molecule in regulating
the tumourigenic capacity of MSCs is not well understood.
In the present study, the expression of CTLA-4 in
MSCs in melanoma cell lines were examined and the function of
CTLA-4 in maintaining the characteristics of MSCs were
investigated. The results demonstrated that the CTLA-4 molecule
participated in the tumourspheres proliferation in vitro and
apoptosis of melanoma cells. Furthermore, CTLA-4 was expressed in
MSCs and was involved in enhancing the aldehyde dehydrogenase
(ALDH) activity and tumourigenic capacity of MSCs.
Materials and methods
Cell culture
The mouse melanoma B16-F0 and B16-F1 cell lines
(American Type Culture Collection, Manassas, VA, USA) were
maintained in RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 10% foetal bovine serum (ScienCell
Research Laboratories, Inc., San Diego, USA), 100 U/ml penicillin
(Gibco; Thermo Fisher Scientific, Inc.) and 100 µg/ml streptomycin
(Gibco; Thermo Fisher Scientific, Inc.). Cells were cultured at
37°C, 95% humidity and 5% CO2.
Flow cytometry
To determine the expression of CTLA-4 in melanoma
cells, B16-F0 and B16-F1 cells were surface stained for CTLA-4. A
total of 1×106 cells/ml B16-F0 or B16-F1 cells were suspended in
PBS at room temperature. To one tube of cells, 5 µl anti-CTLA-4
antibody (cat. no. 553720; dilution 1:200PE; BD Pharmingen; BD
Biosciences) was added, and to one tube of cells 5 µl
immunoglobulin G isotype-matched control (BD Pharmingen; BD
Biosciences) was added as a negative control at room temperature.
The tubes were incubated for 30 min at 4°C. Following incubation,
centrifugation was performed at 4°C for 10 min at 12,000 × g and
the pellets were re-suspended with 500 µl of Assay Buffer prior to
data acquisition. Samples were analysed using a FACSCalibur flow
cytometer (BD Biosciences, Franklin Lakes, NJ, USA). To investigate
the expression of CTLA-4 in MSCs, the ALDEFLUOR kit (Stemcell
Technologies, Inc., Vancouver, BC, Canada) was used. The ALDEFLUOR™
reagent used the enzyme bodipy-aminoacetaldehyde (BAAA) as
fluorescent substrate for ALDH, which freely diffused into intact
and viable cells. BAAA was converted into a polar fluorescent
product (BODIPY™-aminoacate) by ALDH and was retained inside the
cells. Dead cells were excluded based on light scatter
characteristics. A total of 1×106/ml cells were resuspended in an
Assay Buffer (Stemcell Technologies, Inc., Vancouver, BC, Canada)
at room temperature. A tube of cells was immediately quenched with
5 µl specific inhibitor of the enzyme ALDH, with
diethylaminobenzaldehyde (DEAB) as the negative control at room
temperature. To all tubes, 5 µl ALDEFLUOR™ reagent was added and
incubated for 45 min at 37°C. In a number of experiments, cells
were labelled with CTLA-4 subsequent to being incubated with
ALDEFLUOR. Data analysis was conducted using Cell Quest Pro
(version 5.1; BD Biosciences).
Apoptosis detection
Viable and dead cells of B16-F0 and B16-F1 with or
without anti-CTLA-4 antibody were assessed using double staining
with fluorescein isothiocyanate (FITC)-annexin V and propidium
iodide (BD Pharmingen; BD Biosciences) for 30 min at room
temperature, following the manufacturer's protocol. Analysis was
performed using flow cytometry, and the apoptotic percentages of
annexin V+/propidium iodide- and annexin V+/propidium iodide+ cells
were calculated.
Tumoursphere culture
In tumoursphere culture, 1×106 cells of
B16-F0 or B16-F1 were plated as single cells in ultralow attachment
six-well plates (Corning, Lowell, MA, USA) without anti-CTLA-4
following the protocol of Duarte et al (20). Cells were briefly cultured for 24 h at
37°C in RPMI 1640 containing 6 mg/ml glucose (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany), 1 mg/ml NaHCO3
(Sigma-Aldrich; Merck KGaA), 5 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (Sigma-Aldrich;
Merck KGaA), 4 µg/ml heparin (Sigma-Aldrich; Merck KGaA), 4 mg/ml
bovine serum albumin (Sigma-Aldrich; Merck KGaA), 20 pg/ml insulin
(Sigma-Aldrich; Merck KGaA), and N2 supplement (Invitrogen; Thermo
Fisher Scientific, Inc.) in addition to 10 ng/ml basic fibroblast
growth factor (PeproTech, Inc., Rocky Hill, NJ, USA) and 20 ng/ml
epidermal growth factor (PeproTech, Inc.). The second day following
seeding, cells were treated with 10 µg anti-CTLA-4 antibody (cat.
no. 16-1521; 1:100; eBioscience; Thermo Fisher Scientific, Inc.)
for 14 days at 37°C. Tumourspheres were observed under a optical
microscope (magnification, ×40) 14 days later. Individual spheres
>100 µm from each replicate well were counted under an inverted
microscope.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
B16-F0 and B16-F1 cells were cultured with or
without anti-CTLA-4 antibody in RPMI-1640 for 48 h at 37°C. RNAiso
Plus (1 ml; Takara Bio, Inc., Otsu, Japan) was added to all the
cultured B16-F0 and B16-F1 cells for total RNA extraction according
to the manufacturer's protocol. cDNA was synthesized using the
PrimeScriptRT Master Mix (Takara Bio, Inc.) following the
manufacturer's protocol. The temperature protocol of reverse
transcription was as follows: 37°C for 15 min, 85°C for 5 sec and
4°C for 10 min. A total of 20 ng of resulting cDNA were subjected
to RT-qPCR, in a 10 µl final reaction volume and analysed in
triplicates. The cycling conditions were as follows: 95°C for 30
sec, 95°C for 5 sec 40 times and 60°C for 34 sec 40 times. Gene
expression was detected using the ABI 7900 sequence detection
system (Thermo Fisher Scientific, Inc.). The gene expression level
of each target gene was normalized using the endogenous gene GAPDH
and quantified using the real-time quantitative PCR and
2−ΔΔCq method (21).
Relative value of sample=2-(ΔCqinternal parameter-ΔCqsample). PCR
primers were as follows: ALDH1A1 forward,
5′-GGCACTCAATGGTGGGAAAGTC-3′ and reverse,
5′-ATGCCAGGTGAAGAGCCGTG-3′; ALDH1A3 forward,
CCACGGTCTTCTCAGATGTTACG-3′ and reverse,
5′-GCTGTGAGTCCATAGTCGGTGC-3′; ALDH2 forward,
5′-TTAACAATGAGTGGCACGACG-3′ and reverse, 5′-CGCCAATCGGTACAACAGC-3′;
and GAPDH forward, 5′-AGGAGCGAGACCCCACTAACA-3′ and reverse,
5′-AGGGGGGCTAAGCAGTTGGT-3′. Baseline and threshold for the
2−ΔΔCq calculation were set automatically using Stepone
plus 2.3 software. (Thermo Fisher Scientific, Inc., Waltham, MA,
USA).
Animals and tumour model
For the in vivo experiments, 20 adult
specific-pathogen-free C57BL/6 female mice (22±4 g) were provided
by the Experimental Animal Center of Tongji Medical College of
Huazhong University of Science and Technology. All mice were housed
in the same facilities (temperature and humidity), consumed the
same diet (Lab Diet) and water ad libitum, and were kept on an
identical 12-h light/dark cycle. All animals were randomly assigned
to two groups (n=10). The C57 mice were injected with
1×106 B16-F0 (F0 group) or B16-F1 (F1 group) tumour
cells subcutaneously, and the tumour size was monitored with
callipers every other day and sacrificed at the 28th day. Fourteen
days after tumour cells injection, F0 or F1 group was injected
intraperitoneally with anti-CTLA-4 antibody or PBS three times
every other day. All surgical procedures and the handling of
animals were performed in accordance with institutional guidelines.
All procedures were approved by the Institutional Animal Care and
Use Committee at Tongii Medical College of Huazhong University of
Science and Technology (Hubei, China).
Statistical analysis
Statistical analyses were performed using Graph Pad
Prism 5.0 software (GraphPad Software, Inc., La Jolla, CA, USA).
All data were expressed as the mean ± the standard error of the
mean (SEM). Statistical analysis was performed using a two-tailed
paired Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Melanoma cell-intrinsic CTLA-4
regulates cell apoptosis
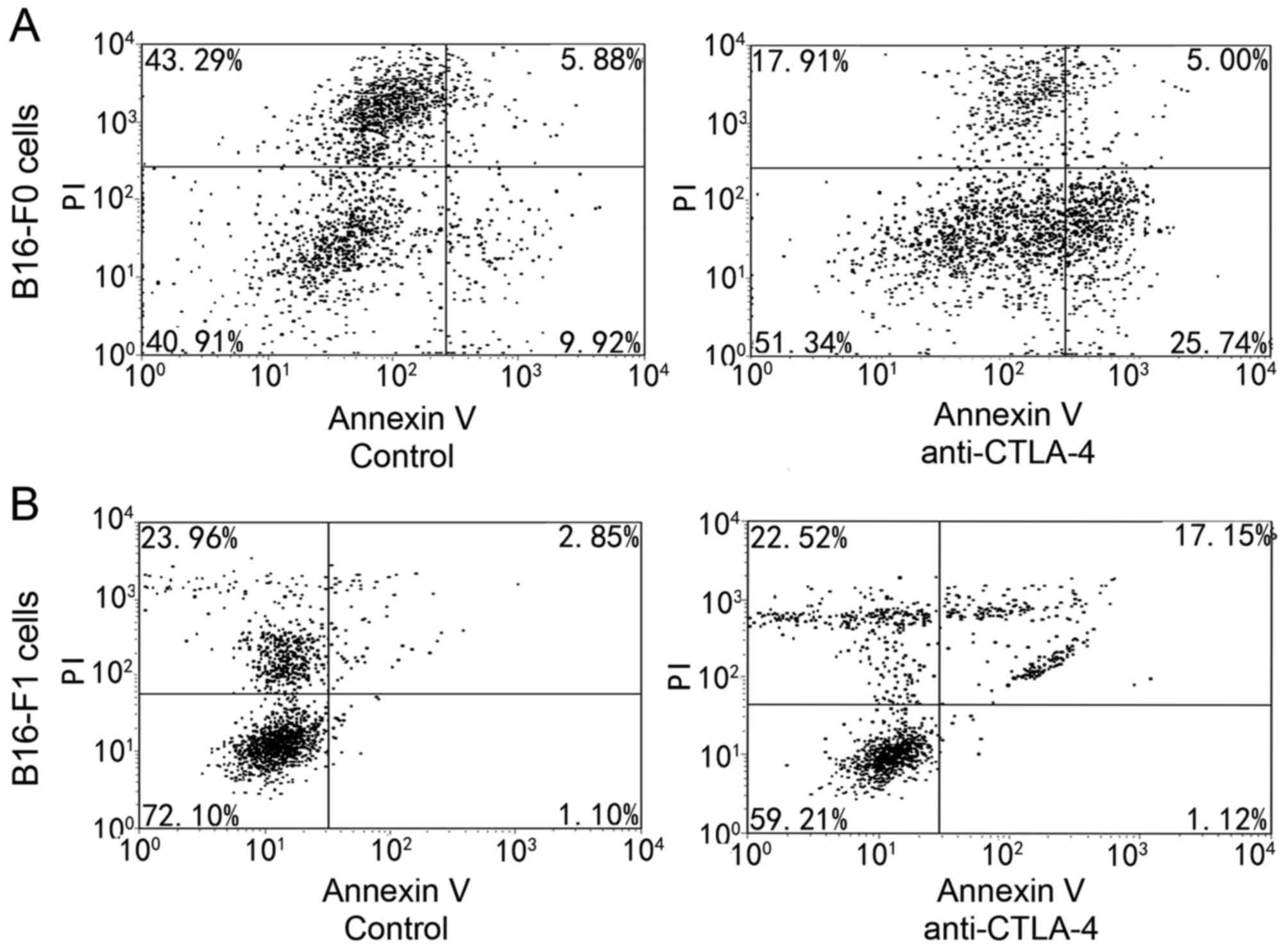
Following a 48-h incubation with anti-CTLA-4, cells
were harvested and annexin V/PI staining was assessed using flow
cytometry. As presented in Fig. 1,
exposure to 10,000 ng/ml anti-CLTA-4 was associated with marked
apoptosis induction. Cells that were in early apoptosis were
FITC-annexin V-positive and PI-negative, and cells that were in
late apoptosis were FITC-annexin V-positive and PI-positive. The
rate of apoptosis meant the ratio of cells in early and late
apoptosis to the total cell number. The rate of apoptosis increased
by 14.68±4.11% in B16-F0 cells and to 15.92±2.42% in B16-F1 cells
compared with the control.
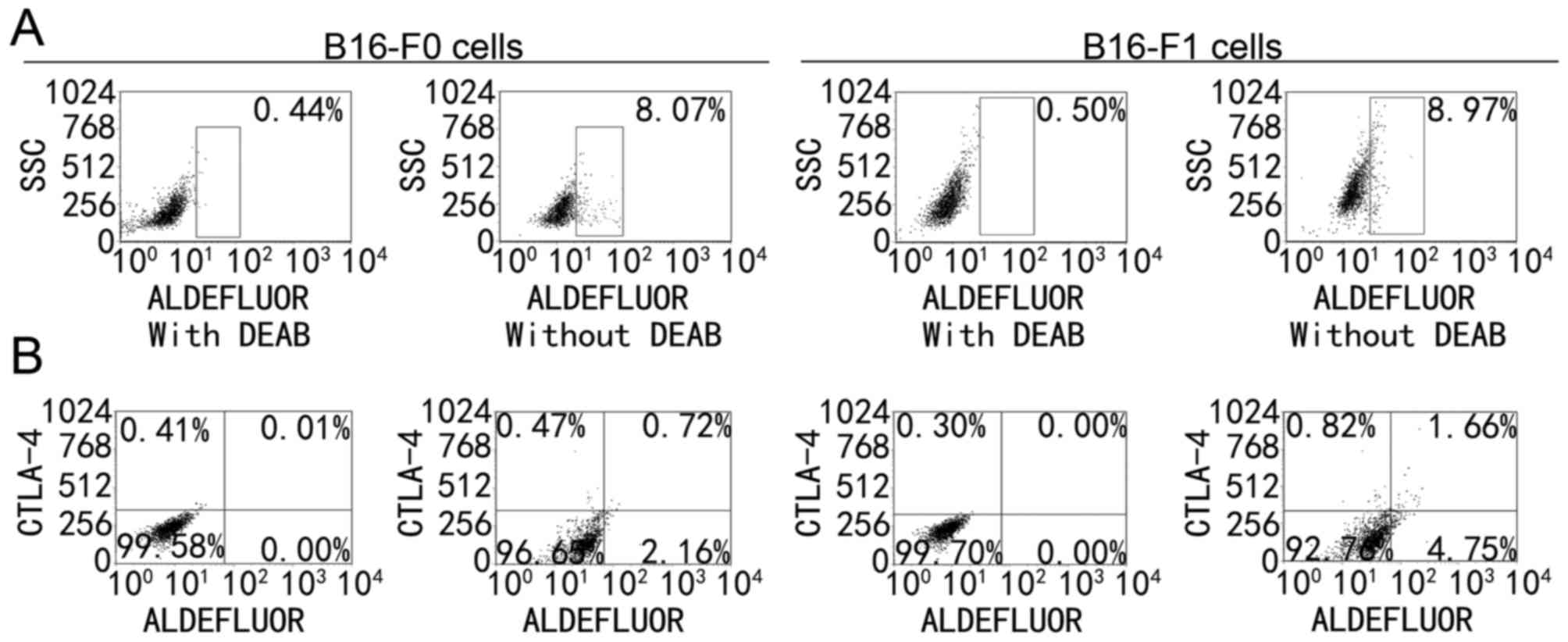
Expression of CTLA-4 in MSCs. On the basis of the
data from melanoma cells, the prevalence of CTLA-4 expression in
MSCs was examined. One previous study reported that 5–10% of B16-F0
and B16-F1 cells belonged to MSCs (22). ALDH+ cells were ALDEFLUOR
positive and ALDH+CTLA-4+ cells were
ALDEFLUOR positive and CTLA-4 positive. In the present study,
9.02±1.24% of the B16-F0 melanoma cells and 9.13±0.81% of B16-F1
melanoma cells were ALDH+ cells (Fig. 2A) and 2–5% of the cells were
ALDH+CTLA-4+ cells (Fig. 2B). Of the ALDH+ cells,
30–45% expressed CTLA-4, indicating that CTLA-4 was mainly
expressed in MSCs in melanoma cells.
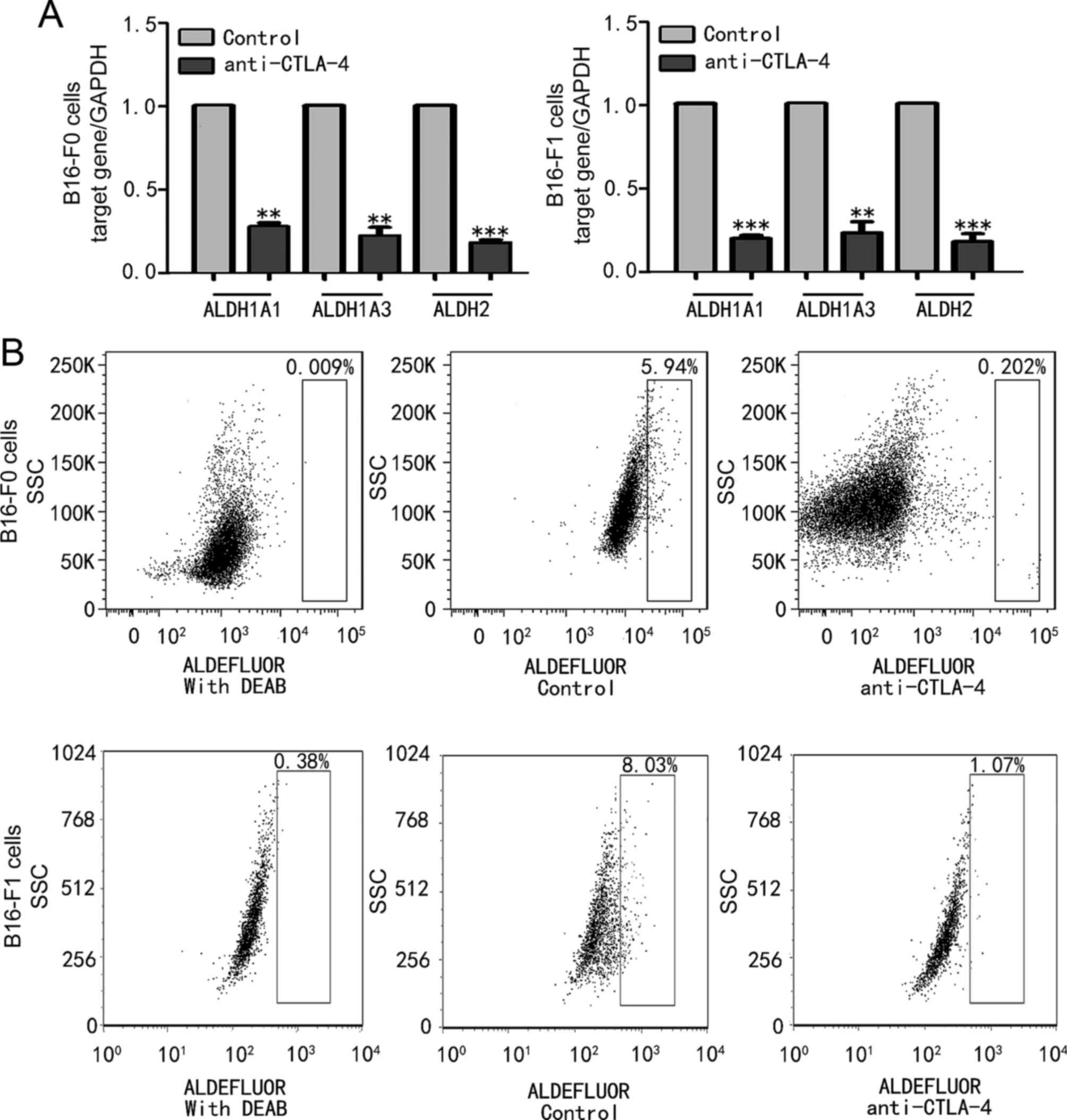
CTLA-4 regulated the activity of ALDH
in melanoma cells
To investigate the biological activity of CTLA-4 on
MSCs, the effect of CTLA-4 suppression on the expression of
ALDH1A1, ALDH1A3 and ALDH2 in B16-F0 and B16-F1 melanoma cells was
examined. Fig. 3A reveals that
anti-CTLA-4 significantly downregulated ALDH1A1 (P<0.01),
ALDH1A3 (P<0.01) and ALDH2 (P<0.001) mRNA expression levels
compared with their respective controls in B16-F0 cells, and
significantly downregulated ALDH1A1 (P<0.001), ALDH1A3
(P<0.01) and ALDH2 (P<0.001) mRNA expression levels compared
with their respective controls in B16-F1 cells. The expression
levels of ALDH1A1, ALDH1A3 and ALDH2 mRNA were inhibited by ~80% in
B16-F0 and B16-F1 melanoma cells (Fig.
3A). These results were confirmed by flow cytometry. Once the
cells were treated with anti-CTLA-4, the expression of ALDH was
decreased to 0.202% (Fig. 3B), and
the inhibition rate of ALDH expression in B16-F0 and B16-F1 cells
was 81.12±5.842 and 79.35±4.438%, respectively. These results
indicate that CTLA-4 is involved in the regulation of ALDH activity
and affects the biological behaviour of MSCs.
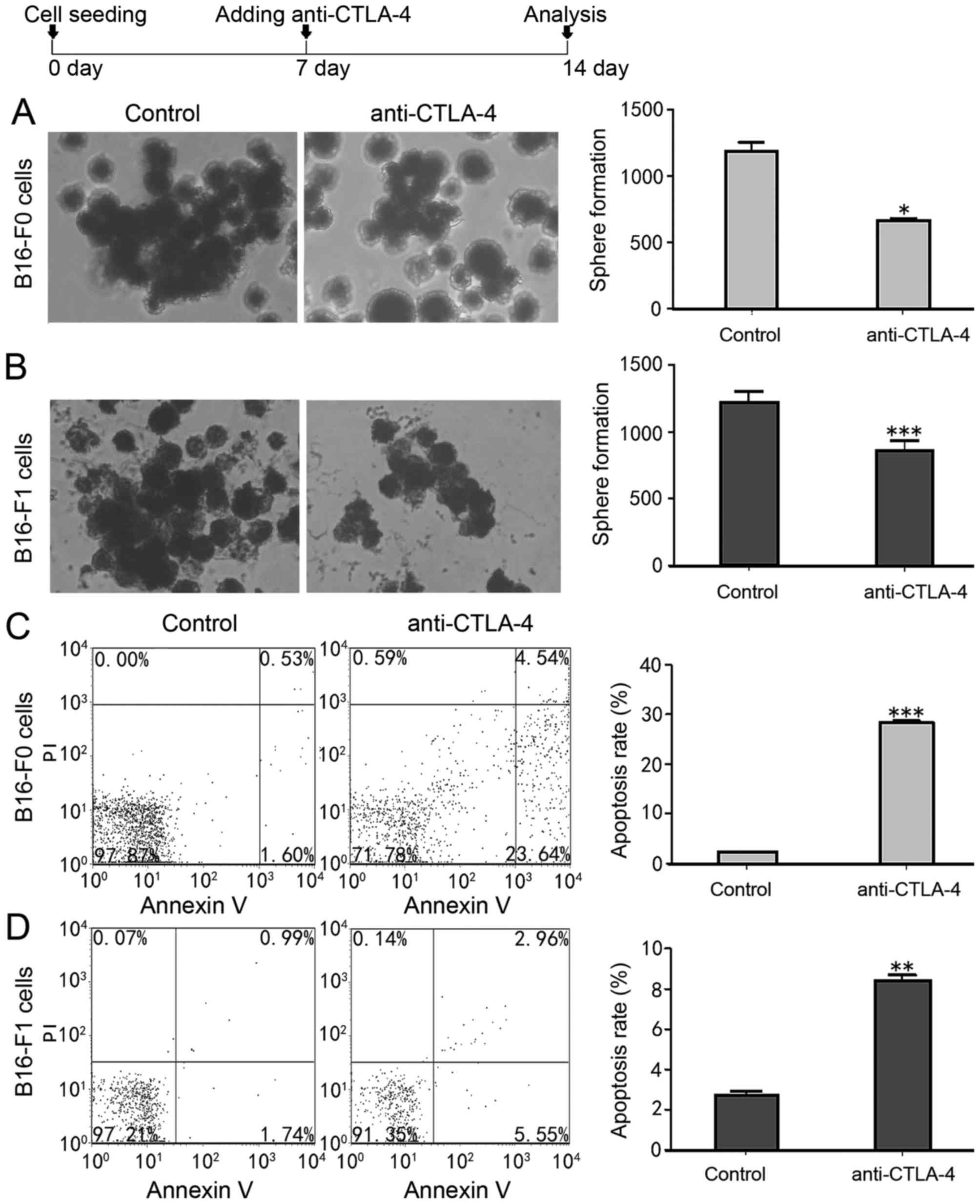
CTLA-4 affected sphere formation by
melanoma cells
To assess whether CTLA-4 expression affects the
self-renewal of MSCs, melanoma cell lines were cultured with
anti-CTLA-4. It was revealed that anti-CTLA-4 significantly reduced
the tumoursphere-forming efficiency of B16-F0 and B16-F1 melanoma
cells compared with the control groups (Fig. 4A and B). Cancer stem cells may be
serially passaged to generate secondary spheres with a cellular
composition resembling that of the primary sphere (23). The secondary tumourspheres generated
from the anti-CTLA-4 cultured melanoma cell-derived tumourspheres
were significantly decreased, by ~2-fold in B16-F0 (P<0.05) and
~1.4-fold in B16-F1 (P<0.001) melanoma cells in number and size,
compared with the control groups (Fig. 4A
and B). The effect of anti-CTLA-4 on melanoma cancer stem-like
cell apoptosis was detected. The results revealed that anti-CTLA-4
significantly increased the rate of apoptosis in B16-F0
(P<0.001) and B16-F1 (P<0.01) cells, and resulted in a
4–15-fold increase in the rate of apoptosis in melanoma cancer
stem-like cells (from 28.32±0.43 to 2.19±0.83% in B16-F0 cells and
from 8.42±0.27 to 8.42±0.27% in B16-F1 cells; Fig. 4C and D). Therefore, these results
indicate that CTLA-4 promoted the tumoursphere forming capacity of
MSCs and inhibited their apoptosis.
CTLA-4 affected the tumourigenic
capacity of melanoma cells
To confirm the function of CTLA-4 in the
self-renewal capacity of MSCs, whether CTLA-4 affected the
tumourigenic capacity of melanoma cells in vivo was
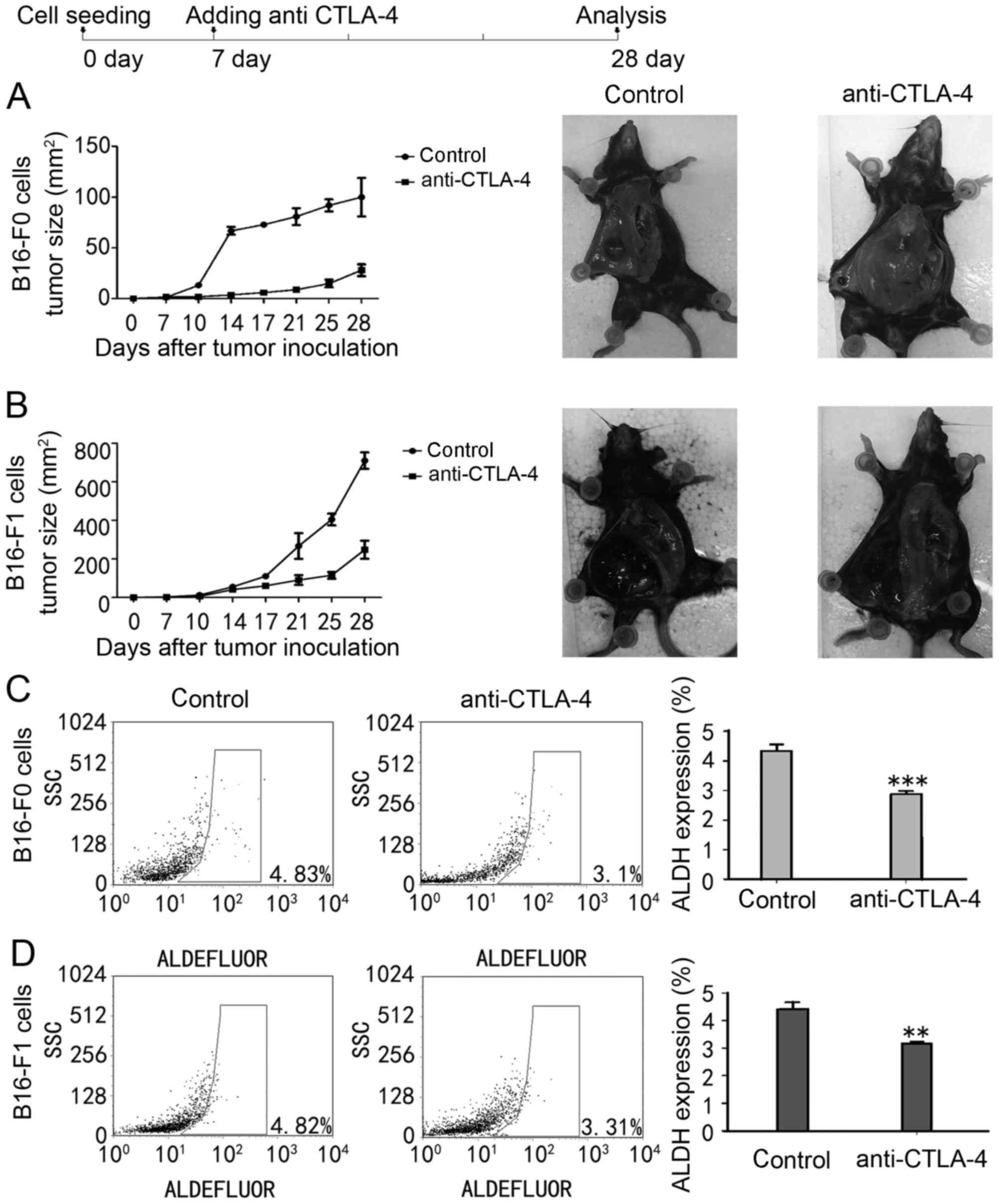
evaluated (Fig. 5A and B). Compared
with PBS-injected mice, anti-CTLA-4 suppressed tumour growth in two
mice models, which is supported by previous observations (24). In addition, residual ALDH+
MSCs were observed within the tumour. As presented in Fig. 5C and D, anti-CTLA-4 significant
decreased the residual ALDH+ MSCs in the two mice models
compared with PBS-injected animals (from 4.35±0.22 to 2.85±0.13% in
B16-F0-injected mice models, P<0.001; from 4.41±0.25 to
3.15±0.28% in B16-F1-injected mice models, P<0.01). These
results depict a potential mechanism for the
anti-tumour effects of anti-CTLA-4 associated with its ability
to suppress the tumourigenic capacity of MSCs.
Discussion
Extensive clinical evidence supports that CTLA-4
targeting agents demonstrate a notable benefit in patients with
advanced melanoma due to the function of CTLA-4 in negatively
regulating T cell activation (8,9). In
addition to T cells, CTLA-4 is expressed on cell lines derived from
a variety of human malignant solid tumour types, including
melanoma, neuroblastoma, oesophageal carcinoma, lung cancer,
gastric and colorectal cancer types (13,15,17,18).
CTLA-4 was revealed to serve a function in the protection against
apoptosis (18). These findings are
consistent with the results from a study by Contardi et al
(18) regarding the association
between CTLA-4 expression in tumour cells and tumour cell
apoptosis. These studies indicate the function of the CTLA-4
molecule in tumour cells and the notable function of CTLA-4 in stem
cells. Dai et al (19)
reported that CTLA-4 was over expressed in allogeneic mesenchymal
stem cells and promoted osteogenic differentiation. Shinoyama et
al (25) revealed that CTLA-4
promoted neuronal differentiation in the grafts of embryonic stem
cell-derived neural precursor cells. These findings confirmed that
CTLA-4 regulated the differentiation of stem cells.
Studies have demonstrated that MSCs drive the
initiation and progression of a melanoma based on the capacity for
self-renewal, multiple differentiation, chemoresistance and immune
evasion (2,3). Thus, targeting the survival mechanisms
utilized by MSCs may be the most effective way to cure melanoma.
Previous studies have successfully used ALDEFLUOR/ALDH as a marker
to isolate MSCs from mice (22,26).
However, the mechanisms underlying the biological effects in
ALDH+ MSCs remain unclear. In the present study, it was
revealed that CTLA-4 was expressed in ALDH+ MSCs, in
which it regulated the activity of ALDH in melanoma cells. These
results indicated that CTLA-4 induced melanoma progression and may
contribute to its function in regulating the stemness of MSCs.
To elucidate the function of CTLA-4 signalling
pathways in ALDH+ MSCs further, the effect of
anti-CTLA-4 on tumourigenic capacity was examined. The results of
the present study demonstrated, to the best of our knowledge for
the first time, that anti-CTLA-4 may efficiently inhibit the
capacity for tumoursphere formation and induce apoptosis in
melanoma cancer stem-like cells in vitro. In vivo,
the results demonstrated that treatment with anti-CTLA-4 suppressed
tumour growth and significantly decreased residual ALDH+
MSCs. These results indicate that the blockage of CTLA-4 may target
ALDH+ MSCs, which is a novel non-immunological mechanism
for the anti-tumour activity of anti-CTLA-4. Ipilimumab (an
anti-CTLA-4 monoclonal antibody) confers a long-term survival
benefit in ~20% of patients with advanced melanoma in an unselected
population (27), and identifying
those patients may prove to be valuable. One previous study
revealed that immune activation in responding patients increased
the absolute lymphocyte count and upregulated the T-cell activation
marker inducible T-cell co-stimulator and the tumour antigen New
York oesophageal squamous cell carcinoma 1 (28). The tumour mutation volume and high
baseline expression levels of immune-associated genes, including
C-C motif chemokine receptor 2 (CCR2), CCR-like 2 and CCR5,
additionally aid prediction of the activity of immune checkpoint
blockade (29). In fact, no
predictive biomarkers for the selection of patients with melanoma
able to receive ipilimumab are sufficiently robust enough for
clinical use. However, previous studies have illustrated that, in
breast and oesophageal cancer, CTLA-4 expression on tumour cells
was associated with a poor prognosis (6,12).
According to the biological characteristics of MSCs, the expression
of CTLA-4 on tumour cells, indicating poor prognosis, may be
associated with the regulation of CTLA-4 on cancer stem cells. The
present study has revealed that the regulation of CTLA-4 on MSCs
may serve as a predictive biomarker for selecting patients with
melanoma to receive anti-CTLA-4 treatment.
In conclusion, the present findings illustrated that
melanoma cell-intrinsic CTLA-4 induced tumourigenesis and inhibited
apoptosis, in addition to its pro-tumourigenic function.
Identifying melanoma-CTLA-4-driven tumourigenesis critically
improves the current understanding of the mechanism underlying
melanoma progression and may benefit CTLA-4-targeted therapy by
improving the long-term outcomes in patients with advanced-stage
melanoma.
Acknowledgements
The authors would like to thank the Center for Stem
Cell Research and Application for providing flow analysis and Dr
Wen Lin (Department of Pediatrics, Affiliated Union Hospital,
Wuhan, China) for their insights.
Funding
The present study was supported by the National
Nature Sciences Foundation of China (grant no. 81301954) and the
Hubei Provincial Health and Family Planning Scientific Research
Project (grant no. 2015060101010043).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BZ performed the flow analysis and analysed all
data, and was a major contributor in writing the manuscript. FZ
helped to design the stufy, performed the flow analysis and was a
important contributor in writing the manuscript. JD cultured the
cells. DB made the animal model. CW performed the western blot. JH
performed the RT-qPCR. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Animal Care and Use Committee at Tongii Medical College of Huazhong
University of Science and Technology (Hubei, China).
Patient consent for approval
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MSCs
|
melanoma stem cells
|
|
CTLA-4
|
cytotoxic T lymphocyte antigen-4
|
|
ALDH
|
aldehyde dehydrogenase
|
References
|
1
|
Gray-Schopfer V, Wellbrock C and Marais R:
Melanoma biology and new targeted therapy. Nature. 445:851–857.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Murphy GF, Wilson BJ, Girouard SD, Frank
NY and Frank MH: Stem cells and targeted approaches to melanoma
cure. Mol Aspects Med. 39:33–49. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hirohashi Y, Torigoe T, Tsukahara T,
Kanaseki T, Kochin V and Sato N: Immune responses to human cancer
stem-like cells/cancer-initiating cells. Cancer Sci. 107:12–17.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brunner MC, Chambers CA, Chan FK, Hanke J,
Winoto A and Allison JP: CTLA-4-mediated inhibition of early events
of T cell proliferation. J Immunol. 162:5813–5820. 1999.PubMed/NCBI
|
|
5
|
Wu L, Yun Z, Tagawa T, Rey-McIntyre K and
de Perrot M: CTLA-4 blockade expands infiltrating T cells and
inhibits cancer cell repopulation during the intervals of
chemotherapy in murine mesothelioma. Mol Cancer Ther. 11:1809–1819.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Whiteside TL: Regulatory T cell subsets in
human cancer: Are they regulating for or against tumor progression?
Cancer Immunol Immunother:. 63:67–72. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lindau D, Gielen P, Kroesen M, Wesseling P
and Adema GJ: The immunosuppressive tumour network: Myeloid-derived
suppressor cells, regulatory T cells and natural killer T cells.
Immunology. 138:105–115. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hodi FS, O'Day SJ, McDermott DF, Weber RW,
Sosman JA, Haanen JB, Gonzalez R, Robert C, Schadendorf D, Hassel
JC, et al: Improved survival with ipilimumab in patients with
metastatic melanoma. N Engl J Med. 363:711–723. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Robert C, Thomas L, Bondarenko I, O'Day S,
Weber J, Garbe C, Lebbe C, Baurain JF, Testori A, Grob JJ, et al:
Ipilimumab plus dacarbazine for previously untreated metastatic
melanoma. N Engl J Med. 364:2517–2526. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Suwalska K, Pawlak E, Karabon L,
Tomkiewicz A, Dobosz T, Urbaniak-Kujda D, Kuliczkowski K, Wolowiec
D, Jedynak A and Frydecka I: Association studies of CTLA-4, CD28
and ICOS gene polymorphisms with B-cell chronic lymphocytic
leukemia in the Polish population. Hum Immunol. 69:193–201. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Monne M, Piras G, Palmas A, Arru L,
Murineddu M, Latte G, Noli A and Gabbas A: Cytotoxic T-lymphocyte
antigen-4 (CTLA-4) gene polymorphism and susceptibility to
non-Hodgkin's lymphoma. Am J Hematol. 76:14–18. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu H, Yang J, Jiao S, Li Y, Zhang W and
Wang J: Cytotoxic T lymphocyte antigen 4 expression in human breast
cancer: Implications for prognosis. Cancer Immunol Immunother.
64:853–860. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Khaghanzadeh N, Erfani N, Ghayumi MA and
Ghaderi A: CTLA4 gene variations and haplotypes in patients with
lung cancer. Cancer Genet Cytogenet. 196:171–174. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Laurent S, Queirolo P, Boero S, Salvi S,
Piccioli P, Boccardo S, Minghelli S, Morabito A, Fontana V, Pietra
G, et al: The engagement of CTLA-4 on primary melanoma cell lines
induces antibody-dependent cellular cytotoxicity and TNF-alpha
production. J Transl Med. 11:1082013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hadinia A, Hossieni SV, Erfani N,
Saberi-Firozi M, Fattahi MJ and Ghaderi A: CTLA-4 gene promoter and
exon 1 polymorphisms in Iranian patients with gastric and
colorectal cancers. J Gastroenterol Hepatol. 22:2283–2287. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pawlak E, Karabon L, Wlodarska-Polinska I,
Jedynak A, Jonkisz A, Tomkiewicz A, Kornafel J, Stepien M,
Ignatowicz A, Lebioda A, et al: Influence of CTLA-4/CD28/ICOS gene
polymorphisms on the susceptibility to cervical squamous cell
carcinoma and stage of differentiation in the Polish population.
Hum Immunol. 71:195–200. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang XF, Pan K, Weng DS, Chen CL, Wang
QJ, Zhao JJ, Pan QZ, Liu Q, Jiang SS, Li YQ, et al: Cytotoxic T
lymphocyte antigen-4 expression in esophageal carcinoma:
Implications for prognosis. Oncotarget. 7:26670–26679.
2016.PubMed/NCBI
|
|
18
|
Contardi E, Palmisano GL, Tazzari PL,
Martelli AM, Falà F, Fabbi M, Kato T, Lucarelli E, Donati D, Polito
L, et al: CTLA-4 is constitutively expressed on tumor cells and can
trigger apoptosis upon ligand interaction. Int J Cancer.
117:538–550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dai F, Zhang F, Sun D, Zhang ZH, Dong SW
and Xu JZ: CTLA4 enhances the osteogenic differentiation of
allogeneic human mesenchymal stem cells in a model of immune
activation. Braz J Med Biol Res. 48:629–636. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Duarte S, Momier D, Baqué P, Casanova V,
Loubat A, Samson M, Guigonis JM, Staccini P, Saint-Paul MC, De Lima
MP, et al: Preventive cancer stem cell-based vaccination reduces
liver metastasis development in a rat colon carcinoma syngeneic
model. Stem Cells. 31:423–432. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C (T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ning N, Pan Q, Zheng F, Teitz-Tennenbaum
S, Egenti M, Yet J, Li M, Ginestier C, Wicha MS, Moyer JS, et al:
Cancer stem cell vaccination confers significant antitumor
immunity. Cancer Res. 72:1853–1864. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Choi AR, Park JR, Kim RJ, Kim SR, Cho SD,
Jung JY and Nam JS: Inhibition of Wnt1 expression reduces the
enrichment of cancer stem cells in a mouse model of breast cancer.
Biochem Biophys Res Commun. 425:436–442. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li F, Guo Z, Yu H, Zhang X, Si T, Liu C,
Yang X and Qi L: Anti-tumor immunological response induced by
cryoablation and anti-CTLA-4 antibody in an in vivo RM-1 cell
prostate cancer murine model. Neoplasma. 61:659–671. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shinoyama M, Ideguchi M, Hayashi H, Doi D,
Hashimoto N, Suzuki M and Takahashi J: Cytotoxic T lymphocyte
antigen 4 immunogloblin promotes neuronal differentiation in the
grafts of embryonic stem cell-derived neural precursor cells.
Neuroscience. 202:484–491. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Luo Y, Dallaglio K, Chen Y, Robinson WA,
Robinson SE, McCarter MD, Wang J, Gonzalez R, Thompson DC, Norris
DA, et al: ALDH1A isozymes are markers of human melanoma stem cells
and potential therapeutic targets. Stem Cells. 30:2100–2113. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Topalian SL, Taube JM, Anders RA and
Pardoll DM: Mechanism-driven biomarkers to guide immune checkpoint
blockade in cancer therapy. Nat Rev Cancer. 16:275–287. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Callahan MK, Postow MA and Wolchok JD:
Immunomodulatory therapy for melanoma: Ipilimumab and beyond. Clin
Dermatol. 31:191–199. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ji RR, Chasalow SD, Wang L, Hamid O,
Schmidt H, Cogswell J, Alaparthy S, Berman D, Jure-Kunkel M,
Siemers NO, et al: An immune-active tumor microenvironment favors
clinical response to ipilimumab. Cancer Immunol Immunother.
61:1019–1031. 2012. View Article : Google Scholar : PubMed/NCBI
|